
Abstract
The scope and species content of the pterioid bivalve Monotis Bronn with almost global distribution in Norian (Late Triassic) marine strata are reconsidered. Various infra-generic groupings have been proposed in the last 50 years and all have been subject to some criticism. A scheme which allocates all species to one of eight subgenera (Monotis, Entomonotis Marwick, Eomonotis Grant-Mackie, Inflatomonotis Grant-Mackie, Maorimonotis Grant-Mackie, Pacimonotis Grant-Mackie & Silberling, Baikalomonotis Bychkov and Praemonotis Bychkov) is favoured here and, in line with current trends, these eight are afforded generic status. There is also a growing move away from recognition of subspecies and, when applied here, allows most to be accepted at species level with a few names synonymised.
Introduction
Monotis Bronn, 1830 is a genus-level name in use for more than 180 years for a group of pterioid bivalves widespread in the Late Triassic of the Tethys and the circum-Pacific but so far unknown from rocks of this age in Africa, Australia or Antarctica, where Late Triassic marine strata are rare (Westermann Citation1973). Two major problems have affected the terminology applied to the 50-odd infra-generic groupings within Monotis: identification and classification.
This group has received much taxonomic attention globally in the last 30 years or more, so an update on the identifications should be of some value to students of Triassic marine palaeontology and biodiversity. It also provides the opportunity to review some of the nomenclatural changes and propose new nomenclature where considered necessary or helpful.
The first alteration to the nomenclature of Monotis in the last 50 years was by Westermann (Citation1962), who put forward a ‘species group’ scheme. Some of these groups were formalised as subgenera by Grant-Mackie (1Citation978a) who resurrected Entomonotis (proposed by Marwick, Citation1935 as a subgenus, which quickly dropped out of use however). Grant-Mackie (1Citation978a) proposed three additional subgenera, somewhat paralleling Westermann's scheme. Two of these, Eomonotis and Maorimonotis, were treated as genera by Tozer (Citation1980) but have not yet found general acceptance at this level. Grant-Mackie (1Citation978a) recognised that more work was required on some northern Pacific species of Monotis, which he tentatively included in Entomonotis. This problem was partially addressed by Grant-Mackie & Silberling (Citation1990), who erected the subgenus Pacimonotis for Westermann's (Citation1962) Subcircularis Group. Two additional subgenera have since been proposed by Bychkov (Citation2007), one for some species included by Grant-Mackie (1981) in Eomonotis and the other for members of Westermann's Zabaikalica Group.
Despite some criticisms of the taxonomic scheme developed by Grant-Mackie (Citation1978a,b,c,d), it has been applied by various workers to China (JAG-M, unpublished), North and South America and Southwest Pacific faunas for more than 30 years without any serious nomenclatural or classificatory problems being encountered, and the chronostratigraphic scheme based upon it (Grant-Mackie Citation1985) has proven robust.
Payevskaya (Citation1985) concluded that species of Monotis could not be readily arranged into subgenera and, from the basis of numerical taxonomy, recognised six major ‘groups’ within the genus. These groups show considerable relationship to Westermann's (Citation1962) species groups. She claimed that the composition of these groups differs significantly from subgenera formulated by Grant-Mackie (1Citation978a), the main objection being that Grant-Mackie had included in the single subgenus (Entomonotis) the ochotica, subcircularis and zabaikalica groups. However, Grant-Mackie (1Citation978a) himself had already stated that Entomonotis was being used in a wide sense and that future work would be likely to separate some species into other (at that stage unnamed) subgenera. Subsequently, the subcircularis group was included in the new subgenus Pacimonotis (Grant-Mackie & Silberling Citation1990) and the zabaikalica group in the subgenus Baikalomonotis (Bychkov Citation2007), leaving the ochotica group in Entomonotis.
The content of Payevskaya's groups does closely parallel both Westermann's (Citation1962) groups and the subgenera; her conclusion (Citation1985, p. 32) that it is not advisable to give subgeneric status to groups within Monotis therefore has little basis. Her study has, in fact, arrived at groupings essentially similar to those subgenera, while in doing so using a valuable alternative approach to Monotis nomenclature.
Ando (Citation1987) also reviewed the nomenclature of Monotis in his study of the genus in Japan. He tentatively rejected the use of subgenera but went further. Using population analysis and character variability Ando synonymised many taxa, reducing the named Monotis of Japan from 23 or more taxa to 5 (4 species and a subspecies). He concluded (Citation1987, p. 63) that ‘… all the specimens of Monotis in each fossil bed seem to represent a single fossil population … Therefore it is reasonable to regard the wide morphological variations as intrapopulational.’ The claim that collections of Monotis from single levels are monospecific certainly does not hold for all Southwest Pacific faunas. For example, Monotis richmondiana Zittel, Citation1864 occurs in different slump packets at Kiritehere with M. maniapotoi Grant-Mackie, 1978 or M. routhieri Avias, 1953; M. acutecostata Techmann, 1918 is found with M. awakinoensis Grant-Mackie, 1978 or M. maniopotoi Grant-Mackie; and M. calvata Marwick, Citation1953 has been collected with specimens of Entomonotis at the top of the Monotis beds at Kiritehere and with Pacimonotis in New Caledonia (see Grant-Mackie Citation1981b; Campbell et al. Citation1985). In northwest North America, M. (Pacimonotis) subcircularis Gabb, Citation1864 and M. (Monotis) alaskana Smith, Citation1927 each occurs with M. (Eomonotis) jakutica (Teller, Citation1886) (Westermann Citation1962; Silberling et al. Citation1997). Other species associations have been identified in northeast China (unpublished fieldwork by JAG-M et al.). It can be noted that co-occurring Monotis species are placed in different subgenera and therefore become members of different genera in the proposed scheme.
