
Abstract
The recent finding that the formation of staphylococcal enterotoxins in food is very different from that in cultures of pure Staphylococcus aureus sheds new light on, and brings into question, traditional microbial risk assessment methods based on planktonic liquid cultures. In fact, most bacteria in food appear to be associated with surfaces or tissues in various ways, and interaction with other bacteria through molecular signaling is prevalent. Nowadays it is well established that there are significant differences in the behavior of bacteria in the planktonic state and immobilized bacteria found in multicellular communities. Thus, in order to improve the production of high-quality, microbiologically safe food for human consumption, in situ data on enterotoxin formation in food environments are required to complement existing knowledge on the growth and survivability of S. aureus. This review focuses on enterotoxigenic S. aureus and describes recent findings related to enterotoxin formation in food environments, and ways in which risk assessment can take into account virulence behavior. An improved understanding of how environmental factors affect the expression of enterotoxins in foods will enable us to formulate new strategies for improved food safety.
Acknowledgments
We would like to acknowledge the Swedish research council for environment, agricultural sciences and spatial planning (FORMAS) for financial support. G.C.B. acknowledges financial support from a competitive strategic grant from the BBSRC, United Kingdom.
Figures and Tables
Figure 1 Regulation of se transcription in S. aureus. Black arrows represent identified direct regulation, while dashed purple arrows indicate potential regulatory pathways. The individual regulators and regulatory pathways are described in the text.Citation66 AIP, autoinducing peptide.
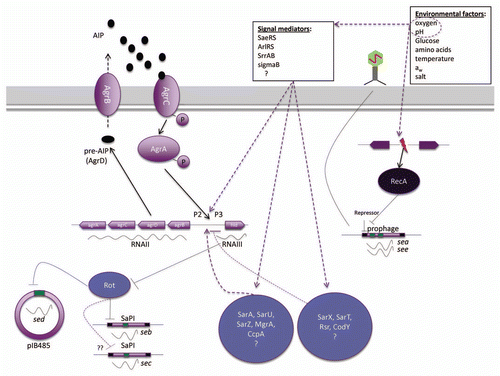
Table 1 Factors affecting Staphylococcus aureus growth and enterotoxin formationCitation4
Table 2 The staphylococcal enterotoxins