Abstract
The Late Miocene–Early Pleistocene (Tongaporutuan–Nukumaruan) sedimentary succession in the forearc basin in central and western Hawke's Bay, encompassed by the Tolaga and Mangaheia Groups, comprises a wide variety of variably fossiliferous lithofacies, ranging from non-marine greywacke-derived conglomerates to bathyal mudstone and flysch beds. Thirty molluscan biofacies, inferred to have accumulated in estuarine to outer shelf palaeoenvironments, have been identified and represent both in situ and transported assemblages. Although distributed through the late Neogene succession, most biofacies occur within the Late Pliocene–Early Pleistocene part of the Mangaheia Group. Using the spatial distribution of the different biofacies, we have constructed detailed palaeoenvironmental reconstructions for the Plio-Pleistocene of central Hawke's Bay. The stratigraphic occurrence of particular biofacies has been primarily controlled by relative sea-level positions and variations in sediment input during high-frequency glacio-eustatic sea-level oscillations. The distribution of biofacies reflects proximity to contemporaneous shorelines, localised sources of carbonate sediment or dominance of siliciclastic sedimentation, and records the interplay between tectonic and eustatic drivers of relative sea-level change.
Introduction
Shallow-marine macrofossil associations are the preserved remnants of palaeocommunities, and their occurrence usefully aids the identification of palaeoenvironmental change through geological time (Brett Citation1998; Brett et al. Citation2007). In particular, such biofacies have found wide usage in evaluating variations in marine palaeoenvironments within depositional systems of sedimentary sequences, helping distinguish transgressive (TST), highstand (HST), regressive (RST) and lowstand (LST) systems tracts (e.g. Abbott & Carter Citation1997; Beu & Kitamura Citation1998; Hendy & Kamp Citation2004, Citation2007). Understanding the spectrum and environmental preferences of biofacies enable the rates and magnitudes of palaeoenvironmental change to be estimated quantitatively. Furthermore, where strata are preserved along and across depositional strike, biofacies may be used to elucidate spatial variations in water depth and types of sediment, factors that are key to interpreting sedimentary basin evolution.
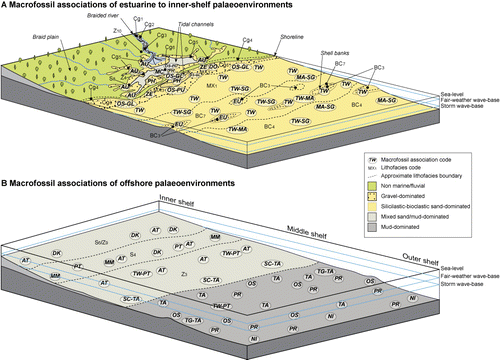
This study presents the results of biofacies analysis from late Neogene rocks cropping out in central and western Hawke's Bay. Macrofossil biofacies of late Neogene successions in Taranaki and Wanganui basins in western North Island have been assessed in some detail previously, particularly with respect to their palaeoenvironmental significance (e.g. Abbott & Carter Citation1997; Kondo et al. Citation1998; Hendy & Kamp Citation2004). However, this is the first time that such a study has been undertaken on age-equivalent rocks in eastern North Island. The succession of interest is of Late Miocene–Early Pleistocene age (Tongaporutuan–Nukumaruan) and encompasses the Tolaga and Mangaheia Groups (Bland Citation2006; Bland et al. Citation2007; Lee et al. Citation2011). The lithofacies identified within this succession and their inferred environments of deposition are summarised in ; full details are described in Bland (Citation2006). The primary focus of this paper is the character of the molluscan biofacies preserved within the Pliocene–Early Pleistocene Mangaheia Group, a shelf-dominated succession that occurs extensively throughout Hawke's Bay (, ; e.g. Bland et al. Citation2007; Lee et al. Citation2011) and continues north to the Raukumara Peninsula (e.g. Mazengarb et al. Citation1991; Mazengarb & Speden Citation2000). Due to their widespread distribution in outcrop and fossil-rich horizons, special attention is given here to the Mangapanian–Nukumaruan-aged Pohue, Matahorua, Petane, and Okauawa formations (). The stratigraphy of this area was described in detail by Bland et al. (Citation2007), and is supported by a series of 1:50 000 geological maps in Bland (Citation2006) and stratigraphic columns and interpretations in Kamp et al. (Citation2007); reference is made to some of these columns in this paper. The regional geology of the study area is presented in Lee et al. (Citation2011).
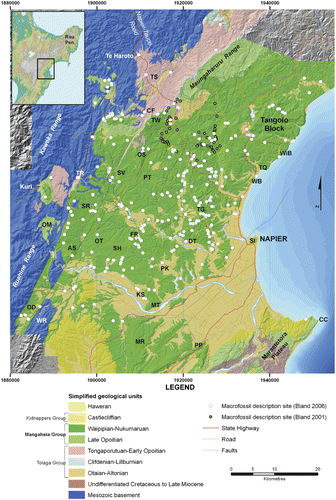
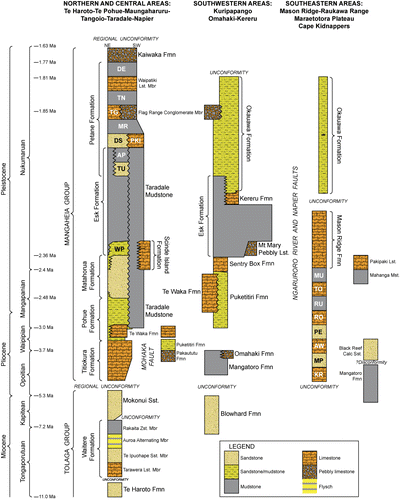
Table 1 Summary of lithofacies in the Neogene succession cropping out in western and central Hawke's Bay (refer to Bland Citation2006 for discussion on individual lithofacies). Lithofacies are grouped into siliciclastic, bioclastic or mixed bioclastic-siliciclastic groups. Siliciclastic lithofacies are further sub-divided into four assemblages, primarily on the basis of grain size (sandstone, siltstone, conglomerate and volcaniclastic assemblages). A, abundant; C, common; R, rare; S, some; UC, uncommon. Faunal associations are summarised in .
Macrofossil assemblages preserved in the study area reflect tectonically and glacio-eustatically driven controls on the distribution of sedimentary lithofacies (Haywick Citation1990; Beu Citation1995; Caron et al. Citation2004a; Bland et al. Citation2008a). The documentation of the various macrofossil assemblages in the Hawke's Bay area therefore allows some controls to be placed on the geological evolution of this region. Central Hawke's Bay presently occupies a forearc position within the convergent Hikurangi subduction margin of eastern North Island (Ballance Citation1993; Field & Uruski et al.1997; Lee et al. Citation2011). The forearc basin, which has developed in this region since the Middle Miocene, lies between the inboard part of the accretionary prism that involves the hill country south of Cape Kidnappers and the high country of the axial ranges in the west (Ruahine, Kaweka and Ahimanawa ranges; ) (Pettinga, Citation1982; Lewis & Pettinga Citation1993). The growth of these tectonostratigraphic features during the late Neogene has exerted a strong control on the distribution of biofacies and their enclosing rocks in central Hawke's Bay (e.g. Kamp et al. Citation1988; Beu Citation1995; Caron et al. Citation2004a,Citationb; Bland et al. Citation2008a; Trewick & Bland Citation2012). The basin contains a siliciclastic-dominated Neogene succession >5 km thick that was deposited in palaeoenvironments ranging from braided river systems to deep marine slope fans (Grindley Citation1960; Field & Uruski et al. Citation1997; Bland et al. Citation2007; Lee et al. Citation2011).
Data acquisition and analysis
Macrofaunal data were acquired during the course of geological mapping and stratigraphic logging (Bland Citation2006; for stratigraphic columns see Kamp et al. Citation2007) from c. 350 outcrops (), or in the laboratory from analysis of 111 bulk samples. Map co-ordinates in this paper are presented in New Zealand Transverse Mercator (NZGD 2000) units, converted from New Zealand Map Grid 260 series units. All identification, census counts and collation of data from bulk samples were undertaken by a single worker, following the methodology of Hendy & Kamp (Citation2004). Field observations provide a combination of presence-absence and semi-quantitative data for assemblages across central Hawke's Bay (). As far as possible, the identification and relative abundances of macrofossils were documented at the outcrop (see appendix data in Bland Citation2006). Specimens that could not be identified in the field were brought back to the laboratory. The determination of biofacies from field observations is a legitimate process for the identification of biofacies where bulk sampling is not possible (Hendy & Kamp Citation2004, Citation2007). This is often necessary because macrofossils are sparsely distributed through a unit making collection of a bulk sample impractical, or the rock is so cemented that a suitable sample cannot be removed.
