
ABSTRACT
The Early Miocene Waitemata and Waitakere group strata from Auckland and lower Northland, New Zealand encompass shallow rocky-shore, shallow soft-sediment and shallow-water through to outer-shelf facies. They contain an interesting brachiopod fauna comprising eight genera in six families and include the type localities for three species. Terebratella neozelandica is transferred to the genus Magasella and is confirmed as a separate species based on morphology and paleoecology. For the first time the type specimens of the species Liothyrella gravida and Magasella neozelandica are figured, Magadina browni and ‘Pachymagas’ are described and figured from North Island strata and Terebratulina suessi is described and figured from the Waitemata and Waitakere groups. The brachiopod fauna described here inhabited the margins of a subsiding marine basin in the Auckland and lower Northland area for c. 5 Ma in marginally tropical conditions in the Early Miocene. Six genera were also living in the warm–temperate Canterbury–Otago Basin at this time suggesting brachiopods may not be a reliable indicator of faunal provinces in New Zealand in the Early Miocene.
Introduction
Cenozoic brachiopods of New Zealand
The South Island of New Zealand has one of the best-documented Cenozoic brachiopod faunas in the world, with numerous publications describing faunas, including many of Miocene age, from localities in Canterbury, North Otago and Southland (Dawson Citation1990). In comparison, the Cenozoic brachiopod fauna of the North Island is poorly documented. The modest number of publications with full descriptions of faunas or individual species of brachiopod includes Lee and Wilson (Citation1979), who figured Early Miocene (Otaian) Notosaria antipoda (Thomson, Citation1918a) from Waitemata Group localities, Hiller (Citation2000, Citation2011) and Hiller et al. (Citation2008a, Citation2008b), who described Early Miocene (Otaian) brachiopods from Northland, Neall (Citation1970, Citation1972) who reviewed the taxonomy and described the paleoecology of Late Miocene (Kapitean)–Pleistocene (Castlecliffian) Neothyris Douville, Citation1879 from localities on the east and west coast of the North Island and Grant-Mackie and Chapman-Smith (Citation1971) who described and figured Pleistocene (Castlecliffian) Magasella haurakiensis (Allan, Citation1931) from East Cape.
This study documents a North Island Early Miocene (Otaian–Altonian) brachiopod fauna from shallow rocky-shore, shallow soft-sediment and shallow through to outer-shelf environments. In publications on the geology and paleontology of the Auckland area, the presence of fossil brachiopods was often noted and a wide variety of names assigned to them. This study includes full synonymies of the brachiopod genera and species names used in publications on the Waitemata and Waitakere groups. Two brachiopods at the genus level and five at the species level are described and figured. An eighth species (from the Waitakere Group) was comprehensively described and figured by Hiller et al. (Citation2008a) and Hiller (Citation2011) and is mentioned only briefly here. This study provides a map of localities from which brachiopods have been collected and includes a summary of their paleoecology. Whether Early Miocene brachiopods are reliable indicators of faunal province is discussed and the fauna of the Waitemata and Waitakere groups is compared with rocky-shoreline and Early Miocene brachiopod faunas from the South Island.
At least five New Zealand Cenozoic brachiopod genera appear to have an unwarranted number of named species: Liothyrella Thomson, Citation1916, Rhizothyris Thomson, Citation1915a, Waiparia Thomson, Citation1920, Magadina Thomson, Citation1915a and ‘Pachymagas’ (Allan Citation1957). Early taxonomists used a typological approach to brachiopod species, seemingly unaware of the possible morphological variability of a single species. For example, Thomson (Citation1920) described eight species of Pachymagas Ihering, Citation1903 from a single locality at Weka Pass, Canterbury. Two further New Zealand Cenozoic brachiopod genera with taxonomies in need of revision are Magasella Dall, Citation1870 and Magella Thomson, Citation1915a. The taxonomy, stratigraphic ranges and geographical ranges of these all genera need to be reviewed. This study makes a modest contribution to the review process of four of these genera. The type specimens of Liothyrella gravida (Suess, Citation1864) and Magasella neozelandica (Ihering, Citation1903), held in Vienna, Austria () are figured for the first time. The status of M. neozelandica as a species is confirmed based on morphology and paleoecology and localities in Southland and Fiordland where this species may occur are noted. The stratigraphic range of Magadina browni Thomson, Citation1915a is discussed, its geographical range is extended from north Canterbury to Auckland and a synonymous species name is suggested. Specimens of ‘Pachymagas’ from the North Island are described and figured for the first time.
Table 1. Fossil brachiopod specimens from the Waitemata Group strata collected as part of the Novara Expedition (Flügel 1959). These are held in the Natural History Museum, Vienna, Austria.
Some of the earliest work on the geology and paleontology of New Zealand was carried out around the sea cliffs of Auckland, on strata now included in the Waitemata Group. The first paleontological study of Waitemata Group invertebrates (Jones in Heaphy Citation1860) mentioned brachiopod specimens identified as Terebratulae. Hochstetter (Citation1864, Citation1867) and Hochstetter et al. (Citation1865) was the first geologist to explore New Zealand (Beu et al. Citation2012) and he took specimens of three brachiopod genera from Waitemata Group strata back to Vienna. These were described and illustrated (with pencil drawings) by Suess (in Zittel Citation1864) using ‘improved’ versions with missing parts of the original specimens replaced (Beu et al. Citation2012). These ‘improved’ illustrations were reproduced in Park (Citation1910) and Beu et al. (Citation2012). However, in the figure captions of both Park (Citation1910) and Beu et al. (Citation2012) the illustrations of ‘Terebratella dorsata’ are misnamed as ‘Rhynchonella nigricans’ and the illustrations of ‘Rhynchonella nigricans’ are misnamed as ‘Terebratella dorsata’.
