
Abstract
The femoral hemispherical articular head was regarded as autapomorphic of Mauisaurus haasti Hector, 1874 from the upper Campanian of New Zealand. Review of three Maastrichtian taxa, Aristonectes quiriquinensis from Chile, Aristonectes sp. from Antarctica and Kaiwhekea katiki from New Zealand, reveals that the femoral hemispherical head is a common character among aristonectines. This feature can therefore be discarded as an autapomorphy of Mauisaurus haasti. Propodials of A. quiriquinensis are shown to be the same as those in two coeval Chilean specimens previously referred to Mauisaurus sp.; in consequence, these are now referred to A. quiriquinensis. Additionally, specimens referred to A. quiriquinensis allow confirmation that during ontogeny the humerus and the femur change from a flat capitulum in juveniles to hemispherical heads of both the humeri and femora in the adult stage. The evidence shows that the presence of Mauisaurus along the southeastern Pacific margin cannot be verified to date.
Introduction
Late Cretaceous records of elasmosaurids (Sauropterygia, Plesiosauria) within the Weddellian Biogeographic Province (sensu Zinsmeister Citation1979; WBP hereafter) includes records from Antarctica, Argentina, Chile and New Zealand (Welles & Gregg Citation1971; Gasparini & Goñi Citation1985; Cruickshank & Fordyce Citation2002; Hiller et al. Citation2005; Wilson et al. Citation2005; Consoli & Stilwell Citation2009; O’Gorman Citation2012, Citation2013; Otero et al. Citation2012, 2014a,b; Hiller et al. Citation2014; O’Gorman et al. Citation2014). In New Zealand, extensive descriptions of several proposed taxa have been presented since the late 19th century. New Zealand elasmosaurids include at least ten different former species within two genera, most of them currently considered invalid taxa due the lack of diagnostic elements (see Welles & Gregg Citation1971).
Among these, Mauisaurus haasti Hector, 1874 is a ‘historical survivor’. It is a taxonomic concept originally based on eight specimens from different localities in New Zealand (Hector Citation1874; Hiller et al. Citation2005). The original description by Hector (Citation1874) includes a table indicating a single specimen with ‘coracoids, scapula and paddles’ from the Jed River, Cheviot Hills and seven other specimens from Amuri (now known as Haumuri Bluff) and Waipara River. Hector based the description of Mauisaurus haasti on the ‘coracoids’ (actually pubes), the ‘scapula’ (ilium) and the ‘forelimb’ (actually the hind limb). On the last, a very unusual feature noted by Hector was the presence of a propodial ‘greatly thickened at the proximal end to form a hemispherical articular surface and an expanded trochanter-like process, which are separated by a deep bicipital groove, giving it more the look of a cetacean than a reptilian limb-bone’ (Hector Citation1874, pp. 347–348). The distinctive morphology of the propodial was later noted by Welles (Citation1962) who commented on the hypodigm of the taxon and selected as the lectotype the same specimen (8a) of Hector comprising the pubes, ilium and hind limb. Welles (Citation1962) also indicated that the species Mauisaurus brachiolatus Hector, 1874, originally based on a large humerus, should be considered as nomen dubium. In addition, Welles (Citation1962) realised that the specimen chosen as the lectotype of Mauisaurus haasti Hector, 1874 was indeed a pelvic girdle and a hind limb instead of a pectoral girdle and a forelimb as described originally. Subsequently, this author considered this specimen as unique, due to the femur being ‘quite different from other plesiosaurs in having a hemispherical capitulum with a large trochanter that dips steeply posteriorly and is separated from the capitulum by a broad groove’ (Welles Citation1962, p. 75). A better formalisation of the lectotype was included in Welles & Gregg (Citation1971) who mentioned for the first time the catalogue number of the Hector’s specimen (DM R1529) and indicated a Haumurian (Campanian–Maastrichtian) age for the material. This was later emended by Hiller et al. (Citation2005) through stratigraphic correlations that indicate a middle Campanian age for DM R1529. One of the specimens included in the original hypodigm (8g) was also regarded by Welles & Gregg (Citation1971) as a paralectotype (CM Zfr 92), while three other specimens (8b, c, f) were considered non-diagnostic and left as indeterminate elasmosaurids. The remaining three specimens (8d, e, h) of the original hypodigm have been lost. Finally, Welles & Gregg (Citation1971) referred to Mauisaurus haasti 11 additional specimens from the Campanian–Maastrichtian of New Zealand (DM R878; CM Zfr 88; CM Zfr 89; CM Zfr 90; CM Zfr 103; BM R830; CM Zfr 30; CM Zfr, 95; CM Zfr 102; CM Zfr 104; and CM Zfr 109).
The proposal of Welles & Gregg (Citation1971) regarding the lectotype, paralectotype and additional specimens referred to M. haasti was subsequently used as the basis for further identifications of Late Cretaceous plesiosaur specimens from the WBP. Fostowicz-Frelik & Gaździcki (Citation2001) described a probable associated, postcranial skeleton from middle–upper Maastrichtian beds of Seymour Island (= Isla Marambio), Antarctica, suggesting affinities to the genus Mauisaurus based on the presence of the hemispherical head of the femur. Later, Gasparini et al. (Citation2003) referred two specimens from upper Maastrichtian beds of northern Argentinean Patagonia to cf. Mauisaurus sp., based on morphological affinities of the humerus with those humeri included in referred materials of Welles & Gregg (Citation1971). Hiller et al. (Citation2005) described a new fairly complete skeleton of probable late Campanian age from Ngaroma Station, New Zealand, referring it to M. haasti based on the hemispherical head of the femur.
Following the revision of the genus Mauisaurus by Hiller et al. (Citation2005), Gasparini et al. (Citation2007) reassessed the two specimens previously assigned to cf. Mauisaurus sp. as Elasmosauridae indet. Martin et al. (Citation2007) described a juvenile specimen from Maastrichtian levels of Vega Island, Antarctica, referring it to Mauisaurus sp. based on the presence of a rounded end of the femur, conical ventral process of the coracoids and fork-like gastralia. The latter was present in one of the specimens described by Hector (Citation1874), which was not referred to Mauisaurus haasti but to ‘Plesiosaurus australis’ Owen, Citation1861. Later, Otero et al. (Citation2010) referred to Mauisaurus sp. two putative femora from upper Maastrichtian beds of central Chile that bear distinctive hemispherical heads; finally, Novas et al. (Citation2011) tentatively referred to Mauisaurus a new, relatively complete skeleton from the Maastrichtian of Argentinean Patagonia.
