Abstract
Fossil organic-walled dinoflagellate cysts (dinocysts) have been used to constrain the age of the Maungataniwha Sandstone and associated phosphatic and calcareous concretions from the Mangahouanga Stream, northwest Hawke's Bay, New Zealand. These concretions often contain dinosaur and marine reptile fossils, including the mosasaur Moanasaurus mangahouangae, first discovered within the study area in a calcareous concretion. The Maungataniwha Sandstone contains low to moderately diverse dinocyst assemblages corresponding to the Vozzhennikovia spinulosa – Isabelidinium pellucidum zonal interval (lower to upper Haumurian). Recovered dinocyst assemblages from in situ concretions are similar to those from the surrounding sediments, suggesting that the concretions have grown in place. Calcareous float concretions containing the mosasaur Moanasaurus mangahouangae, as well as the two phosphatic concretions (one of which also contained a mosasaur fossil), are all of a lower Haumurian age. Plesiosaur fossil-bearing calcareous concretions exhibit a range of ages from lower to upper Haumurian (early Campanian to early Maastrichtian), spanning the age range of the Maungataniwha Sandstone section within the study area. The two dinosaur fossil-bearing calcareous concretions can only be dated as no younger than the middle upper Haumurian (early Maastrichtian).
Introduction
The first evidence of Mesozoic reptiles in New Zealand was found in 1974 by WL Moisley, T Crabtree, and J Wiffen. The fossil was recovered from a calcareous concretion in the Mangahouanga Stream, a major tributary of the Te Hoe River, northwest Hawke's Bay (). It was later identified as the caudal vertebrate of a theropod dinosaur (Molnar Citation1981). Since the initial discovery, several terrestrial and marine Mesozoic reptile fossils have been collected from the area, including the new genus of mosasaur Moanasaurus mangahouangae (McKee & Wiffen Citation1998). The vertebrate fossils were largely recovered as fragments within phosphatic and calcareous concretions. These bone-bearing concretions appear to be derived from the Maungataniwha Sandstone, a Piripauan to Haumurian (Santonian to Maastrichtian) shallow marine deposit found along the East Coast of the North Island, New Zealand (Crampton & Moore Citation1990). The Maungataniwha Sandstone has since proved to be New Zealand's most important source for marine and terrestrial vertebrate fossils, and a summary of the fossil findings is presented by McKee & Wiffen (Citation1998).
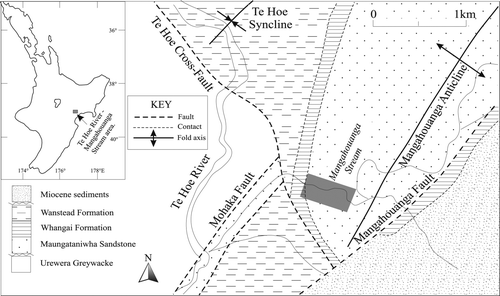
Fig. 1 Location map of the study area (grey box), lower Mangahouanga Stream section, northwest Hawke's Bay.
The Hawke's Bay fossils, along with a theropod bone found in the Huriwai Measures (late Tithonian) south of the Waikato River (Molnar et al. Citation1998), and discoveries from Marlborough, North Canterbury, North Otago, and Chatham Islands (Wilson et al. Citation2005) indicate that terrestrial vertebrates existed in New Zealand both before and subsequent to the separation of New Zealand from Australia and Antarctica at around 81 Ma (Lawver et al. Citation1991).
This paper presents results from a study of dinocysts from samples obtained from loose (float) concretions and in situ sediments and concretions of the Maungataniwha Sandstone. The aim of this work is to improve the age control for the Mesozoic marine reptile and dinosaur fossils found within the concretions, and to determine the potential source of the concretions themselves.