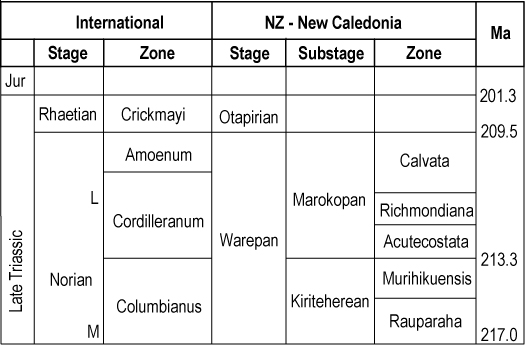
Ando's rejection of subgenera was based upon finding that morphological characters were ‘extremely and continuously variable’, with many ‘inadequate as diagnostic criteria for subgeneric separation’ (Ando Citation1987, p. 82). In a couple of cases at least, morphologic differences do not intergrade: presence or absence of radial sculpture on the posterior ear is one example, another is the presence of the growth stop (or step) in species of Maorimonotis but in no other ‘subgenus’. As noted above, the subgeneric divisions introduced over the last 30 years have stood the test of time and have proven useful. Utility is one of the subsidiary criteria for judging the acceptability of such divisions, especially for fossil groups, and this is relevant to the chronostratigraphic scheme ().
A review of the characteristic morphology of each generic grouping provides the opportunity to reconsider the nomenclature, its rank and application. Few fossil molluscan genera have had more subspecies proposed than Monotis (v., e.g. in M. salinaria, ochotica, scutiformis and zabaikalica in Payevskaya Citation1985), and it is here proposed to follow the modern trend towards a simplified nomenclature in which subgenera and subspecies are less favoured than previously. Using this approach with Monotis, all subgenera are elevated to generic status and subspecies either raised to species level or synonymised.
This change raises the question of a collective name for species that have previously been included under ‘Monotis’, and the simplest solution is to use the informal term ‘monotis’. The family name Monotidae (or monotid) is unsatisfactory for this purpose because it includes other genera (e.g. Otapiria Marwick). This will be the procedure followed below in discussing the group as a whole.
Class Bivalvia Linnaeus, 1758
Subclass Autobranchia Grobben, 1984
Superorder Pteriomorphia Beurlen, 1944
Order Pectinida Gray, 1854
Superfamily Monotoidea Fischer, 1886
Family Monotidae Fischer, 1886
Genus Monotis Bronn, Citation1830
Type species (by SD). Pectinites salinarius Schlotheim, Citation1820.
Diagnosis. Thin-shelled, of small to medium size, subequivalved, weakly inflated with right valve less so than left, with unsculptured well-separated posterior ear with broad moderately deep posterior sinus; ornament of fine rounded radial ribs (after Grant-Mackie Citation1978a for subgenus Monotis).
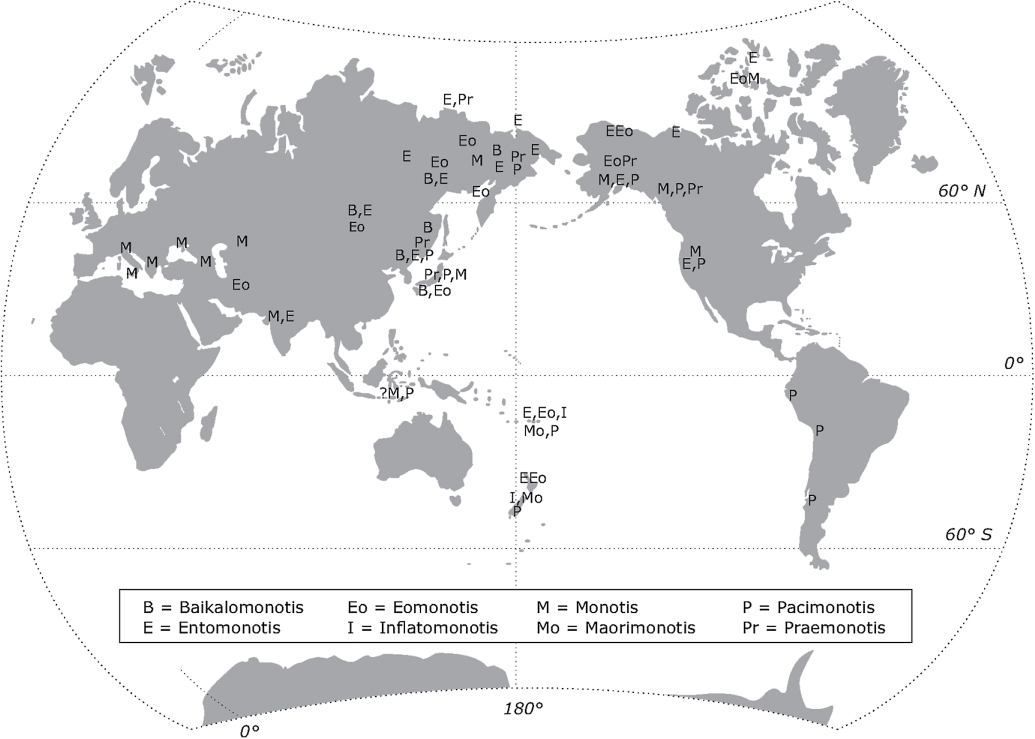
What was the type subgenus is widespread across the Northern Hemisphere from the European Alps to Arctic Canada (see for distribution of this and the other monotis genera).