These records appeared to be biogeographically consistent due the common age and distribution of all the specimens, indicating that the genus Mauisaurus was present in New Zealand, Antarctica and southern South America over a time span of 5 Ma (late Campanian–late Maastrichtian). Furthermore, such a palaeobiogeographic distribution of Mauisaurus was commented on by several authors as being consistent with the WBP model (Consoli & Stilwell Citation2009; Roberts et al. Citation2014).
A recently prepared aristonectine from the upper Maastrichtian beds of the López de Bertodano Formation exposed on Seymour (= Marambio) Island, Antarctica proves for the first time the presence of a femur with a hemispherical head in the genus Aristonectes (O’Gorman Citation2013). A re-examination of the holotype of Kaiwhekea katiki Cruickshank & Fordyce, Citation2002 from the early Maastrichtian of New Zealand also revealed the same morphology of the femoral head. Additionally, the recent description of two fairly complete skeletons from central Chile, the holotype (SGO.PV.957, young adult) and referred specimen (SGO.PV.260, juvenile) of Aristonectes quiriquinensis Otero et al. (Citation2014a), and comparison with other Chilean specimens (SGO.PV.135 and SGO.PV.169), provide further evidence that a hemispherical femoral head and a prominent trochanter, as well as a deep fossa between them, are also present in the femur of this species. The Chilean specimens also provide information about humeral morphological changes during ontogeny.
Based on this information, the aims of this paper are: (1) to show that the hemispherical femoral head, previously regarded as an autapomorphy and diagnostic feature of M. haasti, should be rejected since this feature is present in at least two other genera (Aristonectes and Kaiwhekea); (2) to show that the hemispherical head is present not only in femora but also in humeri, and its development depends on the ontogenetic stage of the examined individuals; (3) to re-examine the Chilean specimens previously referred to Mauisaurus sp. and show that they belong to a femur and a humerus of the species Aristonectes quiriquinensis; and (4) to use the new evidence to positively identify the actual laterality of the femora of K. katiki.
Institutional Abbreviations: SGO.PV.: Área de Paleontología, Museo Nacional de Historia Natural, Santiago, Chile; MLP: Museo de La Plata, La Plata, Argentina; CM: Canterbury Museum, Christchurch, New Zealand; DM: Museum of Te papa Tongarewa (former Dominion Museum), Wellington, New Zealand; OU: Department of Geology Museum, University of Otago, Dunedin, New Zealand.
Anatomical Abbreviations: f: fossa; ff: facet for fibula; fh: femoral head; fu: facet for ulna; g: gastralia; hh: humeral head; t: tuberosity of the humerus; tr: trochanter; vt: ventral tuberosity.
Locality and geological setting
The three Chilean specimens discussed here (SGO.PV.135, SGO.PV.169 and SGO.PV.260) were recovered from Las Tablas Bay in the northern part of Quiriquina Island, Biobío Region, central Chile. A fourth specimen (SGO.PV.957) was recovered from Cocholgüe, a small hamlet located on the mainland opposite Quiriquina Island. Las Tablas Bay and Cocholgüe have been considered the type and paratype localities of the Quiriquina Formation, respectively (Biró-Bagóczky Citation1982). The age of the unit is currently considered as late Maastrichtian based on biostratigraphic correlations (Salazar et al. Citation2010). Another specimen discussed here (MLP 89-III-3-1) was recovered from Seymour Island (= Marambio Island), Antarctica. The fossil-bearing level is the uppermost part of the López de Bertodano Formation (Rinaldi et al. Citation1978), currently assigned to the upper Maastrichtian based on biostratigraphic correlations (Crame et al. Citation2004). One specimen reviewed from New Zealand (OU 12649) was recovered from lower levels of the Katiki Formation exposed at Shag Point, north of Dunedin (Cruickshank & Fordyce Citation2002), early Maastrichtian in age. The localities of the reviewed specimens are summarised in .
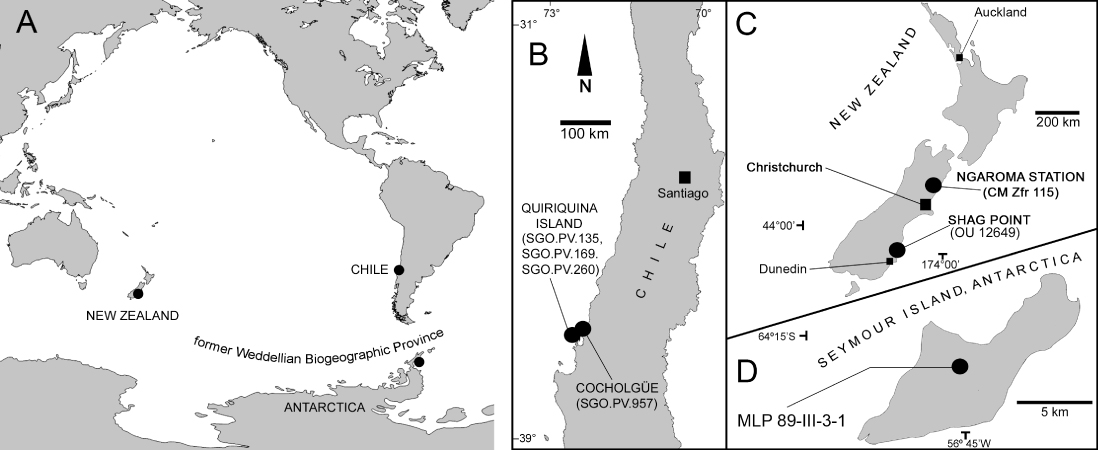
Systematic palaeontology
Plesiosauria de Blainville Citation1835
Plesiosauroidea Welles Citation1943
Xenopsaria Benson & Druckenmiller Citation2014
Elasmosauridae Cope Citation1869
Mauisaurus Hector, 1874
Type species
Mauisaurus haasti Hector 1874. Amuri Bluff, Waipara district (as originally mentioned by Hector Citation1874), New Zealand. Cretaceous.