Stratigraphy
A section profile of the study area is shown in . Stratigraphy of the Maungataniwha Sandstone has been described in detailed by Moore (Citation1987) and Crampton & Moore (Citation1990). The Maungataniwha Sandstone is a moderately soft, massive or very poorly bedded, fine to medium grained quartz-rich sandstone. Pyrite is present as small (< 5 cm) irregular nodules in both the concretions and the sandstone. Moore (Citation1987) noted a slight increase in glauconite up through the unit. Although the sandstone is non-calcareous, it does contain both calcareous and phosphatic concretions. The extremely hard calcite-cemented concretions range in size from a few centimetres to c. 2 m in diameter. They are common both in situ and as floats along the lower Mangahouanga Stream. Phosphatic concretions are generally smaller in size, ranging from 5 to 20 cm in diameter, and found as float samples only. The occurrence of reptile fossils within these phosphatic concretions is less common than from calcareous concretions. Diagnostic surface features and delicate structures of the fossilised bones are well preserved within the concretions, indicating limited transportation has occurred prior to burial and incorporation of the bones into the sediment. Early concretionary formation around the bone nuclei has also helped prevent deformation of the fossils.
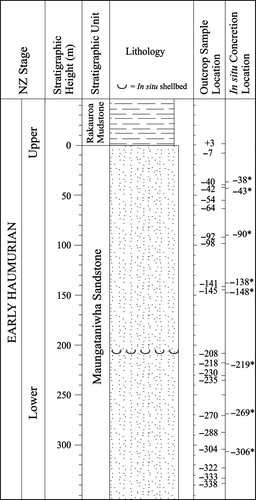
Fig. 2 Measured section profile for the lower Mangahouanga Stream section, showing outcrop (Maungataniwha Sandstone and Rakauroa Mudstone) and in situ calcareous concretion sample locations (the latter are indicated by an asterisk).
The Maungataniwha Sandstone rests unconformably on late Jurassic Urewera Greywacke basement (Moore Citation1987) and the contact is exposed in the upper Te Hoe River and along the lower reaches of the Mangahouanga Stream (see ). The sandstone is in turn unconformably overlain by the Rakauroa Mudstone member of the Whangai Formation. The contact between the two units is gradational, and just below the contact the sandstone becomes softer and more weathered.
Based on calcareous and organic walled microfossils, the Maungataniwha Sandstone has been dated as late Piripauan to early Haumurian (Santonian to Maastrichtian) with the upper 200–250 m of the unit being restricted to Haumurian age (Crampton & Moore Citation1990; Isaac et al. Citation1991; Moore Citation1987). Although the concretions have not been dated directly, Moore (Citation1987) has suggested they may be more common within the upper part of the sandstone.
The abundance of terrestrial (spores, pollen, dinosaur bones, plant remains, and wood fragments) and marine material (turtle bones, marine vertebrates, and shellbeds) recovered from the Maungataniwha Sandstone indicates a shallow marine environment, possibly close to a river mouth or delta (Crampton & Moore Citation1990).
NZ cretaceous dinoflagellate biostratigraphy
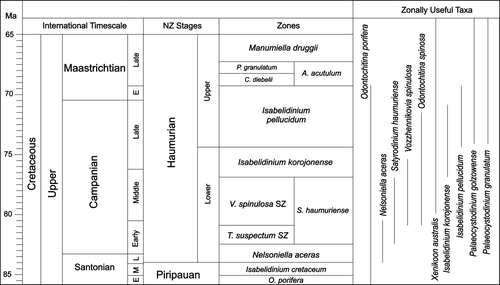
Owing to faunal endemism and the consequent difficulties of correlation with international stages, Mesozoic and Cenozoic strata are subdivided in NZ using a system of local series and stages (Crampton et al. Citation2000). The resolution of the primary dinoflagellate zonation of Wilson (1984) has been enhanced through the ongoing Timescale Programme of the Institute of Geological and Nuclear Sciences (e.g., Schi⊘ler & Wilson Citation1998; Roncaglia et al. Citation1999; Crampton et al. Citation2000). As a result of this work, the late Coniacian to Maastrichtian is currently subdivided into 14 dinoflagellate zones and subzones based on detailed studies in north Canterbury, Marlborough, and Raukumara Penninsula (Schi⊘ler & Wilson Citation1998; Roncaglia et al. Citation1999; Crampton et al. Citation2000, Citation2004). Late Cretaceous New Zealand dinocyst zonation used in this study is based on Roncaglia et al. (Citation1999) and Crampton et al. (Citation2004), as presented in .
