
ABSTRACT
An early Miocene dolphin (ZMT 73: partial skull and ear bones) from Kaikoura, New Zealand, is the sister taxon to the previously named Early Miocene Papahu taitapu. The new specimen, although insufficient to justify a new species name, includes an informative squamosal and periotic that help move the Papahu clade from the stem Odontoceti to either the Platanistoidea, or to become the sister taxon to the Ziphiidae+Eurhinodelphinidae+Delphinida. The fossils usefully add to the taxonomic and structural diversity of Cetacea during the important but poorly known Early Miocene.
Introduction
The fossil history of dolphins and whales (Cetacea) starts in the Ypresian, 50–53 Ma. Early cetaceans were subtropical during the Ypresian and Lutetian (Gingerich Citation2015), but in the Bartonian spread to temperate latitudes as far south as New Zealand (Köhler & Fordyce Citation1997). The later global stratigraphic record for cetaceans is generally well-known and fairly continuous (for example, see: fossilworks.org), except for three problematic intervals: Rupelian (Early Oligocene), Aquitanian (Early Miocene) and Gelasian–Ionian (Pleistocene). In each case, fossils from the adjacent stages, and/or phylogenetic reconstructions, imply that cetacean lineages are under-represented by fossils (e.g. Marx & Fordyce Citation2015). The Rupelian low diversity has been explained by the erosion of bone-bearing shelf strata when the sea level fell in the middle of the Oligocene (Fordyce Citation2003). Similarly, sea level fluctuations and repeated erosion of bone-bearing shelf strata might account for the patchy Pleistocene record. Pleistocene fossil Cetacea might also be hard to date because of the lack of marine pelagic biostratigraphic zonal species. The Aquitanian is more problematic, because the Early Miocene has been mentioned widely, albeit with stage often not specified, as representing the rise of early ‘modern’ cetaceans. Notable assemblages () include those from the ‘Patagonian marine formation’ of Argentina (Cozzuol Citation1996), from the Jewett Sand and Vaqueros Formation of California (Barnes Citation1977) and the Libano Sandstone of Belluno, northern Italy (Pilleri Citation1985; Bianucci & Landini Citation2002). However, the Patagonian cetaceans, which are now ascribed formally to the Gaiman Formation, appear to be no older than Burdigalian (Cione et al. Citation2011). The Belluno cetaceans may be upper Aquitanian or lower Burdigalian, but are not dated by calcareous pelagic microfossils, and the grade of cetaceans has been suggested to indicate Aquitanian age (Bianucci & Landini Citation2002). The Jewett–Vaqueros cetaceans are reportedly Arikareean (Land Mammal Age, equivalent to later Rupelian to Aquitanian) (Barnes & Reynolds Citation2009), and possibly Aquitanian, but occurrences lack datable calcareous pelagic microfossils. In summary, although Aquitanian ages are possible, they have often not been demonstrated.
Figure 1. Stratigraphy of cetacean-bearing units relevant to fossil dolphin ZMT 73. For international occurrences, only the key Early Miocene formations are indicated; see text for details. For New Zealand, the key zonal species of foraminifera for Oligocene to Early Miocene stages are based on Hornibrook et al. (Citation1989) and references discussed in the text. Schematic lithostratigraphy for the southern Canterbury Basin is based on Hornibrook et al. (Citation1989), modified by Fordyce’s field observations of vertebrate-bearing strata. The selected species of Odontoceti are shown in approximate age for the Canterbury Basin formations, although ZMT 73 and Papahu taitapu are from other sequences, as discussed in the text.
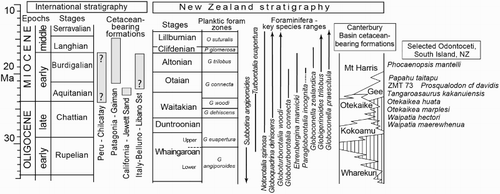
To consider the stratigraphy of New Zealand Early Miocene Cetacea (), the Aquitanian spans the upper Waitakian Stage to about the middle Otaian Stage, while the Burdigalian spans middle Otaian to the top of the Altonian (Raine et al. Citation2012, Citation2015). Morgans et al. (Citation1999, Citation2002) reviewed the key zonal planktic foraminifera (see also Jenkins Citation1965; Hornibrook et al. Citation1989); selected species are given in . Such foraminifera often provide zonal dates for cetacean matrix. However, there is no middle Otaian planktic foraminiferal datum that is a demonstrated proxy for the Aquitanian–Burdigalian boundary in New Zealand. Hornibrook et al. (Citation1989) noted that Globorotalia kugleri does not range to the top Otaian and, elsewhere, the species is used as a zonal indicator (Paragloborotalia kugleri, of Wade et al. Citation2011), but Morgans et al. (Citation1999) questioned most of the New Zealand reports for the species. In the southern Canterbury Basin, Aquitanian cetacean-bearing units include the uppermost Otekaike Limestone, and the lower part of the overlying Mount Harris Formation, sometimes with intervening Gee Greensand. These represent the lower part of the Globoturborotalita connecta zone (). The upper part of that zone overlaps with the lowest occurrences of Paragloborotalia incognita (see Morgans et al. Citation1999), near the base of the Burdigalian.
Recently described fossil dolphins from New Zealand’s Waitakian, Otaian and Altonian stages add to the structural and taxonomic diversity of the Odontoceti (toothed whales and dolphins) (Fordyce Citation1994; Aguirre-Fernández & Fordyce Citation2014; Tanaka & Fordyce Citation2014, Citation2015a, Citation2015b). One species, the Early Miocene Papahu taitapu Aguirre-Fernández and Fordyce (Citation2014), includes a skull, periotic, mandible and a few postcranial bones from northwest Nelson. All parts are damaged, but the dolphin is complete enough to form the basis for a new genus and to allow detailed phylogenetic analysis. Importantly, this fossil is probably Otaian (based on Globoturborotalita connecta; see Aguirre-Fernández & Fordyce Citation2014: figure 1), in the range middle Aquitanian to middle Burdigalian, 23–19 Ma, making Papahu one of few odontocetes world-wide of apparent earliest Miocene age. In spite of the dearth of specimens, the Aquitanian–Burdigalian was important in the rise of modern pelagic dolphins (Aguirre-Fernández & Fordyce Citation2014). Aguirre-Fernández and Fordyce (Citation2014) placed P. taitapu among a paraphyletic cluster of Oligocene to Early Miocene stem odontocetes such as Waipatia and Prosqualodon, and basal to the Ziphiidae, Physeteridae, Platanista and Delphinida. Our later analyses of the relationships of the dolphin Otekaikea moved Papahu to become the sister taxon to the crown-Odontoceti (Tanaka & Fordyce Citation2015a: figure 21, implied weights analysis).
The purpose of this article is to further reassess the phylogenetic position of P. taitapu using the hitherto undescribed specimen ZMT 73 (Canterbury Museum): a partial skull, teeth, tympanic bulla and periotic of one individual. ZMT 73 includes a better preserved squamosal and periotic than in P. taitapu, and also preserves the tympanic bulla and single-rooted teeth. In our preferred implied weights analysis using molecular constraints, the new specimen and P. taitapu are sister species. Their clade is the sister clade of beaked whales + eurhinodelphinids + Delphinida. ZMT 73 is, however, too incomplete to warrant naming as a new species.