As a byssally attached bivalve, members of this genus and most other monotids have been suggested to have lived on the laminae of marine algae, given the presence throughout life of a functional byssal notch and in the absence of evidence for any other local substrate (Grant-Mackie Citation1980a).
Various subspecies have been included by previous authors in the type species. Payevskaya (Citation1985) listed s. taurica (Moiseev, Citation1939), s. haueri (Kittl, Citation1912), s. anjuensis (Bychkov, in Kiparisova et al. Citation1966), and s. alaskana (Smith, Citation1927). Pseudomonotis caucasica var. taurica resembles M. salinaria in nearly all characters; it is said by Payevskaya (1958) to differ by having finer ribbing and less inflated valves. She also stated that shell convexity may, however, vary from slightly to moderately convex. This ‘subspecies’ is therefore synonymised with M. salinaria. The other taxa are here accepted as full species, as already regarded by other authors.
The following species are those here regarded as members of the genus Monotis (in this and following generic lists, ‘syn’ = junior synonym, ‘type’ = type species, ‘?’ = uncertain as to generic location or validly separate status): M. alaskana (Smith, Citation1927); M. anjuensis Bychkov & Efimova, 1966; M. digona Kittl, 1912; M. haueri Kittl, 1912; M. hoernesi Kittl, 1912; M. kaukasica (Wittenburg, 1923); M. limaeformis Gemmellaro, 1882; ?M. megalota Mojsisovics, 1873; M. salinaria (Schlotheim, Citation1820) (type) (syn. M. taurica (Moiseev, Citation1939).
Genus Entomonotis Marwick, Citation1935
Type species (OD). Monotis salinaria richmondiana Zittel, Citation1864.
Diagnosis. Thin-shelled, medium to large size, obliquely suboval outline, strongly inequivalved with moderately to highly inflated left valve and nearly flat right valve; posterior auricle moderate to large, unsculptured, well separated from disc; with well-differentiated radial sculpture, primary ribs varying from narrow, high and sharp to broad, low and rounded (amended from Grant-Mackie Citation1978a).
This genus characterises the middle and upper portions of the monotis sequence, correlated by Grant-Mackie (Citation1985) with the late Norian Cordilleranus Zone. It is represented in the Southwest Pacific region by the type species and by E. acutecostata (Trechmann, Citation1918), formerly regarded (e.g. Grant-Mackie Citation1978d) as a chronosubspecies of E. richmondiana. It is here accorded full specific status. E. acutecostata is still accepted as a direct ancestor of E. richmondiana and it characterises a zone immediately preceding that of E. richmondiana in both areas (Grant-Mackie Citation1985; ).
This genus includes E. ochotica (Keyserling) which has included subspecies ambigua, densistriata, eurhachis, pachypleura and sparsicostata (all of Teller, Citation1886). Some authors have already treated the latter two as full species (Kobayashi & Ichikawa Citation1949; Tamura Citation1965; Payevskaya Citation1985). All except densistriata were submerged in E. ochotica by Ando (Citation1987). E. ambigua and eurhachis were regarded as infraspecific forms of ochotica, rather than subspecies; Grant-Mackie (1Citation978a) viewed them as doubtfully separable from ochotica, and they are here submerged in that species. In addition, E. o. longa (Kiparisova, Citation1932) was synonymised with ochotica as a valve elongated by lateral compression (Grant-Mackie, Citation1978a). The inadequately known E. o. wingia (Bando, Citation1961) has some features that differ from those of E. ochotica and E. densistriata and may be a separate taxon, but it requires more study.
This genus is widespread in Asia, North America and the southwest Pacific; it could be expected in Indonesia but its presence there is unconfirmed so far. It includes the largest known specimens of Monotidae, E. richmondiana, which are up to 122 mm long.
Species regarded as belonging to the genus Entomonotis are: E. acutecostata (Trechmann, Citation1918); E. densistriata (Teller, Citation1886); E. intermedia (Trechmann, Citation1918); E. ochotica (Keyserling, Citation1848) (syn. E. ambigua (Teller, Citation1886), E. eurhachis (Teller, Citation1886), E. o. longa (Kiparisova, Citation1932)); E. pachypleura (Teller, Citation1886); E. posteroplana (Westermann, Citation1962); E. richmondiana (Zittel, Citation1864) (type); E. sparsicostata (Teller, Citation1886); and ?E.wingia (Bando, Citation1961).