Where bulk sample collection was achievable, as many lithofacies as possible were sampled. However, the dataset reveals that samples from coarse-grained shell-rich beds of Mangapanian and Nukumaruan age are over-represented. The reasons for this are twofold. First, the highly fossiliferous nature of most of these horizons makes them obvious targets for sampling. Second, most of these horizons are more cemented than their siliciclastic counterparts; they are therefore better exposed in the landscape, making them more accessible to investigation and collection. See Hendy (Citation2009) and Sessa et al. (Citation2009) for a discussion of the biases imposed on diversity and palaeoecological analyses due to lithification of host sediments.
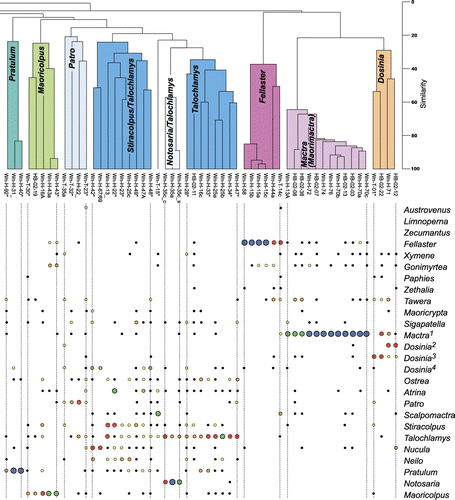
Analyses of bulk samples were based mainly on census counts of bivalves and gastropods, but also included brachiopod and echinoderm specimens. Fragments of bivalves were only counted if they included the umbo, or if the complete posterior and anterior margins were present. For gastropods, specimens were only counted if the aperture was present. The number of disarticulated bivalve and brachiopod specimens was halved, as bivalved organisms can produce twice the number of countable skeletal parts compared to other groups such as gastropods (Gilinsky & Bennington Citation1994). Echinoderm ossicle counts were divided by five, to partly account for the multiple skeletal elements that these taxa contribute to death assemblages (Hendy & Kamp Citation2004). Census count data were analysed using Q-mode cluster analysis to determine biofacies using the computer programme PRIMER, following the methodology of Hendy & Kamp (Citation2004). The Q-mode analysis used the arithmetic averages of a Bray–Curtis distance matrix to produce a dendrogram classification of samples ().
Figure 3 Q-mode cluster analysis of Pleistocene (Nukumaruan) macrofossil bulk samples in the study area. These clusters form the basis for identifying many of the macrofossil associations in Mangaheia Group in western and central Hawke's Bay.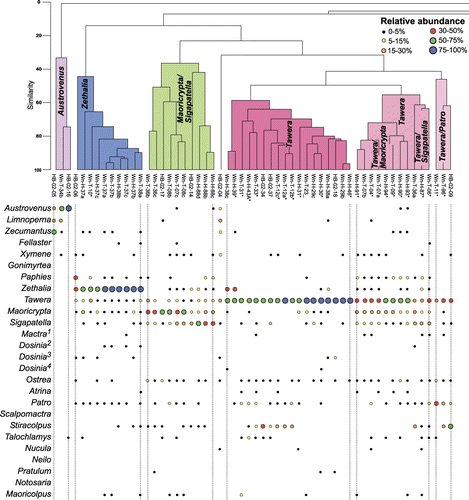
Macrofossils were identified to the finest taxonomic scale possible. This was usually to species level, but at many localities the degree of cementation meant that identification to genus level only was possible. Consequently, data analysis in this study was undertaken where possible at genus or subgenus level. Taxonomic nomenclature follows Beu & Maxwell (Citation1990) and more recent revisions (e.g. Beu Citation2004, Citation2006). Interpretations of the inferred environment of accumulation of the molluscan biofacies have been determined from a combination of macrofaunal autecology, sedimentary structures and lithofacies of the enveloping sedimentary rocks and, where possible, comparisons of the association with modern equivalents. Because of the geologically young age of the Mangaheia Group, many taxa and biofacies in the study area have modern analogues with which direct comparisons can be made (e.g. Morton & Miller Citation1968; McKnight Citation1969; Luckens Citation1972; Beu & Kitamura Citation1998).
Data on fossils collected during this study are lodged in the New Zealand Fossil Record Electronic Database (FRED). More detailed descriptions of biofacies and lithofacies within the study area are contained in Bland (Citation2006), which can be downloaded from the University of Waikato Research Commons (http://researchcommons.waikato.ac.nz/handle/10289/2222).
Results
Thirty faunal associations and six sub-associations were identified (), reflecting the large variety of lithofacies in the study area and the wide age range of the Late Miocene–Early Pleistocene succession studied. The reader is referred to Bland (Citation2006) for more detailed descriptions of these associations, including discussion of their palaeoenvironmental assignments.
Table 2 Summary of Neogene macrofaunal associations identified in this study for Late Miocene–Pleistocene rocks cropping out in western and central Hawke's Bay. Biofacies defined from field observations are denoted †. Lithofacies are summarised in . Tt, Tongaporutuan (early Late Miocene); Wo, Opoitian (Early Pliocene); Wm, Mangapanian (Late Pliocene); Wn, Nukumaruan (Early Pleistocene). Refer to Bland (Citation2006) for additional discussion and supporting evidence for interpretations.
Sixteen of these associations and sub-associations were identified from Q-mode cluster analysis (comprising three or more samples; ), while the remainder were defined from field observations where bulk sampling was not possible. The faunal associations are represented by a combination of in situ (life position) and within-habitat time-averaged assemblages. We infer that the faunal composition of the biofacies has suffered little in the way of modification from taphonomic processes.
Three major subsets can be delineated on the basis of the abundance of the bivalve genera Dosinia/Mactra/Fellaster, Tawera and Talochlamys. The Tawera-dominated subset consists of three discrete clusters characterised by: (1) the overall dominance of Tawera subsulcata; (2) the dominance of Tawera subsulcata with common Maoricrypta profunda and Sigapatella novaezelandiae (Tawera-Maoricrypta sub-association); and (3) the dominance of Tawera subsulcata with common Patro undatus and Sigapatella spp (Tawera-Sigapatella sub-association).
The Talochlamys-dominated subset consists of three smaller clusters characterised by: (1) ubiquitous Talochlamys gemmulata; (2) common Talochlamys gemmulata and Stiracolpus spp.; and (3) common Talochlamys gemmulata and the brachiopod Notosaria nigricans. Nine smaller subsets are recognised that are distinctly separated from the larger subsets. These include clusters of samples dominated by Dosinia, Mactra (Maorimactra), Fellaster, Patro, Maoricolpus, Pratulum, Maoricrypta-Sigapatella, Zethalia and Austrovenus. With the exception of the Patro and Pratulum-dominated clusters, most of these smaller subsets are present in shallow-water sandstone, conglomerate and shellbed lithofacies (, ). The Pratulum and Patro associations are typical of offshore (shelf) siltstone to silty sandstone lithofacies.
In the following sections we describe the biofacies identified in this study, their constituent taxa and their inferred environments of accumulation. They are grouped according to their general depositional settings, from those of estuarine/shoreface environments through to nearshore/inner shelf settings and finally associations more representative of outer shelf environments.
Biofacies of estuarine and shoreface palaeoenvironments
Most of these associations have fairly low faunal diversity due to their restricted inferred palaeoenvironmental positions (e.g. ). The Austrovenus association (AU) is dominated by the extant species Austrovenus stutchburyi, the New Zealand ‘cockle’, which is extremely abundant in semi-enclosed embayments and estuaries around the modern New Zealand coastline (Beu & Maxwell Citation1990; Beu Citation2006). The Austrovenus association is therefore inferred to have been deposited in an estuarine environment on a substrate of silt to silty sand. The association typically has a low to monospecific faunal diversity reflecting the inferred estuarine depositional setting of the association.
The Paphies association (PH) is similar to the Austrovenus association in its low diversity, although the former is almost entirely dominated by Paphies australis (the modern-day ‘pipi’). This species is presently abundant in very shallow-water intertidal environments such as the Ahuriri Lagoon at Napier, the Firth of Thames and Tauranga Harbour, where it has a preference for tide-race channels (A.G. Beu, pers. comm., 2009). The Paphies association probably accumulated in marginal-marine to shoreface settings and is often represented by a combination of in situ and reworked valves. In situ valves are inferred to have accumulated in highly fossiliferous shell banks in outer estuarine and upper to middle parts of semi-enclosed embayments, consistent with the modern distribution of Paphies australis. Such palaeoenvironments are inferred to represent the submergence of a fluvial fan-delta system during the early stage of eustatic sea-level rise. The Paphies association is restricted in outcrop to the Glengarry Road (BJ39/240282, column 96) and Hedgley Station areas (BJ39/266299, column 98) east of Dartmoor. Lithofacies containing the Paphies association are usually very pebbly, although sandy lithofacies occur in the Napier–Taupo Road area (e.g. BJ38/212422). The association is observed to pass laterally into the shallow-water Zethalia, Eumarcia and Glycymeris associations.
