Figures & data
Figure 1. Targeting of genes encoding key enzymes of fungal aromatic amino acids biosynthesis. (A) Schematic outline of aromatic amino acid biosynthesis in Aspergillus. Starting reagents erythrose-4-phosphate (E4P) and phosphoenolpyruvate (PEP) as well as intermediates chorismic acid (CA), prephenic acid (PA), and anthanilate (AA) are indicated, together with encoding genes of key enzymatic activities, such as aroM (multifunctional enzyme, encoded by A. fumigatus annotated locus AfuA_1G13740), aroB (chorismate synthase, AfuA_1G06940), aroC (chorismate mutase, AfuA_5G13130), and trpA (anthranilate synthase I, AfuA_6G12580). (B) and (C) Schematic outlines of the genomic situation for the targeted gene loci aroC and trpA together with Southern hybridization analyses indicating successful deletion, marker excision, and reconstitution.
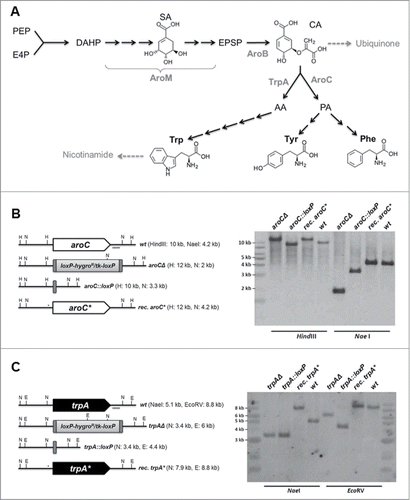
Figure 2. A. fumigatus strains that are auxotrophic for phenylalanine/tyrosine or tryptophan are severely attenuated in virulence. (A) Strains deleted for aroC or trpA are auxotroph for tyrosine and phenylalanine or tryptophan, respectively, as demonstrated by their growth phenotype on supplemented media. 1 × 103 conidia of deletion strains AfS155 (aroCΔ) or AfS157 (trpAΔ) and their reconstituted derivatives AfS156 (aroC*) or AfS158 (trpA*) were spotted on Aspergillus minimal medium (AMM) supplemented with phenylalanine (F), tyrosine (Y), tryptophan (W), or anthranilate (AA) and growth was monitored after 3 days of incubation at 37°C. (B) Germination of an aroCΔ deletion strain is significantly impaired in liquid culture, as assessed by microscopic inspection of inoculated conidia in supplemented culture medium after 18 hours incubation at 37°C: while germ tubes and elongated hyphae became evident for the wild-type progenitor strain or the trpAΔ mutant, could almost none of these structures be seen for the tyrosine/phenylanine auxotroph. (C) Inhibitor studies indicate that deletion of aroC or trpA does not significantly interfere with respiratory capacities of A. fumigatus. Shown are dilution series of indicated strains on culture medium supplemented with the inhibitors CCCP and sodium azide, respectively, after 3 days of growth at 37°C. (D) Virulence of aromatic amino acid biosynthesis deletants in neutropenic mice after pulmonary (left, cohorts of n=15) or intravenous (right, n=5) inoculation with 2 × 105 and 1 × 105 conidia, respectively. Kaplan-Meier plots clearly indicate a significant attenuation of each deletion mutant that is further corroborated by low competitive indices (CI) as determined from mixed pulmonary inoculations applying each deletant together with the progenitor strain AfS35 (middle panel).