Geological age and setting
The Waitemata Group is Otaian and the Waitakere Group is Otaian to mid Altonian. The Otaian stage ranges from 21.7 to 18.7 Ma, and the Altonian stage from 18.7 to 15.9 Ma (Raine et al. Citation2015). The Waitemata Group is a series of marine strata, mostly unfossiliferous sandstones and siltstones formed by turbidity flows that were deposited in a rapidly subsiding, elongate basin in the underlying greywacke basement with a row of stratovolcanoes emerging from the sea to the west and the north (Hayward Citation1979). The Waitemata Group extends from Whangarei in the north to Mt Pirongia in the south and from Kaipara in the west to the tip of the Coromandel Peninsula in the east (Ballance Citation1976). It has been divided into four subgroups; a basal shallow-water (0–20 m) Kawau Subgroup (Hayward & Brook Citation1984) and three bathyal (1000–2000 m) subgroups, the Bream Subgroup to the north, the Meremere Subgroup to the south and the Warkworth Subgroup in the centre (Isaac et al. Citation1994). The Kawau Subgroup is unconformably overlain by the Warkworth, Bream and Meremere subgroups (), reflecting the rapid tectonic subsidence of the Waitemata Basin and a period of sediment starvation (Isaac et al. Citation1994). Hayward (Citation1993) stated that the Kawau Subgroup is early Otaian and the overlying Warkworth Subgroup is middle Otaian.
Table 2. The groups, subgroups and formations mentioned in this study.
The basal Kawau Subgroup contains a wide range of lithologies, some richly fossiliferous localities including distinct faunas that developed in different marine settings (Grant-Mackie Citation1965) and is divided into three formations: Cape Rodney Formation, Tipakuri Sandstone Formation and Papakura Limestone Formation, each of which contain one or more members (Hayward & Brook Citation1984). Outcrops are found over the greater Auckland area, from Cape Rodney in the north to Raglan Harbour in the south and to Cape Colville, Coromandel Peninsula in the east (, ).
Figure 1. Localities mentioned in this study.
Table 3. Latitude and longitude for localities in Figure 1.
The Cape Rodney Formation to the north contains lenses of greywacke boulder breccias, cobble and pebble conglomerates and coarse sandstones with a shallow-water rocky-shoreline fauna (Hayward & Brook Citation1984). The Tipakuri Sandstone occurs throughout the Kawau Subgroup as lenses of coarse to fine calcareous sandstone (Hayward & Brook Citation1984). They contain mostly shallow-water soft-sediment faunas that are sparsely to richly fossiliferous (Eagle et al. Citation1995). The Papakura Limestone Formation occurs as lenses throughout the Kawau Subgroup and includes sandy, pebbly, gritty or shelly limestones, sometimes glauconitic, and shelly sandstones and conglomerates (Hayward & Brook Citation1984). The fauna ranges from shallow-water to outer-shelf assemblages. Detailed locality maps can be found in Hayward and Brook (Citation1984), Rickets et al. (Citation1989) and Campbell et al. (Citation2004). The Warkworth Subgroup is largely unfossiliferous, but includes the Parnell Grit, a widespread but discontinuous series of lenses of slumped volcaniclastic material from the unstable flanks of the Manukau and Kaipara volcanoes (Isaac et al. Citation1994) that often carried shallow-water fauna, including brachiopods (Fox Citation1902), into deep water.
The Waitakere Group also contains a wide range of lithologies, including igneous and volcanogenic sedimentary rocks, formed by volcanoes erupting both underwater and subaerially (Hayward Citation1976a). The Waitakere Group lies to the west and north of the Waitemata Group and comprises the Waipoua Subgroup of the Hokianga–Kaihu area, the Hukatere Subgroup of the Kaipara area, and the Manukau Subgroup, which makes up the Waitakere Ranges. In the Hukatere Subgroup, the fossiliferous Pakaurangi Formation is found on the Hukatere, Puketotara and Okahukura peninsulas (all in Kaipara Harbour, ) and contains volcaniclastic shelf sediments (Isaac et al. Citation1994), a range of sandstones, silty sandstones and thin micritic limestones (Jones Citation1969) and fossil faunas ranging from shallow-water to mid-shelf assemblages (Jones Citation1970). Scott (Citation1965), Beu (Citation1966) and Hiller (2008b) listed the Pakaurangi Formation as Otaian. Hayward (Citation1976b) noted that in the Manukau Subgroup most fossils occur in the fine-grained volcaniclastics of the Nihotupu Formation, with fossil faunas ranging from shallow-water to inner-shelf assemblages, gave an age range of late Otaian to mid Altonian for the Nihotupu Formation and included a detailed fossil locality map.
Materials and methods
The material is generally poorly preserved; most specimens are distorted and have matrix cemented to them internally and externally. Specimens showing internal features are rare. This study is based on collections held in the Naturhistorisches Museum, Vienna, Austria, the Auckland Museum, the University of Auckland School of Environment, Te Papa Tongarewa, the National Museum of New Zealand, Wellington, the Canterbury Museum, Christchurch and the Geology Museum, University of Otago, Dunedin (–). As far as the author is aware, this includes most of the brachiopod material collected from the Waitemata and Waitakere groups. The paleontological collections of Jones (Citation1970), Hayward (Citation1976b) and Brook (Citation1979, Citation1983) are held by the Auckland School of Environment and include a wide range of taxa; unfortunately, none of Hayward’s (Citation1976b) brachiopod specimens, and only a few of the specimens collected by Jones (Citation1970) and Brook (Citation1979, Citation1983) could be found. Note that Jones (Citation1970) and Brook (Citation1979, Citation1983) only provided fossil record file numbers of their localities and the names of their Hukatere Subgroup localities used in this study are taken from the Fossil Record Electronic Database (FRED, provided by GNS New Zealand).