Genus Eomonotis Grant-Mackie, Citation1978a
Type species (by OD). Pseudomonotis scutiformis var. typica Kiparisova, Citation1936.
Diagnosis. Small to medium size, subequivalve to moderately inequivalve, with small- to medium-sized obliquely truncated, poorly separated costate posterior auricle, posterior sinus weak or absent; ornament of numerous fine radial ribs (after Grant-Mackie Citation1978a).
This covers a large group of species and its members are generally quite clearly differentiated from those of the other ‘subgenera’ by their small size, fine ribbing and costate ear. It was first given generic status by Tozer (Citation1980).
Members of this genus are distributed widely in Asia, North America and the southwest Pacific in strata of the Columbianus Zone of the Norian; in addition, E. jakutica extends up into the Cordilleranus Zone in North America.
The type species, E. typica, has been regarded as a subspecies of E. scutiformis by many earlier workers (e.g. Kiparisova Citation1936; Vozin & Tikhomirova Citation1964) but as a full species by Hase (Citation1961) and Westermann (Citation1966). Payevskaya (Citation1985) listed E. typica as a ‘forma’ of E. scutiformis. This latter taxon has also been listed with subspecies daonellaeformis (Kiparisova, Citation1961), kolymica (Kiparisova, Citation1936), and setakanensis (Kiparisova, Citation1961). E. daonellaeformis was originally described as a full species, reduced to a subspecies of Monotis typica by Kiparisova et al. (Citation1966), and is here restored to the species level. Eomonotis kolymica is represented by only a few left valves, with no known right valves (Payevskaya Citation1985); it is said by her to be similar to E. daonellaeformis, differing in outline and with coarser ribs, and can tentatively be submerged in E. typica. E. setakanensis is separated from E. scutiformis, with sparser ventrally flattening ribs and shorter hingeline than E. daonellaeformis, and appears to warrant separate species status.
Eomonotis aotearoa (Grant-Mackie, Citation1978a) was originally described as a subspecies of Monotis scutiformis pinensis Westermann, Citation1962, which Bychkov (Citation2007) selected as type for his new genus Praemonotis. E. aotearoa cannot be accommodated in this genus because of its well-ribbed posterior ear and strong ornament on the posterior area of the valve; it fits without problem in Eomonotis. However, the species is known from only a few valves (four right, one left) and its valid separation is uncertain.
Monotis jakutica (Teller, Citation1886) has included two subspecies in addition to the nominate form, E. j. mabara and multistriata, both of Kobayashi & Ichikawa (Citation1949). The latter was submerged in jakutica by Westermann (Citation1973) and in ochotica aequicostata (Kiparisova, Citation1940) by Kiparisova et al. (Citation1966). It is accepted as an invalidly separated form here. E. j. mabara was erected as a subspecies of M. tenuicostata, then regarded by Westermann (Citation1973) as a subspecies of E. jakutica from which it differs only in size; it is here regarded as a synonym of that species. E. jakutica was raised to full specific rank by Ando (Citation1987) and Silberling & Grant-Mackie (Citation1987).
Grant-Mackie (1Citation981a) erected subspecies within E. murihikuensis and E. rauparaha. In the former, it is recommended that E. murihikuensis remain separated from E. taringatura (each raised to species level) because subsequent work has confirmed their stratigraphic separation and there seems little or no blurring of the original morphologic basis for separation. However, I have some doubts about the validity of their separation, having noted that the more finely ribbed E. murihikuensis is generally found in fine-grained rocks (siltstones and fine sandstones), with E. taringatura occurring more frequently in medium sandstones in which the finest elements of the ribbing could have been destroyed during preservation; ribbing numbers, outline, inflation, and size are the same. Future more detailed comparisons could lead to the synonymising of these two forms, despite the stratigraphic separation.
In the case of E. rauparaha, three subspecies were originally proposed (Grant-Mackie Citation1981a), mokaui defining the basal zone of the Monotis sequence and the other two forms also confined to this zone. One subspecies, E. r. aries, is represented by a dozen incomplete and poorly preserved valves from one locality only, and I have long been uncertain of its valid separation. All three ‘subspecies’ are very similar morphologically and there seems little justification for continued separation. Accordingly I recommend the suppression of the names E. rauparaha aries and E. r. mokaui in favour of the nominate subspecies and, as a consequence, renaming the Mokaui Zone as Rauparaha Zone.
Grant-Mackie (1Citation980a) suggested that these small light shells occupied the distal-most parts of the algal fronds on which they may have lived.
