Figures & data
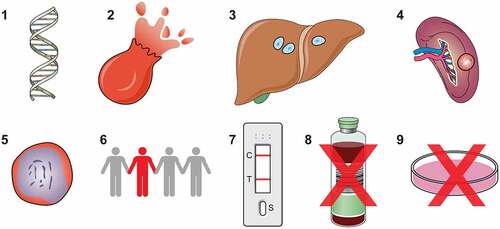
Figure 1. Key pathogenic factors that contribute to severe malaria. High parasite biomass and parasite products released into circulation during cell lysis are stimulants for inflammation and may directly contribute to severe anaemia by reducing erythropoiesis. A strong inflammatory response, including Th1 type cytokine bias and sequestration of innate immune cells, such as monocytes, can activate and damage endothelial cells, enhance sequestration and contribute to severe anaemia by excessive complement mediated lysis of uninfected erythrocytes. A lack of antibodies targeting IE surface antigens may contribute to high parasite biomass and sequestration, although the key targets of antibodies are unclear. Sequestration of IE, mediated by PfEMP1, allows the parasite to evade clearance by the spleen and sequestration in the brain of young children is key to the pathogenesis of cerebral malaria. Sequestration in the placenta is strongly associated with poor pregnancy outcomes. Sequestration in the placenta is mediated by PfEMP1 expressed from VAR2CSA and sequestration in the brain microvasculature is associated with Group A PfEMP1 with domains that bind to ICAM-1 and EPCR. Protective genetic variants can reduce PfEMP1 expression, sequestration and rosetting.
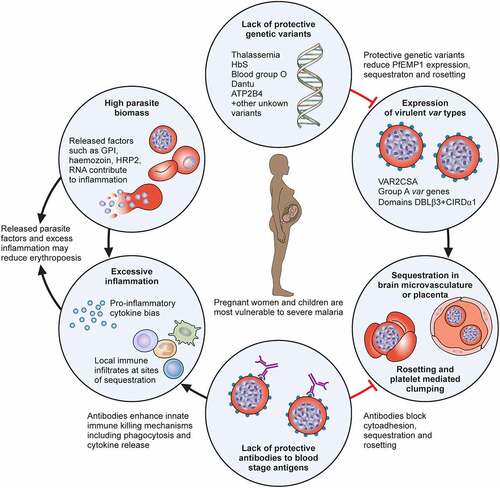
Table 1. Key PfEMP1 types associated with clinical outcomes and evidence of antibody-mediated protection from any clinical outcome. Summarizes studies included in the text only.
Figure 2. Sequestration in Plasmodium falciparum. Reading left to right, the brain colour changes from light tan to a “slate grey” colour in cases of fatal cerebral malaria. Mature infected erythrocytes (IEs) sequester in the deep vasculature, including cerebral vessels. Receptors on endothelial cells include endothelial protein C receptor (EPCR), ICAM-1 and heparan sulphate proteoglycans. Platelets can clump IEs, and bridge between CD36 binding IEs and endothelium. IE can activate monocytes and neutrophils to release cytokines and immune mediators, which can upregulate ICAM-1 expression, and induce damage to the blood–brain barrier. Fluid leakage through and between cerebral endothelial cells can lead to neuronal activation and may result in potentially fatal brain swelling, associated with brain discolouration.
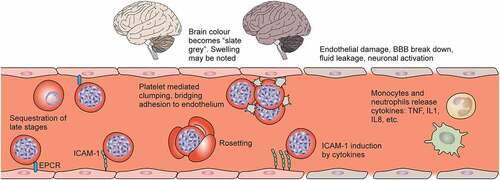
Figure 3. Challenges in Plasmodium vivax. Vivax malaria presents a unique set of challenges for elimination. 1. Genetic mutations: CYP2D6 polymorphisms decrease effectiveness of radical cure, G6PD deficiency predisposes to 2. Drug-induced haemolysis. 3. Hypnozoite reactivation in the liver causes most new episodes of P. vivax malaria. 4. Reservoirs of hidden infection include the spleen (illustrated) and bone marrow. 5. P. vivax forms gametocytes early in infection, increasing transmissibility. 6. Asymptomatic carriers of vivax whose infection may be submicroscopic (figure in red) are common. 7. Low-density infections may not be detected by RDTs. 8. There are no imminent prospects for a P. vivax vaccine. 9. Robust in vitro culture systems are yet to be developed.