Table 4. Material examined in this study.
Table 5. Material examined in this study.
The author visited many known fossil localities in the Kawau Subgroup but found few specimens (–, OU numbers). The author also collected bulk sediment samples from two coastal localities on the southern Hukatere Peninsula (Pakaurangi Formation, Waiteroa and Hollands members), the cliffs and wave-cut platforms that featured in Jones (Citation1969, Citation1970) and where Hiller (Citation2011) collected material. The bulk samples were dried at 90 °C, soaked with white spirits and then hot soapy water to disaggregate the clay fraction. The residue was again dried, then twice soaked in melted sodium thiosulfate that was allowed to cool and set to disaggregate the remaining matrix from the bioclastic material. The bioclastic residues were picked through under a binocular microscope. Only a very small amount of fossil brachiopod material was recovered, and only from the sample collected from the Hollands Member type section (–). Photographs of the type specimens of two species were kindly provided by the Naturhistorisches Museum Vienna, Austria. Photographs were taken at the University of Auckland on a Nikon D300 and at the University of Otago on a Nikon D70 camera and a Canon EOS1100D camera attached to a Nikon SMZ1000 microscope. Digital photographs of specimens were taken as multiple images and focus-stacked with ZereneStacker software. Two specimens of ‘Pachymagas’ were X‐rayed with a Skyscan 1172 Bruker-Micro CT scanner at the Otago Centre for Microscopy and the data processed with the open source computer software program ImageJ64 to produce sections through the valves.
Systematic paleontology
Note that only synonyms from publications describing or listing brachiopods from the Waitemata and Waitakere groups are given.
Phylum Brachiopoda
Class Rhynchonellata
Order Rhynchonellida Kuhn, 1949
Family Notosariidae Mancenido & Owen, 2002
Genus Notosaria Cooper, 1959
Notosaria antipoda (Thomson, 1918a)
()
Rhynchonella nigricans: Suess (in Zittel Citation1864), Hutton (Citation1873), Cox (Citation1881), Park (Citation1886a), Fox (Citation1902), Clarke (Citation1905), Park (Citation1910), Flügel (Citation1959), Beu et al. (Citation2012)
Rhynchonella: Buchanan (Citation1870), Cox (Citation1882), Park (Citation1886b)
Hemithyris antipoda: Thomson (Citation1918a), Firth (Citation1929)
Tegulorhynchia nigricans: Laws (Citation1931)
Tegulorhynchia cf. antipoda: Skinner (Citation1969), Grant-Mackie and Moore (Citation1970)
Notosaria sp.: Jones (Citation1970), Brook (Citation1979)
Tegulorhynchia squamosa: Lee (Citation1980), Brook (Citation1983)
Notosaria sp. cf. N. antipoda: Dawson (Citation1990)
Notosaria antipoda: Lee and Wilson (Citation1979), Hayward and Brook (Citation1984), Eagle et al. (Citation1994, Citation1995, Citation1999), Campbell et al. (Citation2004), Beu et al. (Citation2012)
Suess (Citation1864) first described and illustrated specimens from the Waitemata Group strata as Rhynchonella nigricans, based on material collected by Mr Charles Heaphy from south of Cape Rodney and presented to Hochstetter (Beu et al. Citation2012). Thomson (Citation1918a) transferred the species to the genus Hemithyris Orbigny, Citation1847 and gave it the new species name antipoda. The type material of Hemithyris (from the Rakaia River, Canterbury) is held in Te Papa Tongarewa, the National Museum of New Zealand (holotype BR001262). Lee and Wilson (Citation1979) placed this species into the genus Notosaria, and figured the holotype together with specimens from three Kawau Subgroup localities, Te Kohuroa/Mathesons Bay, Motutapu Island and Cape Rodney.
Lee (Citation1980) stated that a single specimen of Tegulorhynchia squamosa was known from the Otaian of the Hukatere Peninsula (the only known North Island occurrence), but gave no details of locality or collection number. Brook (Citation1983) listed Tegulorhynchia squamosa from Strawberry Bay, Hukatere Peninsula. Both studies may refer to the same specimen. The Fossil Record Electronic Database lists Notosaria from Brook’s locality. The specimen is unavailable, but it may have been a morphological variant of Notosaria antipoda.
Stratigraphic range
Whaingaroan (latest Eocene–Early Oligocene) to Otaian (Early Miocene) (Lee & Wilson Citation1979).
Otaian age specimens have been collected at localities in Southland, Canterbury and Auckland (Lee & Wilson Citation1979).
Kawau, Warkworth and Hukatere subgroup localities for Notosaria antipoda
Published Kawau Subgroup localities include Cape Rodney (Hayward & Brook Citation1984), Te Kohuroa/Mathesons Bay (Hayward & Brook Citation1984; Eagle et al. Citation1999), Kawau Island (Hayward & Brook Citation1984; Eagle et al. Citation1994), Motuketekete Island (Hayward & Brook Citation1984; Campbell et al. Citation2004), Double‐U Bay, Waiheke Island (Eagle et al. Citation1995), Motutapu Island (Grant-Mackie & Moore Citation1970; Lee & Wilson Citation1979), Little Omaha Quarry (Clarke Citation1905), Waikopua Stream (Clarke Citation1905; Firth Citation1929), Hays Stream, Papakura (Park Citation1886a; Eagle & Hayward Citation1992) and Cape Colville, Coromandel (Skinner Citation1969; Grant-Mackie & Moore Citation1970). Fox (Citation1902) listed specimens from the Warkworth Subgroup locality Cheltenham Beach (). Unpublished Kawau Subgroup localities include Goat Island and Te Motu-a-Ihenga/Motuihe Island (, ). The single unpublished Hukatere Subgroup locality is Strawberry Bay, Hukatere Peninsula (Brook Citation1979, Citation1983).