Species now considered to belong in the genus Eomonotis are: ?E. aotearoa (Grant-Mackie, 1981); E. daonellaeformis (Kiparisova, Citation1961); E. inaequivalvis (Bronn, 1830); E. iwaiensis (Ichikawa, Citation1951); E. jakutica (Teller, Citation1886) (syn. E. j. multicostata (Kobayashi & Ichikawa, Citation1949), E. j. mabara (Kobayashi & Ichikawa, Citation1949)); E. murihikuensis (Grant-Mackie, 1981); E. obtusicostata (Westermann, Citation1966); E. rauparaha (Grant-Mackie, 1981) (syn. E. aries (Grant-Mackie, 1981), E. mokaui (Grant-Mackie, 1981)); E. scutiformis (Teller, Citation1886); E. setakanensis (Kiparisova, Citation1961); ?E. taringatura (Grant-Mackie, 1981); E. typica (Kiparisova, Citation1936) (type) (syn. E. kolymica (Kiparisova, Citation1936)); and E. wairakae (Grant-Mackie, 1981).
Genus Maorimonotis Grant-Mackie, Citation1978c
Type species (by OD). Monotis routhieri Avias, 1953; Norian, Cordilleranus Zone, New Zealand–New Caledonia.
Diagnosis. Shells equivalve to subequivalve, obliquely oval to suboval, with small to obsolete unsculptured posterior ear, relatively thick-shelled; radial ribbing strong to obsolete or absent, broad and blunt when present (after Grant-Mackie Citation1978a).
Monotis routhieri was introduced with minimal description but a good figure by Avias (Citation1953), and is more fully described by Brûlez (Citation1957) in an unpublished thesis and by Grant-Mackie (1Citation978c).
This group was first raised to generic status by Tozer (Citation1980). It is a group of four species, confined to the New Caledonia–New Zealand region. They form a coherent evolutionary and stratigraphic sequence characterised by progressive weakening of the ribbing, increase in inflation of the right valve, and later development of a ‘growth stop’. The latter constitutes a change in shell profile considered by Grant-Mackie (Citation1979, Citation1978a) as having been associated with an ecologic trend from epifaunal to a semi-infaunal mode of life. These features clearly separate this group from all other species formerly placed in the genus Monotis. Although the oldest species M. maniapotoi is least differentiated from Entomonotis, from which it probably sprang, full generic separation is clearly warranted.
The following species are those regarded here as belonging in the genus Maorimonotis: M. awakinoensis (Grant-Mackie, 1978); M. calvata (Marwick, Citation1953); M. maniopotoi (Grant-Mackie, 1978); and M. routhieri (Avias, 1953) (type).
Genus Pacimonotis Grant-Mackie & Silberling, Citation1990
Type species (by OD). Monotis subcircularis Gabb, Citation1864; Norian, Cordilleranus Zone, North America.
Diagnosis. Moderate to large size, subcircular to posteroventrally elongate; subequivalve, gently inflated, RV less than LV; umbones low, not prominent; posterior auricle of medium size, unsculptured, relatively acutely truncated and with broad deep sinus behind; well separated from disc; byssal ear small; radial sculpture moderately well differentiated, with narrow, high, sharp or rounded primaries and markedly weaker subsidiary orders; most members also possess fine dense concentric striae, especially in interstices between primary plicae (after Grant-Mackie & Silberling Citation1990).
Pacimonotis subcircularis has had three subspecies included in it, apart from the nominate form – gigantea (Avias, 1953), sibirica (Bychkov, 1966), and discordans (Grant-Mackie, Citation1978b) –and these are raised to species level.
This genus is widespread in the circum-Pacific region.
The following species are here regarded as members of the genus Pacimonotis: P. callazonensis (Westermann, Citation1962); P. discordans (Grant-Mackie, 1978); P. gigantea (Avias, 1953); P. sibirica (Bychkov, 1966); and P. subcircularis (Gabb, Citation1864) (type).
Genus Inflatomonotis Grant-Mackie, Citation1978a
Type species (by OD). Monotis salinaria var. hemispherica Trechmann, Citation1918; Middle Norian, New Zealand–New Caledonia.
Diagnosis. Small, subcircular to suboval, markedly inequivalve, with strongly inflated left valve and nearly flat right valve, posterior ear smooth; with strong broad radial ribbing (after Grant-Mackie Citation1978a).
The type species was included in Entomonotis by Ando (Citation1987).
In New Zealand this species is confined to the Murihikuensis Zone (Norian, Columbianus Zone); it has recently also been recorded from New Caledonia (unpublished record by P. Maurizot, Nouméa) but is unknown elsewhere.
Westermann (Citation1962) reported Monotis sp. cf. M. hemispherica from Canada. Grant-Mackie (1Citation980b) agreed that the specimen, a left valve, was very similar to the New Zealand material and could represent the only member of the genus so far known outside the southwest Pacific.
Listed below are those species regarded as being members of the genus Inflatomonotis: hemispherica (Trechmann, Citation1918) (type); and warepana (Grant-Mackie, Citation1980b).
Genus Praemonotis Bychkov, Citation2007
Type species (by OD). Monotis scutiformis pinensis Westermann, Citation1962; Middle Norian, British Columbia, Canada.
Diagnosis. Shell small, from weakly to strongly inflated, moderately to markedly inequivalve, with a weakly to well-separated small posterior auricle with weakly angled posterior; ornament moderately strong and rounded but fading posteriorly, weak or absent on posterior auricle (after Bychkov Citation2007, translated from the Russian by G-M).