Acronyms
AMNH: American Museum of Natural History, New York, USA
IRSNB: Institut Royal des Sciences Naturelles de Belgique, Brussels, Belgium
MNHN: Muséum National d’Histoire Naturelle, Paris, France
MUSM: Museo de Historia Natural, Universidad Nacional Mayor de San Marco, Lima, Peru
OU: Geology Museum, University of Otago, Dunedin, New Zealand
PGN: Paleontological collection of Montpellier University, France
USNM: Department of Paleobiology, National Museum of Natural History, Smithsonian Institution, Washington, DC, USA
ZMT: Fossil mammals catalogue, Canterbury Museum, Christchurch, New Zealand
Material and methods
Material was prepared in 1976 by R. Ewan Fordyce using a pneumatic scribe and immersion of blocks in dilute (4–5%) acetic acid. Yoshihiro Tanaka finished preparation in 2012–2014 using a microjack pneumatic scribe under a Zeiss SR binocular microscope, supplemented with localised use of dilute acetic acid (4%). The positions of most sutures were confirmed using the binocular microscope. Bone was glued with cyanoacrylate and/or polyvinyl butyral solution, the latter also used as a consolidant. Photographs were taken with a Nikon D700 DSLR camera and a 105 mm micro lens. Most views show the specimen coated with sublimed ammonium chloride, with lighting from the upper left.
The initial ‘immersion’ acid preparation in 1976 freed bone from matrix, but also seriously weakened bone along pre-existing fine (structural) joints. That treatment led to continuing later breakage, particularly of the naturally dense ear bones including the malleus and incus, in spite of consolidation with polyvinyl butyral in 3:1 acetone/ethanol solution. For such structurally affected specimens, we now maximise physical preparation with pneumatic scribes, with localised use of dilute acetic acid for final preparation.
Systematic palaeontology
Cetacea Brisson, 1762
Neoceti Fordyce & de Muizon, 2001
Odontoceti Flower, 1867
Papahu Aguirre-Fernández & Fordyce, Citation2014
cf. Papahu sp.
(– and )
Figure 3. Skull materials of ZMT 73. A, Dorsal view. B, Ventral view (see for detailed basicranium). C, Left lateral view. D, Posterior view.
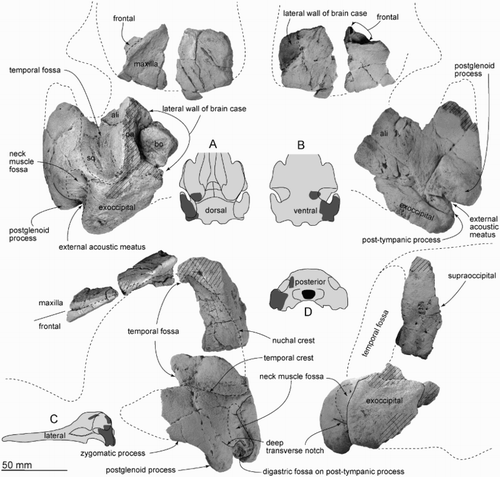
Figure 4. Details of partial left basicranium of ZMT 73. Ventral view.
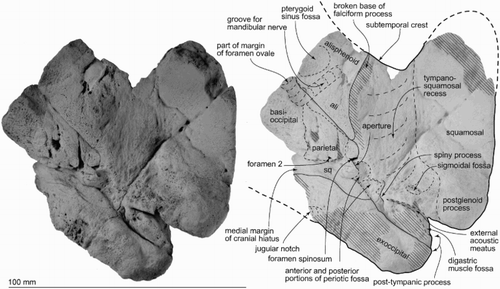
Figure 5. Teeth of ZMT 73. A–D, One complete tooth. E–G, Three crowns lacking roots. H–J, Crown with a denticle. A, Anterior view. B, Posterior view. C, Buccal view. D, Lingual view. E–I, Buccal or lingual views. J, Distal (apical) view.

Figure 6. Right periotic of ZMT 73. A, Ventral view. B, Dorsomedial view showing foramina in the internal acoustic meatus. C, Lateral view. D, Medial view. E, Posterior view. F, Dorsal view. G, Anterior view.
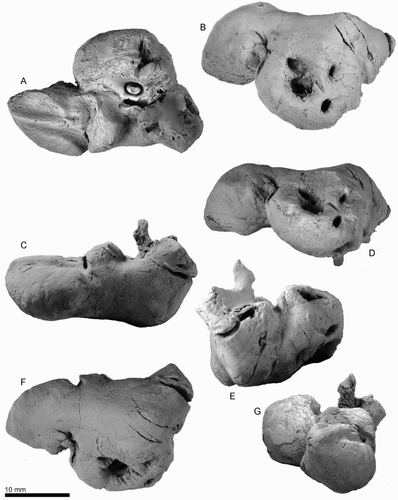
Figure 7. Key features of the right periotic of ZMT 73. A, Ventral view. B, Dorsomedial view showing foramina in the internal acoustic meatus. C, Lateral view. D, Medial view. E, Posterior view. F, Dorsal view. G, Anterior view.
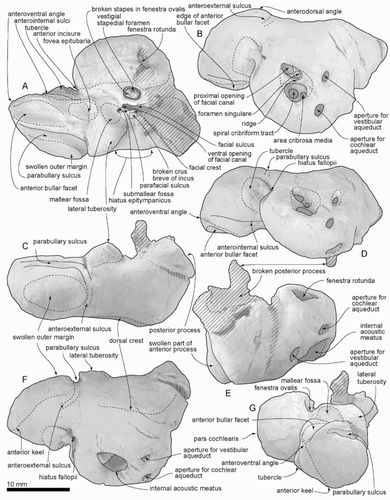
Figure 8. Right bulla of ZMT 73. A, Dorsal view. B, Lateral view. C, Posterior view. D, Medial view. E, Ventral view. F, Anterior view.
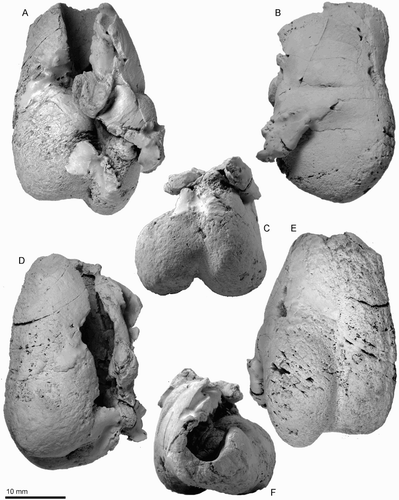
Figure 9. Key features of the right bulla of ZMT 73. A, Dorsal view. B, Lateral view. C, Posterior view. D, Medial view. E, Ventral view. F, Anterior view.
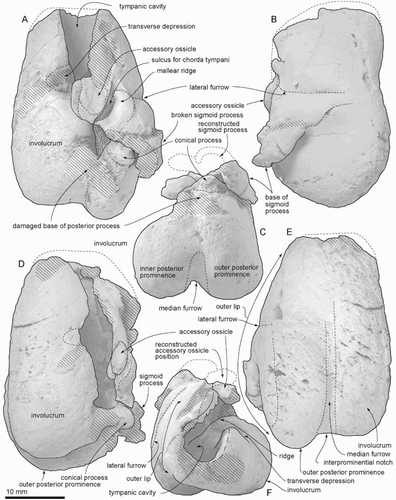
Figure 10. Incus and malleus of ZMT 73. A, Lateral view. B, Dorsal view. C, Anterior view. D, Posterior view. E, Medial view. F, Ventral view.
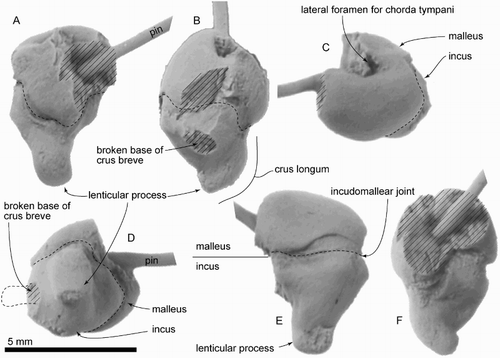
Figure 11. Provisionally identified supraoccipital and rostrum of ZMT 73. A, B, Part of the putative supraoccipital. C–E, Part of the putative rostrum. C, Putative rostral elements in original preserved contact. D, E, Putative rostral elements separated, showing both sides.