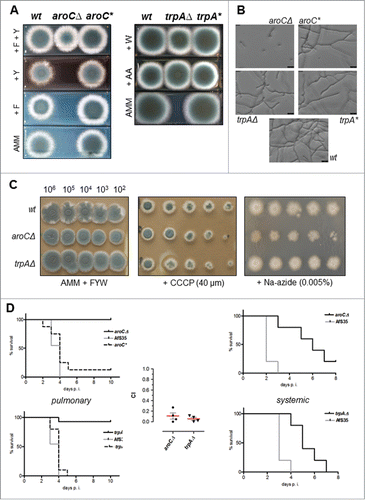
Figure 3. The shikimate pathway is essential for growth of A. fumigatus. (A) Schematic outline of the conditional promoter replacement for aroB by the Tet-ON module to achieve doxycycline-dependent transcription. The recombination event between the gpdA promoter and the identical sequence of the minimal promoter (pmin) resulting in constitutive expression of the gene under Tet-ON control is indicated. (B) Schematic outline together with molecular analyses of the aroB genomic locus after Tet-ON conditional promoter replacement and recombination in the Tet-ON module by Southern analysis with the indicated probe after EcoRV digestion and diagnostic PCR using oligonucleotides that anneal at the specified positions, respectively. (C) Growth of the conditional promoter replacement strain AfS159 [Tet-ON::aroB<ptrA>] depends on the presence of doxycycline, as validated on supplemented medium containing 1 μg·ml-1 of this antibiotic, whereas supplementation of aromatic amino acids does not rescue the growth defect that is caused by silenced aroB transcription in the absence of doxycycline. (D) In vivo repression of aroB gene expression during pulmonary (upper panel, n=11) and systemic (lower panel, n=5) inoculation of susceptible mice is achieved in the absence of doxycycline, demonstrated by significant virulence attenuation of the Tet-ON::aroB strain in the respective cohort. In mice of the -Dox cohort that succumbed to pulmonary aspergillosis, a recombination event in the Tet-ON::aroB cassette had caused constitutive expression of the otherwise silenced gene to reconstitute full virulence. Histopathological inspections revealed hyphal proliferation and invasion in the murine lungs under doxycycline supplementation but no mycelial elements in its absence.
![Figure 3. The shikimate pathway is essential for growth of A. fumigatus. (A) Schematic outline of the conditional promoter replacement for aroB by the Tet-ON module to achieve doxycycline-dependent transcription. The recombination event between the gpdA promoter and the identical sequence of the minimal promoter (pmin) resulting in constitutive expression of the gene under Tet-ON control is indicated. (B) Schematic outline together with molecular analyses of the aroB genomic locus after Tet-ON conditional promoter replacement and recombination in the Tet-ON module by Southern analysis with the indicated probe after EcoRV digestion and diagnostic PCR using oligonucleotides that anneal at the specified positions, respectively. (C) Growth of the conditional promoter replacement strain AfS159 [Tet-ON::aroB<ptrA>] depends on the presence of doxycycline, as validated on supplemented medium containing 1 μg·ml-1 of this antibiotic, whereas supplementation of aromatic amino acids does not rescue the growth defect that is caused by silenced aroB transcription in the absence of doxycycline. (D) In vivo repression of aroB gene expression during pulmonary (upper panel, n=11) and systemic (lower panel, n=5) inoculation of susceptible mice is achieved in the absence of doxycycline, demonstrated by significant virulence attenuation of the Tet-ON::aroB strain in the respective cohort. In mice of the -Dox cohort that succumbed to pulmonary aspergillosis, a recombination event in the Tet-ON::aroB cassette had caused constitutive expression of the otherwise silenced gene to reconstitute full virulence. Histopathological inspections revealed hyphal proliferation and invasion in the murine lungs under doxycycline supplementation but no mycelial elements in its absence.](/cms/asset/480efb73-4406-4744-a7d8-00343e69eefd/kvir_a_1109766_f0003_oc.gif)
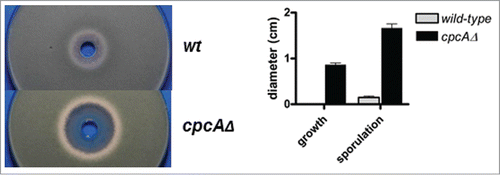
Figure 4. Growth inhibition studies with the shikimate pathway inhibitor glyphosate demonstrate susceptibility of A. fumigatus in a CpcA-dependent manner. Shown are inoculated culture plates (left), to which the herbicide was applied in a centrical reservoir at a concentration of 150 mg/ml together with a quantitative analysis (right) from 3 independent experimental setups presenting the halos' diameters corrected for the punch hole.