This genus includes a couple of species formerly allotted to Eomonotis. P. pinensis was described by Westermann (Citation1962) as a subspecies of E. scutiformis with poorly or un-ornamented well-separated posterior auricle, but was raised to full specific rank by Vozin & Tikhomirova (Citation1964).
Entomonotis iwaensis Ichikawa, Citation1951, synonymised with E. densistriata by Ando (Citation1987), was included in Praemonotis by Bychkov (Citation2007) but its possession of a costate posterior auricle excludes it from that genus. This character had earlier led Grant-Mackie (1Citation978a) to place it more appropriately in Eomonotis; this character also excludes it from Praemonotis.
Among the species included by Bychkov (Citation2007) in this genus is the New Zealand form P. diptonensis, which Grant-Mackie (1981) had described under Eomonotis. Its features include moderately strong radials across the disc and only slightly weaker on the posterior ear, which is half the postumbonal length, characters not typical of Praemonotis. The species is therefore returned to Eomonotis. No valid member of this genus has been recorded from New Zealand or New Caledonia.
Listed below are species here regarded as being members of the genus Praemonotis: P. pinensis (Westermann, Citation1962) (type); and P. versicostata (Bychkov, 1966).
Genus Baikalomonotis Bychkov, Citation2007
Type species (by OD). Pseudomonotis zabaikalica Kiparisova, Citation1936; Norian, Trans-Baikal, Siberia.
Diagnosis. Shell small to large size, obliquely oval to subcircular in outline; sculpture strong but fading, with concentric wrinkles and short shallow radial ribs and grooves not always developed; posterior auricle of moderate size, poorly separated from disc; small anterior auricle on right valve directed forwards and upwards (after Bychkov Citation2007, translated from the Russian by G-M).
Some previous authors have included in Baikalomonotis zabaikalica the central member of this genus, subspecies or varieties planocostata (Kiparisova, 1960) and semiradiata (Ichikawa, Citation1958). Both were regarded by Payevskaya (Citation1985) as full species and this is followed here. B. notabilis (Oleynikov & Payevskaya Citation1980), synonymised with zabaikalica by Ando (1987), is another member of the genus.
Bychkov (Citation2007) listed two further species in his genus, B. cycloidea and B. sublaevis, both of Teller, Citation1886 and regarded by Kiparisova et al. (Citation1966) as related to B. zabaikalica. Westermann (Citation1973) suggested the two were synonyms, with sublaevis having page priority. Their identity is an issue still to be clarified. Each was based upon few specimens and they have similar morphology.
Monotis mukaihatensis Hase, Citation1961 was accepted as a valid taxon by Grant-Mackie (1Citation978a) but was unable to be allotted confidently to any of his subgenera. Given its occurrence in Japan, its small and inequivalve nature with poorly separated posterior auricle, concentric wrinkles and poorly developed radial ornament (see Ando 1987), allocation to Baikalomonotis must be considered. Against this is its possession of a small smooth posterior auricle and its occurrence low in the monotis sequence. Until further morphological details are known it should be included tentatively in Baikalomonotis.
This genus is confined in its distribution to eastern Asia.
Species now included in Baikalomonotis are: ?B. mukaihatensis (Hase, Citation1961); B. notabilis (Oleynikov & Payevskaya Citation1980); B. planocostata (Kiparisova, 1960); B. semiradiata (Ichikawa, 1957); ?B. sublaevis (Teller, Citation1886) (syn.?B. cycloidea (Teller, Citation1886)); and B. zabaikalica (Kiparisova, Citation1936) (type).
Many other species, subspecies and varieties have been included in monotis but have subsequently been synonymised or transferred to other genera. They are therefore not considered here.
Summary
From the various nomenclatural schemes recently recommended for application to species included in Monotis, that favoured here is use of the subgenera of Marwick (Citation1935), Grant-Mackie (1Citation978a), Grant-Mackie & Silberling (Citation1990) and Bychkov (Citation2007) raised to generic level, with subspecies raised to species level or synonymised (taurica (Moiseev, Citation1939) with Monotis salinaria, ambigua Teller, eurhachis Teller, and longa Kiparisova withEntomonotis ochotica, kolymica Kiparisova with Eomonotis typica, mabara Kobayashi & Ichikawa and multistriata Kobayashi & Ichikawa with Eomonotis jakutica, aries Grant-Mackie and mokaui Grant-Mackie with Eomonotis rauparaha).There remain several taxonomic challenges still to be addressed however, especially including consideration of the status of some names used in the Northern Hemisphere, namely Entomonotis ochotica wingia (Bando, Citation1961), Baikalomonotis cycloidea (Teller, Citation1886), Baikalomonotis sublaevis (Teller, Citation1886), Baikalomonotis mukaihatensis (Hase, Citation1961) and, in the southwest Pacific, Eomonotis aotearoa, murihikuensis and taringatura (Grant-Mackie, Citation1981a).
