Abstract
The New Zealand continental shelf has a rich bryozoan fauna, and Bryozoa are relatively common as fossils in local marine-deposited strata. We record 103 species of Bryozoa occurring in the Wanganui Basin sequence spanning the Plio-Pleistocene (3.0–0.3 Ma), 77 of which belong to the order Cheilostomata and the remainder to the order Cyclostomata. The majority are conspecifics of the modern New Zealand shelf fauna. Palaeoecological characteristics of Wanganui Basin bryozoan faunas including diversity, taxonomic and colony growth-form combinations are identified, defining three biofacies groupings corresponding to depositional sequences (systems tracts). Water depth, sedimentation, substratum characters and taphonomic processes have all influenced the distribution and preservation of bryozoans in the local fossil record. Broader stratigraphic variations in Wanganui Basin faunas are explained by ‘habitat tracking’: a lateral shifting of bryozoan palaeocommunity zones in response to changing environments related to Plio-Pleistocene glacio-eustatic sea-level fluctuations.
Introduction
Bryozoans are colonial, suspension-feeding, mostly marine organisms that usually attach to a variety of hard or soft substrata including rock, shell, hard anthropogenic surfaces, seaweeds and other organisms from the intertidal to the deep sea. Bryozoan colonies range from simple weakly integrated uniserial encrusting forms to large multiserial structures, composed of hundreds of thousands of individuals that are highly integrated (McKinney & Jackson Citation1989). Erect colonies can be large enough to create seafloor habitat for a variety of other organisms (Bradstock & Gordon Citation1983; Batson & Probert Citation2000; Gordon et al. Citation2009). Bryozoans contribute to hardgrounds and reefs, as summarised by Taylor & Wilson (Citation2003). The carbonate skeletons of bryozoan colonies are precipitated from seawater and may be calcite (mostly intermediate-Mg content, but can vary) or aragonite or a mixture (Smith et al. Citation1998).
Bryozoans invade available substratum space by means of sexually produced larvae that respond to a variety of environmental cues, hence surface nature (topography, orientation, hydrophilicity, etc.) rather than the type of substratum per se can be important for determining diversity and distribution (Ward & Thorpe Citation1989). Once a colony is established, the driving non-genetic factors involved in growth and form seem to be spatial limitation and competitors and/or water flow and feeding (see Jackson et al. Citation2001). In terms of surface area, relative diversity increases on larger substrata and the variety of interactions, including overgrowth, potentially increase as colonisation by different epibionts takes place (Kay & Keough Citation1981).
At least 953 species of Recent marine bryozoans have so far been discovered in the New Zealand Exclusive Economic Zone (EEZ) (Gordon et al. Citation2009), with the greatest diversity occurring in shallow-coastal and shelf waters (Gordon Citation1987). A high proportion (probably greater than 61%) of the bryozoans found in the New Zealand region are endemic (Gordon et al. Citation2009), with the majority of living marine species belonging to the order Cheilostomata (class Gymnolaemata) plus representatives of the order Cyclostomata (class Stenolaemata).
Bryozoans can be the single-most abundant skeletal component in New Zealand shelf carbonate sediments and dominate Cenozoic limestones, forming 30–70% of the carbonate fraction (Nelson et al. Citation1988). Recent taxonomic study of modern bryozoan species (e.g. Gordon Citation1984, Citation1986, Citation1989) aids reliable identification of Cenozoic taxa. In a recent overview of New Zealand bryozoan diversity, Gordon et al. (Citation2009) noted there are >368 recorded bryozoan species found in New Zealand Cenozoic rocks, with many fossil taxa currently undescribed. The fauna contains many component species that appear to have relatively long stratigraphic ranges, extending over at least ten million years (Gordon Citation1994; Gordon et al. Citation1994, Citation2009). Current knowledge of the New Zealand fossil record suggests that c. 1% of the Eocene, c. 25% of the Oligocene, c. 45% of the Miocene and some 65% of the Pliocene bryozoan species have survived to the present (Gordon et al. Citation2009), but taxonomic revision is likely to reduce these percentages.
The fossil record of bryozoans has the potential to add much to the understanding of New Zealand marine-deposited Cenozoic strata, especially in conjunction with results from other studies. In this article, we record the first detailed analysis of bryozoans occurring through the important, well-studied Wanganui Basin Plio-Pleistocene sequence, highlighting their usefulness in sequence stratigraphy and palaeoenvironmental interpretations. Our results are compared with known features of Wanganui Basin sedimentological and molluscan data to assess how bryozoan distribution and preservation relates to local variations in habitat, or basin-wide processes. This study identifies, for the first time, changes in New Zealand bryozoan faunas through this significant period known to be affected by cyclic sea-level fluctuations.
Geological setting
The Wanganui region is situated in the southwest of the North Island, New Zealand, at approximately 40°S latitude. Wanganui Basin itself is a 200×150 km ovoid sedimentary basin situated in a ‘behind-arc’ or ‘proto back-arc’ position with respect to the active Hikurangi subduction margin, and originated as an extensional regime related to the wider evolution of the Australia–Pacific plate-boundary zone (Kamp et al. Citation2004; Proust et al. Citation2005). The basin is bounded to the west by the Taranaki fault and associated Patea–Tongaporutu High and the Kapiti–Manawatu Fault systems to the east (see Proust et al. Citation2005). Sediments have been uplifted and exposed on land due to gentle upwarping of the eastern margin of the basin nearest the plate boundary, tilting strata at a low angle towards the south. Plio-Pleistocene strata are well exposed today in the river valleys and as coastal cliffs northwest of Wanganui city, from Castlecliff and Kai Iwi Beach to Nukumaru, which form the stratotype sections for the Castlecliffian and Nukumaruan Stages of the New Zealand Plio-Pleistocene (Thompson Citation1916; Cooper Citation2004).
Wanganui Basin contains a 4–5 km thick record of predominantly shelf and shallow-water siliciclastic sediment, the terrigenous input supplied from erosion of the Southern Alps and North Island ranges. There are localised areas with a notable carbonate component (largely bryozoan remains) as on the modern Wanganui Shelf (Gillespie et al. Citation1998). Occasional influxes of volcanic material from the central North Island provide tephrachronology and palaeomagnetic analyses for correlation (e.g. Pillans et al. Citation2005; Turner et al. Citation2005). Recognised sedimentary facies include coastal plain, shore-face and shallow shelf environments. Major contributions to the description and understanding of Wanganui geology were made by Fleming (Citation1947, Citation1953), who emphasised the repetitive nature of depositional sequences at Wanganui. Following workers recognised the implied sea-level changes as the result of global sea-level cycles (Beu & Edwards Citation1984; Abbott & Carter Citation1994). Intensive mapping and logging of lithologic facies and sampling of units for macro- and microfossils (e.g. Abbott Citation1997; Abbott & Carter Citation1997) revealed repeating cyclothem systems tracts and localised motifs (e.g. Carter & Naish Citation1998). Multidisciplinary geologic, sequence-stratigraphic and palaeontological studies now provide a high-resolution chronostatigraphic framework for the Plio-Pleistocene sequence (Carter & Naish Citation1998; Naish et al. Citation1998).