Lectotype
DM R1529, designated by Welles (Citation1962) and better formalised in Welles & Gregg (Citation1971). It comprises a right hind limb, incomplete illium and a fragmentary pubis. Specimen 8a of Hector (Citation1874). Jed River, Gore Bay, North Canterbury, New Zealand. Probable middle Campanian age based on dinoflagellates (Wilson et al. Citation2005).
Aristonectinae O’Keefe & Street Citation2009 (sensu Otero et al. Citation2012)
Aristonectes Cabrera Citation1941
Type species
MLP 40-XI-14-6 (holotype) Aristonectes parvidens Cabrera Citation1941 (p. 114), part of a skull attached to the mandible, atlas-axis and 21 other cervical vertebrae of which the anterior 16 are articulated, eight caudal vertebrae and an incomplete limb. Cañadón del Loro, northwestern Chubut Province, Argentina. Paso del Sapo Formation, Lefipan Member, Maastrichtian (Gasparini et al. Citation2003).
Synonymy
Gasparini et al. (Citation2003) considered the genus and species Morturneria seymourensis (Chatterjee & Small, 1989; emend. Chatterjee & Creisler 1994), based on a juvenile skull and cervicals from upper Maastrichtian levels of Seymour Island (Antarctica), as a junior synonym of A. parvidens.
Aristonectes sp.
Material
MLP 89-III-3-1 (referred, O’Gorman Citation2013), very large adult, postcranial specimen including 19 cervical, 1 pectoral and 10 caudal vertebrae, left scapula, incomplete left forelimb, incomplete left hind limb preserving the complete femur, ribs and gastroliths. Seymour (=Marambio) Island, Antarctica. López de Bertodano Formation, upper Maastrichtian.
Aristonectes quiriquinensis Otero et al. Citation2014a
Mauisaurus sp.: Otero et al. 2010
()
Holotype
SGO.PV.957. A fairly complete skeleton preserving the partial skull, most of the neck, pectoral girdle, anterior portion of the trunk, both front limbs, and a partial right hind limb with the femur lacking its proximal part. Cocholgüe, Biobío Region, central Chile. Upper levels of the Quiriquina Formation, upper Maastrichtian. SGO.PV.260. Referred specimen. A fairly complete postcranial skeleton. Las Tablas Bay, Quiriquina Island, Biobío Region, central Chile. Quiriquina Formation, upper Maastrichtian.
New referred specimens
SGO.PV.135. Complete left femur; SGO.PV.169. Proximal portion of a left humerus. Las Tablas Bay, north of Quiriquina Island, Biobío Region, central Chile. Quiriquina Formation, upper Maastrichtian.
Remarks
SGO.PV.135 is here confirmed as a left femur, as previously identified by Otero et al. (Citation2010). On the other hand, SGO.PV.169 is anatomically emended, being the proximal portion of a left humerus. Both specimens are a perfect match in size and morphology with the respective femur and humerus of the holotype of A. quiriquinensis and are therefore referred to that species. Prior to this research, the most diagnostic features of Mauisaurus were the presence of a prominent hemispheric articular head of the femur, a large trochanter separated from the articular head by a deep fossa and the presence of a large tuberosity in the ventral surface of the diaphysis (Hector Citation1874; Welles Citation1962; Welles & Gregg Citation1971; Hiller et al. Citation2005). Based on this, Otero et al (Citation2010) referred to Mauisaurus two specimens (SGO.PV.135 and SGO.PV.169) from the upper Maastrichtian of Chile.
Kaiwhekea katiki Cruickshank & Fordyce Citation2002
Type species
OU 12649 (holotype), a fairly complete adult skeleton including casts of both femora. Shag Point, north of Dunedin, New Zealand. Katiki Formation, lower Maastrichtian.
Remarks
The original description of OU 12649 included detailed description and figures of one femur (Cruickshank & Fordyce Citation2002; ). Due to its taphonomic placement in the fossil, a right laterality was assigned for this element. The presence of a large trochanter diagonally oriented over the diaphysis is a feature observed in other elasmosaurids such as the lectotype of M. haasti and the femur SGO.PV.135 referred to A. quiriquinensis. A similar structure is also present in MLP 89-III-3-1 from Antarctica and referred to Aristonectes sp. All these specimens confirm that the diagonal extension of the trochanter is only visible in dorsal view and that it extends from the anteroproximal part of the femur to the posteromedial part of the diaphysis. Such anatomical landmarks are useful for recognising the anatomical identification of the femur, as well as the orientation of the latter. Based on this, the femur of K. katiki (Cruickshank & Fordyce Citation2002; ) can be identified as a left rather than a right femur, while the whole left hind limb is mostly visible in ventral view on the holotype.