The Fellaster association (FE), dominated by the echinoid ‘sand dollar’ Fellaster zelandiae, is considered to represent a less-restricted marine setting than either the Austrovenus or Paphies associations. The Fellaster association probably occupied near-fully marine nearshore to shoreface environments, in keeping with its modern distribution (Grace Citation1966; McKnight Citation1969; Grange Citation1979). In most localities, the Fellaster association occurs within scour-based event-concentrated shell lenses enclosed by well-sorted sandstone (lithofacies S3), indicating that it is usually a transported association in the Mangaheia Group. We infer the scour-base of shell lenses containing the Fellaster association represents remobilisation of sediments during storm events.
The Zethalia association (ZE) is most common in sandy to pebbly lithofacies of Mangapanian and Nukumaruan age. Fauna are dominated by Zethalia coronata (Wm rocks) or Z. zelandica (Wn rocks). Lithofacies containing the Zethalia association are typically sandy to pebbly (Tables 1, 2). Although the Zethalia association is most common in fossiliferous conglomerate beds, it also occurs in many sandstone beds where taxa are concentrated in reworked event-concentrated shell lenses. The sandy and pebbly lithofacies are interpreted as having accumulated in tidal channels within enclosed bays or in nearshore environments, proximal to a river system that was actively supplying large volumes of greywacke pebbles (Bland Citation2006). In modern environments, Zethalia zelandica is abundant in sandy environments on open marine beaches at 3–5 m water depth (Beu & Maxwell Citation1990). The Zethalia association therefore likely accumulated in a shoreface to nearshore setting, and probably on coarser-grained substrates and in more exposed settings than the Fellaster association.
Biofacies of enclosed embayment, nearshore and inner shelf palaeoenvironments
Many associations representing palaeo-water depths of c. 5–50 m have been identified in this study. The Maoricrypta-Sigapatella association (MA-SG) is dominated by Maoricrypta profunda and Sigapatella novaezelandiae, and is particularly prominent in the Waipatiki Limestone Member cropping out in the Matapiro and Pukehamoamoa areas, to the west of Hastings (see geological maps in Bland Citation2006 and Lee et al. Citation2011). It is inferred that the association passes laterally into the Tawera association and its sub-associations (see below), and accumulated on a seafloor with a coarser-grained substrate than the Tawera association. The Maoricrypta-Sigapatella association probably accumulated at nearshore to inner shelf depths on a firm-ground substrate of coarse-grained shellhash subjected to a strong tidal regime that is inferred to have kept the seafloor relatively free of fine-grained siliciclastic sediment.
The Tawera association (TW), one of the most common faunal associations in Nukumaruan Mangaheia Group rocks (Bland Citation2006), is dominated by the extinct ‘venus clam’ Tawera subsulcata along with common Ostrea chilensis. The latter becomes more common in progressively offshore sub-associations and it is likely that the Tawera association grades into the Ostrea association. The Tawera association is most often observed in sandy to coarse-grained, moderately- to highly-fossiliferous shelly and sandy lithofacies, although it is also common in coarse-grained shelly conglomerate, fine sandstone and sandy siltstone. At some localities the Tawera association is represented by fossils preserved in life position or with minimal reworking and it passes laterally into the transported accumulations of the Maoricrypta-Sigapatella association, among others. Tawera subsulcata is extremely abundant in the fossil record and is characteristic of very shallow-water lithofacies (Beu & Maxwell Citation1990). The Tawera association described here probably accumulated on a sandy to shell-gravel substrate subject to strong tidal or wave influence in either very localised settings such as tidal channels in partially enclosed embayments, or offshore from sandy or gravelly beach settings at inner-shelf (10–50 m) water depths.
Three sub-associations have been identified within the Tawera association. The Tawera-Sigapatella sub-association (TW-SG) is restricted to coarse-grained, late Nukumaruan limestone beds of the Petane Formation, and in many places is observed to grade vertically and laterally into assemblages of the Tawera-Maoricrypta sub-association. The biofacies has moderately high faunal diversity, but is characterised by Tawera subsulcata with common Sigapatella novaezelandiae and Maoricrypta profunda. A tide-dominated nearshore to innermost-shelf depositional environment with a coarse-grained shell-gravel substrate is inferred for the Tawera-Sigapatella sub-association. The significant amount of fragmentation and abrasion observed on skeletal material indicates sediment winnowing and condensation, and suggests that the association comprises reworked assemblages. Deposition occurred above storm wave-base, allowing siliciclastic sediments to be winnowed from bioclastic debris. This sub-association is observed in many shellbed lithofacies in the Waipatiki Limestone Member in the Matapiro area, and near Waipunga Road in the eastern Tangoio Block.
The Tawera-Maoricrypta sub-association (TW-MA) has been observed only in coarse-grained highly fossiliferous limestone beds of late Nukumaruan age. It is characterised by the dominance of Tawera subsulcata and common Maoricrypta profunda. Sigapatella novaezelandiae is also common, and where it dominates over Maoricrypta the association is classified as the Tawera-Sigapatella sub-association. The Tawera-Maoricrypta association probably accumulated in a tide-dominated nearshore to innermost shelf setting on a substrate of coarse shell-gravel under the influence of tidal and wave currents, in a position below the lowstand shoreline. The Tawera-Maoricrypta sub-association passes laterally into the Maoricrypta-Sigapatella, Tawera-Sigapatella, Tawera and Zethalia associations. The Tawera-Maoricrypta sub-association occurs in lithofacies comprising almost exclusively coarse shell-gravel with a limited contribution from siliciclastic sand and gravel (). As with other Tawera associations, it is inferred that tidal currents played a strong influence in the accumulation of this sub-association, keeping the seafloor relatively free of siliciclastic sediments and also reworking shell debris.
By contrast, the Tawera-Patro (TW-PT) sub-association is characterised by the co-dominance of Tawera subsulcata and the extinct false oyster Patro undatus. It has been defined from cluster analysis of bulk samples collected from sandy to silty lithofacies (e.g. Z3, Z4), and occurs sporadically through the study area. It is inferred to have accumulated at lower-inner to upper-middle shelf water depths on a semi-firm-ground sandy to silty substrate, in an environment slightly further offshore than other Tawera sub-associations.
The Dosinia association (DO) occurs within sandy lithofacies, as opposed to the shell-gravel lithofacies typical of most of the Tawera-dominated associations. It is characterised by the occurrence of the extant bivalves Dosinia (Phacosoma) subrosea and Dosinia (Austrodosinia) anus. Today, Dosinia (Phacosoma) subrosea commonly lives near the low tide mark in a variety of soft-bottom lithofacies off sandy beaches in enclosed embayments and, less commonly, on open coasts. Dosinia (Austrodosinia) anus is an open-ocean sandy beach species (Beu & Maxwell Citation1990). The Dosinia association is inferred to have accumulated on sandy substrates at shoreface to inner shelf depths (1–30 m). The Dosinia association is most common in well-sorted, clean sandstone lithofacies, although it has been recognised in sandy interbeds in conglomeratic lithofacies of the Flag Range Conglomerate Member (Petane Formation) in Kikowhero Stream (BK38/092110). Valves in this association are typically arranged in a disarticulated, concave-down pattern in moderately to highly fossiliferous, scour-based event-concentrated shellbeds (Bland Citation2006). Concentrations of valves probably occurred during storm events when sediments were winnowed from around in situ shells with a minimal degree of remobilisation of the valves themselves. Evidence of deposition in a tidal channel within a semi-enclosed embayment for some beds of the Dosinia association is present in Kikowhero Stream, on the basis of enclosing lithofacies (Bland Citation2006). In most other cases accumulation of this association occurred in open-marine settings.
The Maoricolpus association (MC) is dominated by the extant gastropod Maoricolpus roseus, a species widespread all over the modern New Zealand shelf including in very shallow water depths such as intertidal environments in Manukau Harbour (Beu & Maxwell Citation1990). Powell (Citation1937) and McKnight (Citation1969) described a modern Maoricolpus association from the Waitemata and Manukau harbours, which lives on hard-bottomed shelly to sandy substrates composed of coarse shell debris redeposited from shallower environments by tidal currents, at water depths of 5–20 m. Such a depositional setting is inferred for the accumulation of the Maoricolpus association. It is suggested that the Maoricolpus association accumulated in a more shoreward setting in association with strong tidal currents than the Tawera association. The Maoricolpus association is observed to pass laterally into the Glycymeris, Tawera and Maoricrypta-Sigapatella (sub)-associations.
The Ostrea-glycymeridassociation (OS-GL) occurs in rocks of Opoitian to late Nukumaruan age and is characterised by an abundance of the extant ‘bluff oyster’ Ostrea chilensis in association with glycymerid bivalves such as Glycymeris spp. and Tucetona laticostata. It occurs most commonly in coarse-grained pebbly to conglomeratic lithofacies, and passes laterally into several other shallow-water faunal associations such as the Austrovenus, Maoricolpus, Ostrea-Purpurocardia, Eumarcia, Tucetona and Tawera associations. The Ostrea-glycymerid association probably accumulated at nearshore to innermost shelf water depths on a firm sandy to gravelly substrate, as indicated by the modern distributions of similar assemblages (e.g. Beu & Maxwell Citation1990). Strong tidal currents kept the seafloor relatively free of siliciclastic sediment. Abraded valves of estuarine restricted bivalves such as Austrovenus stutchburyi and Paphies australis are common in this association at many places and suggest close proximity to estuaries during the period of accumulation.