Wanganui Basin Plio-Pleistocene stratigraphy
Wanganui Basin strata spanning the Middle Pliocene to Pleistocene (3.0–0.3 Ma; OIS G–OIS 9) consists of some 48 lithostratigraphic units comprising a 2.5 km thick vertical stack of some 50 cyclic unconformity-bounded depositional sequences (Abbott et al. Citation2005; Naish et al. Citation2005). These are correlated with sea-level cycles on the deep marine oxygen isotope curve, revealing that basin architecture has been controlled by 41 and 100 ka duration (5th and 6th order) glacio-eustatic sea-level fluctuations. Sequence stratigraphic facies successions represented include the transgressive systems tract (TST), highstand systems tract (HST) and regressive systems tracts (RST) of each rise, highstand and early fall of sea level ().
Figure 1 An idealised Pleistocene Wanganui Basin shallow-marine depositional sequence, showing some sequence stratigraphic features (modified from Abbott and Carter Citation1994; Abbott et al. Citation2005), inferred relative changes in sea level and associated bryozoan-based biofacies with some commonly occurring taxa. Insert: map of North Island, New Zealand showing location of Wanganui.
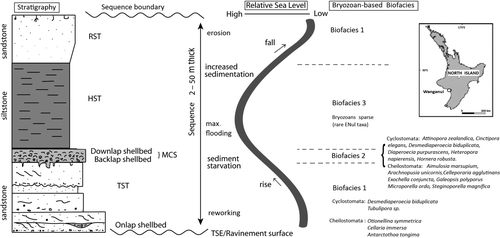
Four key Mangapanian Stage (3.0–2.4 Ma) facies associations are identified by Kamp & McIntyre (Citation1998): siltstone, siliciclastic sandstone, carbonate shellbeds and conglomerate. The shellbeds often occur at the base of sequences (above transgressive surfaces of erosion) and are mostly compound in nature. They may contain numerous Phialopecten and the last of some warmer-water mollusc genera: Maoricardium, Polinices and Zeatoma (Beu Citation2004). Mangapanian highstand and regressive systems tracts are typically represented by thick sandstones.
The local Nukumaruan Stage (2.4–1.63 Ma) encompasses the Pliocene–Pleistocene boundary (1.81 Ma; in OIS 65). Nukumaruan transgressive, regressive and highstand systems tracts are dominated by extensive shallow-marine, siliciclastic sand-rich facies. Shellbeds are present in the sequences but vary in their origin, thickness and composition. The onlap shellbeds are often cross-bedded and often contain the molluscs Paphies, Austrovenus and Zethalia, while backlap shellbeds contain significant Tiostrea and Tawera in situ (Naish et al. Citation2005). During Nukumaruan lowstands, the shoreline was situated perhaps many tens of kilometres to the west of the present Wanganui coast. Substantial non-marine systems tracts composed of lignites and palaeosols were deposited (see Fleming Citation1953; Naish et al. Citation2005).
A comprehensive summary of Castlecliffian stage (1.63–0.34 Ma) strata identified some 27 lithofacies grouped into three associations: mudstones, sandstones and shellbeds (Naish et al. Citation2005, p. 104, ). Deposited in a variety of environments from coastal swamps and sand dunes, shoreface and beach settings to inner- and possibly mid-shelf depths; sea-level (water-depth) changes of up to 50 m are suggested through the sequence (Abbott Citation1997; Naish et al. Citation2005).
Table 1 Bryozoan fauna beta-diversity (number of species recorded per unit) and biofacies represented in the Wanganui Basin Plio-Pleistocene sequence sampled by Rust (Citation2009) (for key features of bryozoan biofacies, see ).
Analysis of bryozoans from Wanganui Basin strata
Brown (Citation1952) described a significant number of New Zealand Cenozoic cheilostome bryozoans including specimens from Wanganui, mostly collected by Fleming during the 1940s. Brown (Citation1952) examined six samples from the Castlecliff section, including the Tainui Shellbed which contains bryozoans ‘… concentrated in lens-like colonies or bioherms’. He recorded 18 species from the unit, 7 of which exhibited an encrusting habitat (Brown Citation1952). Brown described species taxonomically and noted the variation in forms collected from different beds at Castlecliff, but provided little detail on the nature of the specimens, substratum or geological field information and did not include the cyclostome fauna (cf. Waters 1887).
This study utilises a comprehensive record of bryozoan abundance and diversity through a set of systematic samples spanning the Wanganui Basin Plio-Pleistocene sequence. Each unit was carefully studied in outcrop and bulk-sampled from at least one field site; some horizons were supplemented by the inclusion of associated bryozoan material in previous collections. Bryozoans were identified using Scanning Electron Microscope (e.g. ) and all taxa and colony forms were counted on tally sheets with details on the nature of specimens (e.g. degree of preservation, substratum encrusted) also recorded. Species counts were made for each sample and unit to give a measure of alpha- and beta-diversity, respectively. Palaeoecological interpretations were made using ecological data from extant representatives of bryozoan taxa and analogous modern habitats on the New Zealand shelf (e.g. Gordon Citation1984, Citation1986, Citation1989; Gillespie et al. Citation1998). Further statistical analysis of the fossil assemblages from Wanganui provided definitive, visual and comparable measures of the observed stratigraphic patterns and faunal differences between samples. Features of the dataset such as bryozoan diversity, presence/absence of taxa, relative abundance of taxa and percentages of morphological characters, including growth form, were represented numerically in a spreadsheet database. They were organised into matrices which were constructed and analysed by Rust (Citation2009) using the software PAST version 1.38 (Hammer et al. Citation2006). Due to the large dataset involved, multivariate techniques were used including R-mode (taxon-centred) cluster analysis to find commonly co-occuring taxa and ordination and Q-mode (sample-centred) cluster analysis to classify samples on the basis of their faunal composition and subdivide the data into bryozoan palaeocommunities/biofacies. This paper provides an overview of results; dendograms and further details on analyses are provided in Rust (Citation2009). Locations, grid references and New Zealand fossil record (FR) numbers for sampled sites can be accessed via the Fossil Record Electronic Database (www.fred.org.nz). Auckland University (AU) numbers have been assigned to assemblages collected, and specimens are housed in the collections of the Geology Department, University of Auckland, New Zealand.
Figure 2 SEM images of fossil bryozoans from the Wanganui Basin. (A) Crepidacantha parvipora, Lower Kai Iwi Shellbed, Kai Iwi Beach, Wanganui (Castlecliffian Stage, Pleistocene), scale bar: 0.2 mm. (B) Valdemunitella cf. fraudatrix, Lower Castlecliff Shellbed, Castlecliff, Wanganui (Castlecliffian Stage, Pleistocene), scale bar: 0.2 mm. (C) Desmediaperoecia biduplicata, Lower Kai Iwi Shellbed, Kai Iwi Beach, Wanganui (Castlecliffian Stage, Pleistocene), scale bar: 0.5 mm. (D) Mobunula bicuspis growing on the exterior of Chlamys sp. from the Tainui Shellbed, Castlecliff, Wanganui (Castlecliffian Stage, Pleistocene), scale bar: 1.0 mm. (E) Aimulosia marsupium, Lower Castlecliff Shellbed, Castlecliff, Wanganui (Castlecliffian Stage, Pleistocene), scale bar: 0.2 mm. (F) Rogicka biserialis from the Tainui Shellbed, Castlecliff, Wanganui (Castlecliffian Stage, Pleistocene), scale bar: 1 mm. (G) Otionellina squamosa from the Landguard Formation, Landguard Bluff, Wanganui (Haweran Stage, Pleistocene), scale bar: 1.0 mm.