Presence of a hemispherical femoral head among elasmosaurids from the WBP
Aristonectes sp. (MLP 89-III-3-1)
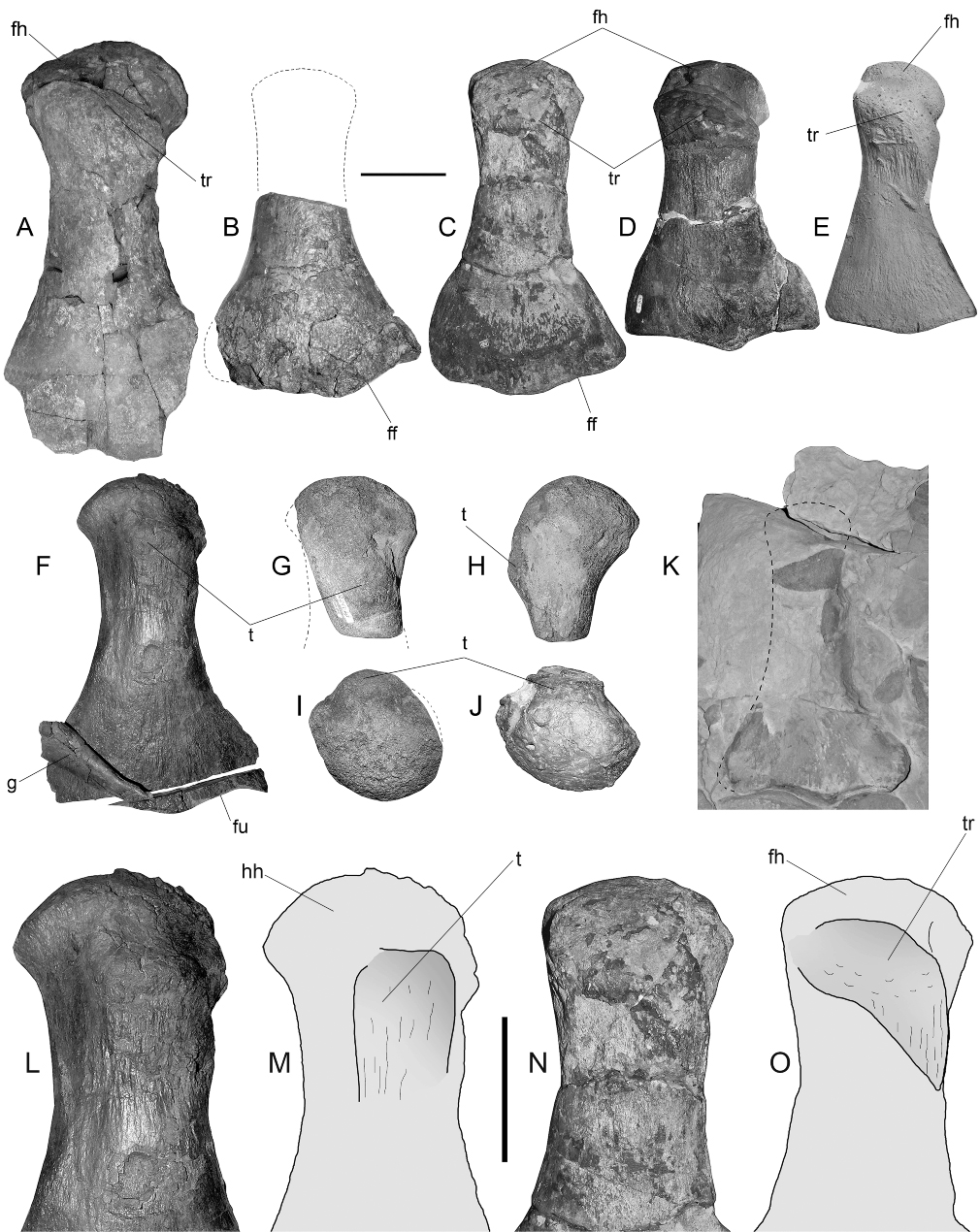
This very large specimen has short cervical vertebrae that are more similar to those of Aristonectes than Kaiwhekea; it is therefore kept as Aristonectes sp. for the moment, awaiting more detailed studies (in progress by JPO’G). The femur of this specimen () exhibits a clear hemispherical head, and the presence of this feature in the genus Aristonectes is therefore verified.
Aristonectes quiriquinensis (SGO.PV.957 and SGO.PV.260)
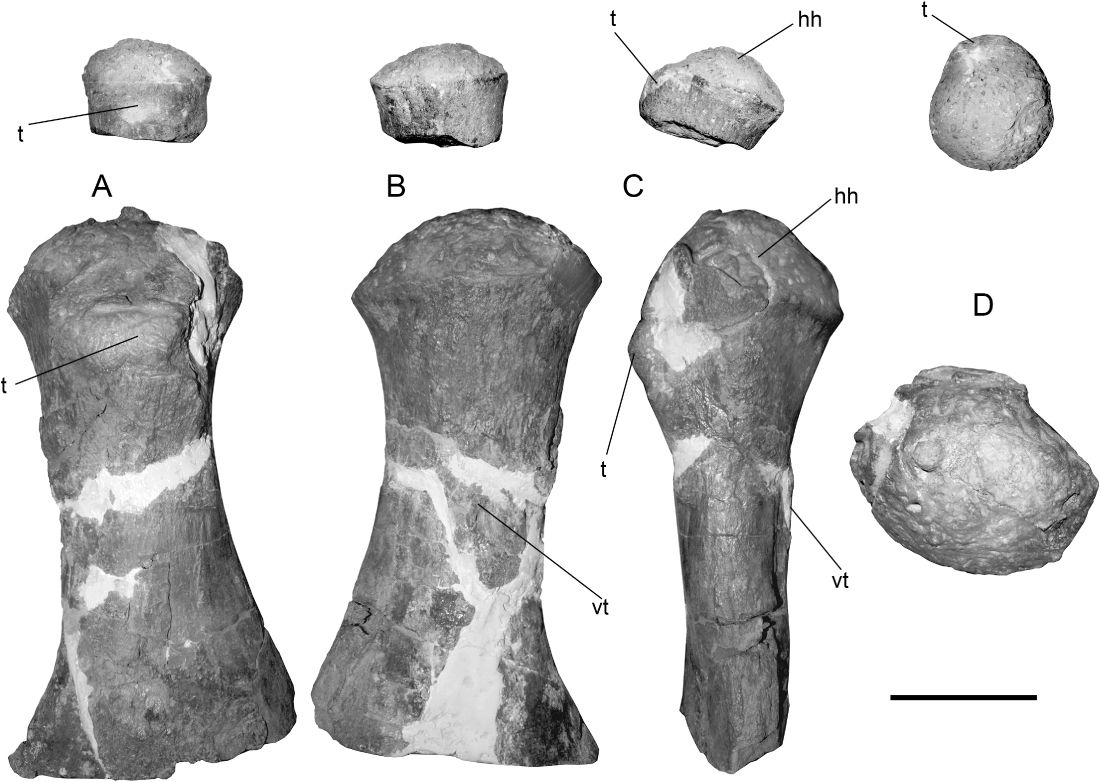
The holotype of A. quiriquinensis does not preserve the proximal portion of its only known femur (). On the other hand, the referred juvenile specimen SGO.PV.260 has incomplete humeri lacking part of the diaphysis while the femur, although fractured, is almost complete but preserved in two separate blocks. The articular heads of the humeri and the femur are convex, with the tuberosity/trochanter clearly differentiated from the articular head as well as from the diaphysis.
SGO.PV.135 and SGO.PV.169 (previously referred to Mauisaurus sp.)
Direct comparison of SGO.PV.135 with the femur of the holotype SGO.PV.957 of A. quiriquinensis shows a perfect match of the distal portion, both in morphology and size (). The distinctive articular facets and posterodistal projection are present in both specimens. Also, the breadth of the shaft (at least the portion preserved in SGO.PV.957) is identical. Based on this, we identify SGO.PV.135 as a femur of an adult A. quiriquinensis. These two femora differ from the specimen DM R1529, the lectotype of M. haasti (), only sharing the presence of the hemispherical femoral head.