The Ostrea-Purpurocardia association (OS-PU) is common and widespread in conglomerate lithofacies of late Nukumaruan age in central and southern parts of the study area, especially within the Flag Range Conglomerate Member. Faunal diversity is high, with most species comprising robust, thick-shelled, semi-infaunal and epifaunal species such as Purpurocardia purpurata, Lutraria grandis and Alcithoe arabica, with less common Glycymeris shrimptoni and Tucetona laticostata. Lithofacies containing the Ostrea-Purpurocardia association are exclusively conglomeratic (lithofacies Cg6, MX3). The association likely accumulated in nearshore to innermost shelf environments on gravelly substrates, where strong tidal currents kept the seafloor relatively free of fine siliciclastic sediment.
The Eumarcia association (EU) is characterised by an abundance of the large extinct bivalve Eumarcia plana, and is one of the most distinctive Nukumaruan associations in the basin. The association contains a wide variety of extant shallow-water marginal-marine to nearshore taxa such as Paphies australis, Fellaster zelandiae, Mactra discors, Zethalia spp., Circomphalus spp., Dosinia (Fallartemis) lambata, Glycymeris shrimptoni, Gari lineolata, Scalpomactra scalpellum, Divaricella cumingi and Myadora waitotarana. Other notable molluscs present include Purpurocardia purpurata and Tawera subsulcata. The Eumarcia association probably passes laterally into the Austrovenus, Fellaster and Zethalia associations in a shoreward direction and the Ostrea-glycymerid, Maoricrypta-Sigapatella and Tawera associations in an offshore direction. The Eumarcia association likely accumulated in a shallow-water, nearshore to innermost shelf environment under a regime of strong tidal currents. The Maoricolpus association probably accumulated in a similar setting to the Eumarcia association, but tends to occur in lithofacies with coarser-grained conglomeratic sediment.
The Tucetona association (TU) is most common in Early Pliocene rocks adjacent to the Kaweka Range (Bland et al. Citation2003, Citation2007; Bland Citation2006) and is characterised by the abundance of the extant dog cockle Tucetona laticostata. The association contains a moderately diverse range of robust epifaunal and semi-infaunal taxa including Ostrea chilensis, Crassostrea ingens, Oxyperis elongata, Phialopecten spp. and Eucrassatella sp. Tucetona-dominated associations are widespread on the modern New Zealand shelf and occur most commonly on coarse sandy and gravelly substrates. Morton & Miller (Citation1968), McKnight (Citation1974) and Gillespie & Nelson (Citation1996) documented similar modern benthic communities. The Tucetona association likely accumulated in nearshore settings on very coarse sandy to gravelly substrates, probably in tidal channels, and close to a rocky greywacke coastline.
The Zeacolpus association (ZC) is common in coarse-grained pebble-rich limestone and some sandstone of Late Miocene (Tongaporutuan) age, especially around Te Haroto (BH38/ 096649) and Tarawera Station (BH38/170608) near the Napier–Taupo Road. The association is invariably present in beds above pronounced unconformities. It is almost monospecific at most localities, being characterised by the turritellid gastropod Zeacolpus spp. The Zeacolpus association is commonly interbedded with, and passes laterally into, the Struthiolaria (Callusaria) association.
The Struthiolaria (Callusaria) association (ST) is also restricted to rocks of Tongaporutuan–Opoitian age in western parts of the study area (e.g. Te Haroto Formation, BH38/096649; Te Ipuohape Sandstone Member, BJ38/105508; Omahaki Formation, BJ37/913251). The association is most common in proximity to Mesozoic basement rocks, and is dominated by the gastropod Struthiolaria (Callusaria) spp. with common Zenatia acinaces and Zeacolpus sp. Lithofacies containing the Struthiolaria (Callusaria) association are concretionary fine- to medium sandstone to less-common pebbly limestone and shelly conglomerate. Both the Zeacolpus and Struthiolaria (Callusaria) associations probably accumulated in nearshore to inner shelf environments on sandy to slightly gravelly substrates, in settings that were in close proximity to a rocky greywacke coastline.
The Crassostrea association (CS), common in Early–Middle Pliocene (Opoitian–Mangapanian) rocks in western parts of the study area, is dominated by the extinct large thick-shelled oyster Crassostrea ingens. The Crassostrea association is mostly restricted to settings in close proximity to basement rocks, although it does occur on hard-ground surfaces representing unconformities. The fauna of the Crassostrea association is dominated by epifaunal or robust semi-infaunal taxa such as Crassostrea ingens, Maoricardium spatiosum, Tucetona laticostata and occasional Phialopecten spp. In some localities, such as Opau Stream (BJ38/107433; Katz 1973), the association contains abundant flabellid corals. Lithofacies containing the Crassostrea association are usually very coarse conglomerate to breccia that directly overlie greywacke basement. Where the association is separated from greywacke basement by other Cenozoic beds, the lithofacies containing the association still comprise conglomerate at most sites. The Crassostrea association accumulated in high-energy shoreface to nearshore settings on hard-ground substrates adjacent to a rocky greywacke coastline.
The Crassostrea-Phialopecten sub-association (CS-PP) is restricted to western parts of the study area to limestone lithofacies of the Middle Pliocene Te Waka Formation in the Patoka and Puketitiri area, although it is common in Te Aute limestone lithofacies in the eastern hill country of Hawke's Bay (e.g. Beu Citation1995). The sub-association is characterised by the abundance of Crassostrea ingens and the scallop Phialopecten thomsoni. Patro undatus is also a very common component of the sub-association, as are abundant barnacle plates. The Crassostrea-Phialopecten sub-association accumulated at nearshore to inner shelf water depths on a gravelly mixed bioclastic-siliciclastic hard-ground substrate. The sub-association probably represents the development of carbonate factories: sites of high bioclastic carbonate productivity typically located atop a structural high such as an anticlinal crest or other form of submarine seafloor relief (e.g. Kamp et al. Citation1988; Nelson et al. Citation2003; Caron et al. Citation2004a).
The Neothyris association (NE) is restricted to Pliocene (Waipipian–Mangapanian) Te Waka and Titiokura Formation limestone beds along the western margin of the study area. Although often dominated by barnacle fragments, it is the abundance of intact specimens of the brachiopod Neothyris aff. obtusa that distinguishes the biofacies. Assemblages of this biofacies also commonly contain Phialopecten marwicki or P. thomsoni and Ostrea chilensis, and it may pass laterally into the Crassostrea-Phialopecten sub-association. The Neothyris association is inferred to have accumulated in shallow water environments between lower-inner to middle shelf water depths on firm-ground shell-gravel to sandy substrates. Strong tidal currents are assumed to have kept the palaeo-seafloor relatively free of fine siliciclastic sediment as occurs in present-day habitats for Neothyris, such as in Cook Strait and Foveaux Strait (e.g. Neall Citation1970, Citation1972; Beu & Maxwell Citation1990). Such a depositional setting is interpreted for western Hawke's Bay during the Pliocene, as encompassed by the ‘Ruataniwha’ and ‘Kuripapango’ straits (Pettinga Citation1980; Beu et al. Citation1980; Browne Citation2004a; Bland et al. Citation2008a; Trewick & Bland Citation2012).
Biofacies of offshore shelf palaeoenvironments
Offshore shelf palaeoenvironments are widespread in Neogene rocks in central Hawke's Bay. Faunal diversity varies from low to very high, with the highest diversity usually in rocks of middle to outer shelf palaeoenvironments.
The Atrina association (AT) contains a sparse macrofauna dominated almost exclusively by the common extant horse mussel Atrina pectinata. Atrina zelandica is widespread in a variety of semi-enclosed bays and inner to middle shelf environments on the modern New Zealand shelf, and it is also commonly exposed on tidal flats (A.G. Beu, pers. comm., 2009). Lithofacies containing the Atrina association are typically silty sandstone to sandy siltstone, and the association is inferred to have accumulated at inner shelf to upper to middle shelf water depths on and in silty sand substrates with a reasonably high sediment input.
The Mactra (Maorimactra) association (MM) is dominated by the small bivalve Mactra (Maorimactra) spp. and occurs in fine sandstone to silty sandstone lithofacies of Late Pliocene–Early Pleistocene (Mangapanian–Nukumaruan) age. Mactra (Maorimactra) ordinaria is an extant species that occupies a broad bathymetric range (10–170 m), although it is most commonly present in water depths of less than 80 m (Hendy & Kamp Citation2004). Modern communities resembling the Mactra (Maorimactra) association are common on the New Zealand shelf in water depths of 20–60 m (Fleming Citation1953; McKnight Citation1969). Beu & Maxwell (Citation1990) regarded Mactra (Maorimactra) as an indicator of shallow-water soft-bottom lithofacies. The Mactra (Maorimactra) association is inferred to have accumulated on soft fine sand to muddy sand substrates at inner to upper-middle shelf water depths.