Results
Bryozoans occurred in samples from 29 of the 48 Wanganui Basin units studied (). Lists of bryozoans identified in each unit, a total faunal list and biostratigraphic range chart were compiled (see Rust Citation2009). The gamma diversity of the entire collection, 103 bryozoan taxa (), was determined from an estimated 1200 colonies. About 75% (77 species) of these belong to the order Cheilostomata. The remainder (26 species) are members of the order Cyclostomata. This result is consistent with the reported global trend, where cheilostomes are dominant at an average level of 80% for the Pliocene and Pleistocene (Lidgard et al. Citation1993).
Table 2 Taxonomic list of cheilostome and cyclostome bryozoans recorded from the Plio-Pleistocene sequence of the Wanganui Basin
Most fossil bryozoan taxa at Wanganui were represented by the encrusting colony form. Some 692 bryozoan colonies were recorded encrusting fossil oyster and pectinid valves alone. Other notable occurrences include fragments of the erect branching cyclostomes Cinctipora elegans, Attinopora zealandica and Cylindropora areolata, all members of the family Cinctiporidae (Boardman et al. Citation1992) which is endemic at the present day. Further known New Zealand endemics (see Gordon et al. Citation2009) include Tretosina flemingi, Eurystomella biperforata, Mobunula bicuspis, Desmediaperoecia biduplicata and Odontoporella bishopi.In addition, three bryozoan species (Chiastosella sp., Microporella sp. and Smittina sp.) collected from Wanganui lack formally validated species names although they may have been recognised previously in New Zealand material, namely in the study by Brown (Citation1952). No true origination events were recognised for bryozoans in the Wanganui sequence. The majority of taxa are relatives of the modern New Zealand shelf fauna (Gordon et al. Citation2009) with stratigraphic ranges that extend right through the Pliocene to the Recent. However, closer analysis of time ranges reveal that most taxa have a locally interrupted (episodic) fossil record and were perhaps at times absent from the Wanganui region.
Bryozoan-encrusted substrata in the Wanganui Basin sequence include lithic clasts and pebbles (of varying lithology), bivalve and gastropod mollusc shells (e.g. A, B, D–F), brachiopod shells, barnacles, rare echinoid fragments, rare corals and also the colonies of other bryozoans. The brachiopod Calloria inconspicua is found in several Pleistocene units at Castlecliff; interestingly, exterior surfaces of valves are often encrusted by the cheilostome Crepidacantha crinispina. Other bryozoans occur on Calloria, but infrequently.
Figure 3 Fossil specimens from the Wanganui Basin. (A) An example of the diverse bryozoan fauna seen in the Nukumaru Limestone: here numerous bryozoan colonies encrust the exterior of a large Patro undatus valve from Waiinu Beach (Nukumaruan Stage, Pliocene). (B) Desmediaperoecia biduplicata encrusting the interior of a Patro undatus valve, Nukumaru Limestone, Waiinu Beach (Nukumaruan Stage, Pliocene). (C) Hornera robusta branches from the Tainui Shellbed, Castlecliff, Wanganui (Castlecliffian Stage, Pleistocene). (D) Bryozoan colonies encrusting a fragment of a Patro undatus valve from the Tewkesbury Formation, Nukumaru Beach (Nukumaruan Stage, Pliocene), bryozoans include Chaperia sp., Platonea sp. (inside oyster spat), Exochella conjucta (several colonies) and Fenestrulina reticulata. (E) Cosciniopsis vallata, growing on a bioeroded specimen of the turritellid gastropod Maoricolpus sp., Lower Castlecliff Shellbed, Castlecliff, Wanganui (Castlecliffian Stage, Pleistocene). (F) Steginoporella magnifica, growing on a bioeroded Ostrea valve from the Lower Castlecliff Shellbed, Castlecliff, Wanganui (Castlcliffian Stage, Pleistocene). (G) Bryozoan-encusted interior surface of a Pecten valve from the Landguard Formation, Landguard Bluff, Wanganui (Haweran Stage, Pleistocene). All scale bars: 10 mm.

Overall, the valves of bivalve molluscs, especially ‘oysters’ (e.g. Ostrea sp., Patro undatus, Anomia trignopsis), pectininds (Pecten sp., Phialopecten sp.), mesodesmatids (Paphies sp.), carditids (Purpurocardia sp.), glycymerids (Tucetona sp.) and mactrids (Mactra sp.) supported the greatest abundance and diversity of encrusting bryozoans. Abundances of mollusc valves varied through sequences; the richest units sampled included onlap, backlap or compound shellbeds; the latter typically formed mid-cyclothem during somewhat sediment-starved deposition, for example the Tainui Shellbed (e.g. Kondo et al. Citation1998). Most of the well-preserved Pecten specimens are found in the mid-cycle shellbeds (Beu Citation2006) and hence usually form part of the ‘Ostrea Association’ of Abbott & Carter (Citation1997), along with a rich oyster and bryozoan fauna. Of bryozoans recorded encrusting oysters and pectinid shells by Rust (Citation2009), 86% (579 colonies) are cheilostomes belonging to 55 taxa (85% of diversity) and 14% (97 colonies) are cyclostomes comprising 10 taxa (15% of diversity).
Variation in spatial occurrences of different bryozoan genera present on mollusc valve surfaces was noted in the Wanganui samples following studies by Ward & Thorpe (Citation1989, Citation1991). Some taxa occurred as widely dispersed generalists while others appeared to have preferred distributions. Occurrence of bryozoans (and other epibionts) on interior surfaces of shells from many horizons (especially TST deposits) indicate molluscs were deceased before encrustation (cf. McKinney Citation1996). Overall, orientation, topography and taphonomic features of mollusc valves appear to influence suitability for bryozoan establishment and growth. Ultimately, the distribution of bryozoan colonies is influenced by local interrelated ‘ecological controlling parameters’ (see Hageman et al. Citation1997) such as nutrient availability, light intensity, local water energy, sedimentation rate and substratum type, as well as broader environmental factors.
Colony growth-form analysis
Bryozoan colony morphology (growth form) has been used as a tool in environmental interpretation of both modern and fossil faunas. For sessile bryozoan colonies, the diversity of forms represents an adaptive strategy with different costs and benefits to the organism in/on a particular environment and substratum (see McKinney & Jackson Citation1989). Researchers categorised a suite of recurring colony forms (see Hageman et al. Citation1997 for a review), although the categories and relative abundance of these growth form variations does not always correspond to taxonomic groupings. Most studies show that certain bryozoan growth forms characterise, but are not exclusive to, certain environments (Hageman et al. Citation1997).
Analysis of growth forms present at Wanganui reveal that the unilaminar encrusting (ENul) form (sometimes termed ‘membraniporiform’ for cheilostomes and ‘stomatiporiform’ for cyclostomes) is the most taxonomically diverse colony morphology in the sequence, represented by members of 82 taxa. The encrusting growth form (EN) component often constitutes >60% of the colonies counted in samples containing bryozoans.
