Figures & data
Figure 1. Bar diagrams of myosin heavy chains that have roles in cell junctions in vertebrates and/or Drosophila. See the text for descriptions of the different myosins and their key structural features.

Figure 2. The apical junctional complex and calcium-switch model of junction assembly. (A) The tight junction and adherens junction, and their associated actin cytoskeleton, comprise the apical junctional complex. The tight junction strands form a semi-permeable barrier, and ZO-1 functions as a tight junction scaffolding protein. At the adherens junction, the cadherin-catenin complex contributes to cell adhesion. The apical junctional complex separates the apical and basolateral domains. (B) In the calcium-switch model, junctions are disassembled by removing calcium (left, low Ca2+). Upon calcium re-addition (center), the junctions begin to assemble, and radial actin cables are observed at early cell-cell contacts. Cells are fully polarized in a mature monolayer (right).
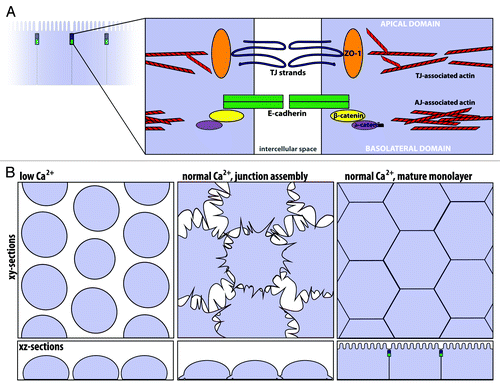
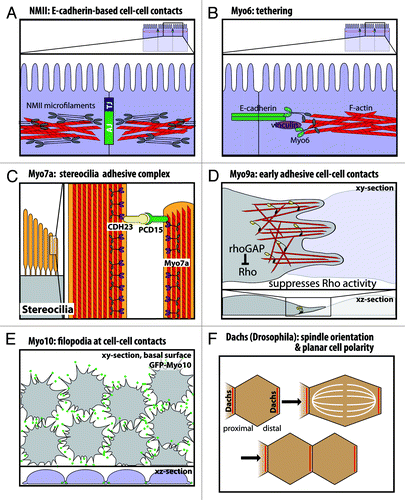
Figure 3. Functional roles of myosins in cell junctions and cell-cell contacts. (A) Nonmuscle myosin-II localizes to the adherens junction-associated circumferential actin belt, and knockdown on nonmuscle myosin-II disrupts E-cadherin-based cell-cell contacts. (B) Myo6 and vinculin have been suggested to tether E-cadherin to the perijunctional actin cytoskeleton. (C) In stereocilia, Myo7a binds cadherin-23 and protocadherin-15 in a stereocilia adhesive tip complex. (D) Myo9a is a RhoGAP that localizes to cell-cell contacts and suppresses Rho activity. Myo9a knockdown shows defects in formation and stabilization of early cell-cell contacts, resulting in a cell scattering phenotype. (E) Myo10 localizes to the tips of filopodia at nascent cell-cell contacts during junction assembly, and Myo10 knockdown delays junction assembly. (F) Dachs has a planar polarized distribution in Drosophila wing disc epithelia. Loss of Dachs disrupts orientation of the mitotic spindle and cell division along the proximal-distal axis.