Kaiwhekea katiki (OU 12649)
A direct review of the holotype of this species (OU 12649) shows the presence of femora with hemispherical heads (). This feature is clearly observed in the mould of the proximal portion on the better-preserved femur. As illustrated by Cruickshank & Fordyce (Citation2002; ), the femur of K. katiki has a trochanter clearly aligned diagonally to the shaft axis. The distal end of the femur is comparatively narrower than those of A. quiriquinensis specimens.
Comparisons
SGO.PV.169 was directly compared with the left humerus of the holotype (SGO.PV.957) of A. quiriquinensis. Both match in size, as well as in the hemispherical outline of the articular head (). Further, the putative ‘tuberosity’ is displaced from the midline into the posterior half but does not extend diagonally over the shaft, which is one of the features here considered useful for distinguishing the humerus from the femur of A. quiriquinensis (see the ‘Femur’ section). In consequence, due to the morphological affinities, similar ontogenetic stages and because of the precise match in size, SGO.PV.169 is now identified as a humerus of the latter species. Interestingly, the humerus of A. quiriquinensis has a distal portion which is very similar to that of the humerus of K. katiki (). Both taxa have concave facets for the epipodials and rounded distal margins surround the epipodial articulations. They also share the presence of radius and ulna axially longer than broad, with a lunate shape.
Morphological disparities between the adult humeri and femora of Aristonectes quiriquinensis with comments regarding other aristonectines
Among elasmosaurid specimens recovered from the WBP, isolated propodials are common. This could be a consequence of the compactness of the bone, which gives it good resistance to the burial conditions; also, paddles are often found displaying bite marks and could reflect the loss of the extremities resulting from the action of predators. Independent of the causes for isolated propodials, it is relevant to recognise distinctive morphologies that could allow identification of their anatomical position. Further, knowing the ontogenetic stage is relevant for understanding how the general propodial shape changes during growth. Finally, recognition of morphological features with taxonomic value could lead to identification of propodials, even in isolated occurrences.
Based on the complete left humerus and the partially preserved femur of the holotype SGO.PV.957 of A. quiriquinensis, we can recognise some features useful in differentiating humerus from femur even when comparing fragmentary specimens.
Shaft features
In dorsal view, the humerus has a straight shaft, which is comparatively thicker than that of the femur SGO.PV.135. This confers to the humerus a more robust aspect.
Tuberosity/Trochanter shape and position
In the humerus, the tuberosity is slightly displaced from the midline into the post-axial half and is subrectangular to square, whereas in the femur of K. katiki and specimen MLP 89-III-3-1 referred toAristonectes sp., the trochanter is more elongated, extending into the mid portion and is diagonally oriented. The tuberosity of SGO.PV.957 has the same relative position on the humerus as in the juvenile SGO.PV.260, indicating that this is the actual position and not a taphonomic artefact. On the other hand, the femur SGO.PV.135 has a trochanter extending diagonally from the proximal portion to the postaxial, distal margin. Similarly, the trochanter of K. katiki and that of MLP 89-III-3-1 both extend diagonally in the same direction, but the trochanter of SGO.PV.957 is comparatively shorter and constrained to the proximal third, while in K. katiki this extends closer to the middle of the shaft. As a result, the femur can be distinguished by the presence of a larger tuberosity placed diagonally and fading into the postaxial margin of the mid part of the diaphysis ().
Distal end of the propodials
Another relevant difference is observed on the distal articular facets of the propodials of A. quiriquinensis. The humerus has a facet for the ulna with a small projection that covers the postero-proximal margin of the ulna, while in the femur only a posterior projection occurs, leaving the posterodistal margin of the femur open over the fibula.
Size
Finally, the comparative size of the articular heads is different, being larger in the humerus. This is consistent with the typical larger size of this element with respect to the femur among elasmosaurids (O’Keefe Citation2001).
Propodial ontogeny in Aristonectes quiriquinensis
Humerus
Since the right humerus of SGO.PV.957 has good tridimensional preservation, we chose it for comparison, in different views, with the right humerus of the juvenile SGO.PV.260. In dorsal view (), the latter bears an incipient tuberosity which is low but separated from the articular humeral head. In the adult, SGO.PV.957, the tuberosity becomes large and elevated, although no proper fossa between this and the humeral head is present. In ventral view (), it can be noted than even in juvenile stages the humerus already has a marked limit between the diaphysis and the articular surface, which is slightly more prominent than the diaphysis. In comparison, the adult bears a strongly convex hemispherical head that is markedly wider than the diameter of the diaphysis. In anterior view, both specimens (juvenile and adult) have a humeral head with rounded articular outline (). The proximal view () shows that the articular head of the juvenile has a rounded contour with the dorsal tuberosity slightly displaced from the midline towards the posterior half. In the adult specimen SGO.PV.957, the distal portion of the right humerus is slightly crushed; the left humerus is more deformed, hence it was not considered for comparison here. Despite this, the right adult humerus shows a slight posterior displacement of the tuberosity, but the rounded outline is hard to evaluate due to the crushing.
Femur
The femur and humerus of the juvenile SGO.PV.260 () are very similar. In ventral view, the juvenile femoral head is comparatively more prominent than the respective humeral head, a condition that is reversed in the adult stage as is revealed by the new referred specimens SGO.PV.135 and SGO.PV.169, where the humerus acquires an articular head that is comparatively more prominent than that of the adult femur (SGO.PV.135). The distal portion of the juvenile bears a posterior extension for the articular facet of the fibula, which is also present in the adult. In cross-section, the bone shows a distinctive outline with the dorsal surface convex and the ventral surface even more convex, and having its pre-axial margin narrower than the rest of the diaphysis section (). The distal enlargement of the fibular facet, as well as the singular cross-section outline, is present in the available portion of the femur of the adult holotype of A. quiriquinensis (). Due to the remarkable match in morphology and size, the femur of SGO.PV.957 and the femur of SGO.PV.135 are here considered to belong to the same species. The adult femur of A. quiriquinensis therefore has a hemispherical head, a high trochanter not extending far diagonally over the shaft, a deep fossa between the latter and the femoral head and a strong and high tuberosity placed in the mid ventral portion of the diaphysis.