Many units (such as the Tainui Shellbed, Lower Castlecliff Shellbed) contain a variety of other growth forms, including abundant erect (ER) types. These tree-like branching forms are represented mainly by cyclostomes, especially members of the Cinctiporidae and Horneridae (e.g. C). Delicate-branching erect growth forms (ER) and erect flexible (EF) forms break down and disarticulate more readily during taphonomic processes; hence, they are often over-represented in abundances based on fragment counts. However, erect delicate colonies appear to dissolve readily and may be under-represented (as biomass) in certain chemical conditions (Smith et al. Citation1998).
Small, discoidal, free-living (FL) forms (e.g. G) belong to the genus Otionellina (Otionellidae) (Cook & Chimonides Citation1984), while local erect flexible forms are represented by fragments of Cellaria (Cellariidae). The radiate (ERra) form is moderately abundant, usually represented by the small cyclostomes Disporella novaehollandiae and D. wanganuiensis. Interestingly, erect fenestrate (ERfe), and foliose (ERfo) bryozoan colonies are rare in the Wanganui Basin; however, one would expect them to occur (if only as fragments) in the compond shellbeds. Perhaps the taxa known to produce these growth forms were rare or locally absent in the Wanganui Basin at this time. Erect colonies of Adeonellopsis species have not been found, even although the genus is abundant on the modern Wanganui Shelf (see Gillespie et al. Citation1998). This may partly be a consequence of chemical dissolution of the largely aragonitic skeleton (Smith et al. Citation1998) in some deposits; this likelihood notwithstanding, aragonitic Otionellina squamosa is frequently preserved at Wanganui (Brown Citation1952).
Hermit-crab symbiont tubular bryozoan colonies were collected from the Landguard Formation (Odontoporella bishopi), Lower and Upper Castlecliff Shellbeds and Tainui Shellbed (Akatopora circumsaepta) and Nukumaru Limestone (Disporella pristis, Tetrocycloecia parapelliculata). Although both paguran hermit crabs and their symbiont bryozoans are most commonly encountered in shallow shelf depths today (Taylor et al. Citation1989), they can vary widely in habitat and do not seem to be heavily influenced by temperature. They therefore add little, unfortunately, to palaeoenvironmental interpretations (Taylor et al. Citation2004).
Wanganui faunal associations and palaeocommunities
Particular sedimentary facies and systems tracts generally exhibit a distinctive suite of fossils defined as a biofacies, reflecting an assemblage (palaeocommunity) which lived in a certain habitat and subject to taphonomic processes (e.g. Abbott & Carter Citation1997; Hendy & Kamp Citation2004, Citation2007). In the Wanganui Basin the fossil record of molluscs is well studied, comprehensive, rich and diverse (Beu & Maxwell Citation1990; Johnson & Curry Citation2001). Typically, units deposited during the transgressive parts of each sea-level cycle (TSTs) have a high diversity of molluscs, while the highstand (HST) deposits contain only sparse macrofossils (Abbott & Carter Citation1997; Hendy & Kamp Citation2004, Citation2007).
Field observations at Wanganui suggest that characteristics of bryozoan faunas also vary through each depositional sequence and across the basin. Vertical changes in benthic assemblages through a sequence section, or across a gradient perpendicular to the shoreline, typically reflect bathymetric changes. Lateral faunal changes across a horizon or area parallel to the shoreline may indicate microhabitat variations, differences in grain size, substratum or disturbance. In the Wanganui Basin as a whole, the richest bryozoan habitats were found to occur in areas with hard substrata, suggestive of low sedimentation.
Multivariate statistical analyses of the recorded bryozoan data identified several clusters of samples (based on similar faunal content and colony growth form) that appear in accordance with particular sedimentary facies (Rust Citation2009). Three significant clustered bryozoan faunal associations are distinguished as relating to systems tracts, and described here as Wanganui Basin bryozoan biofacies 1, 2 and 3 (summarised in & ). These bryozoan-based associations lack the stratigraphic and taxonomic resolution of the distinctive local mollusc-based macrofossil palaeocommunities of Abbott & Carter (Citation1997) and Hendy & Kamp (Citation2004), however.
Table 3 Palaeoecological characteristics of three bryozoan-based, systems tract-related biofacies identified in the Wanganui Basin Plio-Pleistocene sequence. Bryozoan colony growth form categories (after Nelson et al. Citation1988) are discussed in text
Biofacies 1
At the base of each Plio-Pleistocene depositional sequence (above the basal unconformity representing low-stand sea level), onlap shellbeds and basal TST deposits formed in shoreline to shallow-shelf settings generally contain reworked and sorted bryozoan and molluscan material. Condensed, highly time-averaged, shelly onlap deposits from higher energy settings contain specimens affected by physical processes and display high frequencies of disarticulation, fragmentation, bryozoan encrustation of interior surfaces, valve ratio biasing (of molluscs), corrosion and/or abrasion (cf. Kidwell & Bosence Citation1991). The typically sandy or conglomeratic samples of bryozoan biofacies 1 from the Wanganui Basin are characterised by a moderate or low diversity of bryozoans. These include deposits from frequently ‘disturbed’ near-shore environments, which may be dominated by rapidly establishing taxa or ‘pioneer’-type bryozoan assemblages. Common encrusters include Antarctothoa tongima and Valdemunitella spp. In addition, a high proportion of disarticulated Cellaria fragments occur, and Otionellina is generally present. In samples from the Butlers Shell Conglomerate, Kaikokopu Formation, Okehu Shell Grit and Seafield Sand, erect bryozoan colonies are represented by occasional broken, disarticulated, transported and worn fragments (which are probably allochthonous material reworked and mixed into these TST units).
Biofacies 2
Shellgrounds were often developed with increasing sea-level and decreasing sedimentation up-sequence, forming stable substrata available for epibiont encrustation. Uppermost TST-MCS (mid-cycle or compound shellbed) samples such as the Lower Castlecliff Shellbed and Nukumaru Limestone are rich in representatives of bryozoan biofacies 2. They are characterised by a high diversity and abundance of in situ bryozoans with many specialised genera present, especially branching cyclostomes (such as Cinctipora, Diaperoecia and Hornera) which are common. Biofacies 2 samples from shellbeds exhibit in situ encrustation; typically, fragments of branching erect colonies are well preserved (e.g. Fig. 3C). These deposits tend to display higher proportions of articulated material in autochthonous position with little or no breakage, intact delicate forms and little or no biasing of mollusc valve ratios (cf. Kidwell & Bosence Citation1991), suggesting deposition in lower energy settings. Often evidence of the history of encrustation for an individual mollusc valve is preserved in a layered sequence of epibiont organisms, which may include a variety of interacting taxa including those from ‘climax’-type communities. In a form of positive taphonomic feedback (Kidwell & Jablonski Citation1983), bryozoan colonies themselves may support other epibiont taxa, including other bryozoans. Interactions between bryozoan colonies mostly take the form of overgrowth, where one occupant of a substratum grows over the surface of another (e.g. Jackson Citation1979; Barnes & Dick Citation2000). The relative abundance of these interactions reflects the presence of large, stable substrata (namely mollusc shells) and longevity of exposure. Examples of bryozoan encounter and overgrowth are quite common in biofacies 2 samples from Wanganui; 465 colony–colony interactions were recorded by Rust (Citation2009). Many factors influence the outcomes of bryozoan colony interference, as investigated by Jackson (Citation1979).