Diagnosis
ZMT 73 shares with P. taitapu the following unique combination of periotic features: a V-shaped parabullary sulcus on the periotic, with a swollen outer margin of the anterior process; tapered and short anterior process; narrow anteromedial sulcus; robust lateral tuberosity and large flat space between the internal acoustic meatus and faint dorsal crest. ZMT 73 differs from P. taitapu in a deeper neck muscle fossa (Character 114), more prolonged anteromedially; shallower posterior portion of the periotic fossa (Character 155); abruptly deflected anterior process of the periotic (Character 173); and thinner pars cochlearis (Character 193). Other differences, not included in cladistic analysis, are: the anteroposteriorly thicker exoccipital, shallower fossa of tympano-squamosal recess descending onto the postglenoid process, deeper groove in roof of external acoustic meatus, rhomboidal post-tympanic process of the squamosal, more robust postglenoid process, more widely open squamosal fossa bounded posteriorly and posterolaterally by a more-elevated nuchal crest, and more robust and strongly arched (lateral view) zygomatic process.
Material
Canterbury Museum specimen ZMT 73, an incomplete cranium (left squamosal, exoccipital, parietal and alisphenoid), right bulla, periotic, malleus, incomplete incus, stapes in situ, 18 teeth and presumed fragments of rostrum. These were prepared from associated blocks with limited contacts; no field photos were available to show original positions.
Locality
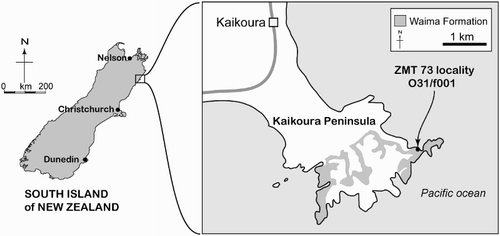
ZMT 73 was collected by I. Mannering (Department of ZMT 73 was collected by I. Mannering (Department of Zoology, University of Canterbury) on 16 January 1966 from an outcrop of Waima Formation on the shore platform, informally known as Seal Reef, toward the eastern end of Kaikoura Peninsula. The site is c. 3 km southeast of central Kaikoura, Marlborough (), and is some 700 m west from the east end of the peninsula (Point Kean). The Geosciences Society of New Zealand fossil record number is O31/f0001, with the coordinates NZMS260 grid (NZGD49) O31:691646, or approximately latitude 42°25′31.3″S, longitude 173°43′04.7″E (Google Earth). Fordyce prospected the locality in 1976, but found no further cetacean fossils.
Figure 2. Locality map and stratigraphy, based on Rattenbury et al. (Citation2006).
Horizon and age
The Cretaceous–Cenozoic sedimentary sequence at Kaikoura Peninsula (Rattenbury et al. Citation2006) includes c. 400 m thickness of Waima Formation on the coast both east and west of the ZMT 73 locality. The exact horizon for ZMT 73 is uncertain, but is presumably toward the base, given the planktic foraminiferal zone discussed below. The lithology of ZMT 73 agrees with published descriptions of the unit as greenish to bluish-grey calcareous silty mudstone (Browne & Field Citation1985; Rattenbury et al. Citation2006). McKay (Citation1887) recorded fragments of cetacean or possibly reptile bone from the ‘grey marls,’ of the Waima Formation, at Kaikoura (Fordyce Citation1980).
The matrix of ZMT 73 is light green–grey massive calcareous glauconitic siltstone. We identified the zonal planktic foraminiferan Globoturborotalita connecta (taxonomy follows Aze et al. Citation2011), in matrix from ZMT 73, indicating the Globoturborotalita connecta planktic foraminiferal zone of Jenkins (Citation1965) (see also Hornibrook et al. Citation1989) (). The latter zone indicates the upper Waitakian to Otaian New Zealand stages, c. 23 to 18.7 Ma, early Aquitanian to middle Burdigalian. Morgans et al. (Citation1999: figure 13) showed the base of the Globoturborotalita connecta zone provisionally at 22.8 Ma. Hornibrook (pers. comm. to RE Fordyce) identified other foraminifera from the ZMT 73 matrix in 1976: Globoturborotalia. woodi woodi, Globoquadrina dehiscens, Globigerinita dissimilis and Globorotalia semivera, consistent with upper Waitakian to Otaian stages.
General description
Morphological terminology follows Mead and Fordyce (Citation2009), unless stated. Surfaces are eroded in places, noted when major. The ventral surfaces of the squamosal and adjacent bones are worn, presumably by bioerosion shortly after death; such wear is consistent with a ventral-up post-mortem position, as is common for fossil Cetacea. The larger parts are dissociated from each other. The tympanoperiotic shows fissures that are probably fine structural joints. No features clearly indicate ontogenetic age, but the few sutures are firm, rather than open, suggesting submature to physically mature. Some of the teeth have pulp cavities not fully infilled.
Maxilla and frontal
Only the posteromedial part of the left ascending process of the maxilla and frontal is preserved; the suture is closed but still evident (). The maxilla shows thick anterolateral and thinner posterior edges, and the medial part of the ascending process. The posterior edge of the ascending process of the maxilla is curved in dorsal view, which implies that the ascending process was semicircular like P. taitapu, and the edges have weak striae. The frontal preserves some of the brain case medially, but reveals no more than a hemispherical endocranial cavity.
Possible rostrum or mandible
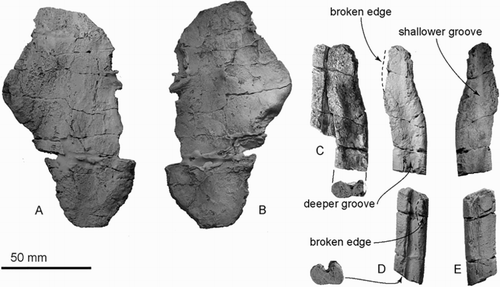
Two elongate fragments of superficially rib-like bones were found together, joined by matrix and preserved in slightly overlapping parallel contact (C–E). Because of their symmetry, these bones are interpreted as possible rostrum or mandible rather than rib. The bones were associated no later than the early stages of burial, as indicated by shared fine structural joints that overprint the two bones (C); the structural joints would have formed later in burial history. The bones suggest an originally long and, together, flat structure, but neither preserves an apparent natural end, and the original association with other cranial parts of ZMT 73 is uncertain. One side is more slender, the other side wider and flatter. Each bone has a compressed oval cross section, with deep and shallow longitudinal grooves on the opposing surfaces. The deeper groove, which is 4 mm deep and an open U‐shape, may be an alveolar groove, identifying the bone as maxilla. One longitudinal side has slightly flatter area, which might be an unfused suture for the vomer. Mandible seems less likely because each bone is dorsoventrally thin. We have not been able to find a match among ribs from other archaic odontocetes. A postcranial position further seems unlikely because of the lack otherwise of pectoral bones, vertebrae and definite ribs.
Parietal
The parietal is preserved in two parts (), which form the lateral wall of the braincase. The parietal/squamosal suture is not evident and is perhaps fused. The parietal/supraoccipital suture is closed but still evident dorsally. The reconstructed temporal fossa is relatively high, and the posterodorsal part is rounded.
Squamosal
The squamosal is robust and well extended laterally. The zygomatic process is broken anteriorly, but the preserved posterior part is wide (46.5 mm at the level of the subtemporal crest) and high (28.3 mm) with a smooth and rounded lateral body; its shape is consistent with an originally short blunt tip. The floor of the temporal fossa is short ventrally, and widely open, between the subtemporal crest and temporal crest at the posterior end of the temporal fossa. (The floor of the fossa was termed squamosal sulcus by Miller Citation1924, and squamosal fossa by Barnes Citation1985.) The glenoid fossa is excavated strongly, markedly arched in lateral view, with a long postglenoid process with a robust and smoothly rounded apex. Slightly dorsal to the postglenoid process, a deep transverse notch runs obliquely forward from the deep narrow slit of the external acoustic meatus. Posterior to the lateral opening of the external acoustic meatus, the rhomboidal post-tympanic process has a prominent, laterally facing fossa, presumably at least partly for the digastric muscle. Dorsal to the external acoustic meatus and the post-tympanic process, on the lateral face of the squamosal, a long open fossa for neck muscles rises dorsally and slightly forward; the posterior border is formed by the exoccipital.