Fig. 3 New Zealand dinoflagellate zonations for the Late Cretaceous (Haumurian) and estimated ranges of key taxa (based on Roncaglia et al. Citation1999 and Crampton et al. Citation2004).
Material and methods
In total, 43 samples were collected for palynological analysis. The samples consist of the following:
-
19 Maungataniwha Sandstone outcrop samples covering an interval of 341 m. The limited sampling was due to the extensive vegetation and inaccessibility to certain sections along the Mangahouanga Stream.
-
1 sample from the overlying Rakauroa Mudstone.
-
8 in situ calcareous concretions from the Maungataniwha Sandstone.
-
15 ‘float’ samples from concretions recovered along the stream bed: these included bone-bearing and non-bone-bearing calcareous and phosphatic concretions ().
Table 1 Float concretion types and recovered dinosaur and marine reptile fossils from each sample
In situ calcareous concretions were sampled along with the surrounding sediments to determine if they grew in place and were not present as a result of reworking. The stratigraphic locations for samples from the Maungataniwha Sandstone are shown in , with the stratigraphic height given as metres relative to the contact with the overlying Rakauroa Mudstone.
All samples were processed at the Palynological Laboratory at Victoria University of Wellington following standard laboratory procedures as detailed in Roncaglia et al. (Citation1999). Oxidation via Cold Schulze Treatment was carried out on all samples for 10 minutes, followed by washing in 10% NH4OH. The organic residue was separated using sodium polytungstate, and the resulting residue then mounted in glycerol gelatin for microscopic analysis. The residue was not sieved at any stage of processing due to the low palynomorph content recovered from samples. The slides are lodged with Victoria University of Wellington. Nomenclature follows Fensome et al. Citation2008.
Results
Outcrop samples
Dinocyst counts from the Maungataniwha Sandstone and in situ concretions and from the Rakauroa Mudstone are presented in . Assemblages are generally sparse and poorly preserved, limiting the precision with which ages can be attributed to the samples. Furthermore, because of the strong possibility of intense reworking affecting all species, last appearance datums cannot be used with any degree of confidence in the Mangahouanga Stream section. Determinations of zonal/stage boundaries have been made on first occurrences (FO) only. Three of the 20 samples were barren of palynomorphs. Dark brown wood fragments and amorphous organic matter dominated the majority of residues from both the sandstone unit and the concretions. Although dinocysts are scarce, similar assemblages are found in both the concretion and the surrounding sediment.
Fig. 4 Stratigraphic distribution and counts (number of specimens from two slides) of dinocyst taxa from in situ samples collected along the lower Mangahouanga Stream. Samples marked by an asterisk are from concretions; the remainder are from the surrounding sediment. Suggested zonal assignment is shown on the right-hand side.
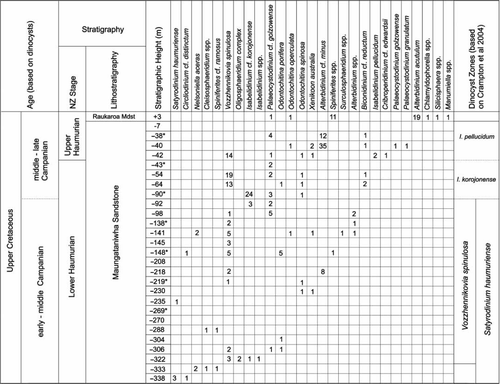
The presence of Satyrodinium haumuriense in the lowest stratigraphic sample (−338 m) and Nelsoniella aceras in the sample immediately above (−333 m) suggests that this interval can be assigned to the Satyrodinium haumuriense zone (lower Haumurian) (Roncaglia et al. Citation1999, Crampton et al. Citation2004). Crampton et al. (Citation2000) uses the first occurrence of N. aceras to define the base of the Haumurian Stage, indicating that the lowest sampled section is no older than basal Haumurian (Late Santonian–Early Campanian) in age.