Acknowledgements
Louise Cotterall, University of Auckland School of the Environment, is gratefully acknowledged for the illustrations. Thanks are due to: Alan Beu, GNS Science, Lower Hutt for helpful comments on the modern understanding of the subspecies issue and for constructive criticism of the text; Hamish Campbell, GNS Science; Neville Hudson, Auckland University School of the Environment; Donald MacFarlan, New Plymouth for helpful comments on earlier versions of the manuscript; an anonymous reviewer; and Kristin Garbett and Pauline Muir, GNS Science library, for the provision of references.Associate Editor: Associate Professor Kathy Campbell.
Related Research Data
References
- Ando H 1987. Paleobiological study of the Late Triassic bivalve Monotis from Japan. University Museum, University of Tokyo, Bulletin 30. 109 p
- Avias J 1953. Contribution à l’étude stratigraphique et paléontologique des formations antécretacées de la Nouvelle Calédonie centrale. Science de la Terre 1: 1–267.
- Bando Y 1961. Note on the upper Triassic Monotidae from the Nariwa basin, Okayama Prefecture, Japan. Kagawa Daigaku Gakugei Gakubu Kenkyu Hokoku (Research Report of the Kagawa Liberal Arts Faculty) 102: 1–9.
- Bronn H 1830. Über die Muschel—Versteinerungen des süddeutschen Steinsalzgebirges, welchesbisher dem namen Pectinites salinarius zusammenbegriffen wurden. Jahrbuch für Mineralogie 1: 279–285.
- Brûlez C 1957. Étude monographique des Pseudomonotidae Newell du Trias supérieur de Nouvelle-Calédonie. Unpublished thesis. Nancy, France, University of Nancy. 82 p.
- Bychkov Iu. 2007. Some new genera and subgenera of the Late Triassic bivalves and ammonoids of north-east Asia. Tezisy Dokladov Vserossiisokoi Nauchnoi Konferentsii, Magadan: 43–44. ( in Russian)
- Campbell HJ, Grant-Mackie JA, Paris J-P 1985. Geology of the Moindou-Téremba area, New Caledonia. Stratigraphy and structure of the Téremba Group (Permian–Lower Triassic) and Baie de St-Vincent Group (Upper Triassic–Lower Jurassic). Géology de la France 1: 19–36.
- Gabb WM 1864. Description of the Triassic fossils of California and adjacent territories. Geological Survey of California, Palaeontology 1: 19–35.
- GNS Science 2012. New Zealand geological timescale v. 2012/1. [Chart]. Lower Hutt, Institute of Geological & Nuclear Sciences Ltd. 2 p.
- Grant-Mackie JA 1978a. Subgenera of the Upper Triassic bivalve Monotis. New Zealand Journal of Geology & Geophysics 21: 97–111.
- Grant-Mackie JA 1978b. Systematics of New Zealand Monotis (Upper Triassic Bivalvia): subgenus Entomonotis. New Zealand Journal of Geology & Geophysics 21: 483–500.
- Grant-Mackie JA 1978c. Sytematics of New Zealand Monotis (Upper Triassic Bivalvia): subgenus Maorimonotis. Journal of the Royal Society of New Zealand 8: 293–322.
- Grant-Mackie JA 1978d. Status and identity of the New Zealand Upper Triassic bivalve Monotis salinaria var. richmondiana Zittel, 1864. New Zealand Journal of Geology & Geophysics 21: 375–402.
- Grant-Mackie JA 1979. Environment and mode of life of the Triassic bivalve Monotis (abstract). Abstracts 49th ANZAAS Congress, Auckland 1: 139.
- Grant-Mackie JA 1980a. Mode of life and adaptive evolution in the cosmopolitan Triassic bivalve Monotis. Journal of the Malacological Society of Australia 4: 242–243.
- Grant-Mackie JA 1980b. Systematics of New Zealand Monotis (Upper Triassic Bivalvia): subgenus Inflatomonotis. New Zealand Journal of Geology & Geophysics 23: 629–637.
- Grant-Mackie JA 1981a. Systematics of New Zealand Monotis (Upper Triassic Bivalvia): subgenus Eomonotis. New Zealand Journal of Geology and Geophysics 23: 639–663.
- Grant-Mackie JA 1981b. New Zealand Warepan (Upper Triassic) sequences: Murihiku Supergroup of the North Island. Journal of the Royal Society of New Zealand 11: 231–256.
- Grant-Mackie JA 1985. The Warepan Stage of the Upper Triassic: redefinition and subdivision. New Zealand Journal of Geology and Geophysics 28: 701–724.
- Grant-Mackie JA, Silberling NJ 1990. New data on the Upper Triassic bivalve Monotis in North America, and a new subgenus. Journal of Palaeontology 64: 240–254.
- Hase A 1961. A find of Monotis (Entomonotis) from eastern Yamaguchi Prefecture, Japan. Transactions & Proceedings of the Palaeontological Society of Japan, n.s. 42: 79–87.
- Ichikawa K 1951. Note on the Entomonotis-bearing Triassic formation at Iwai near Itsukaichi, Tokyo Prefecture, with description of a new Entomonotis. Transactions & Proceedings of the Palaeontological Society of Japan, n.s. 2: 43–48.