Biofacies 3
Higher in the sequences at Wanganui, sedimentation usually increased and thick siltstones and mudstones were deposited. In some of these HST units, bryozoans (and molluscs) are sparse (i.e. the ‘impoverished’ biofacies 3 fauna); in others they were not recorded at all. As sea level fell once more, RST deposits (where preserved) are often similar to those found in the lower TST (recording a return to bryozoan biofacies 1).
Discussion
Early workers were optimistic about the usefulness of bryozoans in determining palaeoenvironments (Stach Citation1937; Schopf Citation1969; Cuffey Citation1970). Summaries on the use of bryozoans for habitat interpretation have been provided by McKinney & Jackson (Citation1989), Smith (Citation1995) and Taylor (Citation2005).
Studies on the environmental requirements of modern bryozoans enable some interpretations of the ecology of past habitats. Plio-Pleistocene bryozoan palaeocommunities and biofacies recorded from Wanganui have modern counterparts on the New Zealand shelf, examples of habitats with the same species or closely related taxa occurring in what are considered equivalent environments to the fossil deposits. For example, species often found in low-sedimentation upper TST-MCS (biofacies 2) deposits at Wanganui, such as Cinctipora elegans, Celleporaria agglutinans, Cellaria immersa, Tubulipora sp. and Otionellina sp., are commonly encountered on the Otago Shelf (Probert & Wilson Citation1979; Batson & Probert Citation2000).
Use of bryozoans to determine water depth was tested in a study by Taylor et al. (Citation2004) who looked at four New Zealand species: Attinopora zealandica, Cinctipora elegans, Celleporaria emancipata and Diaperoecia purpurascens, all of which are common as fossils at Wanganui. However, these species were all found to have wide bathymetric ranges in modern habitats, limiting their usefulness in depth interpretation. Likewise, in a test of the relationship between depth and branch diameter in C. elegans, D. purpurascens and Erkosonea sp., correlation was found only in C. elegans and only at a low level of significance (Taylor et al. Citation2007).
Due to the broad bathymetric ranges of most modern relatives of Wanganui Basin bryozoan taxa, isolated use of their occurrence in samples to determine palaeodepth is cautioned. We note that extreme physical conditions (such as very high water energy) inhibit bryozoan growth; turbid and muddy environments are often avoided by most taxa (Taylor Citation2005). Sedimentation rates, substratum characters and taphonomic processes are all likely to have significantly influenced the distribution and preservation of bryozoans in the Wanganui Basin fossil record.
Habitat-tracking, migrating bryozoan faunas
The recurring pattern of bryozoan diversity and the cyclothem sequences at Wanganui suggest a relationship between bryozoan fauna and sedimentary facies. Distributional changes of bryozoans in the Wanganui Basin could be explained by a lateral shift, perhaps only of the order kilometres to tens of kilometres, of palaeocommunity zones or biofacies belts in response to shifting environments (i.e. ‘habitat tracking’; Brett Citation1998; Brett et al. Citation2007). During sea-level rise the zones of marine benthos, including the bryozoan palaeocommunities, typically move landwards (perpendicular to the shoreline) and shift back during regression. As commented by Brett et al. (Citation2007), the recurring community replacement patterns in the sequence result from parts of the fauna shifting into, or out of, the frame of reference provided by a local outcrop section. Such consistent patterns of replacement have been recorded for mollusc-based biofacies in some Wanganui Basin strata (e.g. Hendy & Kamp Citation2004). Through the Pliocene and Pleistocene sequences, this process has a recurrence interval correlating to the 41,000- or 100,000-year (6th and 5th order) glacio-eustatic cycles of sea-level changes, perhaps 30–120 m in amplitude (Naish et al. Citation2005).
We observe (see ) that as one passes up-section through the lower part (TST) of each Castlecliffian cyclothem (and corresponding sea-level rise), bryozoan biofacies 1 grades into biofacies 2 (upper TST-MCS) before being reduced to biofacies 3 near the deepest part of each cycle (HST). Due to erosion of the RSTs in many of these cycles, the sea-level-fall part of the sequence is often not preserved. Hence we see a sudden shift in the record back to biofacies 1 and the start of a new depositional cycle. The incomplete cyclic sequences therefore produce an ‘asymmetrical’ record of shifting faunas, an artefact of preservation (cf. Brett Citation1998).
In the overall Wanganui sequence the transgressive parts of each sea-level cycle, in particular the more advanced later stages, are favourable for many bryozoans (). The highstand and regressive parts of each cycle appear to be less favourable for bryozoan faunas, probably reflecting higher sedimentation rates. This mirrors the results seen for a number of molluscan faunas in the Wanganui Basin strata (e.g. Hendy & Kamp Citation2007). Plio-Pleistocene bryozoan faunas were probably linked to the migration of molluscs across the basin and changes in the mollusc taxa available as associated habitat/substrata through the sequence.
Biostratigraphic patterns
Some 29 molluscan genera are noted as having disappeared at the end of Nukumaruan time in the Wanganui Basin by Beu (Citation2004). Local extinctions are also apparent in the large dataset of Johnson & Curry (Citation2001), who noted a peak in the last occurrences of molluscs at the Late Pliocene (2.5–2.0 Ma). Beu (Citation2004, Citation2006), however, suggested the marine fauna of the Wanganui Basin was affected by changes in oceanic circulation, rather than directly by a climatic event.
Interestingly, cyclostome bryozoans appear more abundant and diverse taxonomically in the Pliocene shellbeds than those from the Pleistocene; cheilostomes are still dominant, however. Cyclostome diversity peaks in the Nukumaru Limestone, where 16 taxa are recognised (32% of the bryozoan diversity in that unit) and in the Wilkies Shellbed where 9 cyclostome taxa make up 36% of the bryozoan diversity. A possible Late Pliocene increase in the extinction rates of cyclostomes has been noted previously elsewhere (McKinney & Taylor Citation2001). Several cyclostomes do not reappear in the Wanganui Basin records after the Nukumaruan Stage. The endemic cinctiporid Cylindropora areolata is thought to have become extinct at the end of the Pliocene (Boardman et al. Citation1992). The robust erect branching specimens of Heteropora cf. napierensis and a fasciculiporid cyclostome are reasonably common in the Mangapanian shellbeds but have not been found post-Nukumaru Limestone. Cyclostomes Tetrocycloecia parapelliculata, Disporella wanganuiensis and Platonea sp. are among those notably more abundant in the Pliocene deposits. Modern populations of these latter taxa can still be found living on the New Zealand shelf today (Gordon & Taylor Citation2001; Gordon et al. Citation2009); hence their paucity during the Castlecliffian reflects local absence or decline in numbers rather than true extinction. The majority of Pleistocene oyster and bryozoan-rich shellbeds sampled probably represent slightly deeper settings than those from the Pliocene. It is uncertain just how much taxonomic variation is a simple consequence of increased depth. Other influences may relate to sedimentation in the Pleistocene beds; perhaps some of the cyclostome bryozoans were more susceptible to increased input of fine sediment.