Ventrally (), the zygomatic process has a thin lateral edge just anterior to the glenoid fossa. The latter merges forward and medially into the wide shallow tympano-squamosal recess without a clear boundary. The recess separates the zygomatic process from the falciform process and the periotic fossa, occupying the squamosal from the subtemporal crest to the external acoustic meatus and spiny process. A shallow branch of the tympano-squamosal recess descends onto the postglenoid process. The apex of the spiny process is worn and indistinct. A worn, shallow, mediolaterally long oval fossa between the spiny and postglenoid processes may be the sigmoidal fossa sensu Geisler et al. (Citation2005). The falciform process shows only a broken base, leaving the original proportions uncertain; the base of the falciform process has a firm suture with the alisphenoid. Posterior to the falciform process, shallow anterior and posterior portions of the periotic fossa lie just anterior to the exoccipital. Medial to the anterior portion of the periotic fossa, a small pocket continues to the small foramen spinosum. The external acoustic meatus is a narrow deep slit, rising onto the lateral face of the squamosal. The anterior meatal crest appears to be naturally low and rounded; there is no evidence of a posterior meatal crest, and no obvious suture for the posterior process of the bulla on the post-tympanic process. The posterior portion of the periotic fossa extends laterally, narrowing to end posterior to the rhomboidal post-tympanic process.
Supraoccipital and exoccipital
A large thin (c. 5 mm) plate without clear symmetry (A,B) is, by process of elimination, probably part of the transversely wide braincase roof (B). The element is not part of the scapula, or a facial crest. The flat and rugose left lateral part of the supraoccipital overlies the parietal. Ventral contacts with the exoccipital are lost. The left exoccipital is preserved lateral to the jugular notch; the bone has a closed but visible suture with the squamosal. In ventral view, the exoccipital is anteroposteriorly thick (18.5 mm) and extends laterally at least to the limit of the adjacent post-tympanic process of the squamosal. Posteromedially, the rounded ventral surface could be the lateral wall of the jugular notch.
Alisphenoid
The alisphenoid forms the posteromedial margin of the subtemporal crest. On the basicranium, the alisphenoid is thick with firm sutures with the squamosal laterally and posterolaterally. Medially, there is a shallow subcircular pterygoid sinus fossa. A shallow groove for the mandibular nerve runs forward (partly consolidated with a plug of plaster to stablise fractured bone), between the pterygoid sinus fossa and the base of the falciform process of the squamosal.
Teeth
ZMT 73 preserves one complete tooth and at least 17 incomplete teeth (A–D and E–I show examples). All the preserved teeth include a variably recurved conical cusp with blunt anterior and posterior keels. One crown (H, I) shows a tiny nodular vestigial denticle at the base on the posterior keel. No double-rooted teeth are preserved. All broken cross sections show open pulp cavities. One well-preserved tooth has a long and relatively straight root (12.2+ mm) and has oval cross section (3.8 mm mediolateral and 4.2 anteroposterior diameters at the base of the crown), with a low crown (6.6 mm height).
Periotic
The periotic is a structurally complex and phylogenetically informative bone, and is considered in detail. When the periotic is in situ in archaic odontocetes, the ventral foramen of the facial canal can be seen in ventral view, but the fenestra ovalis is partially obscured by the pars cochlearis (see, for example, the genus Otekaikea). We infer a similar in situ orientation for ZMT 73, in which, however, the (right) periotic cannot be fitted to the (left) squamosal.
The periotic (–; )<<ts: Please flag Table 1 here>> has a short and wide anterior process, skewed medially, and a dorsoventrally weakly inflated pars cochlearis, which is slightly anteroposteriorly longer than the anterior process. Most of the posterior process is broken, but the preserved base of the posterior process is skewed laterally. Thus, both the anterior and posterior processes are oblique to the axis of the body of the periotic.
Table 1. Measurements in mm of ZMT 73: periotic and tympanic bulla. Dimensions follow Tanaka & Fordyce (2015a).
The anterior process is prolonged anteriorly and weakly medially, with a pointed anteroventral angle. The anterior process is deep and laterally swollen, with the medial tubercle of the anterior process (new term) (see discussion) and a swollen outer margin. Dorsally, a very weak anteroexternal sulcus runs from just anterior to the lateral tuberosity, passing into the parabullary sulcus, which is deep and weakly curved laterally. The anteroventral angle, as seen anteriorly, is a small sharp tip, just anterior to the anterior bullar facet. The anterior keel runs posterodorsally from the anteroventral angle, and stops anterior to the anteroexternal sulcus. The medial tubercle is pointed medially. Just behind the medial tubercle, the anterior incisure separates the anterior process and pars cochlearis, and runs to the level of the mallear fossa. The anterior bullar facet is a deep and slender groove, c. 10.0 mm long, with raised parallel-sided margins (3.6 mm width). The shallow fovea epitubaria is anteroposteriorly long, with a posterior border very close to the mallear fossa. The accessory ossicle is fused with the bulla, not the periotic.
The pars cochlearis is hemispherical in ventral view, transversely compressed and long (16.8 mm) relative to width (9.0 mm). The smooth anteromedial corner passes back into medial margin, with widening slightly toward the obtuse posteromedial angle. A tiny slit-like foramen for the hiatus Fallopii opens dorsal to the anterior incisure. The circular internal auditory meatus (maximum length, 5.3 mm, width, 5.5 mm) opens on the centre dorsomedially (B). The meatus contains four notable structures: (1) the proximal opening of the facial canal, (2) the foramen singulare, (3) the spiral cribriform tract, and (4) the area cribrosa media. A thin transverse ridge separates the large and rounded proximal opening for the facial canal (2.0 mm diameter) from the smaller foramen singulare (1.2 mm diameter). A larger, anteroposteriorly long and thick ridge separates the opening of the facial canal and foramen singulare from the spiral cribriform tract and area cribrosa media. The tract and pit for the area cribrosa media are rounded and almost the same size (2.0 mm diameter). The spiral cribriform tract is broken in its deeper reaches, and does not show the expected screw-like structure.
The fenestra rotunda is an anteroposteriorly short compressed reniform opening (transverse diameter 4.1 mm, anteroposterior length 2.8 mm). The aperture for the vestibular aqueduct and aperture for the cochlear aqueduct are both transversely long and elliptical (1.8 and 2.2 mm transverse diameters respectively).
On the body of the periotic, ventrally, the mallear fossa is deep, wide and slightly reniform in shape; the rim of the fossa connects with the fovea epitubaria. The lateral tuberosity is well developed both laterally and ventrally. Immediately behind the mallear fossa is a tiny deep submallear fossa (Tanaka & Fordyce Citation2016), not to be confused with the larger and more-posteriorly placed fossa incudis. The submallear fossa is associated with two posteriorly developed structures: the facial crest of the periotic and the parafacial sulcus. The parafacial sulcus (new term) is an anteroposterior groove between the facial crest (laterally) and the facial sulcus (medially). Earlier (Tanaka & Fordyce Citation2014), we used the term tympanohyal sulcus for the equivalent parafacial sulcus in Otekaikea marplesi, which ends at the anteromedial angle of the posterior process. The facial crest of the periotic arises at the base of the mallear fossa, lateral to the parafacial sulcus, and descends posteriorly to the apex of the posterior process. On the facial crest, the broken crus breve of the incus is cemented by matrix to the fossa incudis. The fenestra ovalis is a wide ellipse in profile, elongated anteromedially, small (2.5 mm) and filled by the footplate of the stapes. The hiatus epitympanicus is widely triangular and is clearly separated by the lateral tuberosity and the broken posterior process.
The rather flat dorsal surface of the periotic has an indistinct dorsal crest posteriorly, far from the internal acoustic meatus (8.8 mm). Such features differ from fossil platanistoids (Waipatia, Otekaikea, Notocetus), in which the dorsal crest lies close to the internal acoustic meatus.