Description
Small to medium-sized brachiopod, wider than long, transversely sub-oval to sub-pentagonal (A–B). Valve surface strongly costellate with concentric growth lines, costae increase in number by bifurcation and intercalation. Notosaria antipoda may be spinose; Lee and Wilson (Citation1979, p. 466) described the spines as ‘short, tent-like projections’ of the growing shell edge. Spines are not preserved on this material, but the characteristic broken edges of growth increments (A) suggest that some specimens had spines. The foramen is large and hypothyrid, the deltidial plates narrow and disjunct. The dorsal valve has a fold and the ventral valve has a corresponding sulcus, the anterior commissure is weakly to strongly unisulcate (Lee & Wilson 1976, figure 5.2, 5.6). Crura were not preserved in any Waitemata Group material. The cardinal process is bi-lobed and there is a low dorsal septum (C–D).
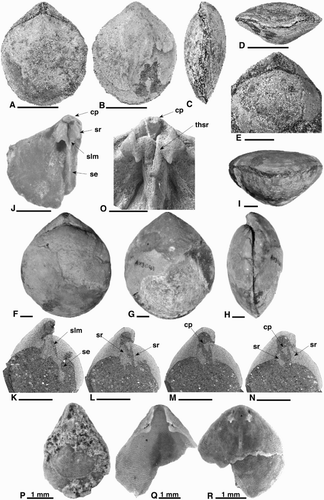
Figure 2. A–D, Notosaria antipoda. A, MA34672a. Dorsal exterior. B, B629. Dorsal exterior. C–D, MA34672b. C, Dorsal interior. D, Close up of sockets, socket ridges and septum. E, F, Thecidellina cf. maxilla CM 2006.64.92. E, Dorsal interior. F, Dorsal interior, posterior tilted forward. G–I, Liothyrella gravida. Type specimens. G, H, NHMW‐1959-0335-0012a. G, Ventral exterior. H, Lateral exterior (ventral valve only). I, NHMW‐1865-0037-0105a. Dorsal exterior of partial specimen. J–N, Terebratulina suessi. J, OU45764. Dorsal exterior of juvenile. K–N, MA77312. K, Dorsal exterior. L, Ventral exterior. M, Lateral exterior. N, Anterior commissure. E–G photographed by Alice Schumacher of the NHMW. All scale bars 5 mm unless indicated otherwise. se, septum; sp, broken spine base. B = University of Auckland School of Environment, CM = Canterbury Museum, Christchurch, MA = Auckland Museum, NHMW = Naturhistorisches Museum Vienna, OU = University of Otago Geology Museum, Dunedin.

Remarks
Notosaria has been found in four other New Zealand Cenozoic rocky-shore faunas (Carter Citation1972; Lee et al. Citation1983, Citation2014; Bosel & Coombs Citation1984) and appears to have occupied this habitat since at least the Late Oligocene (Duntroonian) (Lee et al. Citation1983). Holocene Notosaria nigricans (Sowerby, Citation1846) appears to be the direct descendant of Early Miocene N. antipoda (Lee & Wilson Citation1979) and still occupies rocky shoreline habitats in many parts of New Zealand and deeper water habitats down to nearly 800 m (Lee Citation1978).
Order Thecideida Elliot, 1958
Family Thecididae Gray, 1840
Genus Thecidellina Thomson, 1915b
Thecidellina cf. maxilla
Thecidellina sp. Hiller (2011)
Thecidellina is very rare in the Waitakere Group strata and unknown in the Waitemata Group. A single dorsal valve was collected, described and figured by Hiller (Citation2011, A) from the Pakaurangi Formation (Waiteroa Member) at Hollands Point, Hukatere Peninsula (). The author collected bulk samples from within several hundred metres of Hiller’s locality (at the Hollands Member type section) but found no more thecideid specimens. The original figure of Hiller (Citation2011) was very small so the specimen is refigured here. In the original figure the specimen is more complete but has more matrix on it, presumably it was damaged while being cleaned.
The single valve, although damaged, is similar to Thecidellina maxilla (Hedley, Citation1899), a specimen in a similar state of preservation is shown in Lee and Robinson (Citation2003, figure 34). Lee and Robinson (Citation2003) placed Thecidellina specimens from the Early Miocene of Java, the Miocene of Fiji (Cooper Citation1978), the ‘post-Miocene’ of Eniwetok and Bikini Atolls (Cooper Citation1964) and the Pleistocene of Vanuatu (Cooper Citation1978) into T. maxilla, extending the stratigraphic range of this species back to the Early Miocene. Lee and Robinson (Citation2003) figured Pliocene material identified as Thecidellina maxilla from the Chatham Islands and Brook (Citation1998) found Pleistocene specimens in the Kermadec Islands attached to corals.
Holocene T. maxilla is widespread in the Pacific, being known from the Marshall Islands (Cooper Citation1954), New Caledonia (Laurin Citation1997; Lee & Robinson Citation2003; Bitner Citation2010), Fiji and the Marquesas Islands (Bitner Citation2005), the Austral Islands (Bitner Citation2007), Norfolk Ridge (Bitner Citation2009), the Great Barrier Reef, Australia (Hoffman et al. Citation2009) and French Polynesia (Bitner Citation2014). Lee and Robinson (Citation2003) also described and figured Thecidellina japonica (Hayasaka, Citation1938) from Japan, and suggested that this species may be synonymous with T. maxilla. However, since then, there has been a lot of research into the morphology and taxonomy of thecideids and Hoffman et al. (Citation2009) suggested that specimens identified as T. maxilla from various localities in the Pacific show previously undescribed morphological variations and may represent a species-complex. If further research proves this to be correct, the fossil material in the Pacific will also need re-examination.