In the Pliocene units, the thick prismatic right valve of the anomiid Patro undatus is often very abundant (see Beu & Maxwell Citation1990; Beu Citation2006) and is a significant host substratum for bryozoans (e.g. A, B). Most of the 50 species of bryozoan recorded from the Nukumaru Limestone were found to encrust Patro valves. Small juveniles of P. undatus appear to have been attached to hard substrata, while larger specimens may have been free-lying on the seabed in areas where current activity kept sedimentation rates low (Beu & Maxwell Citation1990; Beu Citation2006). For bryozoans, the Patro undatus beds in the Nukumaruan may have provided opportunities not matched by Ostrea chilensis in the Castlecliffian.
Johnson & Curry (Citation2001) noted a peak of molluscan first occurrences during the middle Pleistocene (0.75–0.5 Ma). Beu (Citation2006, Citation2004) noted a possible Mid-Pleistocene Transition (MPT) correlation with the first appearances of several mollusc taxa in the Wanganui Basin, including Pecten. The MPT interval also marks the return of many bryozoan taxa into the local fossil record, many of which have not been recorded since the macrofossil-rich Nukumaruan stage shellbeds (Rust Citation2009). Rather than a single event, the middle Pleistocene period is important here as it marks the return of more frequent in situ shellbed facies locally, sufficient to preserve the reoccupation of hard substrata in the Wanganui Basin by bryozoans. It is likely that the establishment of Pecten sp. in the basin (Upper Westmere Shellbed, OIS 19) was significant, its large valves providing an additional hard, shelly substratum upon which bryozoans could settle, grow and be preserved.
Conclusions
In the Wanganui Basin a diverse fauna of bryozoan colonies encrust hard substrata, especially mollusc valves, in cyclothemic deposits of near-shore and shelf origin. During the Plio-Pleistocene, the processes of sea level, climate and sediment-supply fluctuation that created the (5th and 6th order) depositional sequences had strong ecological impacts on the local benthic faunas including the colonial suspension-feeding bryozoans.
To summarise, palaeoecological characteristics of the bryozoan faunas (including diversity and the relative abundance of colony growth forms) vary through each depositional sequence in a recurring pattern of distribution related to lateral faunal migrations or habitat tracking across the Wanganui Basin. Rather than solely due to changes in water depth, this repetitive stratigraphic pattern is linked to many local environmental factors associated with cyclic sea-level fluctuation such as mollusc migration and hard substrate availability, sedimentation rates and water currents (all of which are likely to have influenced bryozoan colony growth).
Statistical analysis identified three recurring bryozoan-based biofacies that correlate with local sequence-stratigraphic systems tracts. Some bryozoan taxa and growth forms were found to characterise certain environments (e.g. free-living Otionellina spp. in shallow-shelf sandy settings), but most were not exclusive in their distribution. In terms of diversity, unilaminar encrusting species dominate the assemblages overall. Taphonomic effects on assemblages are evident, particularly those of biofacies 1 where bryozoan remains are typically disarticulated, transported and worn. In mid-cycle or compound shellbeds of biofacies 2, however, the in situ encrusting bryozoan colonies show little taphonomic alteration with frequent overgrowth interactions, and remains of erect-branching bryozoans such as Cinctipora, Diaperoecia and Hornera are important. Highstand siltstones of biofacies 3 contain only sparse bryozoan remains.
These recurring bryozoan-based biofacies and growth-form associations allow for general environmental interpretations, but are not necessarily strict indicators of habitat limitations for the specific taxa involved. We conclude that bryozoan remains can help define systems tract-related facies; rather than simply recording the presence/absence of most taxa, however, they are best used with sedimentological data and alongside other groups (mainly molluscs). We agree with Smith (Citation1995) that multiple lines of enquiry are advantageous for palaeoenvironmental reconstruction.
Other sequences have potential for comparison with the Wanganui Basin; strata from Hawkes Bay represent the contemporaneous faunas to the east and are rich in molluscs and bryozoans (Brown Citation1952). As additional Plio-Pleistocene faunas are examined with increasing detail, they will allow for sequence stratigraphic and biogeographic correlation. Currently, it appears that few bryozoan species are truly cosmopolitan; the modern New Zealand bryozoan fauna and the late Cenozoic record are largely endemic (Brown Citation1952; Gordon et al. Citation2009). Despite local periodic fluctuations related to faunal migration, the overall high diversity recorded from the Wanganui Basin, in addition to records from the Eocene of Oamaru (Lee et al. Citation1997) and the Paleocene–Eocene of Chatham Islands (Gordon & Taylor Citation1999), suggests that the rich diversity of bryozoans in the New Zealand region has existed through most of the Cenozoic.
Acknowledgements
Jack Grant-Mackie and Kathy Campbell supervised S. Rust's PhD project at the University of Auckland (with advisor K. Tilbrook), and commented on drafts of the manuscript. A University of Auckland PhD Scholarship helped fund the research. Dennis Gordon was supported by New Zealand Foundation for Research, Science & Technology Contract C01X0502. Many fruitful discussions were held with IBA members (P. Taylor, A. Smith, etc.). N. Hudson, L. Cotterall and C. Hobbis (University of Auckland) assisted in the preparation and presentation of fossil specimens. The manuscript was kindly proofread by D. R. Yanakopulos and reviewed by C. Reid.
Additional information
Notes on contributors
S Rust
Present address: 6702 S.H. 12,RD3, Kaikohe, 0473, New ZealandRelated Research Data
References
- Abbott , ST . 1997 . Foraminiferal paleobathymetry and mid-cycle architecture of mid-Pleistocene depositional sequences, Wanganui Basin, New Zealand . Palaios , 12 : 267 – 281 .
- Abbott , ST and Carter , RM . 1994 . “ The sequence architecture of mid-Pleistocene (0.35–0.95 Ma) cyclothems from New Zealand ” . In Orbital forcing and cyclic sequences , Edited by: de Boer , PL and Smith , DG . Vol. 19 , 367 – 394 . International Association of Sedimentologists Special Publication .
- Abbott , ST and Carter , RM . 1997 . Macrofossil associations from mid-Pleistocene cyclothems, Castlecliff, New Zealand . Palaios , 12 : 188 – 210 .
- Abbott , ST , Naish , TR , Carter , RM and Pillans , BJ . 2005 . Sequence stratigraphy of the Nukumaruan Stratotype (Pliocene-Pleistocene, c. 2.08–1.63 Ma), Wanganui Basin, New Zealand . Journal of the Royal Society of New Zealand , 35 : 123 – 150 .
- Barnes , DKA and Dick , MH . 2000 . Overgrowth competition in encrusting bryozoan assemblages of the intertidal and infra-littoral zones of Alaska . Marine Biology , 136 : 813 – 822 .
- Batson PB , Probert PK 2000 . Bryozoan thickets off Otago Peninsula . New Zealand Fisheries Assessment Report , Ministry of Fisheries , Wellington , 31 p.
- Beu , AG . 2004 . Marine Mollusca of oxygen isotope stages of the last 2 million years in New Zealand. Part 1: Revised generic positions and recognition of warm-water and cool-water migrants . Journal of the Royal Society of New Zealand , 34 ( 2 ) : 111 – 265 .
- Beu , AG . 2006 . Marine Mollusca of oxygen isotope stages of the last 2 million years in New Zealand. Part 2: Biostratigraphically useful and new Pliocene to Recent bivalves . Journal of the Royal Society of New Zealand , 36 : 151 – 338 .