Bulla
The right bulla (, ; ) is small, long and narrow heart-shaped, with a distinctive transverse depression on the involucrum. For purpose of description, the dorsal view is defined as the position of the bulla when the ventral face is sitting on a flat plane (, A).
The apex of the bulla is lost, but the anterolateral margin of the involucrum is tapering smoothly. The outer lip is widened to the level of the lateral furrow, which is a vertical crease. Medial to the prominent lateral furrow, a wide and inflated mallear ridge curves around the deep sulcus for the chorda tympani. The large, in situ, accessory ossicle has a gently (anteroposteriorly) curved suture with faintly raised edges, for the fovea epitubaria. The slender sigmoid process is depressed posteriorly, because of burial compaction, and the apex is lost. The prominent conical process is a rounded triangular nodule. Two broken bases of the posterior process lie just posterior and lateral to the conical process. It is not clear whether an elliptical foramen opened between the posterior process bases. The involucrum is narrow anteriorly but widens posteriorly, dramatically so at the middle of the involucrum. A transverse depression separates the narrow anterior of the involucrum from the wide posterior parts. The posterior end of the involucrum is blunt rather than rounded. Ventrally, a deep and narrow median furrow runs from the interprominential notch to the level of the lateral furrow. The mediolaterally thin outer posterior prominence projects strongly beyond the level of the inner posterior prominence.
Stapes
The right stapes, in situ, lacks the head. A depression on the medial surface presumably represents the non-patent vestigial stapedial foramen; the lateral face is smooth. The footplate is an elongate ellipse.
Incus and malleus
The incus and malleus were preserved in original articulation (). (During handling, after photography and after descriptive notes were made, both the malleus and incus broke along microscopic shears, and could not be reconstructed.) The incus is low and subconical, with a robust base around the facet for the malleus and a slender and tapered body and crus longum. The crus breve is broken; the apex is cemented by matrix in the fossa incudis of the periotic. The crus longum is perpendicular to the body, and its end has the lenticular process. On the incus, the incudomallear joint comprises a larger facet for the part of the manubrium of the malleus and smaller facet for the part of the anterior process of the malleus.
The malleus is larger than the incus, and is smoothly subspherical, with a prominent lateral depression near the broken base of the anterior process. The depression, which has been termed the foramen for the chorda tympani (e.g. Kellogg Citation1936, p. 193), appears imperforate. The anterior process is broken away. The facet for the incus has a thick lip.
Phylogenetic analysis
The phylogenetic position of ZMT 73 was analysed using the matrix of Tanaka and Fordyce (Citation2015a), slightly modified as below. Tables S1 and S2 show the datamatrix in nexus and TNT formats, respectively; Table S3 is the character list and Table S4 lists modified codings. Tables S5 and S6 are tree files for analyses 1 and 2, respectively. Figures S1 and S2 show the full trees which were used to produce the trees with collapsed clades of . Originally, the matrix was modified from that of Murakami et al. (Murakami et al. Citation2012b), which was designed to focus on phylogeny of Delphinoidea within the Odontoceti. The Tanaka and Fordyce (Citation2015a) matrix includes 83 extant and extinct taxa, all named and described except for OU 22670, and 287 characters (with 31 soft tissue characters) cited and/or modified from previous studies (Barnes Citation1985, Citation1990; Heyning Citation1989, Citation1997; Fordyce Citation1994; Arnold & Heinsohn Citation1996; Messenger & McGuire Citation1998; Geisler & Sanders Citation2003; Lambert & Post Citation2005; Lambert Citation2005b; Fajardo-Mellor et al. Citation2006; Lambert et al. Citation2008; Aguirre-Fernández et al. Citation2009; Geisler et al. Citation2011; Murakami et al. Citation2012a, Citation2012b, Citation2014).
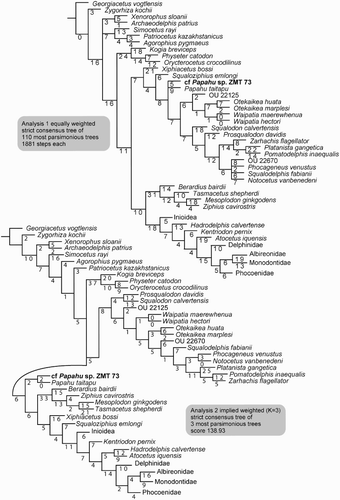
Figure 12. Phylogenetic analysis of ZMT 73 and Papahu taitapu. The clades Inioidea, Phocoenidae and Delphinidae are collapsed. (Cladograms with all clades shown are in Figures S1 and S2.) Top, Strict consensus tree of equally weighted Analysis 1 with branch length labelled. Bottom, Strict consensus tree of implied weighting Analysis 2 with branch length labelled.
The Tanaka and Fordyce (Citation2015a) matrix was modified in minor ways. Two taxa were added: ZMT 73 of this study, and OU 22670 of Fordyce et al. (Citation2015). Percentages of missing data of ZMT 73 were 86.0% (includes soft tissue characters) and 84.3% (excludes soft tissue). The ear bones are well coded; the periotic shows 16 of 31 characters, 54%, and the bulla 10 of 19 characters, 53%. Some other characters and codings were modified and deleted (see Table S4).
Character data and tree data were managed using Mesquite 2.75 (Maddison & Maddison Citation2011). Two different analyses were performed with TNT, 1.1 (Goloboff et al. Citation2008). All characters were treated as unweighted and unordered (Analysis 1), or implied weights with K=3 and unordered (Analysis 2). Both analyses used New Technology Search with the setting: recover minimum length trees = 1000 times with a backbone constraint of extant taxa, based on the topology of the McGowen et al. (Citation2009) molecular phylogeny. To measure node stability, we used the decay index method (Bremer Citation1994) for the strict consensus trees. After the analyses, species in some taxa among the Delphinida (Inioidea, Phocoenidae and Delphinidae) were collapsed for ease of illustrating.
Phylogenetic relationships
Analysis 1. Unweighted and unordered
The phylogenetic analysis shows 110 shortest trees of 1881 steps each. The strict consensus tree (, upper) shows a similar topology for Platanistoidea and Delphinida as for the Tanaka and Fordyce (Citation2015a: figure 21, upper) unweighted and unordered strict consensus tree. However, physeterids are now basal, and P. taitapu and cf. Papahu sp. ZMT 73 appear in a polytomy in the clade Platanistoidea. The Ziphiidae and Delphinida are more crownward.
Analysis 2. Implied weighting
The analysis recovers three shortest trees with a score of 138.93. The strict consensus tree (, lower) now excludes Papahu from the Platanistoidea. The major clades are: Physeteroidea in the base, followed by Platanistoidea, with the clade Papahu + cf. Papahu sp. ZMT 73 as the sister group to Ziphiidae + Eurhinodelphinidae + Delphinida.
Discussion
Amongst the included taxa, ZMT 73 is related closely to P. taitapu both in Analysis 1 and Analysis 2. Features on the periotic, bulla and teeth of ZMT 73 indicate morphologies not preserved on the type specimen of P. taitapu, for example, single-rooted teeth predicted from the empty alveoli in P. taitapu. As summarised in the diagnosis and elaborated below, ZMT 73 is not conspecific with P. taitapu, but the former is too incomplete to justify naming a new species. Papahu is discussed further below.
The positions of some higher-level clades of odontocetes differ from the Tanaka and Fordyce (Citation2015a) analyses because of the use of a constraints tree based on the McGowen et al. (Citation2009) molecular phylogeny. Of note, our current phylogenies reflect the molecular constraint tree in that the Platanistoidea lies between the more-basal Physeteroidea and the more-crownward Ziphiidae. Analysis 1 places P. taitapu and cf. Papahu (ZMT 73) in a polytomy toward the base of the Platanistoidea. Analysis 2 agrees with Tanaka and Fordyce (Citation2015a: figure 21) in that Papahu clade lies outside the Platanistoidea and immediately basal to the large clade comprising mainly Ziphiidae and Delphinida. We prefer the result of Analysis 2, rather than Analysis 1, because the implied weights approach deals better with homoplasy. Homoplasy is common with large data sets, and probably influenced the Tanaka and Fordyce (Citation2015a) morphological phylogenies, in which Physeteroidea and Ziphiidae formed a clade that disagrees with the molecular constraint tree. The implied weighting methodology of Analysis 2 is better, because it reduces the weight of the homoplastic characters.