The Dosinia (Kereia) association (DK) is widespread in silty sandstone to sandy siltstone lithofacies and is often dominated by in situ and articulated Dosinia (Kereia) greyi, an extant species that prefers sheltered inner shelf settings (Hendy & Kamp Citation2004). The Dosinia (Kereia) association is inferred to have accumulated on silty sand substrates at inner shelf (10–50 m) water depths. The environment of deposition for the M. (Maorimactra) association is thought to have been more open-marine than that for the Dosinia (Kereia) association, although both are likely to have inhabited similar water depths and substrates.
In contrast to the diversity often present within the Mactra (Maorimactra) and Dosinia (Kereia) associations the Ostrea association (OS) is usually monospecific, containing only Ostrea chilensis. The Ostrea association likely accumulated on silty firm-ground substrates at lower-inner to middle shelf water depths, probably no deeper than about 60 m (Hendy & Kamp Citation2004). It is inferred that periods of accumulation of this biofacies marked periods of sediment starvation and minor biostrome and reef development.
The Patro association (PT) has limited occurrence and differs from the Tawera-Patro sub-association and Talochlamys association by the more ubiquitous presence of the false oyster Patro undatus and lack of Tawera and Talochlamys. As Patro undatus is an extinct taxon, its habitat during the Miocene–Pleistocene can only be inferred from co-occurring taxa and enveloping sedimentary lithofacies. It is inferred to have accumulated at lower-inner to middle shelf water depths on fine sandy to silty firm-ground substrates, and represents a transitional grouping from the shallow-water Tawera-dominated associations to the deeper water Pratulum and Talochlamys associations.
The Talochlamys association (TA) is widespread throughout the study area and occurs in a wide variety of generally fine-grained lithofacies. It is characterised by common Talochlamys gemmulata in assemblages of generally high diversity. Talochlamys gemmulata is an extant scallop widespread on the modern New Zealand shelf from the low-tide zone to bathyal water depths on a wide variety of substrates (Beu & Maxwell Citation1990; Dijkstra & Marshall Citation2008). This modern distribution is reflected in the widespread occurrence of Talochlamys in the fossil record. The lack of dominance of any one taxon reflects the high diversity observed in the faunal content of such middle to outer shelf environments. While modern Talochlamys gemmulata can occupy a broad range of palaeoenvironments, most occurrences of the Talochlamys association are inferred to have accumulated on firm-ground silt to sandy silt substrates at middle to outer shelf (50–100 m) water depths. It is possible that these firm-grounds record periods of sediment starvation, or the deposition of thin beds of fine sand.
Two sub-associations have been identified within the Talochlamys association. The Stiracolpus-Talochlamys sub-association (SC-TA) is characterised by the co-dominance of the turritellid gastropod Stiracolpus spp. and Talochlamys gemmulata and occurs relatively commonly, mostly in Z3 siltstone lithofacies (e.g. Beu Citation1965). Faunal diversity in the sub-association is relatively high, and Nucula and Neilo are common constituents. The association is inferred to have accumulated at middle to outer shelf water depths (50–150 m) on silty substrates. By contrast, the Notosaria-Talochlamys sub-association (NO-TA) has been observed at only a limited number of sites and is restricted to late Nukumaruan members of the Petane Formation. This sub-association is characterised by the dominance of the extant brachiopod Notosaria nigricans, and is typically present in Z2 siltstone lithofacies. It is inferred to have accumulated on a firm-ground substrate at lower-inner to middle shelf water depths. The environment of accumulation is inferred to be similar to that of the Neothyris association, although this association developed under a regime with a lesser tidal and storm-current influence and on a finer-grained seafloor. In its modern distribution, Notosaria nigricans often forms part of a hard-bottom community dominated by filter feeders (Lee Citation1978). Accumulation of this sub-association may mark periods of firm-ground development due to sediment starvation or perhaps the deposition of thin layers of fine sand during storm events.
The Neilo association (NI) is an uncommon low diversity assemblage characterised by Neilo spp. Other molluscs common in this association include Notocallista multistriata, Calliostoma sp., Talochlamys gemmulata, Pleuromeris sp. and nuculanids. Lithofacies containing the Neilo association are massive fine sandy siltstone to siltstone, often with a fine pebble content, such as Z2 and Z5 siltstone lithofacies. A middle to outer shelf environment of accumulation is inferred for the Neilo association.
The Pratulum association (PR), conspicuously dominated by the extant cardiid bivalve Pratulum pulchellum, is widespread in the study area in shelf lithofacies of late Mangapanian–late Nukumaruan age. It is most common in lower parts of Z2 and Z8 siltstone lithofacies beds of the Pohue, Matahorua, Petane and Okauawa formations. The Pratulum association may either be monospecific or contain a moderately diverse fauna. The molluscs Talochlamys gemmulata, Ostrea chilensis, Dosinia (Kereia) greyi, Zenatia acinaces, Antalis nana, Pelicaria spp. and Austrofusus spp. occur in most Pratulum association occurrences. The association resembles modern assemblages described by Dell (Citation1951a,Citationb), Estcourt (Citation1967), Luckens (Citation1972), Grace & Hayward (Citation1980) and Hayward et al. (Citation1984), as well as the Nemocardium pulchellum-Pleuromeris zelandica community of McKnight (Citation1974). These commonly occur in muddy sand to mud and occasionally on sand along North Island's west coast at water depths of 20–130 m. The Pratulum association described here likely accumulated in lower-inner to outer shelf water depths on a seafloor characterised by soft, silty sediments. The most common distribution of this association is inferred to be in beds that were deposited at middle to outer shelf water depths during times of sea-level highstand. The Pratulum association passes laterally into the Talochlamys and Notosaria-Talochlamys associations, both of which probably accumulated on a firmer substrate, although at similar water depths to the Pratulum association.
Palaeogeographical distribution of biofacies
Using the mapped spatial distribution of biofacies and lithofacies within central and western Hawke's Bay (Bland Citation2006), we have developed a series of detailed palaeogeographies for the Pliocene–Pleistocene (). These maps illustrate the changing distribution and character of biofacies through time, primarily in response to changing basin and landmass configurations driven by regional tectonics. The maps () have been simplified in that they are drawn for glacio-eustatic lowstand–earliest transgressive conditions (see Bland Citation2006). Because there has been no collapsing of data across multiple sequences, the maps represent a palaeogeographic snap-shot within part of an individual sequence. For example, the 1.8 Ma map is based on the distribution of biofacies and lithofacies within sea-level lowstand during deposition of the Tangoio Limestone Member and Okauawa Formation (see Bland Citation2006; Bland et al. Citation2007).
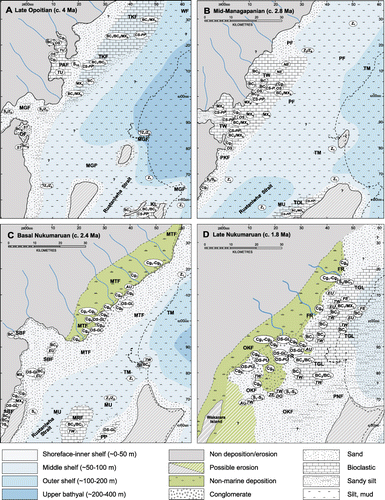
The main factors affecting the distribution of biofacies and lithofacies during deposition of the late Neogene Mangaheia Group included: (1) a general change from an irregular rocky coastline to an indented coastline with embayments and (during the late Nukumaruan) a hinterland comprising a low-gradient coastal plain; (2) an increase in width of the palaeoshelf; (3) the progressive tectonically driven constriction of the basin during the Pliocene because of uplift of adjacent landmasses, resulting in increased strength of tidal flows; (4) the progradation of basement-derived fluvial braid-plain systems during two periods during the Late Pliocene; and (5) geographic variations in sedimentation associated with glacio-eustatically driven oscillations in sea level with durations of c. 41 ka (e.g. Haywick et al. Citation1992; Beu Citation1995; Bland et al. Citation2004, Citation2008a,Citationb; Caron et al. Citation2004a,Citationb). It appears that an increase in the variety of lithofacies and macrofaunal associations occurred through the Pliocene, with greater heterogeneity of lithofacies and biofacies distribution during periods of relative sea-level lowstand.