Description
The specimen (E–F), a single dorsal valve, is very small, 1.9 mm long by 1.9 mm wide, and incomplete, the brachial bridge and calcitic pole are missing. The valve is sub-quadrate, the cardinal process is tri-lobate, the auriform interbrachial lobes have serrated margins and partially developed (or damaged) spicule canopies. The median septum is wide anteriorly but quickly becomes narrow and the sub-peripheral margins have scattered tubercles.
Order Terebratulida Waagen, 1883
Family Terebratulidae Gray, 1840
Genus Liothyrella Thomson, 1916
Liothyrella gravida (Suess, 1864)
(pl. 9, )
Waldheimia gravida: Suess (in Zittel Citation1864), Hochstetter et al. (Citation1865), Hutton (Citation1870), Fox (Citation1902), Flügel (Citation1959), Beu et al. (Citation2012)
?Waldheimia: Fox (Citation1902)
Magellania (Waldheimia) gravida: Clarke (Citation1905)
Magellania gravida: Park (Citation1910)
Liothyrella gravida: Laws (Citation1931), Allan (Citation1960), Dawson (Citation1990), Beu et al. (Citation2012)
Liothyrella boehmi: Hayward (Citation1976b)
Liothyrella cf. elongata: Hayward (Citation1976b)
Liothyrella sp. Jones (Citation1970), Brook (Citation1979; Citation1983)
Suess (Citation1864) first named, described and illustrated specimens from the Papakura Limestone (Waitemata Group) as Waldheimia gravida, based on material collected by Hochstetter from Coopers and Smiths quarries (). These localities are no longer accessible, but a new standard reference section for the Papakura Limestone has been erected nearby in Hays Stream (Waterhouse Citation1974). Thomson (Citation1916) proposed the genus Liothyrella for extant and fossil brachiopod material and Laws (Citation1931) first used the name Liothyrella gravida for Kawau Subgroup material. The only figures of this species previously published are the ‘improved’ illustrations by Suess (Citation1864), reproduced in Park (Citation1910) and Beu et al. (Citation2012). The type specimens are held in the Naturhistorisches Museum Vienna, Austria () and two of these specimens are figured here.
Stratigraphic range
Otaian–Altonian (Early Miocene).
Ten Liothyrella species (and 18 lots of material listed as Liothyrella sp.), ranging in age from Late Eocene to Holocene, have been described or listed from New Zealand (Dawson Citation1990). Many of these names may be synonymous with Liothyrella gravida which is the oldest valid name. The actual stratigraphic and geographical range of L. gravida may be extensive. The missing specimens of Hayward (Citation1976b) from the Waitakere Subgroup are placed in L. gravida.
Kawau, Warkworth, Manukau and Hukatere subgroup localities for Liothyrella gravida
Published Kawau Subgroup localities include Coopers and Smiths quarries (Suess Citation1864), Hays Stream (Hochstetter et al. Citation1865; Clarke Citation1905) and Cape Colville, Coromandel (Skinner Citation1969). Published localities for the Warkworth Subgroup include Cheltenham Beach and Whangaparaoa (Fox Citation1902) and the single published locality for the Manukau Subgroup is Maori Bay (Hayward Citation1976b). Unpublished localities include Waikawau Beach, Motutapu Island and Waikopua Stream (, ). Published localities for the Hukatere Subgroup include Coates Bay, west of Waikupua Bay and north of Pakaurangi Point (Jones Citation1970) and (possibly) Hollands Point (Hiller Citation2011). Unpublished Hukatere Subgroup localities (questionably) include Nihotetea Creek and Waihangaru Creek, Hukatere Peninsula (Brook Citation1979, Citation1983).
Description
Liothyrella is a medium to large brachiopod, elongate oval in outline (G) with a sub-erect to erect beak (H). The foramen is epithyrid and often labiate (I). The commissure is rectimarginate or may be broadly unisulcate in large specimens. The valves are smooth with faint growth lines (G).
Remarks
Liothyrella is rare in the Waitemata and Waitakere group strata. No complete specimens, or specimens showing the interior features, were available in any of the collections. Brook (Citation1979) listed two localities where Liothyrella had been collected and noted (p. 40) that in the ‘lateral equivalent’ to the Pakaurangi Formation on the western side of Hukatere Peninsula Liothyrella is found as ‘scattered solitary valves and rarely with valves conjoined.’ If this locality can produce complete specimens of Liothyrella this could be very useful for the needed revision of fossil Liothyrella species in New Zealand, these being stratigraphically close to the type section of the incomplete type specimens of L. gravida from Papakura. However, the only two specimens of Brook (Citation1979, Citation1983) misidentified as Liothyrella in the collections at the University of Auckland (AU10103a-b from Nihotetea Creek) appear to be ‘Pachymagas’. Hiller (Citation2011) found three valves that might be Liothyrella in the Pakaurangi Formation (Waiteroa Member) at Hollands Point (named as Terebratulidae indet.) but no specimens had a loop. Holocene Liothyrella neozelanica Thomson, Citation1918b is likely to be a direct descendant of Miocene L. gravida.