Relationships between ZMT 73 and Papahu taitapu
cf. Papahu (ZMT 73) is placed with P. taitapu in the implied-weights Analysis 2 with two synapomorphies as below. These taxa share notable periotic features: a V‐shaped parabullary sulcus, a swollen outer margin of the anterior process (Character 176), a tapered and short anterior process, a narrow anteromedial sulcus, a robust lateral tuberosity and a large flat space between the internal acoustic meatus and faint dorsal crest. Some squamosal features are shared between ZMT 73 and P. taitapu: forming a deep suprameatal pit (Character 153), mediolaterally thin anterolateral edge of the zygomatic process, weak border between the anterior and posterior periotic fossae, relatively small post-tympanic process and a deep external acoustic meatus.
cf. Papahu (ZMT 73) differs from P. taitapu in some features in the phylogenetic analysis: a deeper neck muscle fossa (Character 114), shallower posterior portion of the periotic fossa (Character 155), abruptly deflected anterior process of the periotic (Character 173), and thinner pars cochlearis (Character 193). Some other differences were not included in the phylogenetic analyses: the anteroposteriorly thicker exoccipital; the post-tympanic process, which is formed only by a wedge-shaped region of squamosal (on P. taitapu, the anterior part of the exoccipital is topographically continuous with the posterior half of the post-tympanic process); robust postglenoid process (P. taitapu preserves only the base of the process); and more robust zygomatic process. There is no strong reason to consider the differences as ontogenetic variation. The type specimen of P. taitapu has evident sutures for the epiphyses and was identified as physically mature by Aguirre-Fernández and Fordyce (Citation2014), following the criteria of Perrin (1975); the few sutures apparent on ZMT 73 are firm, rather than open, suggesting submature to physically mature.
Comparison of Papahu with other Miocene dolphins
Papahu taitapu and cf. Papahu are phylogenetically distant from better-known Burdigalian and younger dolphins such as the Eurhinodelphinidae and Kentriodontidae. Differences include: both premaxillae extending posterior to nasals (Papahu) vs both premaxillae extending posterior only to the anterior tip of nasals (Kentriodontidae and Eurhinodelphinidae) (Character 59) and presence of the premaxillary cleft (Papahu; relevant bones not preserved in ZMT 73) vs absence (Character 86). The recently described specimens of Kentriodon diusinus Salinas-Márquez et al. Citation2014 and Chilcacetus cavirhinus Lambert et al., Citation2015 became known to us after our study was largely complete, and accordingly they are not included in the cladistic analyses. Ideally, the newly named specimens would be examined directly to confirm cladistic codings and allow reanalysis. For now, compared with Papahu and ZMT 73, Chilcacetus cavirhinus has a more slender anterior process and longer and deeper anterior bullar facet of the periotic. The anterior part of the involucrum of Chilcacetus is flat. Compared with Chilcacetus, the preserved teeth of ZMT 73 have a more pointed crown with a more strongly curved posterior keel, producing a similar condition to K. diusinus. Comparison between K. diusinus and C . cavirhinus shows at least two similarities on the skulls: having an anteriorly invaded medial portion of the frontals, and a long postorbital process.
Phylogenetic position of the genus Papahu
Originally, Aguirre-Fernández and Fordyce (Citation2014) placed P. taitapu among the stem odontocetes. The current analysis corroborates our earlier report on Otekaikea huata (see Tanaka & Fordyce Citation2015a), which placed Papahu as sister to the clade Physeterida + Eurhinodelphinidae + Delphinida. (Note that in the present trees, the molecular constraints helped split the former Physeterida into separate clades Physeteroidea and Ziphiidae.) It is not easy to compare the results from similar yet different data sets that used same analytical methods (both implied and equally weighted analyses). Our matrix here includes the Late Oligocene to earliest Miocene Otekaikea marplesi, O. huata and Waipatia hectori, and the late Early Miocene platanistoid Phocageneus venustus. These additional basal odontocetes probably improved character polarities, helping move P. taitapu from the Platanistoidea (as found earlier by Tanaka & Fordyce Citation2015a), to become the sister taxon for the Ziphiidae + Eurhinodelphinidae + Delphinida. One possible apomorphy of the Papahu clade and the clade Ziphiidae + Eurhinodelphinidae + Delphinida is the depressed involucrum in lateral view, as seen in cf. Papahu ZMT 73. The Eurhinodelphinidae shows depressed anterior and raised posterior parts of the involucrum. The Delphinoidea shows a sigmoidal or saddle-like involucrum. Conversely, fossil platanistoids (e.g. Waipatia, Otekaikea, Notocetus, Zarhachis) lack the sigmoidal involucrum, whereas the extant Platanista may show the feature occasionally. The state also occurs in the Physeteroidea, possibly homoplastic.
Periotic morphologies
The medial tubercle of the anterior process (a new term proposed in the description above) occurs as a flat step among the basal platanistoids, including the type specimens of Squalodon calvertensis, Waipatia maerewhenua, Otekaikea marplesi (earlier labelled as ‘weak tubercle’), O. huata, Notocetus vanbenedeni, Phocageneus venustus, Huaridelphis raimondii and a referred specimen of Zarhachis flagellator (USNM 10485). Waipatia maerewhenua and O. huata show the medial tubercle clearly, and it has a tiny vertical canal, which opens dorsoventrally. In addition, eurhinodelphinids also have the tubercle but more posteriorly (Eurhinodelphis cocheteuxi IRSNB M. 1856 of Lambert Citation2005b), Schizodelphis sulcatus PGN 2 of Benoit et al. (Citation2011) and enigmatic putative eurhinodelphinids (AMNH 102194, 102324 and 102550 of Fordyce Citation1983), but Xiphiacetus cristatus (IRSNB 3241-M. 1893 in Lambert Citation2005a) lacks the tubercle. Among the Ziphiidae, modern species do not show the tubercle well, because they have a swollen medial surface of the anterior process. But the most basal fossil ziphiid, Ninoziphius platyrostris (MNHN SAS 941 of Lambert et al. Citation2013) has the tubercle.
Conclusion
The hitherto undescribed Early Miocene Papahu-like ZMT 73 from Kaikoura is the sister taxon to P. taitapu in the molecular-constraints implied-weights tree. ZMT 73 shows some squamosal and periotic features not seen in P. taitapu, but is not complete enough to warrant a species name. The Papahu clade is sister to the Ziphiidae + Eurhinodelphinidae + Delphinida clade, with the Platanistoidea more basal. ZMT 73 helps reveal dolphin morphology during the poorly known Early Miocene.
Supplementary data
Table S1. Cladistic matrix in nexus format.
Table S2. Cladistic matrix in TNT format.
Table S3. Morphological characters used in the phylogenetic analysis.
Table S4. List of coding modifications.
Table S5. Tree file of Analysis 1.
Table S6. Tree file of Analysis 2.
Figure S1. Full tree of Analysis 1.
Figure S2. Full tree of Analysis 2.
Figure S2. Full tree of Analysis 2.
Download TIFF Image (1.1 MB)Figure S1. Full tree of Analysis 1.
Download TIFF Image (443.1 KB)Table S6. Tree file of Analysis 2.
Download (508 B)Table S5. Tree file of Analysis 1.
Download (148.7 KB)Table S4. List of coding modifications.
Download MS Word (123.2 KB)Table S3. Morphological characters used in the phylogenetic analysis.
Download MS Word (829.5 KB)Table S2. Cladistic matrix in TNT format.