- Beu , AG and Edwards , AR . 1984 . New Zealand Pleistocene and Late Pliocene glacio-eustatic cycles.” Palaeogeography, Palaeoclimatology . Palaeoecology , 46 : 119 – 142 .
- Beu , AG and Maxwell , PA . 1990 . Cenozoic Mollusca of New Zealand . New Zealand Geological Survey Paleontological Bulletin , 58 : 518
- Boardman , RS , McKinney , FK and Taylor , PD . 1992 . Morphology, Anatomy, and Systematics of the Cinctiporidae, new family (Bryozoa: Stenolaemata) . Smithsonian Contributions to Paleobiology , 70 : 81
- Bradstock , M and Gordon , DP . 1983 . Coral-like bryozoan growths in Tasman Bay and their protection to conserve commercial fish stocks . New Zealand Journal of Marine and Freshwater Research , 17 : 159 – 163 .
- Brett , CE . 1998 . Sequence stratigraphy, paleoecology, and evolution: biotic clues and responses to sea-level fluctuations . Palaios , 13 : 241 – 262 .
- Brett , CE , Hendy , AJW , Bartholomew , AJ , Bonelli , JRJ and McClaughlin , PI . 2007 . Response of shallow marine biotas to sea-level fluctuations: a review of faunal replacement and the process of habitat tracking . Palaios , 22 : 228 – 244 .
- Brown DA 1952 . The Tertiary Cheilostomatous Polyzoa of New Zealand. Trustees of the British Museum (Natural History) , London. 405 p.
- Carter , RM and Naish , TR . 1998 . A review of Wanganui Basin, New Zealand: global reference section for shallow marine, Plio-Pleistocene (2.5–0 Ma) cyclostratigraphy . Sedimentary Geology , 122 : 37 – 52 .
- Cook , PL and Chimonides , PJ . 1984 . Recent and fossil Lunulitidae (Bryozoa: Cheilostomata) 1. The genus Otionella from New Zealand . Journal of Natural History , 18 : 227 – 254 .
- Cooper RA 2004 . The New Zealand Geological Timescale . Institute of Geological and Nuclear Sciences Monograph 22 . 284 p.
- Cuffey , RJ . 1970 . Bryozoan–environment interrelationships: an overview of bryozoan paleoecology and ecology . Earth and Mineral Sciences , 39 : 41 – 45 .
- Fleming , CA . 1947 . Standard sections and subdivisions of the Castlecliffian and Nukumaruan Stages in the New Zealand Pliocene . Transactions of the Royal Society of New Zealand , 76 : 300 – 326 .
- Fleming , CA . 1953 . The Geology of Wanganui subdivision . New Zealand Geological Survey Bulletin , 52 : 362
- Gillespie , JL , Nelson , CS and Nodder , SD . 1998 . Post-glacial sea-level control and sequence stratigraphy of carbonate-terrigenous sediments, Wanganui shelf . New Zealand: Sedimentary Geology , 122 : 245 – 266 .
- Gordon , DP . 1984 . The marine fauna of New-Zealand: Bryozoa: Gymnolaemata from the Kermadec Ridge . New Zealand Oceanographic Institute Memoir , 91 : 198
- Gordon , DP . 1986 . The marine fauna of New Zealand: Bryozoa: Gymnolaemata (Ctenostomata and Cheilostomata Anasca) from the western South Island continental shelf and slope . New Zealand Oceanographic Institute Memoir , 95 : 121
- Gordon , DP . 1987 . “ The deep-sea Bryozoa of the New Zealand region ” . In Bryozoa: Present and Past , Edited by: Ross , JP . 97 – 104 . Western Washington University, Bellingham. .
- Gordon , DP . 1989 . The marine fauna of New Zealand: Bryozoa: Gymnolaemata (Cheilostomida Ascophorina) from the western South Island continental shelf and slope . New Zealand Oceanographic Institute Memoir , 97 : 158
- Gordon , DP . 1994 . Tertiary bryozoan genera in the present-day Australasian fauna-implications for classification and biogeography . Invertebrate Taxonomy , 8 : 283 – 298 .
- Gordon , DP and Taylor , PD . 1999 . Latest Paleocene to earliest Eocene Bryozoans from Chatham Island, New Zealand. Bulletin of the Natural History Museum . Geology Series , 55 : 1 – 45 .
- Gordon , DP and Taylor , PD . 2001 . New Zealand Recent Densiporidae and Lichenoporidae (Bryozoa: Cyclostomata) . Species Diversity , 6 : 243 – 290 .
- Gordon , DP , Stuart , IG and Collen , JD . 1994 . Bryozoan fauna of the Kaipuke Siltstone, northwest Nelson: a Miocene homologue of the modern Tasman Bay coralline bryozoan grounds . New Zealand Journal of Geology and Geophysics , 36 : 239 – 247 .
- Gordon , DP , Taylor , PD and Bigey , FP . 2009 . “ Phylum Bryozoa – moss animals, sea mats, lace corals ” . In The New Zealand Inventory of Biodiversity Volume 1 , Edited by: Gordon , D. P. 271 – 297 . Kingdom Animalia : Radiata, Lophotrochozoa, Deuterostomia. Canterbury University Press, Christchurch. .
- Hageman , SJ , Bone , Y , McGowran , B and James , NP . 1997 . Bryozoan colonial growth forms as paleoenvironmental indicators: evaluation of methodology . Palaios , 12 : 405 – 419 .
- Hammer O , Harper DAT , Ryan PD 2006 . PAST: Paleontological Statistics . http://folk.uio.no/ohammer/past/index.html (accessed 15 June 2006) .
- Hendy , AJW and Kamp , PJJ . 2004 . Late Miocene to early Pliocene biofacies of Wanganui and Taranaki Basins. New Zealand: applications to paleoenvironmental and sequence stratigraphic analysis . New Zealand Journal of Geology and Geophysics , 47 : 769 – 785 .
- Hendy , AJW and Kamp , PJJ . 2007 . Paleoecology of the Late Miocene-Early Pliocene Sixth-order Glacioeustatic Sequences in the Manutahi-1 Core, Wanganui-Taranaki Basin . New Zealand. Palaios , 22 : 325 – 337 .
- Jackson , JBC . 1979 . Overgrowth competition between encrusting cheilostome ectoprocts in a Jamaican cryptic reef environment . Journal of Animal Ecology , 48 : 805 – 823 .
- Jackson , JBC , Lidgard , S and McKinney , FK . 2001 . Evolutionary Patterns: Growth, form and tempo in the fossil record , 399 The University of Chicago Press, Chicago .
- Johnson , KG and Curry , GB . 2001 . Regional biotic turnover dynamics in the Plio-Pleistocene molluscan fauna of the Wanganui Basin, New Zealand . Palaeogeography Palaeoclimatology Palaeoecology , 172 ( 1–2 ) : 39 – 51 .
- Kamp , PJJ and McIntyre , AP . 1998 . The stratigraphic architecture of Late Pliocene (2.8–2.4 Ma) asymmetrical shelf sequences, western Wanganui Basin . New Zealand. Sedimentary Geology , 122 : 53 – 67 .
- Kamp , PJJ , Vonk , A , Bland , KJ , Hansen , RJ , Hendy , AJW , McIntyre , AP , Ngatahi , M , Cartwright , SJ , Hayton , S and Nelson , CS . 2004 . Neogene stratigraphic architecture and tectonic evolution of Wanganui, King Country, and eastern Taranaki Basins . New Zealand. New Zealand Journal of Geology and Geophysics , 47 : 625 – 644 .