Family Cancellothyridoidea Thomson, 1926
Genus Terebratulina Orbigny, 1847
Terebratulina suessi (Hutton, 1873)
Terebratulina suessi: Eagle and Hayward (1992), Eagle et al. (1994; 1995)
Suess (Citation1864) first described this species as Terebratulina sp. from specimens collected from Waikato South Head by Hochstetter, but mistakenly named it Terebratella in the illustration. Hutton (Citation1873) briefly described specimens (from localities in the South Island and Chatham Islands) and named them Terebratella suessi (copying the mistake by Suess). Hutton (Citation1904) corrected the name to Terebratulina, and described the internal and external features. Eagle and Hayward (Citation1992) first reported T. suessi from Waitemata Group strata and Eagle et al. (1992, Citation1994, Citation1995) provided small, stylised illustrations of T. suessi as part of fossil community reconstructions. The type material is held in Te Papa Tongarewa (holotype BR001265).
Stratigraphic range
Kaiatan (Late Eocene)–Otaian (Early Miocene) (Dawson 1990).
This species has a long stratigraphic range and has been found at many fossil localities, particularly in the South Island, but also in the North Island and Chatham Islands (Dawson Citation1990).
Kawau and Hukatere subgroup localities for Terebratulina suessi
Kawau Subgroup published localities include Hays Stream (Eagle & Hayward Citation1992), Fossil Point, Kawau Island (Eagle et al. Citation1994) and Fossil Bay, Waiheke Island (Eagle et al. Citation1995). However, specimens from Fossil Point misidentified as T. suessi in the collections of Auckland Museum (MA81968) are here designated Magadina browni. Only a single specimen of T. suessi (from Hays Stream) was held in the various collections examined (at Auckland Museum, ). A second juvenile specimen was collected by the author from the Pakaurangi Formation (Hollands Member type section) at Hukatere Peninsula ().
Description
The juvenile specimen (J) is very small (1.5 mm in length) but clearly resembles the second, larger specimen with fine costellae on the valve exterior. The larger specimen is teardrop-shaped and the valves are ornamented with fine costellae and concentric growth rings (K–L). Although the specimen is small for this species, the small size of the foramen (compared with the shell length) suggests that it is not a juvenile. The foramen is wide and submesothyrid, the deltidial plates are small and disjunct (K). The beak is straight (M) and the anterior commissure is rectimarginate (N).
Remarks
At South Island fossil localities, T. suessi adults range in size from 10 to 27 mm (Allan Citation1932). Three extant species of Terebratulina are found to the north and northeast of New Zealand (unpublished data).
Family Megathyrididae Dall, 1870
Genus Argyrotheca Dall, 1900
Argyrotheca kupei Hiller, Robinson & Lee, 2008a
Argyrotheca kupei: Hiller (2011)
Hiller et al. (2008) first described this new species based on specimens collected from the Pakaurangi Formation (Waiteroa Member) at Hollands Point, Hukatere Peninsula (the type locality) and material from five localities in Northland. The type specimen (and all other material) is held in the Canterbury Museum (holotype CM 2006.64.13). Hiller (Citation2011) figured further specimens from Hollands Point. The descriptions and figures in these two studies cannot be improved and are not repeated here.
Family Terebratellidae King, 1850
Subfamily Terebratellinae King, 1850
Genus Magasella Dall, 1870
Magasella neozelandica (Ihering, 1903)
Terebratella dorsata: Suess (Citation1864), Hutton (Citation1873), Cox (Citation1881), MacKay (Citation1884), Park (Citation1886b), Park (Citation1889), Fox (Citation1902), Clark (Citation1905), Park (Citation1910), Flügel (Citation1959), Beu et al. (Citation2012)
Terebratella cruenta: Fox (Citation1902), Clarke (Citation1905)
Terebratella neozelandica: Ihering (Citation1903), Laws (Citation1931), Allan (Citation1960), Campbell et al. (Citation2004)
Terebratella sanguinea: Laws (Citation1931)
Magasella neozelandica: Grant-Mackie and Moore (Citation1970), Dawson (Citation1990), Eagle and Hayward (Citation1992), Eagle et al. (Citation1994; Citation1999)
Terebratella neozelanica: Hayward and Brook (Citation1984)
Terebratella radiata: Beu et al. (Citation2012)
Suess (Citation1864) first described and illustrated specimens from the Waitemata Group strata as Terebratella dorsata (Gmelin, Citation1790) (a species now considered endemic to southern South America) from material collected south of Cape Rodney by Mr Charles Heaphy and presented to Hochstetter (Beu et al. Citation2012). Ihering (Citation1903) renamed the New Zealand fossil species Terebratella neozelandica. The only figures of this species previously published are the ‘improved’ illustrations by Suess ( Citation1864), reproduced in Park (Citation1910) and Beu et al. (Citation2012). The type specimens are held in the Naturhistorisches Museum Vienna, Austria () and two of these specimens are figured here.
Allan (Citation1949) examined South American brachiopods and stated that New Zealand Terebratella was not congeneric with South American Terebratella and moved the New Zealand extant and fossil species to the genus Magasella. The New Zealand species were transferred back and forth between Terebratella and Magasella five times between 1949 and 1993. Cohen et al. (Citation2011) showed through molecular analysis that Holocene New Zealand Terebratella and Holocene South American Terebratella are not congeneric, finally vindicating Allan. Robinson et al. (Citation2016) transferred the two Holocene species of New Zealand Terebratella back to the genus Magasella. This study returns Otaian Terebratella neozelandica from the Waitemata Group to the genus Magasella.
Stratigraphic range
Duntroonian? (Late Oligocene)–Altonian? (Early Miocene).