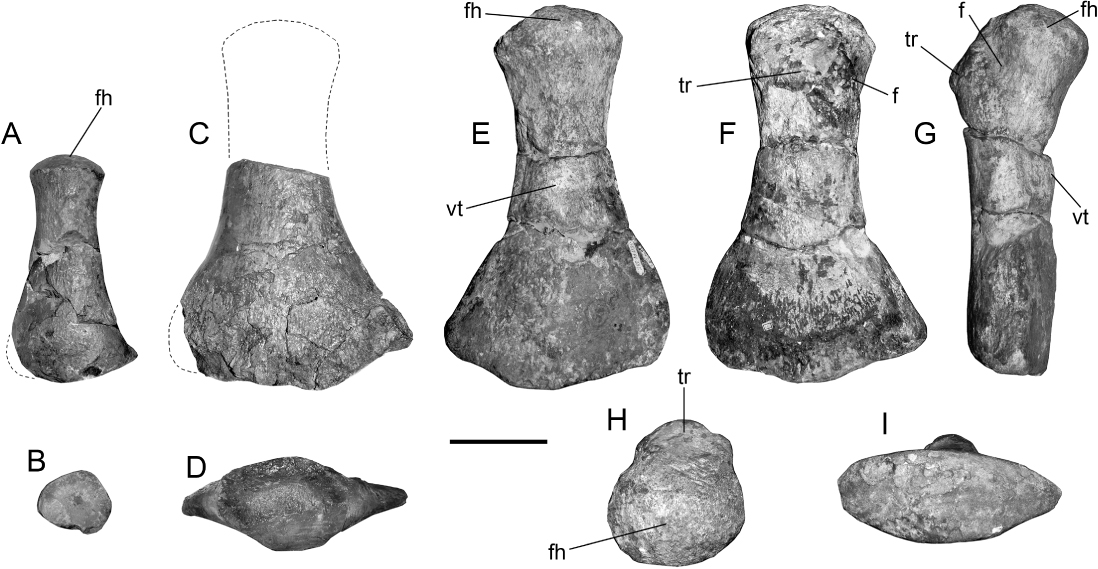
Since SGO.PV.135 has excellent tridimensional preservation, we can add to the description of the femur of A. quiriquinensis the following features: presence of a hemispherical articular head with rounded contour in proximal view and a trochanter shorter than that of K. katiki, with an axial length shorter than one-third of the entire bone (). Also, the distal view of the femur () shows that this has a biconvex outline, with the dorsal surface comparatively flatter than the ventral surface. Based on specimens SGO.PV.260 and SGO.PV.957, the presence of hemispherical heads in the humeri and femora is a feature that depends on the ontogenetic stage of the specimen, being more evident in adult representatives and probably more prominent in even older individuals. In addition, the degree of prominence of the articular head with respect to the shaft when comparing humeri and femora can be variable during ontogeny. During the juvenile stage, SGO.PV.260 has a femoral head comparatively more prominent than its respective humerus; in the adult SGO.PV.957, the humerus becomes comparatively more prominent than the femur. This condition is unusual since the MLP 89-III-3-1 (Aristonectes sp.) possesses a hemispherical femoral head more prominent than that of the humerus.
Anatomical comments on Kaiwhekea katiki
A final observation concerns the laterality of the best-preserved hind limb of K. katiki. The quarry diagram with the original description (Cruickshank & Fordyce Citation2002; ) shows a well-preserved extremity settled in the right side of the skeleton, which was naturally interpreted as the right hind limb, while a putative fragment of the left femur lay over to the left side of the skeleton. The diagram of the femur (Cruickshank & Fordyce Citation2002; ) shows a trochanter diagonally oriented over the shaft and fading posteriorly. Such features are only visible in dorsal view while the trochanter fading occurs towards the posterior margin of the shaft of all the specimens reviewed here. Consequently, the drawing represents a left femur. Since the putative right hind limb is partially preserved as a negative cast, a positive mould obtained and kindly provided by RE Fordyce (May, 2013) was useful for evaluating the anatomy of this portion. Indeed, this reflects the same laterality of the original drawing (Cruickshank & Fordyce Citation2002; ) and positively belongs to a left femur based on the shape of the hemispherical head and the disposition of the trochanter, here shown to occur in posterior situation among aristonectines. The left hind limb must therefore have been displaced prior to burial; soft tissues were still preserved, allowing its transportation from the acetabulum but without suffering disarticulation. In consequence, the fragmentary femur observed in the specimen belongs to the right hind limb.
Conclusions
The results of this contribution prove that the hemispherical head is a feature that lacks enough diagnostic value for genus-level determinations. The description by Otero et al. (Citation2010) noted that the Chilean specimens were larger than the lectotype of M. haasti (Welles Citation1962; Welles & Gregg Citation1971) and, in the case of SGO.PV.169, the articular head was even bigger and more prominent, which led to a first explanation that this latter specimen could belong to an older individual or else to a different species. As explained here, direct comparison of SGO.PV.135 and SGO.PV.169 with the propodials of A. quiriquinensis (holotype, SGO.PV.957) revealed a precise match with their femora and humeri, respectively. Based on these observations, we propose a taxonomic emendation for the Chilean specimens (SGO.PV.135 and SGO.PV.169) from the upper Maastrichtian Quiriquina Formation, previously referred to Mauisaurus sp., now considered as referred specimens of A. quiriquinensis.
Since its first description, the historical taxon Mauisaurus haasti Hector, 1874, from the late Campanian of New Zealand, has been regarded as unique among plesiosaurs due to the presence of a prominent hemispherical head in the femur, which was implicitly considered as an autapomorphy of the species. The very large adult specimen MLP 83-III-3-1 from the late Maastrichtian of Antarctica and referred to Aristonectes sp. (O’Gorman Citation2013), as well as the holotype SGO.PV.957 and referred specimen SGO.PV.135 of A. quiriquinensis, both from the late Maastrichtian of central Chile, demonstrate the presence of this femoral feature in different specimens within the genus Aristonectes. The hemispherical femoral head is here found to be also present in the holotype of K. katiki from the early Maastrichtian of New Zealand. The ontogenetic stage of K. katiki, A. quiriquinensis and DM R1529 allow the comparison of their adult femoral morphologies, showing clear disparities in shape, size and distribution of the relevant anatomical features. Furthermore, the respective chronostratigraphic occurrences of the three specimens, as well as their morphologic differences (especially in the known skulls) prove that all belong to different taxa, therefore precluding disparities due to sexual dimorphism.