- Ichikawa K 1958. Zur Taxonomie und Phylogenie der triadischen ‘Pteriidae’ (Lamellibranch) mit besonderer berücksichtigung der Gattungen Claraia, Eumorphotis, Oxytoma und Monotis. Palaeontographica, A 111: 131–212.
- Keyserling A 1848. Fossile Mollusken. In: Middendorff A ed. Reise in den ousstersten Norden und Osten Siberiens. St. Petersburg, Buchdr, der K. Akademie der Wissenschaften. Pp. 1–20.
- Kiparisova LD 1932. Contribution to the stratigraphy of the marine Trias of eastern Transbaikal. Transactions of the Geological Prospecting Service of the USSR 111. ( in Russian)
- Kiparisova LD 1936. Upper Triassic pelecypods from the Kolyma-Indigurka Land. Trudy Arktichesky Institut 30: 71–136. ( in Russian)
- Kiparisova LD 1940. New faunas of the upper Triassic of Verkhoyansk. Trudy Arktichesky Institut 164. ( in Russian)
- Kiparisova LD 1961. Paleontological fundamentals for the stratigraphy of Triassic deposits of the Primorye region. Trudy Vysesoyuznogo Nauchno-Isselyedovatelskogo Geologicheskogo Instituta (VSEGEI), n.s., 48: 1–278. ( in Russian)
- Kiparisova LD, Bychkov Iu, Polubotko IV 1966. Upper Triassic bivalve molluscs from northeast USSR 8. Magadan, VSEGEI, USSR Ministry of Geology. 312 p. ( in Russian)
- Kittl E 1912. Materialen zu einer Monographie der Halobiidae und Monotidae der Trias. Resultat der Wissenschaftliche Erforschung Balatonsees 1, Paläontologie 2: 1–229.
- Kobayashi T, Ichikawa K 1949. Late Triassic ‘Pseudomonotis’ from the Sakawa basin in Shikoku, Japan. Japanese Journal of Geology & Geography 21: 245–262.
- Marwick J 1935. Some new genera of the Myalinidae and Pteriidae of New Zealand. Transactions & Proceedings of the Royal Society of New Zealand 65: 295–303.
- Marwick J 1953. Divisions and faunas of the Hokonui System (Triassic and Jurassic). New Zealand Geological Survey Paleontological Bulletin 21: 141 p.
- Moiseev AS 1939. New data on the Upper Triassic of north Caucasus and the Crimea. Doklady Akademii Nauk SSSR 23: 816–818. ( in Russian)
- Oleynikov A, Payevskaya EB 1980. New type of Upper Triassic monotid from Primorye. New types of plants and invertebrates in the USSR 5. Moscow, Izd, Nauka. 159 p. ( in Russian)
- Payevskaya EB 1985. The Late Triassic molluscan genus Monotis Bronn (an experiment in logical-mathematical research). A.P. Karpinsky All-Union Geological Research Institute Trudy, n.s., 327. 157 p. ( in Russian)
- Schlotheim EF 1820. Die Petrefaktenkunde aus ihrem jetzigen Standpunkte durch die Beschreibung seiner Sammlung Versteineter und fossiler Ûberneste des Their—und Pflanzenreich erläutet. Gotha, Becker'sche Buchhandlung. 437 p.
- Silberling NJ, Grant-Mackie JA, Nichols KM 1997. The Late Triassic bivalve Monotis in accreted terranes of Alaska. US Geological Survey Bulletin 2151. 21 p.
- Smith JP 1927. Upper Triassic marine invertebrate faunas of North America. US Geological Survey Professional Papers 141. 135 p.
- Tamura M 1965. Monotis (Entomonotis) from Kyushyu, Japan. Memoirs of the Faculty of Education, Kumamoto University 1, 13: 42–69.
- Teller F 1886. Die Pelecypoden-Faunen von Werchojansk in Ostsiberien. Mémoires de l'Académie des Sciences de St-Petersburg, Serie 7, 33: 103–137.
- Tozer ET 1980. Latest Triassic (Upper Norian) ammonoid and Monotis faunas and correlations. Rivista Italiana Paleontologia 85: 843–876.
- Trechmann CT 1918. The Trias of New Zealand. Quarterly Journal of the Geological Society 73: 165–246.
- Vozin VF, Tikhomirova VV 1964. Paleontological field atlas of bivalve and cephalopod molluscs of northeast USSR. Moscow, Institute of Geology, Yakutsk Branch, Siberian Section, USSR Academy of Sciences. Pp. 1–194. ( in Russian)
- Westermann GEG 1962. Succession and variation of Monotis and the associated fauna in the Norian Pine River Bridge section, British Columbia (Triassic, Pelecypoda). Journal of Palaeontology 36: 745–792.
- Westermann GEG 1966. New occurrences of Monotis from Canada (Triassic Pelecypoda). Canadian Journal of Earth Sciences 3: 975–986.
- Westermann GEG 1973. The Late Triassic bivalve Monotis. In: Hallam A ed. Atlas of palaeobiogeography. Amsterdam, London, New York, Elsevier. Pp. 251–258.
- Zittel A 1864. Fossile Mollusken und Echinodermen aus Neuseeland. Reise der österriche Fregatte Novara, Geologische Theil 1: 17–68.