Figures & data
Figure 1. Thermo-regulation of important virulence factors in pathogenic bacteria species. An increase to 37°C (mammalian body temperature) is a universal invasion signal to the bacteria and enables fine tuning of virulence factor expression, promoting survival and proliferation in its host.
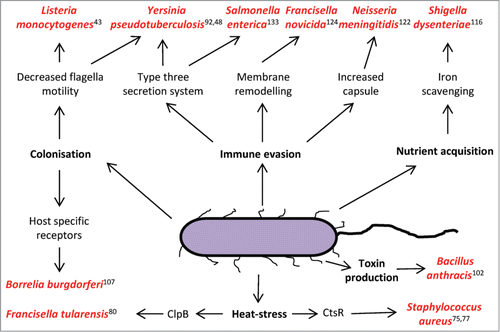
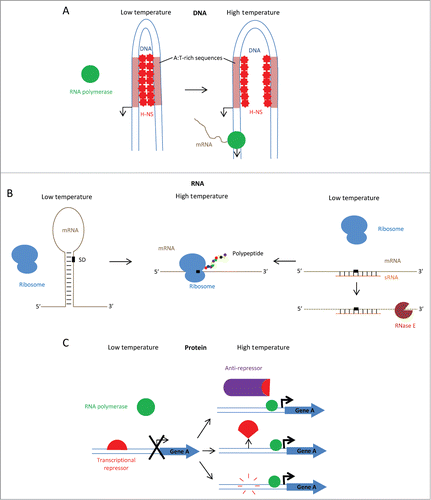
Figure 2. Mechanisms of thermo-regulation (A) At low temperature, the DNA at the promoter region of some virulence genes favors a bent conformation and is stabilised by H-NS, which binds curved DNA at A:T rich sites, blocking access of RNA polymerase to the promoter. At high temperature, homotypic H-NS binding is reduced by relaxation of the DNA bend, enabling access of RNA polymerase to the promoter (e.g. virF in Shigella). (B) At low temperature, mRNA thermometer adopts a stem-loop structure at the 5ʹ position, sequestering the Shine-Dalgarno (SD) sequence from ribosome binding and preventing translation. At high temperature, stem-loop melts to expose the Shine-Dalgarno sequence to ribosome binding and translation occurs. Alternatively, trans-acting small RNAs act by sequestering the SD sequence at low temperature but not at high temperature. Double stranded RNAs are targeted for degradation by RNases. (C) At low temperature, repressor is bound to promoter of Gene A and transcription is prohibited. At high temperature, either: the anti-repressor is stabilised and removes the repressor from the promoter (e.g., GmaR in Listeria monocytogenes), a conformational change causes the repressor to dissociate from the DNA (e.g. RovA in Yersinia spp.) or the repressor is degraded by temperature-dependant proteases which permits the transcription of Gene A (e.g., RovA in Yersinia spp).