Kawau, Warkworth, Hukatere and Manukau subgroup localities for Magasella neozelandica
Published Kawau Subgroup localities include Cape Rodney (Hayward & Brook Citation1984), Te Kohuroa/Mathesons Bay (Hayward & Brook Citation1984; Eagle et al. Citation1999), Kawau Island (Hayward & Brook Citation1984; Eagle et al. Citation1994), Motuketekete Island (Hayward & Brook Citation1984; Campbell et al. Citation2004), Motutapu Island (Grant-Mackie & Moore Citation1970), Cape Colville, Coromandel, (Skinner Citation1969; Grant-Mackie & Moore Citation1970) and Hays Stream (Laws Citation1931; Eagle & Hayward Citation1992). Unpublished Kawau Subgroup localities include Goat Island and Te Motu-a-Ihenga/Motuihe Island. Fox (Citation1902) listed this species from Warkworth Subgroup locality Cheltenham Beach (, ).
Jones (Citation1970) listed Otaian specimens from Pakaurangi Point (Hukatere Subgroup) as cf. Waltonia Davidson, Citation1850 and Hayward (Citation1976b) listed Altonian specimens from a road cutting on Scenic Drive, Waiatarua (Manukau Subgroup), as ?Waltonia. However, Holocene Waltonia is now Calloria Cooper & Lee, Citation1993 and the oldest fossil Calloria known are from the Late Pliocene (Nukumaruan) of Whanganui (Allan Citation1960; Carter Citation1972). Some Kawau Subgroup localities have specimens of M. neozelandica that are smooth, strongly sulcate, with a large foramen and disjunct deltidial plates that are morphologically similar to Holocene Calloria inconspicua (Sowerby, Citation1846) (for example, lot AU11073 contains 23 smooth specimens from Motutapu Island). The specimens of Jones (Citation1970) and Hayward (Citation1976b) may be smooth Magasella neozelandica but the material cannot be found in the University of Auckland collections. The stratigraphic range for this species may or may not extend into the Altonian.
Description
This is a medium-sized brachiopod, the largest ventral valve examined was 35 mm long and 38 mm wide. The outline is sub-oval to sub-pentagonal and the valves may be ribbed (A–B) or smooth (C–D). The anterior commissure is sulcate (E–F), the dorsal valve has a narrow sulcus and the ventral valve a corresponding narrow fold that is strongly raised (E–G). The foramen is large and submesothyrid, the deltidial plates are triangular and disjunct (H). The beak is sub-erect to erect and very robust with strongly developed beak ridges (H–I) but may be very worn (attrite) (B). No specimens examined have a preserved loop, but seven specimens show broken cardinalia. The cardinal process is not preserved. The inner hinge plates form a septalium (J–K), the dorsal septum is low and thin and about half the valve length, and the connecting bands attach to the septum at approximately one-third of the valve length (K).
Figure 3. A–K, Magasella neozelandica. A, NHMW‐1865-0037-0150i. Dorsal exterior. B, MA91104a. Dorsal exterior. C, D, B630. C, Dorsal exterior. D, Lateral exterior. E, NHMW‐1865-0037-0150b. Anterior commissure. F, MA91104b. Anterior commissure. G, B631. Ventral exterior. H, I, B632. H, Strong beak ridges, disjunct deltidial plates. I, Erect beak. J, B633. Cardinalia. K, B634. Cardinalia and septum. L–O, Magasella sanguinea. L–N, OU44534a. L, Dorsal exterior. M, Anterior commissure. N, Less robust beak ridges, conjunct deltidial plates. O, OU44534b. Cardinalia and septum. A and E photographed by Alice Schumacher of the NHMW. All scale bars 5 mm unless indicated otherwise. B = University of Auckland School of Environment, MA = Auckland Museum, NHMW = Naturhistorisches Museum Vienna.

Remarks
Allan (Citation1949, p. 289) stated of fossil Magasella in New Zealand ‘The multicostate Magasella sanguinea (Leach, Citation1814) is a very conservative group. Specimens which are difficult to separate specifically [from extant specimens] occur as early as the Duntroonian stage’. This study examined Duntroonian–Otaian specimens of Magasella from rocky-shore facies and shallow to mid-shelf facies from five South Island localities and specimens of Holocene M. sanguinea from Doubtful Sound, Fiordland (). When compared with Magasella neozelandica, Holocene M. sanguinea and fossil Magasella specimens from inner to mid-shelf limestones and greensands have a wider and shallower dorsal sulcus and ventral fold (L–M), a less robust beak and a foramen with conjunct deltidial plates (N). The cardinalia appear to be similar between M. neozelandica and Holocene M. sanguinea (O).
Holocene Magasella species have been collected at a depth range of 11–260 m (mostly deeper than 30 m, NIWA data) and may be attached to a substrate or may be free-lying on the sea floor (Richardson Citation1981). The paleoecological reconstructions of Eagle et al. (Citation1994, Citation1995, Citation1999) and Campbell et al. (Citation2004) suggest that M. neozelandica collected from the Kawau Subgroup lived in high-energy sub-tidal waters between 0 and 20 m deep. The large foramen, disjunct deltidial plates and often worn beak of M. neozelandica suggest a large pedicle holding the shell tightly against the rocky substrate in turbulent waters. This study suggests that these Otaian specimens are both morphologically distinct from the conservative M. sanguinea fossil lineage and lived in a distinctly different habitat, and confirms M. neozelandica as a separate species.