Although a hemispherical femoral head can be commonly found among aristonectines, such a feature is here proved to be non-diagnostic to genus or species level. Hence, this feature must be rejected as an autapomophy of M. haasti. Furthermore, the hemispherical head of the femur was described in another specimen from New Zealand, CM Zfr 115 (Hiller et al. Citation2005), which clearly belongs to a non-aristonectine elasmosaurid. This implies that the hemispherical femoral head is present in different taxa that are not part of a monophyletic lineage and, in consequence, such a feature should be treated as a homoplasic character.
The two propodials from the upper Maastrichtian of central Chile previously referred to Mauisaurus sp. (SGO.PV.135 and SGO.PV.169) are now identified as a proximal portion of a left humerus and a complete left femur, respectively. Both are here referred to Aristonectes quiriquinensis based on the precise morphological match and adult size of the compared specimens. This allows rejection of the presence of the genus Mauisaurus in the southeastern Pacific, at least on the basis of isolated propodials. Based on the Chilean specimens here studied, such a feature is proved to depend on the ontogeny of the individual and becomes more evident in adult and very mature specimens.
As an additional result of the propodial comparisons carried out here, we can positively identify the left hind limb of K. katiki (originally regarded as the right hind limb based on its placement) which lies over the right side of the skeleton and is exposed in ventral view. The results of this research encourage reassessment of the taxonomic affinities of the specimens from Antarctica, Argentina and New Zealand which have been previously referred to Mauisaurus.
Associate Editor: Associate Professor Kathy Campbell.
Acknowledgements
RAO was supported by the Antarctic Ring Project (Anillos de Ciencia Antártica ACT-105, Conicyt-Chile) and by the Domeyko II UR-C12/1 grant Red Paleontológica U-Chile of the University of Chile. Dr RE Fordyce (University of Otago, New Zealand), Dr P Scofield and Dr C Vink (Canterbury Museum, Christchurch, New Zealand) are especially acknowledged for granting access to the New Zealand specimens reviewed here. JPO’G was supported by projects PICT 2008-0261, PICT 2012-0748, PICTO 2010-0093, PIP 433, UNLP N 677 and UNLP N607. Some materials (MLP 89-III-3-1) were collected with the logistic support from the Instituto Antártico Argentino (Buenos Aires), as well as from the Fuerza Aérea Argentina. JD Stillwell (Monash University, Australia) and a second anonymous reviewer are acknowledged for the review of the manuscript and comments that helped to improve it.
Disclosure statement
No potential conflict of interest was reported by the authors.
Related Research Data
References
- Benson RBJ, Druckenmiller PS 2014. Faunal turnover of marine tetrapods during the Jurassic–Cretaceous transition. Biological Reviews 89: 1–23. [doi:10.1111/brv.12038]
- Biró-Bagóczky L 1982. Revisión y redefinición de los ‘Estratos de Quiriquina’, Campaniano–Maastrichtiano, en su localidad tipo, en la Isla Quiriquina, 36°37′ Lat. Sur, Chile, Sudamérica, con un perfil complementario en Cocholgüe. Congreso Geológico Chileno No 3 (Concepción). Actas 1: A29–A64.
- de Blainville HMD 1835. Description de quelques espèces de reptiles de la Californie précédé de l’analyse d’un système général d’erpétologie et d’amphibiologie. Nouvelles Annales du Muséum d’Histoire Naturelle de Paris, Série 3: 233–296.
- Cabrera A 1941. Un Plesiosaurio nuevo del Cretáceo del Chubut. Revista del Museo de La Plata 2: 113–130.
- Chatterjee S, Creisler BS 1994. Alwalkeria (Theropoda) and Morturneria (Plesiosauria), new names for preoccupied Walkeria Chatterjee, 1987 and Turneria Chatterjee and Small, 1989. Journal of Vertebrate Paleontology 14: 142–142.
- Chatterjee S, Small BJ 1989. New plesiosaurs from the Upper Cretaceous of Antarctica. In: Crame JM ed. Origins and evolution of the Antarctic Biota. Geological Society of London Special Publication 47: 197–215.
- Consoli CP, Stilwell JD 2009. Late Cretaceous marine reptiles (Elasmosauridae and Mosasauridae) of the Chatham Islands, New Zealand. Cretaceous Research 30: 991‒999. [doi:10.1016/j.cretres.2009.02.009]
- Cope ED 1869. On the reptilian orders, Phytonomorpha and Streptosauria. Proceedings of the Boston Society of Natural History 12: 250–266.
- Crame JA, Francis JE, Cantrill DJ, Pirrie D 2004. Maastrichtian stratigraphy of Antarctica. Cretaceous Research 25: 411–423. [doi:10.1016/j.cretres.2004.02.002]
- Cruickshank ARI, Fordyce RE 2002. A new marine reptile (Sauropterygia) from New Zealand: further evidence for a Late Cretaceous austral radiation of cryptocleidid plesiosaurs. Palaeontology 45: 557–575. [doi:10.1111/1475-4983.00249]
- Fostowicz-Frelik L, Gaździcki A 2001. Anatomy and histology of plesiosaur bones from the Late Cretaceous of Seymour Island, Antarctic Peninsula. In: Gaździcki A ed. Palaeontological results of the Polish Antarctic Expeditions. Part III. Palaeontologia Polonica 60. Warsaw, Institute of Paleobiology of the Polish Academy of Sciences. Pp. 7–32.
- Gasparini Z, Bardet N, Martin JE, Fernandez M 2003. The elasmosaurid plesiosaur Aristonectes Cabrera from the latest Cretaceous of South America and Antarctica. Journal of Vertebrate Paleontology 23: 104–115. [doi:10.1671/0272-4634(2003)23[104:TEPACF]2.0.CO;2]
- Gasparini Z, Goñi R 1985. Los plesiosaurios cretácicos de América del Sur y del continente antártico. Congresso Brasileiro de Paleontología, Number 8 (Río de Janeiro), Coletanea de Trabalhos Paleontológicos, Serie Geologie 27: 55–63.
- Gasparini Z, Salgado L, Parras A 2007. Late Cretaceous plesiosaurs from northern Patagonia, Argentina. Geological Journal 42: 185–202. [doi:10.1002/gj.1079]
- Hector J 1874. On the fossil reptiles of New Zealand. Transactions of the New Zealand Institute 6: 333–358.