Early Pliocene (c. 4 Ma)
The distribution of biofacies during the Early Pliocene (late Opoitian; ), as recorded by the Zeacolpus, Struthiolaria (Callusaria), Tucetona, Crassostrea and Crassostrea-Phialopecten associations, suggests that the western coastline of central Hawke's Bay was rocky and highly irregular with embayments, perhaps resembling modern-day Coromandel Peninsula or eastern Northland in character. A shallow (c. 50–100 m deep) seaway across the trend of the modern-day North Island axial ranges around the Kuripapango area connected western Hawke's Bay with eastern Wanganui Basin at this time (Browne Citation2004b; Bland et al. Citation2008a; Trewick & Bland Citation2012). Bioclastic lithofacies along the western margin are rich in greywacke clasts (Bland et al. Citation2007), reflecting the nearby basement exposures and rocky coastline. Coeval lithofacies along eastern margins of the basin record mobilisation of shell debris off structural highs (e.g. Nelson et al. Citation2003; Caron et al. Citation2004a). Towards the axis of the basin, bioclastic beds pass rapidly into sandstone and then into massive mudstone lithofacies (see Bland Citation2006; Bland et al. Citation2008a).
Late Pliocene (c. 2.8 Ma)
By the Late Pliocene (mid Mangapanian; ) progressive uplift along both margins of the basin had induced an increased constriction of tidal flows in the forearc basin (Pettinga Citation1980; Beu et al. Citation1980; Beu Citation1995). Shore-parallel shell banks accumulated along these margins, although massive mudstone lithofacies were still accumulating in the axis of the basin during this time (e.g. Ozolins & Francis Citation2000; Caron et al. Citation2004a; Bland et al. Citation2008a). The western coastline was probably still very irregular and rocky (e.g. Browne Citation2004a,Citationb; Bland et al. Citation2008a), with the Crassostrea and Crassostrea-Phialopecten associations accumulating adjacent to the coastline in association with mixed siliciclastic-bioclastic lithofacies. These associations were also accumulating atop the current-swept fault-bounded hanging-wall highs that were becoming increasingly emergent along the eastern margin of the basin (e.g. Beu et al. Citation1980; Kamp et al. Citation1988; Caron et al. Citation2004a; Bland et al. Citation2005, Citation2007). Most bioclastic-rich lithofacies in western areas contain the Tucetona, Neothyris and Crassostrea-Phialopecten associations. Emergent islands of basement occurred in several places, such as the Opau Stream area (Katz Citation1973; Bland et al. Citation2008a; BJ38/107433, column 73a, 73b), around which pebbly bioclastic beds and fossiliferous breccia were deposited, usually in association with a macrofauna belonging to the Crassostrea association. Along the western margin of the basin, bioclastic-dominated rocks are generally restricted to the area southwest of the Napier–Taupo Road. Fine sandstone and massive mudstone occur to the northeast of this area and, although sparsely fossiliferous, typically contain elements of the Neothyris and Talochlamys associations. The coeval mudstone-dominated rocks that were deposited in central parts of the basin (e.g. Ozolins & Francis Citation2000; Bland et al. Citation2008a) are not exposed at the surface, and their faunal content cannot be assessed. By the late Mangapanian, cyclothemic rocks of the Pohue Formation were accumulating across much of the basin and contain a diversity of biofacies as a result of variations in sediment type and water depth, induced by glacio-eustatic sea-level changes (e.g. Haywick et al. Citation1991, Citation1992; Bland et al. Citation2008b). Most shelf mudstones of Pohue Formation contain the Talochlamys, Atrina, Pratulum, Dosinia (Kereia) and Mactra (Maorimactra) associations, whereas most overlying nearshore sandstones contain the Zethalia and Dosinia associations.
Early Pleistocene (c. 2.4 Ma)
By c. 2.4 Ma (basal Nukumaruan; ) a large embayment surrounded by a rocky basement coastline is inferred to have been present in the Kuripapango and Awapai Station areas. Lithofacies in this embayment comprised coarse-grained fossiliferous conglomerate, shelly sandstone and limestone units primarily containing the Ostrea-glycymerid and Eumarcia associations. Coarse-grained pebbly limestone lithofacies were also deposited in the Ohara Depression (Erdman & Kelsey Citation1992; Beu Citation1995; Bland et al. Citation2007) and are characterised by the Ostrea-glycymerid association. The presence of emergent Torlesse composite terrane greywacke to the east (Wakarara Range) and west (Ruahine Range) of the depression, a consequence of syn-sedimentary vertical offset on the Ruahine, Mohaka and Wakarara faults (Erdman & Kelsey Citation1992; Bland et al. Citation2008a; Trewick & Bland Citation2012), probably constricted tidal flows through the area, winnowing fine siliciclastic sediments and promoting the growth of an epifauna-dominated assemblage.
The localised uplift of structural highs around Scinde Island, Mason Ridge and Pakipaki, which were cored by reverse faults, allowed deposition of limestone lithofacies in these areas (). Here, most biofacies are part of the Tawera and Ostrea-glycymerid associations. Giant cross-bedded limestone at Scinde Island indicates strong tidal currents over this site (Kamp et al. Citation1988). The bioclastic deposits at Scinde Island pass rapidly into massive mudstone lithologies in all directions (e.g. Westech Citation2001). Macrofossils in this mudstone-dominated interval are relatively sparse, but where present are part of the Neilo, Pratulum or Talochlamys associations.
Middle Pleistocene (c. 1.8 Ma)
By the late Nukumaruan (c. 2–1.8 Ma) many of the major faults in the North Island fault system (such as the Kaweka, Ruahine, Mohaka and Wakarara faults) had begun to accommodate increased rates of contractional to oblique slip offset. This had the effect of rapidly forming areas of elevated topography along western parts of the basin (e.g. Erdman & Kelsey Citation1992; Bland & Kamp Citation2006; Bland et al. Citation2008a; Trewick & Bland Citation2012). The distribution of macrofaunal associations during the late Nukumaruan indicates that lowstand shorelines were strongly embayed (e.g. Bland et al. Citation2008a; ), consistent with the nature of the lowstand shoreline inferred for the Tangoio Block by Haywick et al. (Citation1992). A topographically high hinterland, formed from Torlesse composite terrane basement rocks, lay west of the basin (Trewick & Bland Citation2012), and contributed large volumes of coarse-grained sediments via braided river systems. Large semi-enclosed embayments with tidal channels occurred in the Sherenden and Matapiro areas, with lithofacies dominated by shallow-water fossiliferous conglomerates rich in taxa such as Tucetona, Ostrea, Dosinia, Zethalia, Maoricolpus, Purpurocardia and Austrovenus (). These molluscs are common in modern shallow-water high-energy environments such as tidal channels in enclosed embayments around the New Zealand coastline (Morton & Miller Citation1968). Conglomerate lithofacies are inferred to have been less widespread in the Tangoio Block area and pass rapidly from pebbly limestone units into shelly limestone beds (Haywick Citation2000; Bland et al. Citation2007). Most molluscs in limestone beds in this area are part of the Tawera and Eumarcia associations. Overlying mudstone intervals, deposited during sea-level highstands, are highly fossiliferous with the Pratulum, Talochlamys, Dosinia (Kereia) and Atrina associations the most common. Most sandy siliciclastic-dominated lithofacies in late Nukumaruan rocks contain the Fellaster and Zethalia associations, reflecting shoreface and nearshore depositional settings.
Conclusions
The character and distribution of 30 molluscan biofacies in late Neogene strata in Hawke's Bay have been defined from analysis of extensive faunal composition datasets. The spatial and temporal distribution of these biofacies was strongly controlled by interplay between regional tectonics and glacio-eustatic sea-level oscillations, particularly during the Middle Pliocene–Early Pleistocene. Early phases of relative sea-level rise were characterised by biofacies that lived on coarse-grained sediments, typically involving robust semi-infaunal taxa such as the Struthiolaria (Callusaria), Tucetona, Ostrea-glycymerid and Crassostrea associations. Biofacies of offshore palaeoenvironments mostly accumulated during times of high relative sea level, are characterised by fine-grained muddy sedimentation across the shelf and are dominated by suspension-feeding infaunal taxa such as the Pratulum and Neilo associations. Biofacies associated with times of falling sea level, such as the Zethalia and Fellaster associations, contain assemblages of rapid-burrowing infaunal bivalves, gastropods and echinoderms characteristic of high-energy conditions.
Because many of the macrofossils and associations identified have extant analogues off the modern New Zealand coastline it has been possible to reconstruct a series of detailed palaeoenvironmental maps for the central Hawke's Bay region at time periods of c. 4, 2.8, 2.4 and 1.8 Ma. These palaeogeographic reconstructions emphasise the rapid lateral and temporal changes in lithofacies, and therefore habitats, in response to the eustatic and tectonic drivers. Future correlations of the late Neogene macrofossil assemblages between age-equivalent rocks in Wanganui and Taranaki basins and the Hawke's Bay area will result in a better understanding of the palaeogeographic evolution of central and lower North Island during this time, as well as enhancing insight into the dynamics of community replacement under different tectonic and sedimentary regimes.