Download (39.6 KB)Table S1. Cladistic matrix in nexus format.
Download (55.2 KB)Acknowledgements
We thank: Margaret A Bradshaw and Norton Hiller (former Curators of Geology, Canterbury Museum) for allowing research on ZMT 73, Gabriel Aguirre-Fernández (University of Zurich) for constructive comments that helped improve the manuscript, Hiroto Ichishima (Fukui Prefectural Dinosaur Museum) for discussion of the matrix, Naoki Kohno (National Museum of Nature and Science, Tokyo) for advice on analytical methodologies, and Jonathan Geisler (New York Institute of Technology) and an anonymous reviewer for helpful reviewing. Thanks to museum staff, especially David Bohaska, Nicholas Pyenson and Charles Potter of USNM, for access to specimens.
Associate Editor: Professor Kathy Campbell.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
Related Research Data
References
- Aguirre-Fernández G, Barnes LG, Aranda-Manteca FJ, Fernández-Rivera JR. 2009. Protoglobicephala mexicana, a new genus and species of Pliocene fossil dolphin (Cetacea; Odontoceti; Delphinidae) from the Gulf of California, Mexico. B Soc Geo Mex. 61:245–265.
- Aguirre-Fernández G, Fordyce RE. 2014. Papahu taitapu, gen. et sp. nov., an early Miocene stem odontocete (Cetacea) from New Zealand. J Vertebr Paleontol. 34:195–210. doi: 10.1080/02724634.2013.799069
- Arnold PW, Heinsohn GE. 1996. Phylogenetic status of the Irrawaddy dolphin Orcaella brevirostris (Owen in Gray): a cladistic analysis. Mem Queensl Mus. 39:141–204.
- Aze T, Ezard TH, Purvis A, Coxall HK, Stewart DR, Wade BS, Pearson PN. 2011. A phylogeny of Cenozoic macroperforate planktonic foraminifera from fossil data. Biol Rev. 86:900–927. doi: 10.1111/j.1469-185X.2011.00178.x
- Barnes LG. 1977. Outline of Eastern North Pacific fossil cetacean assemblages. Syst Zool. 25:321–343. doi: 10.2307/2412508
- Barnes LG. 1985. Evolution, taxonomy and antitropical distributions of the porpoises (Phocoenidae, Mammalia). Mar Mamm Sci. 1:149–165. doi: 10.1111/j.1748-7692.1985.tb00003.x
- Barnes LG. 1990. The fossil record and evolutionary relationships of the genus Tursiops. In: Leatherwood S, Reeves RR, editors. The bottlenose dolphin. San Diego, (CA): Academic Press Inc; p. 3–26.
- Barnes LG, Reynolds RE. 2009. A new species of early Miocene allodelphinid dolphin (Cetacea, Odontoceti, Platanistoidea) from Cajon Pass, Southern California, U.S.A. In: Albright LBI ed. Papers on geology, vertebrate paleontology, and biostratigraphy in honor of Michael O Woodburne. Flagstaff, Mus North Ariz Bull. 65:483–507.
- Benoit J, Adnet S, Welcomme J-L, Fabre P-H. 2011. New skull of Schizodelphis sulcatus Gervais, 1861 (Mammalia, Odontoceti, Eurhinodelphinidae) from the Lower Miocene of Pignan (Hérault, France) and its implications for systematics of Eurhinodelphinidae. Geobios. 44:323–334. doi: 10.1016/j.geobios.2010.10.006
- Bianucci G, Landini W. 2002. Change in diversity, ecological significance and biogeographical relationships of the Mediterranean Miocene toothed whale fauna. Geobios. 35:19–28. doi: 10.1016/S0016-6995(02)00045-1
- Bremer K. 1994. Branch support and tree stability. Cladistics. 10:295–304. doi: 10.1111/j.1096-0031.1994.tb00179.x
- Browne G, Field B. 1985. Lithostratigraphy of Late Cretaceous to Early Pleistocene rocks of northern Canterbury, New Zealand. N Z Geol Surv Rec. 6:1–63.
- Cione AL, Cozzuol MA, Dozo MT, Acosta Hospitaleche C. 2011. Marine vertebrate assemblages in the southwest Atlantic during the Miocene. Biol J Linn Soc. 103:423–440. doi: 10.1111/j.1095-8312.2011.01685.x
- Cozzuol M. 1996. The record of the aquatic mammals in southern South America. Munchner Geowiss Abh. 30:321–342.
- Fajardo-Mellor L, Berta A, Brownell RL, Boy CC, Goodall NP. 2006. The phylogenetic relationships and biogeography of true porpoises (Mammalia: Phocoenidae) based on morphological data. Mar Mam Sci. 22:910–932. doi: 10.1111/j.1748-7692.2006.00080.x
- Fordyce RE. 1980. The fossil Cetacea of New Zealand. (A catalogue of described genera and species with an annotated literature guide and reference list). Rep Geol Surv N Z. 90:1–60.
- Fordyce RE. 1983. Rhabdosteid dolphins (Mammalia: Cetacea) from the Middle Miocene, Lake Frome area, South Australia. Alcheringa. 7:27–40. doi: 10.1080/03115518308619631
- Fordyce RE. 1994. Waipatia maerewhenua, new genus and new species (Waipatiidae, new family), an archaic Late Oligocene dolphin (Cetacea: Odontoceti: Platanistoidea) from New Zealand. Proc San Diego Soc Nat Hist. 29:147–176. doi: 10.5962/bhl.part.10662
- Fordyce RE. 2003. Cetacean evolution and Eocene-Oligocene oceans revisited. In: Prothero DR, Ivany LC, Nesbitt EA, editors. From greenhouse to icehouse. The marine Eocene-Oligocene transition. New York (NY): Columbia University Press; p. 154–170.
- Fordyce RE, Tanaka Y, Ortega ME. 2015. An Early Miocene dolphin from New Zealand expands the range and diversity of Notocetus-like platanistoids. J Vertebr Paleontol, Program and Abstracts. 2015:125–126.
- Geisler JH, Sanders AE. 2003. Morphological evidence for the phylogeny of Cetacea. J Mamm Evol. 10:23–129. doi: 10.1023/A:1025552007291
- Geisler JH, McGowen MR, Yang G, Gatesy J. 2011. A supermatrix analysis of genomic, morphological, and paleontological data from crown Cetacea. BMC Evol Biol. 11:1–33. doi: 10.1186/1471-2148-11-112
- Geisler JH, Sanders AE, Luo Z-X. 2005. A new protocetid whale (Cetacea: Archaeoceti) from the late middle Eocene of South Carolina. Am Mus Novit. 3480:1–68. doi: 10.1206/0003-0082(2005)480[0001:ANPWCA]2.0.CO;2
- Gingerich PD. 2015. Body weight and relative brain size (encephalization) in Eocene Archaeoceti (Cetacea). J Mamm Evol. 1–15. doi:10.1007/s10914-015-9304-y
- Goloboff PA, Farris JS, Nixon KC. 2008. TNT, a free program for phylogenetic analysis. Cladistics. 24:774–786. doi: 10.1111/j.1096-0031.2008.00217.x
- Heyning JE. 1989. Comparative facial anatomy of beaked whales (Ziphiidae) and a systematic revision among the families of extant Odontoceti. Contrib Sci, Nat Hist Mus LAC. 405:1–64.
- Heyning JE. 1997. Sperm whale phylogeny revisited: analysis of the morphological evidence. Mar Mam Sci. 13:596–613. doi: 10.1111/j.1748-7692.1997.tb00086.x
- Hornibrook NdeB, Brazier RC, Strong CP. 1989. Manual of New Zealand Permian to Pleistocene foraminiferal biostratigraphy. N Z Geol Surv Paleontol Bull. 56:1–175.
- Jenkins DG. 1965. Planktonic foraminiferal zones and new taxa from the Danian to Lower Miocene of New Zealand. N Z J Geol Geophys. 8:1088–1126. doi: 10.1080/00288306.1965.10428156
- Kellogg R. 1936. A review of the Archaeoceti. Carnegie I Wash publ. 482:1–366.
