
Figures & data
Figure 1. Flavivirus genome organization.
The viral RNA genome (vRNA) of flaviviruses has a single open-reading frame (ORF) flanked by highly structured 5´ and 3´ UTRs that possess cis-acting RNA structural elements required for viral translation, RNA replication, and assembly. The Zika virus (ZIKV) genome is depicted. A) In the linear (translation-competent) form of the vRNA, the capped viral genomic RNA contains stem-loop A (SLA) and SLB in the 5´ untranslated region (UTR), which lie just upstream of the capsid coding-region hairpin (cHP). The 3´ UTR contains a variable region, with one or more stem-loop structures (SL-I and SL-II), and a dumbbell (DB) region, containing one or more DB sequences, all of which can form local pseudoknot (PK) structures. These structures are followed by the short hairpin (sHP) and the 3´ stem loop (3´SL) at the 3´ terminus of the genome. B) In the circular (replication-competent) form of the viral genome, long-range RNA-RNA interactions between elements in the 5´ and 3´ UTRs (5´-3´CS, 5´-3´DAR, 5´-3´UAR) facilitate cyclization of the vRNA. These long-range interactions partially disrupt the 3´SL structure, and fully disrupt the SLB and sHP structures. UAR, upstream of AUG region; DAR, downstream of AUG region; CS, cyclization sequence.
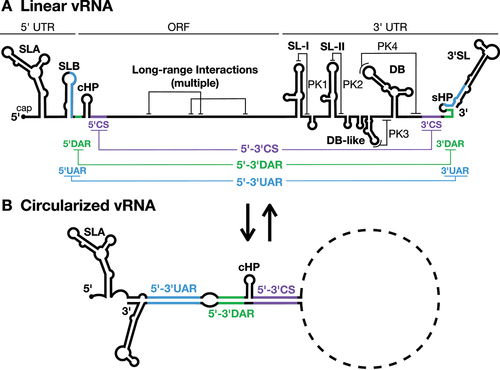
Table 1. RNA structures in the flavivirus genome and their roles in the viral life cycle.
Figure 2. The ‘L/V’ conformations of DENV and ZIKV SLA.
A–C) 2D, 3D and NS5-bound conformations of the DENV ‘L’-shaped SLA structure. The base stem, SSL, and TSL of SLA are coloured in dark green, chartreuse, and pale green, respectively. The AG motif of the TSL is indicated in yellow. The structures of SLA and SLA-NS5 of DENV were generated using PDBs:7LYF and 8GZ9, respectively. In C), the DENV NS5 RdRp and MTase domains are coloured in magenta and pink, respectively; with the Mg2+ ion coordinating addition of the viral cap to the 5´ terminus indicated in red, while the SAM donor is indicated in light purple. The 5´ terminus of the DENV genome (nt 1-3) is depicted in dark blue.D–F) 2D, 3D and modelled NS5-bound conformations of the ZIKV ‘V’-shaped SLA structure. The base stem, SSL, and TL of SLA are coloured in dark blue, teal, and pale blue, respectively. The AG motif of the TSL is indicated in yellow. The structure of ZIKV SLA was generated using PDB:7LYG and the latter SLA-NS5 model was generated by aligning a ZIKV NS5 monomer from PDB:5M2X and the ZIKV SLA structure (PDB:7LYG; nt 4-69) with their counterparts in the DENV SLA-NS5 structure (PDB:8GZP). Notably, as the ZIKV SLA structure was determined via appending the BSL to a tRNA, we have less confidence in the modeled interactions between SLA and the MT domain than those between SLA and the RdRp domain. In F), the ZIKV NS5 RdRp and MTase domains are coloured in magenta and pink, respectively. G) Structural alignment of SLA structures from DENV (PDB:7LYF) and ZIKV (PDB:7LYG).
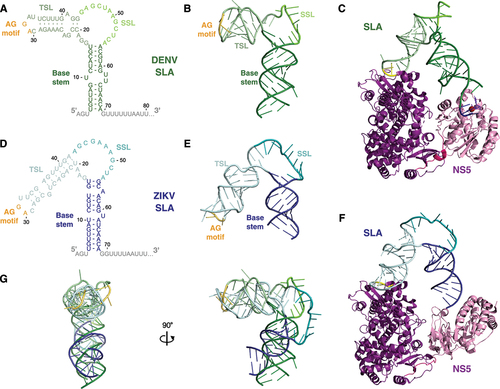
Figure 3. Models for the initiation of negative-strand RNA synthesis.
A) In model 1, NS5 bound to SLA initiates negative-strand RNA synthesis directly on the adjacent 3´ terminus of the positive-strand. SLA is released during the elongation phase of RNA synthesis. B) In model 2, the NS5 bound to SLA initiates negative-strand RNA synthesis directly on the adjacent 3´ terminus, but remains bound to SLA during elongation, only releasing SLA when it is itself unwound for completion of the dsRNA replicative intermediate. C) In model 3, the NS5 bound to SLA is transferred to the 3´SL due to the higher affinity of NS5 for the 3´SL following genome cyclization-induced unwinding of SLB. NS5 then initiates negative-strand RNA synthesis directly on the 3´ terminus of the positive-strand. D) In model 4, the NS5 bound to SLA interacts with a second NS5 bound to the 3´SL, allowing the latter NS5 to initiate negative-strand RNA synthesis off the 3´ terminus of the positive-strand. The 5´ terminus of the genome remains associated with the SLA-bound NS5, only releasing SLA when it is itself unwound for completion of the dsRNA replicative intermediate.
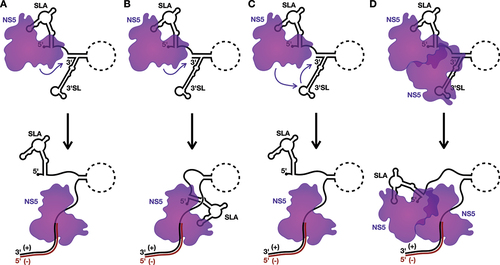
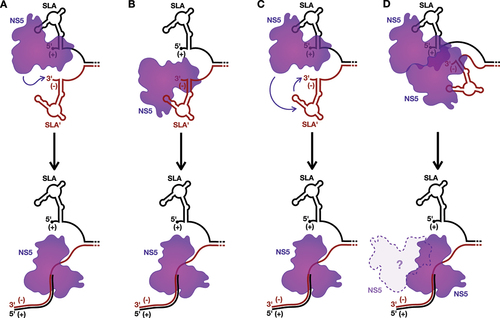
Figure 4. Models for the initiation of positive-strand RNA synthesis.
A) In model 1, NS5 bound to SLA initiates positive-strand RNA synthesis directly on the adjacent 3´ terminus of the negative-strand within the dsRNA replicative intermediate. B) In model 2, NS5 is recruited directly to SLA´ to initiate positive-strand RNA synthesis directly on the 3´ terminus of the negative strand. C) In model 3, the NS5 bound to SLA is transferred to SLA´ and initiates positive-strand RNA synthesis directly on the 3´ terminus of the negative-strand within the dsRNA replicative intermediate. D) In model 4, the NS5 bound to SLA interacts with a second NS5 bound to SLA´, allowing the latter NS5 to initiate positive-strand RNA synthesis off the 3´ terminus of the negative-strand within the dsRNA replicative intermediate. It is unclear whether the NS5 originally bound to SLA would dissociate from or remain bound to the elongating NS5 carrying out positive-strand RNA synthesis. In all cases, the positive-strand is displaced from the dsRNA replicative intermediate during synthesis of the new positive-strand.
Data availability statement
The authors confirm that the data supporting the findings of this study are available within the article.