The FO of Vozzhennikovia spinulosa is used by Roncaglia et al. (Citation1999) to mark the base of the Vozzhennikovia spinulosa Interval Subzone and is considered to be lower Haumurian (Early–Middle Campanian) in age. The first occurrence of this species in the Mangahouanga Stream section is at −332 m, marking the base of the Vozzhennikovia spinulosa sub-zone. Other key first occurrences within this interval include Odontochitina spinosa and Xenikoon australis, both of which occur at −230 m.
Poor preservation prevents confident identification of Isabelidinium korojonense, and therefore determination of the I. korojonense Interval Zone, but there is a marked increase of I. cf korojonense in the concretion sample at −90 m. The first occurrence of Isabelidinium pellucidum is used to mark the lower/upper Haumurian boundary (Crampton et al. Citation2000) and also the base of the I. pellucidum Interval Zone (Roncaglia et al. Citation1999), which is above the I. korojonense zone. In the Mangahouanga Stream section, this event occurs at −42 m. The FO of Palaeocystodinium golzowense and P. granulatum also occur within this zone (Roncaglia et al. Citation1999); they are found in the study area in a concretion from −40 m.
The absence of the FOs of younger zonal indicators (e.g., Cerodinium diebelii and Manumiella druggii) suggests that the topmost section of the Maungataniwha Sandstone (from −42 m upsection) is no older than the Isabelidinium pellucidum zone (upper Haumurian). The single sample from the Rakauroa Member at +3 m contains a different dinocyst assemblage to that recovered from the Maungataniwha Sandstone. However, while the dinocyst assemblage is dominated by the long-ranging Alterbidinium acutulum, the sample age cannot be determined beyond a broad late Cretaceous age.
Concretions in float
Dinocyst assemblages from concretions in float were analysed in order to confirm that these sedimentary structures are of the same age as the concretions found in situ. Also, since the fossil reptiles recovered were from the float material, this more directly dates the vertebrate fossils.
lists the samples analysed and the dinocysts recovered. Dinocyst assemblages from the float material are poorer than those recovered from the outcrop material; however, the assemblages are similar in composition to those recovered from the Maungataniwha Sandstone (see ), suggesting a similar source and age.
Fig. 5 Distribution and counts (number of specimens from two slides) of dinocyst taxa from phosphatic and calcareous concretions collected as ‘float’ from the lower Mangahouanga Stream.
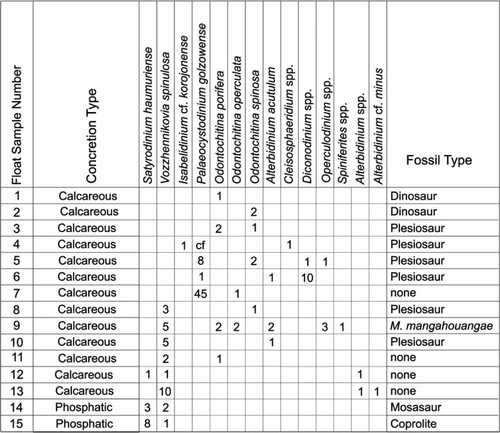
The two phosphatic concretions (Float 14 bearing a mosasaur fossil and Float 15 containing a coprolite) had very low dinocyst counts but can be assigned to the Vozzhennikovia spinulosa subzone (lower Haumurian) due to the presence of the taxa Vozzhennikovia spinulosa and Satyrodinium haumuriense. The non-bone-bearing calcareous concretion Float 11 also contained this species association (along with Odontochitina porifera), indicating a similar age.