Acknowledgements
KJB acknowledges financial assistance from a Tertiary Education Commission TAD Scholarship and GNS Science QMAP field support funding. We acknowledge Public Good Science Funding provided by the New Zealand Government to the University of Waikato and GNS Science. Alan Beu is thanked for palaeontological assistance. We gratefully acknowledge John Simes (GNS Science) for uploading much of the fossil dataset into FRED. Vincent Caron, Arne Pallentin, Dave Francis and Greg Browne are thanked for helpful discussions about East Coast stratigraphy. Rhys Graafhuis, Rachel Baggs and Sarah Dyer (University of Waikato) are acknowledged for their inputs into geological mapping and lithostratigraphy of central Hawke's Bay. Betty-Ann Kamp assisted with preparation of some figures. Reviews from Angela Griffin, Alan Beu, James Crampton and Kari Bassett significantly improved earlier versions of this paper.
References
- Abbott ST, Carter RM 1997. Macrofossil associations from mid-Pleistocene cyclothems, Castlecliff section, New Zealand: implications for sequence stratigraphy. Palaios 12: 188–210. 10.2307/3515306
- Ballance PF 1993. The New Zealand Neogene forearc basins. In: Ballance PF Sedimentary basins of the world 2. South Pacific sedimentary basins. Amsterdam, Elsevier. Pp. 177–191.
- Beu AG 1965. Lower Pleistocene mollusca from Devils Elbow, Hawke's Bay. Transactions of the Royal Society of New Zealand, Geology 3: 139–149.
- Beu AG 1995. Pliocene limestones and their scallops. Lithostratigraphy, pectinid biostratigraphy and paleogeography of eastern North Island Late Neogene limestone. Institute of Geological and Nuclear Sciences Monograph 10. Lower Hutt, Institute of Geological and Nuclear Sciences. 243 p.
- Beu AG 2004. Marine mollusca of oxygen isotope stages of the last 2 million years in New Zealand. Part 1. Revised generic positions and recognition of warm-water and cool-water migrants. Journal of the Royal Society of New Zealand 34: 111–265. 10.1080/03014223.2004.9517766
- Beu AG 2006. Marine mollusca of oxygen isotope stages of the last 2 million years in New Zealand. Part 2. Biostratigraphically useful and new Pliocene to Recent bivalves. Journal of the Royal Society of New Zealand 36: 151–338. 10.1080/03014223.2006.9517808
- Beu AG, Maxwell PC 1990. Cenozoic mollusca of New Zealand. New Zealand Geological Survey Paleontological Bulletin 58. 518 p.
- Beu A, Kitamura, A 1998. Exposed coasts vs sheltered bays: contrast between New Zealand and Japan in the molluscan record of temperature change in Plio-Pleistocene cyclothems. Sedimentary Geology 122: 129–149. 10.1016/S0037-0738(98)00102-X
- Beu AG, Grant-Taylor TL, Hornibrook N de B 1980. The Te Aute limestone facies, Poverty Bay to northern Wairarapa. New Zealand Geological Survey miscellaneous series map 13. 36 p. +2 maps.
- Bland KJ 2001. Analysis of the Pliocene forearc basin succession, Esk River catchment, Hawke's Bay. Unpublished MSc thesis, lodged in the Library, The University of Waikato, Hamilton, New Zealand. 233 p.
- Bland KJ 2006. Analysis of the central Hawke's Bay sector of the late Neogene forearc basin, Hikurangi margin, New Zealand. Unpublished PhD thesis, lodged in the Library, The University of Waikato, Hamilton, New Zealand. 306 p. +8 enclosures and appendices (http://researchcommons.waikato.ac.nz/handle/10289/2222).
- Bland KJ, Kamp PJJ 2006. Geological structure of the forearc basin in central Hawke's Bay, eastern North Island. In: 2006 New Zealand Petroleum Conference proceedings. Wellington, Ministry of Economic Development. 13 p.
- Bland KJ, Nelson CS, Kamp PJJ 2003. Latest Miocene-early Pliocene onlap limestone beds in the western forearc basin, Hawke's Bay. In: Mortimer N, Lee D eds. Geological Society of New Zealand 2003 Annual Conference, Dunedin. Geological Society of New Zealand Miscellaneous Publication116A. Pp. 21.
- Bland KJ, Kamp PJJ, Pallentin A, Graafhuis RB, Nelson CS, Caron V 2004. The early Pliocene Titiokura Formation: stratigraphy of a thick mixed carbonate-siliciclastic shelf succession in Hawke's Bay Basin, New Zealand. New Zealand Journal of Geology and Geophysics 47: 675–695. 10.1080/00288306.2004.9515083
- Bland KJ, Caron V, Nelson CS, Kamp PJJ, Pallentin A 2005. Continent-attached and continent-detached cool-water limestones in an actively deforming forearc basin, Hawke's Bay, New Zealand. In: The Geological Society of America Annual Meeting & Exposition, 16–19 October 2005. SLC 2005, Science, learning, colleagues. Salt Lake City, Utah. Abstracts with programs/Geological Society of America 37(7). Pp. 17.
- Bland KJ, Kamp PJJ, Nelson CS 2007. Systematic lithostratigraphy of the Neogene succession exposed in central parts of the Hawke's Bay Basin, eastern North Island, New Zealand. Ministry of Economic Development New Zealand unpublished petroleum report PR3724. 259 p. +2 enclosures (http://hdl.handle.net/10289/3495).
- Bland KJ, Kamp PJJ, Nelson CS 2008a. Late Miocene-Early Pleistocene paleogeography of the onshore central Hawke's Bay sector of the forearc basin, eastern North Island, New Zealand, and some implications for hydrocarbon prospectivity. In: 2008 New Zealand Petroleum Conference proceedings: Beyond 08. Wellington, Ministry of Economic Development. 22 p.
- Bland KJ, Nelson CS, Kamp PJJ 2008b. Sequence stratigraphic character of late Neogene strata in the onshore East Coast forearc basin, central Hawke's Bay, New Zealand. In: 2008 New Zealand Petroleum Conference proceedings: Beyond 08. Wellington, Ministry of Economic Development.
- Brett CE 1998. Sequence stratigraphy, paleoecology, and evolution: biotic clues and responses to sea level fluctuations. Palaios 13: 241–262. 10.2307/3515448
- Brett CE, Hendy AJW, Bartholomew AJ, Bonelli JR Jnr, McLaughlin PI 2007. Response of shallow marine biotas to sea-level fluctuations: A review of faunal replacement and the process of habitat tracking. Palaios 22: 228–244. 10.2110/palo.2005.p05-028r
- Browne GH 2004a. Late Neogene sedimentation adjacent to the tectonically evolving North Island axial ranges: Insights from Kuripapango, western Hawke's Bay. New Zealand Journal of Geology and Geophysics 47: 663–674. 10.1080/00288306.2004.9515082
- Browne GH 2004b. Field trip 6: The Miocene-Pliocene interior seaway of the central North Island: sedimentary patterns and tectonic styles in the Kuripapango Strait. In: Manville, V.R. ed. Geological Society of New Zealand/New Zealand Geophysical Society/26th New Zealand Geothermal Workshop, 6th–9th December 2004, Taupo: field trip guides. Geological Society of New Zealand miscellaneous publication 117B. Pp. 89–109.
- Caron V, Nelson CS, Kamp PJJ 2004a. Contrasting carbonate depositional systems for Pliocene cool-water limestones cropping out in central Hawke's Bay, New Zealand. New Zealand Journal of Geology and Geophysics 47: 697–717. 10.1080/00288306.2004.9515084
- Caron V, Nelson CS, Kamp PJJ 2004b. Transgressive surfaces of erosion as sequence boundary markers in cool water shelf carbonates. Sedimentary Geology 164: 179–189. 10.1016/j.sedgeo.2003.10.001
- Dell RK 1951a. Some animal communities of the sea bottom from Queen Charlotte Sound, New Zealand. New Zealand Journal of Science and Technology B33: 19–29.
- Dell RK 1951b. A molluscan fauna from Mernoo Bank, New Zealand. New Zealand Journal of Science and Technology B33: 15–18.
- Dijkstra HH, Marshall BA 2008. The Recent Pectinoidea of the New Zealand region (Mollusca: Bivalvia: Propeamussiidae, Pectinidae and Spondylidae). Molluscan Research 28:1–88.
- Erdman CF, Kelsey HM 1992. Pliocene and Pleistocene stratigraphy and tectonics, Ohara Depression and Wakarara Range, North Island, New Zealand. New Zealand Journal of Geology and Geophysics 35: 177–192. 10.1080/00288306.1992.9514512
- Estcourt IN 1967. Distributions and associations of benthic invertebrates in a sheltered soft-bottom environment (Marlborough Sounds, New Zealand). New Zealand Journal of Marine and Freshwater Research 1: 352–370. 10.1080/00288330.1967.9515211
- Field BD, Uruski CI, Beu AG, Browne GH, Crampton J, Funnell R, Killops S, Laird M, Mazengarb C, Morgans HEG, Rait G, Smale D, Strong CP 1997. Cretaceous–Cenozoic geology and petroleum systems of the East Coast region, New Zealand. Institute of Geological and Nuclear Sciences monograph 19. Institute of Geological and Nuclear Sciences Limited, Lower Hutt. 301 p. +7 enclosures.