- Köhler R, Fordyce RE. 1997. An archaeocete whale (Cetacea: Archaeoceti) from the Eocene Waihao greensand, New Zealand. J Vertebr Paleontol. 17:574–583. doi: 10.1080/02724634.1997.10011004
- Lambert O. 2005a. Review of the Miocene long-snouted dolphin Priscodelphinus cristatus du Bus, 1872 (Cetacea, Odontoceti) and phylogeny among eurhinodelphinids. B l Roy Sci Nat Belg, Sci-Earth. 75:211–235.
- Lambert O. 2005b. Phylogenetic affinities of the long-snouted dolphin Eurhinodelphis (Cetacea, Odontoceti) from the Miocene of Antwerp, Belgium. Palaeontology. 48:653–679. doi: 10.1111/j.1475-4983.2005.00472.x
- Lambert O, Bianucci G, de Muizon C. 2008. A new stem-sperm whale (Cetacea, Odontoceti, Physeteroidea) from the latest Miocene of Peru. C R Palevol. 7:361–369. doi: 10.1016/j.crpv.2008.06.002
- Lambert O, de Muizon C, Bianucci G. 2013. The most basal beaked whale Ninoziphius platyrostris de Muizon, 1983: clues on the evolutionary history of the family Ziphiidae (Cetacea: Odontoceti). Zool J Linn Soc. 167:569–598. doi: 10.1111/zoj.12018
- Lambert O, de Muizon C, Bianucci G. 2015. A new archaic homodont toothed cetacean (Mammalia, Cetacea, Odontoceti) from the early Miocene of Peru. Geodiversitas. 37:79–108. doi: 10.5252/g2015n1a4
- Lambert O, Post K. 2005. First European pontoporiid dolphins (Mammalia: Cetacea, Odontoceti) from the Miocene of Belgium and The Netherlands. Deinsea. 11:7–20.
- Maddison WP, Maddison DR. 2011. Mesquite: a modular system for evolutionary analysis. Available at http://mesquiteproject.org. Version 2.73.
- Marx FG, Fordyce RE. 2015. Baleen boom and bust: a synthesis of mysticete phylogeny, diversity and disparity. Roy Soc Open Sci. 2:140434. doi.org/10.1098/rsos.140434 doi: 10.1098/rsos.140434
- McGowen MR, Spaulding M, Gatesy J. 2009. Divergence date estimation and a comprehensive molecular tree of extant cetaceans. Molecular Phylogenetics and Evolution. 53:891–906. doi: 10.1016/j.ympev.2009.08.018
- McKay A. 1887. On the grey-marls and Weka Pass stone in Kaikoura Peninsula and at Amuri Bluff. N Z Geol Surv Rec Geol Explor. 18:74–78.
- Mead JG, Fordyce RE. 2009. The therian skull: a lexicon with emphasis on the odontocetes. Smithson Contrib Zool. 627:1–249. doi: 10.5479/si.00810282.627
- Messenger SL, McGuire JA. 1998. Morphology, molecules, and the phylogenetics of cetaceans. Syst Biol. 47:90–124. doi: 10.1080/106351598261058
- Miller GS. 1924. Some hitherto unpublished photographs and measurements of the blue whale. Proc U S Nat Mus. 66(7):1–4.
- Morgans HEG, Edwards A, Scott G, Graham I, Kamp PJ, Mumme T, Wilson G, Wilson GS. 1999. Integrated stratigraphy of the Waitakian-Otaian Stage boundary stratotype, Early Miocene, New Zealand. N Z J Geol Geophys. 42:581–614. doi: 10.1080/00288306.1999.9514864
- Morgans HEG, Scott G, Edwards A, Graham I, Mumme T, Waghorn D, Wilson G. 2002. Integrated stratigraphy of the lower Altonian (Early Miocene) sequence at Tangakaka Stream, East Cape, New Zealand. N Z J Geol Geophys. 45:145–173. doi: 10.1080/00288306.2002.9514966
- Murakami M, Shimada C, Hikida Y, Hirano H. 2012a. A new basal porpoise, Pterophocaena nishinoi (Cetacea, Odontoceti, Delphinoidea), from the upper Miocene of Japan and its phylogenetic relationships. J Vertebr Paleontol. 32:1157–1171. doi: 10.1080/02724634.2012.677299
- Murakami M, Shimada C, Hikida Y, Hirano H. 2012b. Two new extinct basal phocoenids (Cetacea, Odontoceti, Delphinoidea), from the upper Miocene Koetoi Formation of Japan and their phylogenetic significance. J Vertebr Paleontol. 32:1172–1185. doi: 10.1080/02724634.2012.694337
- Murakami M, Shimada C, Hikida Y, Soeda Y, Hirano H. 2014. Eodelphis kabatensis, a new name for the oldest true dolphin Stenella kabatensis Horikawa, 1977 (Cetacea, Odontoceti, Delphinidae), from the upper Miocene of Japan, and the phylogeny and paleobiogeography of Delphinoidea. J Vertebr Paleontol. 34:491–511. doi: 10.1080/02724634.2013.816720
- Perrin WF. 1975. Variation of spotted and spinner porpoise (Genus, Stenella) in the eastern Pacific and Hawaii. B Scripps I Oceanography. 21:1–206.
- Pilleri G. 1985. The Miocene Cetacea of the Belluno sandstones (eastern southern Alps). Mem Sci Geol. 37:1–250.
- Raine JI, Beu AG, Boyes AF, Campbell HJ, Cooper RA, Crampton JS, Crundwell MP, Hollis CJ, Morgans HEG. 2012. The New Zealand geological timescale. Retrieved 18 April 2014 from http://www.gns.cri.nz/Home/Our-Science/Earth-Science/Fossils/Online-Resources/New-Zealand-s-Geological-Timescale
- Raine JI, Beu AG, Boyes AF, Campbell HJ, Cooper RA, Crampton JS, Crundwell MP, Hollis CJ, Morgans HEG. 2015. Revised calibration of the New Zealand geological timescale. GNS Sci Rep. 2012/39:1–53.
- Rattenbury MS, Townsend D, Johnston MR. 2006. Geology of the Kaikoura area. Scale 1: 250 000. Institute of Geological and Nuclear Sciences geological map 13.
- Salinas-Márquez FM, Barnes LG, Flores-Trujillo JG, Aranda-Manteca FJ. 2014. Una especie de delfín fóssil (Cetacea; Delphinoidea; Kentriodontoidae) del Mioceno Medio de Baja California. B Soci Geol Mex. 66:145–164.
- Tanaka Y, Fordyce RE. 2014. Fossil dolphin Otekaikea marplesi (latest Oligocene, New Zealand) expands the morphological and taxonomic diversity of Oligocene cetaceans. PloS ONE. 9:e107972. doi: 10.1371/journal.pone.0107972
- Tanaka Y, Fordyce RE. 2015a. A new Oligo-Miocene dolphin from New Zealand: Otekaikea huata expands diversity of the early Platanistoidea. Palaeontol Electron. 18.2.23A:1–71.
- Tanaka Y, Fordyce RE. 2015b. Historically-significant late Oligocene dolphin Microcetus hectori Benham 1935: a new species of Waipatia (Platanistoidea). J Roy Soc N Z. doi:10.1080/03036758.2015.1016046
- Tanaka Y, Fordyce RE. 2016. Awamokoa tokarahi, a new basal dolphin in the Platanistoidea (late Oligocene, New Zealand). J Syst Palaeontol. doi:10.1080/14772019.2016.1202339
- Wade BS, Pearson PN, Berggren WA, Pälike H. 2011. Review and revision of Cenozoic tropical planktonic foraminiferal biostratigraphy and calibration to the geomagnetic polarity and astronomical time scale. Earth-Sci Rev. 104:111–142. doi: 10.1016/j.earscirev.2010.09.003