Five calcareous concretions (Float samples 8 and 10, both containing Plesiosaur fossils, Float 9 containing the mosasaur M. mangahouangae, and the non-bone-bearing Float samples 12 and 13) contain dinocyst assemblages with V. spinulosa, indicating a V. spinulosa – I. korojonense zonal range (lower Haumurian). Species of Odontochitina present in two of these samples (i.e., O. porifera and O. operculata in Float 9, and O. spinosa in Float 8) further support the Haumurian age for these concretions.
The calcareous Float samples 4, 5, and 6 (all bearing Plesiosaur fossils), and the non-bone-bearing Float 7, all contain Palaeocystodinium golzowense without V. spinulosa, strongly suggesting they are of a younger age (i.e., no younger than upper Haumurian) than the concretions containing V. spinulosa.
Only broad ages can be assigned to Float 1 and Float 2, the two dinosaur fossil-bearing calcareous concretions. The presence of O. porifera in Float 1 and O. spinosa in Float 2 suggests these concretions are no younger than the middle upper Haumurian (early Maastrichtian). Both O. porifera and O. spinosa were also identified in the Plesiosaur fossil-bearing calcareous Float 3, which suggests this concretion is of a broadly similar age to Float 1 and Float 2 (i.e., lower–upper Haumurian).
Conclusions
The dinocyst assemblages recovered from outcrop sediments and concretions of the Maungataniwha Sandstone are of low diversity and poor preservation. The assemblages from both the concretions and the surrounding sediments are similar, suggesting that the concretions have grown in situ. A provisional zonal assignment strongly suggests that the studied section of the Maungataniwha Sandstone can be assigned to the Vozzhennikovia spinulosa – Isabelidinium pellucidum zonal interval (lower to upper Haumurian). Because of the lack of index species such as C. diebelii and M. druggii, it appears unlikely that the section is younger than that represented by the Isabelidinium pellucidum zone.
The majority of marine reptile fossils recovered to date have been from the calcareous and phosphatic concretions in the float of the river bed. Dinocyst assemblages from the phosphatic and calcareous float concretions were generally less diverse that those from outcrop. However, the same basic species associations identified in the concretions and from the outcrop confirm the Maungataniwha Sandstone as a source for these concretions.
The calcareous and phosphatic float concretions exhibited two distinct assemblages: those containing V. spinulosa (and S. haumuriense), and those containing P. golzowense. This suggests that the concretions may derive from at least two broad horizons: a lower Haumurian, and an upper Haumurian and younger age. The phosphatic concretions (one containing a mosasaur fossil) are derived from the older section of the Maungataniwha Sandstone due to the presence of the taxa Vozzhennikovia spinulosa and Satyrodinium haumuriense.
Dinosaur fossil-bearing calcareous concretions can only be broadly dated as no younger than the middle upper Haumurian (early Maastrichtian), based on the presence of O. porifera and O. spinosa. The mosasaur Moanasaurus mangahouangae is dated as lower Haumurian (early-middle to lower middle-late Campanian) based on V. spinulosa, O. porifera and O. operculata, while the plesiosaur fossil-bearing calcareous concretions exhibit a range of ages from lower to upper Haumurian (early Campanian to early Maastrichtian), encompassing the age range of the Maungataniwha Sandstone in the study area.
Acknowledgements
The work in this paper formed the basis of a Batchelor of Science with Honours project conducted at Victoria University of Wellington. The authors thank the late Joan Wiffen and Trevor Crabtree for their encouragement to work in Mangahouanga Stream, and their assistance in the field. Chris Hollis (IGNS), Poul Schi⊘ler (IGNS), Brad Field (IGNS) and Pi Willumsen provided initial ideas, and John Simes (IGNS) additional sample material. The manuscript benefited considerably from a critical review by Erica Crouch.