- Fleming CA 1953. The Geology of the Wanganui Subdivision. New Zealand Geological Survey Bulletin n.s. 52: 361 p.
- Gilinsky NL, Bennington JB 1994. Estimating numbers of whole individuals from collections of body parts; a taphonomic limitation of the paleontological record. Paleobiology 20: 245–258.
- Gillespie JL, Nelson CS 1996. Distribution and control of mixed terrigenous-carbonate surficial sediment facies, Wanganui shelf, New Zealand. New Zealand Journal of Geology and Geophysics 39: 533–549. 10.1080/00288306.1996.9514731
- Grace RV 1966. The bottom communities of the entrance to the Whangateau Harbour. Tane 12: 63–70.
- Grace RV, Hayward BW 1980. The macrobenthos of the Cavalli Islands, northern New Zealand. Tane 20: 189–209.
- Grange KR 1979. Soft-bottom macrobenthic communities of Manukau harbour, New Zealand. New Zealand Journal of Marine and Freshwater Research 13: 315–329. 10.1080/00288330.1979.9515809
- Grindley GW 1960. Geological Map of New Zealand 1:250 000. Sheet 8 Taupo. New Zealand Geological Survey. Wellington: Department of Scientific and Industrial Research.
- Hayward BW, Grace RV, Bull VH 1984. Soft-bottom macrofauna, foraminifera and sediments of the Chickens Island, northern New Zealand. Tane 30: 141–164.
- Haywick DW 1990. Stratigraphy, sedimentology, paleoecology and diagenesis of the Petane Group (Pliocene-Pleistocene) in the Tangoio Block, central Hawke's Bay, New Zealand. Unpublished PhD thesis, James Cook University of North Queensland, Townsville, Australia.
- Haywick, DW 2000. Recognition and distinction of normal and forced regression in cyclothemic strata: a Plio-Pleistocene case study from eastern North Island, New Zealand. In: Hunt D and Gawthorpe RL eds. Sedimentary responses to forced regressions. Geological Society of London Special Publication 172: 193–215.
- Haywick DW, Lowe DA, Beu AG, Henderson RA, Carter RM 1991. Pliocene-Pleistocene (Nukumaruan) lithostratigraphy of the Tangoio Block, and origin of sedimentary cyclicity, central Hawke's Bay, New Zealand. New Zealand Journal of Geology and Geophysics 34: 213–225. 10.1080/00288306.1991.9514459
- Haywick DW, Carter RM, Henderson RA 1992. Sedimentology of 40 000 year Milankovitch-controlled cyclothems from central Hawke's Bay, New Zealand. Sedimentology 39: 675–696. 10.1111/j.1365-3091.1992.tb02144.x
- Hendy AJW 2009. Lithification and the measurement of biodiversity. Paleobiology 35: 51–62. 10.1666/07047.1
- Hendy AJW, Kamp PJJ 2004. Late Miocene to early Pliocene biofacies of Wanganui and Taranaki Basins, New Zealand: applications to paleoenvironmental and sequence stratigraphic analysis. New Zealand Journal of Geology and Geophysics 47: 769–785. 10.1080/00288306.2004.9515088
- Hendy AJW, Kamp PJJ 2007. Paleoecology of Late Miocene-Early Pliocene sixth-order glacioeustatic sequences in the Manutahi-1 core, Wanganui-Taranaki Basin, New Zealand. Palaios 22: 325–337. 10.2110/palo.2006.p06-084
- Kamp PJJ, Harmsen FJ, Nelson CS, Boyle SF 1988. Barnacle-dominated limestone with giant cross-beds in a non-tropical, tide-swept, Pliocene forearc seaway, Hawke's Bay, New Zealand. Sedimentary Geology 60: 173–195. 10.1016/0037-0738(88)90118-2
- Kamp PJJ, Bland KJ, Caron V, Graafhuis RB, Baggs RA, Dyer SDJ, Boyle SF, Nelson CS 2007. Stratigraphic columns for the Neogene succession exposed in central parts of Hawke's Bay Basin, eastern North Island, New Zealand, New Zealand. Ministry of Economic Development New Zealand unpublished petroleum report PR3725. 483 p. ( http://hdl.handle.net/10289/3502).
- Katz HR 1973. Pliocene unconformity at Opau Stream, Hawke's Bay, New Zealand. New Zealand Journal of Geology and Geophysics 16: 917–925. 10.1080/00288306.1973.10555231
- Kondo Y, Abbott ST, Kitamura A, Kamp PJJ, Naish TR, Kamataki T, Saul GG 1998. The relationship between shellbed type and sequence architecture: examples from Japan and New Zealand. Sedimentary Geology 122: 109–127. 10.1016/S0037-0738(98)00101-8
- Lee DE 1978. Aspects of the ecology and paleoecology of the brachiopod Notosaria nigricans (Sowerby). Journal of the Royal Society of New Zealand 8: 395–417. 10.1080/03036758.1978.10423321
- Lee JM, Bland KJ, Townsend DB, Kamp PJJ (compilers) 2011. Geology of the Hawke's Bay area. Institute of Geological & Nuclear Sciences 1:250 000 geological map 8. Lower Hutt: Institute of Geological & Nuclear Sciences Limited. 93 p. +1 folded map.
- Lewis KB, Pettinga JR 1993. The emerging, imbricate frontal wedge of the Hikurangi Margin. In: Ballance PF ed. Sedimentary basins of the world 2: South Pacific sedimentary basins. Amsterdam, Elsevier. Pp. 225–250.
- Luckens PA 1972. Distribution of Nemocardium (Pratulum) pulchellum (Gray 1843) (Mollusca: Bivalvia: Cardiidae) in the New Zealand region. NZOI Records 1. Pp. 47–63.
- McKnight DG 1969. Infaunal benthic communities on the New Zealand continental shelf. New Zealand Journal of Marine and Freshwater Research 3: 409–433. 10.1080/00288330.1969.9515308
- McKnight DG 1974. Benthic communities from the continental shelf, west coast North Island, Kawhia Harbour to Cape Terawhiti. NZOI Oceanographic Summary 4: 1–7.
- Mazengarb C, Speden IG (compilers)2000. Geology of the Raukumara area. Institute of Geological & Nuclear Sciences 1:250 000 geological map 6: 60 p. +1 map.
- Mazengarb C, Francis DA, Moore PR 1991. Sheet Y16-Tauwhareparae. Geological Map of New Zealand 1:50 000: 52 p. +1 map.
- Morton JE, Miller M 1968. The New Zealand sea shore. London, Collins. 638 p.
- Neall V 1970. Notes on the ecology and paleoecology of Neothyris, an endemic New Zealand brachiopod. New Zealand Journal of Marine and Freshwater Research 4: 117–125. 10.1080/00288330.1970.9515332
- Neall V 1972. Systematics of the endemic New Zealand brachiopod Neothyris. Journal of the Royal Society of New Zealand 2: 229–247. 10.1080/03036758.1972.10429376
- Nelson CS, Winefield PR, Hood SD, Caron V, Pallentin A, Kamp PJJ 2003. Pliocene Te Aute limestones, New Zealand: expanding concepts for cool-water shelf carbonates. New Zealand Journal of Geology and Geophysics 46: 407–424. 10.1080/00288306.2003.9515017
- Ozolins V, Francis DA 2000. Whakatu-1 Well Completion Report. PEP 38328. Indo-Pacific Energy (NZ) Ltd. Ministry of Economic Development Unpublished Petroleum Report PR 2476.
- Pettinga JR 1980. Geology and landslides of the eastern Te Aute district, southern Hawke's Bay. Unpublished PhD thesis, The University of Auckland, Auckland, New Zealand. 3 volumes.
- Pettinga JR 1982. Upper Cenozoic structural history, coastal Southern Hawke's Bay, New Zealand. New Zealand Journal of Geology and Geophysics 25: 149–191. 10.1080/00288306.1982.10421407
- Powell AWB 1937. Animal communities of the sea-bottom in Auckland and Manukau Harbours. Transactions of the Royal Society of New Zealand 66: 354–401.
- Sessa JA, Patzkowsky ME, Bralower TJ 2009. The impact of lithification on the diversity, size distribution, and recovery dynamics of marine invertebrate assemblages. Geology 37:115–118. 10.1130/G25286A.1
- Trewick SA, Bland KJ 2012. Fire and slice: palaeogeography for biogeography at New Zealand's North Island/South Island juncture. Journal of the Royal Society of New Zealand 42: 153–183.
- Westech Energy New Zealand Ltd 2001. Hukarere-1 Well Completion Report. PEP 38325. Westech Energy New Zealand Ltd. Ministry of Economic Development Unpublished Petroleum Report PR 2656.