- Hiller N, Mannering AA, Jones CM, Cruickshank ARI. 2005. The nature of Mauisaurus haasti Hector, 1874 (Reptilia: Plesiosauria). Journal of Vertebrate Paleontology 25: 588–601. [doi:10.1671/0272-4634(2005)025[0588:TNOMHH]2.0.CO;2]
- Hiller N, O’Gorman JP, Otero RA 2014. A new elasmosaurid plesiosaur from the lower Maastrichtian of North Canterbury, New Zealand. Cretaceous Research 50: 27–37. [doi:10.1016/j.cretres.2014.03.026]
- Martin J, Sawyer F, Reguero M, Case J 2007. Occurrence of a young elasmosaurid plesiosaur skeleton from the Late Cretaceous (Maastrichtian) of Antarctica. In: Cooper AK, Raymond CR et al. eds. Antarctica: a keystone in a changing world. Online Proceedings of the 10th ISAES, USGS Open-File Report 2007-1047, Short Research Paper 66. August 26 to September 1, Santa Barbara, CA, US Geological Survey. Pp. 1–4.
- Novas FE, Isasi MP, D’Angelo JS, Busker F, Damato M, Bogan S et al. 2011. Hallazgo de un esqueleto de plesiosaurio asociado con dientes de selaceos en la Formación El Calafate (Cretácico superior) de la Provincia de Santa Cruz. Libro de Resúmenes IV Congreso Latinoamericano de Paleontología de Vertebrados, San Juan, Argentina. Ameghiniana 48: R247.
- O’Keefe FR 2001. A cladistic analysis and taxonomic revision of the Plesiosauria (Reptilia: Sauropterygia). Acta Zoologica Fennica 213: 1–63.
- O’Keefe FR, Street, HP 2009. Osteology of the cryptocleidoid plesiosaur Tatenectes laramiensis, with comments on the taxonomic status of the Cimoliasauridae. Journal of Vertebrate Paleontology 29: 48–57. [doi:10.1671/039.029.0118]
- O’Gorman JP 2012. The oldest elasmosaurs (Sauropterygia, Plesiosauria) from Antarctica, Santa Marta Formation (upper Coniacian? Santonian–upper Campanian) and Snow Hill Island Formation (upper Campanian–lower Maastrichtian), James Ross Island. Polar Research 31: 1‒10.
- O’Gorman JP 2013. Plesiosaurios del Cretácico Superior de Patagonia y Península Antártica, Tomos I y II. Unpublished PhD dissertation, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, Argentina. 527 p.
- O’Gorman JP, Gasparini Z, Salgado L 2014. Reappraisal of Tuarangisaurus? cabazai (Elasmosauridae, Plesiosauria) from the Upper Maastrichtian of northern Patagonia, Argentina. Cretaceous Research 47: 39‒47.
- Otero RA, Soto-Acuña S, Rubilar-Rogers D 2010. Presence of Mauisaurus Hector (Plesiosauroidea, Elasmosauridae) in the Late Cretaceous (Maastrichtian) of central Chile. Brief Report, Acta Paleontologica Polonica 55: 361–364.
- Otero RA, Soto-Acuña S, Rubilar-Rogers D 2012. A postcranial skeleton of an elasmosaurid plesiosaur from the Maastrichtian of central Chile, with comments on the affinities of Late Cretaceous plesiosauroids from the Weddellian Biogeographic Province. Cretaceous Research 37: 89–99. [doi:10.1016/j.cretres.2012.03.010]
- Otero RA, Soto-Acuña S, O’Keefe FR, O’Gorman JP, Stinnesbeck W, Suárez ME et al. 2014a. Aristonectes quiriquinensis sp. nov., a new highly derived elasmosaurid from the upper Maastrichtian of central Chile. Journal of Vertebrate Paleontology 34: 100‒125. [doi:10.1080/02724634.2013.780953]
- Otero RA, Soto-Acuña S, Vargas AO, Rubilar-Rogers D, Yury-Yáñez R, Gutstein CS 2014b. Additions to the diversity of elasmosaurid plesiosaurs from the Upper Cretaceous of Antarctica. Gondwana Research 26: 772–784. [doi:10.1016/j.gr.2013.07.016]
- Owen R. 1861. On the remains of a plesiosaurian reptiles (Plesiosaurus australis) from the Oolitic formation in the middle island of New Zealand. The Geologist 4: 444‒445.
- Rinaldi CA, Massabie A, Morelli J, Rosenman HL, del Valle R 1978. Geología de la Isla Vicecomodoro Marambio. Contribución del Instituto Antártico Argentino 217: 1–37.
- Roberts EM, Lamanna MC, Clarke JA, Meng J, Gorscak E, Sertich JJW et al. 2014. Stratigraphy and vertebrate paleoecology of Upper Cretaceous–?lowest Paleogene strata on Vega Island, Antarctica. Palaeogeography, Palaeoclimatology, Palaeoecology 402: 55–72. [doi:10.1016/j.palaeo.2014.03.005]
- Salazar C, Stinnesbeck W, Quinzio-Sinn LA 2010. Ammonites from the Maastrichtian (Upper Cretaceous) Quiriquina Formation in central Chile. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen 257: 181–236.
- Welles SP 1943. Elasmosaurid plesiosaurs with description of new material from California and Colorado. Memoirs of the University of California 13: 125–254.
- Welles SP 1962. A new species of elasmosaur from the Aptian of Colombia and a review of the Cretaceous plesiosaurs. University of California, Publications in Geological Sciences 44: 1–96.
- Welles SP, Gregg DR 1971. Late Cretaceous marine reptiles of New Zealand. Records of the Canterbury Museum 9: 1–111.
- Wilson GJ, Schiøler P, Hiller N, Jones CM 2005. Age and provenance of Cretaceous marine reptiles from the South Island and Chatham Islands, New Zealand. New Zealand Journal of Geology and Geophysics 48: 377–387. [doi:10.1080/00288306.2005.9515120]
- Zinsmeister WJ 1979. Biogeographic significance of the Late Mesozoic and early Tertiary molluscan faunas of Marambio Island (Antarctic Peninsula) to the final break-up of Gondwanaland. In: Gray J, Boucot AJ eds. Historical biogeography, plate tectonics and the changing environment. Corvallis, Oregon State University Press. Pp. 349–355.