- Kay , AM and Keough , MJ . 1981 . Occupation of patches in the epifaunal communities on pier pilings and the bivalve Pinna bicolour at Edithburg . South Australia. Oecologia , 48 : 123 – 130 .
- Kidwell , SM and Jablonsky , D . 1983 . “ Taphonomic feedback: ecological consequences of shell accumulation ” . In Biotic interactions in Recent and fossil benthic communities , Edited by: Tevesz , MJS and McCall , PL . 195 – 248 . New York : Plenum Press .
- Kidwell , SM and Bosence , DW . 1991 . “ Taphonomy and time-averaging of marine shelly faunas ” . In Taphonomy: Releasing the data locked in the fossil record , Edited by: Allison , PA and Briggs , DEG . 115 – 209 . New York : Plenum Press .
- Kondo , Y , Abbott , ST , Kitamura , A , Kamp , PJJ , Naish , TR , Kamataki , T and Saul , GS . 1998 . The relationship between shellbed type and sequence architecture: examples from Japan and New Zealand . Sedimentary Geology , 122 : 109 – 127 .
- Lee , DE , Scholz , J and Gordon , DP . 1997 . Paleoecology of a Late Eocene mobile rockground biota from North Otago, New Zealand . Palaios , 12 : 568 – 581 .
- Lidgard , S , McKinney , FK and Taylor , PD . 1993 . Competition, clade replacement, and a history of cyclostome and cheilostome bryozoan diversity . Paleobiology , 19 : 352 – 371 .
- McKinney , FK . 1996 . Encrusting organisms on co-occurring disarticulated valves of two marine bivalves: comparison of living assemblages and skeletal residues . Paleobiology , 22 : 543 – 567 .
- McKinney , FK and Jackson , JBC . 1989 . Bryozoan Evolution. Special topics in paleontology , 238 Unwin Hyman, Boston .
- McKinney , FK and Taylor , PD . 2001 . Bryozoan generic extinctions and originations during the last one hundred million years . Palaeontologica Electronica , 4 : 1 – 26 .
- Naish , TR , Abbott , ST , Alloway , BV , Beu , AG , Carter , RM , Edwards , AR , Journeaux , TD , Kamp , PJJ , Pillans , BJ , Saul , G and Woolfe , K J . 1998 . Astronomical calibration of a southern hemisphere Plio-Pleistocene reference section, Wanganui Basin . New Zealand. Quaternary Science Reviews , 17 : 695 – 710 .
- Naish , TR , Field , BD , Zhu , H , Melhuish , A , Carter , RM , Abbott , ST , Edwards , S , Alloway , BV , Wilson , GS , Niessen , F , Barker , A , Browne , GH and Maslen , G . 2005 . Integrated outcrop, drill core, borehole and seismic stratigraphic architecture of a cyclothemic, shallow-marine depositional system, Wanganui Basin, New Zealand . Journal of the Royal Society of New Zealand , 35 : 91 – 122 .
- Nelson , CS , Hyden , FM , Keane , SL , Leask , WL and Gordon , DP . 1988 . Application of bryozoan zoarial growth-form studies in facies analysis of non-tropical carbonate deposits in New Zealand . Sedimentary Geology , 60 : 301 – 322 .
- Pillans , BJ , Alloway , BV , Naish , TR , Westgate , J , Abbott , ST and Palmer , AS . 2005 . Silicic tephras in Pleistocene shallow-marine sediments of Wanganui Basin, New Zealand . Journal of the Royal Society of New Zealand , 35 : 43 – 90 .
- Probert , PK and Wilson , JB . 1979 . Epibenthic macrofauna off southeastern New Zealand and mid-shelf bryozoan dominance . New Zealand Journal of Marine & Freshwater Research , 13 : 379 – 392 .
- Proust , J-N , Lamarche , G , Nodder , SD and Kamp , PJJ . 2005 . Sedimentary architecture of a Plio-Pleistocene proto-back-arc basin: Wanganui Basin . New Zealand. Sedimentary Geology , 181 : 107 – 145 .
- Rust S 2009 . Plio-Pleistocene bryozoan faunas of the Wanganui Basin, New Zealand . Unpublished PhD thesis , University of Auckland , Auckland, , New Zealand . 438 p. plus appendices .
- Schopf , TJM . 1969 . Paleoecology of ectoprocts (bryozoans) . Journal of Paleontology , 43 : 234 – 244 .
- Smith , AM . 1995 . “ Palaeoenvironmental interpretation using bryozoans: a review ” . In Marine Palaeoenvironmental Analysis from Fossils. Geological Society Special Publication 83 , Edited by: Bosence , DW and Allison , PA . 231 – 243 . London .
- Smith , AM , Nelson , CS and Spencer , HG . 1998 . Skeletal carbonate mineralogy of New Zealand Bryozoans . Marine Geology , 151 : 27 – 46 .
- Stach , LW . 1937 . The Application of the Bryozoa in Cainozoic Stratigraphy . Report of the 23rd Meeting Australian and New Zealand Association for the Advancement of Science , 23 : 80 – 83 .
- Taylor , PD . 2005 . Bryozoans and Palaeoenvironmental Interpretation . Journal of the Palaeontological Society of India , 50 : 1 – 11 .
- Taylor , PD and Wilson , MA . 2003 . Palaeoecology and evolution of marine hard substrate communities . Earth Science Reviews , 62 : 1 – 103 .
- Taylor , PD , Schembri , PJ and Cook , PL . 1989 . Symbiotic associations between hermit crabs and bryozoans from the Otago region, southeastern New Zealand . Journal of Natural History , 23 : 1059 – 1085 .
- Taylor , PD , Gordon , DP and Batson , PB . 2004 . Bathymetric distributions of modern populations of some common Cenozoic Bryozoa from New Zealand, and paleodepth distribution . New Zealand Journal of Geology and Geophysics , 47 : 57 – 69 .
- Taylor , PD , Kuklinski , P and Gordon , DP . 2007 . Branch diameter and depositional depth in cyclostome bryozoans: testing a potential paleobathymetric tool . Palaios , 22 : 220 – 224 .
- Thomson , JA . 1916 . On stage names applicable to the divisions of the Tertiary in New Zealand . Transactions of the New Zealand Institute , 48 : 28 – 40 .
- Turner , GM , Kamp , PJJ , McIntyre , AP , Hayton , S , McGuire , DM and Wilson , GS . 2005 . A coherent middle Pliocene magnetostratigraphy, Wanganui Basin, New Zealand . Journal of the Royal Society of New Zealand , 35 : 197 – 227 .
- Ward , MA and Thorpe , JP . 1989 . Assessment of space utilisation in a sub-tidal temperate bryozoan community . Marine Biology , 103 : 215 – 224 .
- Ward , MA and Thorpe , JP . 1991 . Distribution of encrusting bryozoans and other epifauna on the subtidal bivalve Chlamys opercularis . Marine Biology , 110 : 253 – 259 .
- Waters AW 1887 . On Tertiary cyclostomatous Bryozoa from New Zealand . Quarterly Journal of the Geological Society of London 43 : 337 – 350 , pl. 18.