Related Research Data
References
- Crampton , JS and Moore , PR . 1990 . Environment of deposition of the Maungataniwha Sandstone (Late Cretaceous), Te Hoe River area, western Hawke's Bay, New Zealand . New Zealand Journal of Geology and Geophysics , 33 ( 2 ) : 333 – 348 .
- Crampton , J , Mumme , T , Raine , I , Roncaglia , L , Schi⊘ler , P , Strong , P , Turner , G and Wilson , G . 2000 . Revision of the Piripauan and Haumurian local stages and correlation of the Santonian-Maastrichtian (Late Cretaceous) in New Zealand . New Zealand Journal of Geology and Geophysics , 43 : 309 – 333 .
- Crampton JS Hollis CJ Raine JI Roncaglia L Schioler P Strong CP Wilson GJ 2004 Cretaceous (Taitai, Clarence, Raukumara and Mata Series) 102 122 In: Cooper RA The New Zealand Geological Timescale Lower Hutt Institute of Geological & Nuclear Sciences Limited. Institute of Geological and Nuclear Sciences monograph 22
- Fensome RA MacRae RA Williams GL 2008 DINOFLAJ2, Version 1. American Association of Stratigraphic Palynologists, Data Series no. 1
- Isaac , MJ , Moore , PR and Joass , YJ . 1991 . Tahora Formation: the basal facies of a late Cretaceous transgressive sequence, northeastern New Zealand . New Zealand Journal of Geology and Geophysics , 34 ( 2 ) : 227 – 236 .
- Lawver , LA , Royer , J-Y , Sandwell , DT and Scotese , CR . 1991 . “ Crustal development: Gondwana break-up-evolution of the Antarctic continental margins ” . In Geological Evolution of Antarctica , Edited by: Thompson , MRA , Crame , JA and Thompson , J . 533 – 540 . Cambridge University Press .
- McKee JWA Wiffen J 1998 Mangahouanga Stream-New Zealand's Cretaceous dinosaur and marine reptile site Miscellaneous Publication, 96. Geological Society of New Zealand
- Molnar RE 1981 A dinosaur from New Zealand In: Cresswell MM Vella P Gondwana five Fifth International Gondwana Symposium, Wellington, New Zealand. Rotterdam, A.A. Balkema 91 96
- Molnar , RE , Wiffen , J and Hayes , B . 1998 . A probable theropod bone from the latest Jurassic of New Zealand . New Zealand Journal of Geology and Geophysics , 41 ( 2 ) : 145 – 148 .
- Moore , PR . 1987 . Stratigraphy and Structure of the Te Hoe -Waiau River area, western Hawkes Bay . New Zealand Geological Survey Record , 18 : 4 – 12 .
- Roncaglia , L , Wilson , GJ , Field , BD , Raine , JI and Schi⊘ler , P . 1999 . Dinoflagellate biostratigraphy of Piripauan-Haumurian (Upper Cretaceous) sections from northeast South Island, New Zealand . Cretaceous Research , 20 ( 3 ) : 271 – 314 .
- Schi⊘ler , P and Wilson , GJ . 1998 . Dinoflagellate biostratigraphy of the middle Coniacian-lower Campanian (Upper Cretaceous) in south Marlborough, New Zealand . Micropaleontology , 44 ( 4 ) : 313 – 349 .
- Wilson , GJ . 1984 . New Zealand Late Jurassic to Eocene dinoflagellate biostratigraphy-a summary . Newsletters on Stratigraphy , 13 : 104 – 117 .
- Wilson , GJ . 1987 . Dinoflagellate biostratigraphy of the Cretaceous-Tertiary boundary, Mid-Waipara section, north Canterbury, New Zealand . New Zealand Geological Survey Record , : 8 – 15 .
- Wilson , GJ , Schi⊘ler , P , Hiller , N and Jones , CM . 2005 . Age and provenance of Cretaceous marine reptiles from the South Island and Chatham Islands, New Zealand . New Zealand Journal of Geology and Geophysics , 48 : 377 – 387 .