Figures & data
Figure 1. Comparison of the current incidence of P. falciparum malaria with that projected in 2030 under the most optimistic scenario considered by the WHO Global Technical Strategy for Malaria 2016–2030, scaling up existing interventions with some near-term innovation in longer lasting nets and better chemoprevention therapies, assuming no loss-of-effect due to drug or insecticide resistance. From Griffin et al. (Citation2016).
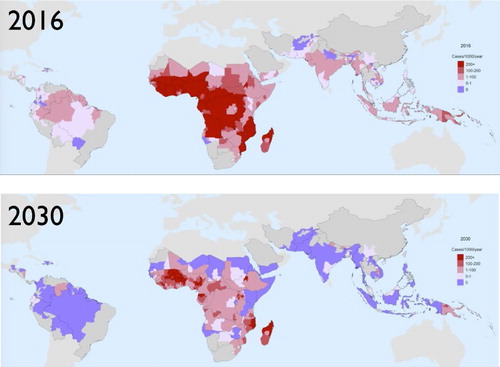
Figure 2. The Ross–McDonald equation for the basic reproductive number for malaria. R0 is the expected number of new infections deriving from a single infection in an otherwise infection-free population; a is the rate at which an adult female bites a human; b is the probability that an infectious mosquito biting a human will transmit the parasite; c is the probability that an infected human will infect a mosquito that bites it; μA is the daily mortality rate for adult female mosquitoes; θE is the probability a female mosquito survives the period between acquiring an infection and becoming infectious; and A is the number of adult female mosquitoes per person. Potential genetic approaches to vector control target one or more of these parameters. r is the rate of recovery of humans, which is unlikely to be affected by vector control. For further details on the equation see Smith and McKenzie (Citation2004).
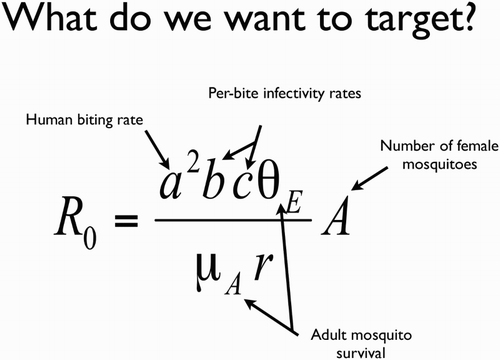
Figure 3. Mendelian transmission of genes does not in itself lead to changes in allele frequency over time, whereas gene drive can. d is the proportion of a heterozygote’s progeny that inherit the driving gene (Mendelian transmission has d = 50%).
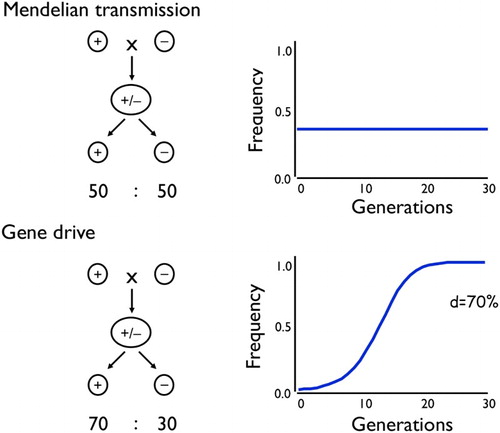
Figure 4. The homing reaction. Diploid cells that are initially heterozygous for the presence of a gene encoding a DNA-cutting enzyme (an ‘endonuclease’) can be converted to homozygotes when the gene is in the middle of its own recognition sequence. The presence of the gene protects the chromosome it is on from being cut, and only chromosomes not containing the gene are cut. The cell’s repair machinery can then use the intact chromosome as a template for repair, resulting in the gene being copied across to the chromosome where previously it was absent. HEG: homing endonuclease gene. After Burt and Trivers (Citation2006).
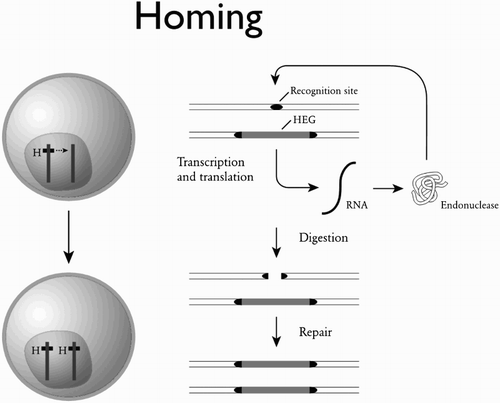
Figure 5. A homing construct for population-wide gene knock-outs. A gene encoding a sequence-specific nuclease that recognises a sequence in a target gene is inserted in the middle of its own recognition sequence, which protects the chromosome it is on from being cut, but also knocks-out the function of the target gene. The nuclease must be expressed in the germline in order to show drive. After Burt (Citation2003).
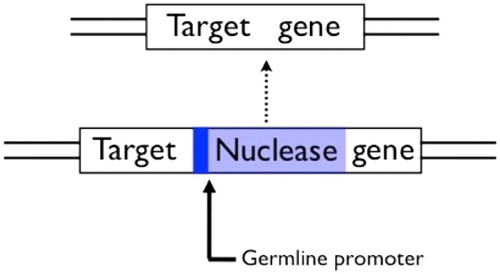
Figure 6. A homing construct for population-wide gene knock-ins. Note the cargo gene will itself need control sequences to ensure expression in the appropriate tissues.
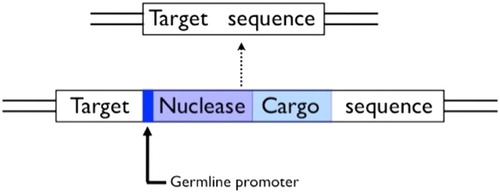
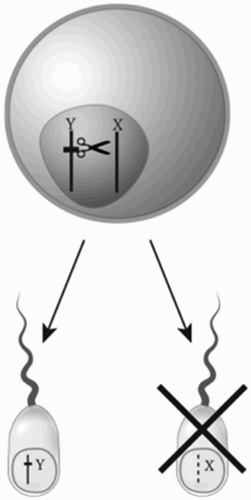
Figure 7. A gene on the Y chromosome that encodes a nuclease that recognises and cleaves a repeated sequence found only on the X chromosome may lead to failure of the X-bearing sperm to develop properly and preferential transmission of the modified Y chromosome – a driving Y.