The Magasella specimens from the Mount Luxmore rocky-shore fauna (Duntroonian–Waitakian) (Lee et al. Citation1983) include one complete specimen (with the foramen obscured by matrix) and five partial valves. The notable features include a narrow and deep sulcus on the dorsal valve and corresponding narrow fold on the ventral valve of the complete specimen, and a strong, narrow ventral fold on four of the partial valves. This material appears to be morphologically closer to M. neozelandica from Waitemata Group strata than to fossil Magasella from shallow to mid-shelf facies. The two Magasella specimens from the Bluff rocky-shore fauna (Waitakian–Otaian) (Bosel & Coombs Citation1984) are juvenile but already have a well-developed sulcus at just 7–9 mm in length. The dorsal valves of Holocene M. sanguinea are faintly sulcate at this size. A full review of New Zealand fossil Magasella is planned by the author and the material of Mount Luxmore and Bluff is not identified beyond the genus level here. However, this study suggests this material may be M. neozelandica and thus the stratigraphic range of this species may extend back to the Duntroonian. The rocky-shore habitat of Early Miocene M. neozelandica is now occupied in many places in New Zealand by Holocene Calloria inconspicua and Calloria variegata Cooper & Doherty, Citation1993.
Genus Magadina Thomson, 1915a
Magadina browni Thomson, 1915a
(Figure 7a–d)
Magadina waiparaensis: Dawson (1990)
Thomson (Citation1915a) first described this genus and named two species from two closely adjacent type localities (Mount Brown Formation, Middle Waipara River, North Canterbury), Magadina browni and Magadina waiparaensis. Thomson (1915, p. 40) stated of M. waiparaensis ‘This species so closely resembles M. browni that the differences only need to be indicated.’ The differences described by Thomson are very minor, well within the range of variability of a single species. Dawson (Citation1990) noted that four specimens held in Te Papa Tongarewa (BR001013) collected from Cape Rodney were identified by Thomson as Magadina cf. waiparaensis. The author has examined these four specimens, specimens from Waitemata strata held in the Auckland Museum and University of Auckland School of Environment and more than 200 specimens from the type section and nearby localities in North Canterbury () and considers M. waiparaensis to be a junior synonym of M. browni, which has page priority. This study places all the specimens from the Kawau Subgroup into M. browni. The type material of M. browni is held in Te Papa Tongarewa (holotype BR0046).
Stratigraphic range
Duntroonian? (Late Oligocene)–Waiauan? (Middle Miocene).
Few publications mention this species and its age range is not well constrained. Most known localities are in the Waipara District, North Canterbury where Magadina browni is found in the Mount Brown beds, which range from Otaian to Waiauan (Newman & Bradshaw Citation1981). Hills (Citation1971) listed this species from the Whiterock Limestone (Otaian), Karetu River, North Canterbury. The Fossil Record Electronic Database records this species from the Duntroonian on the West Coast of the South Island (fossil record file L26/f9501) and the Duntroonian–Altonian in South Canterbury (frf J37/f0035). Thus the stratigraphic range of M. browni may be Duntroonian–Waiauan. The material of M. browni in the Waitemata Group strata (including material reworked from underlying Te Kuiti Group strata) may range in age from the Waitakian (latest Oligocene–Early Miocene) to the Otaian.
Kawau Subgroup localities for Magadina browni
The single published locality is Cape Rodney (Dawson Citation1990). Unpublished localities include Kawau Island, Te Kohuroa/Mathesons Bay, Motutapu Island, Hays Stream and Gibsons Beach (, ). This species is common at Kawau Island, but is uncommon to rare at other localities.
Description
There are a small number of moderately well-preserved specimens, but most material is very worn and with matrix attached. The largest specimen is 12 mm long, 11.4 mm wide and 5 mm deep. The outline is elongate oval, the beak is long and the foramen is epithyrid (A–B). The valves are biconvex (C) and the valve exteriors are smooth with distinct growth lines. The deltidial plates are conjunct forming a concave symphytium (A–B, D). The ventral valve is carinate and the anterior commissure is gently sulcate. The teeth are wide and heavy with deep grooves beneath them where the robust socket ridges fit (functionally forming a second set of teeth and sockets) (D). The muscle scars are incised and there is a low septum at the anterior of the ventral valve (D). The socket ridges are large and heavy in the juvenile interior (E), swollen in the adult interior, meeting medially and closing off the hinge trough (F). The cardinalia extend ventrally well into the ventral valve (G) and the sockets are very deep. The cardinal process curves around the posterior end of the small hinge trough. The dorsal septum runs about three-fifths of the valve length (E–G), is low posteriorly and high anteriorly (not fully preserved in this material) where it extends ventrally to meet the very low anterior septum of the ventral valve. The posterior of both valves is heavily thickened but the anterior of the valves are thin.
Figure 4. A–G. Magadina browni. A, B635. Juvenile dorsal exterior. B, B636. Adult dorsal exterior. C, B637. Lateral exterior. D, B638. Ventral interior. E, B639. Juvenile dorsal interior. F, G, B640. F. Adult dorsal interior. G, Oblique view. All scale bars 2 mm. cp, cardinal process; gr, groove; sr, socket ridges; vs, ventral septum. B = University of Auckland School of Environment.

Remarks
Specimens from Gibsons Beach (AU2939) have a soft, white matrix internally and appear to have been reworked from older strata. Hayward and Brook (Citation1984) stated that the ‘complete brachiopods’ from the Otaian Te Akau Member (Papakura Limestone Formation) are commonly reworked from the underlying Te Kuiti Group strata, the Otorohanga Limestone or Waitomo Sandstone; both are Waitakian (Kear & Schofield Citation1959). Some specimens from Kawau Island (AU6662) also have a soft white matrix internally, these specimens may be from the thin shelly limestone beds that occur within the Cape Rodney Formation at Kawau Island (Hayward & Brook Citation1984). Other specimens from Kawau Island (MA81968) have Cape Rodney Formation matrix internally and externally. The posteriorly directed foramen of Magadina suggests a free-living lifestyle with a motile pedicle similar to that of extant Anakinetica cumingi (Davidson, Citation1852) from eastern Australia (Richardson Citation1987) or New Zealand Oligocene species Rhizothyris amygdala Thomson, Citation1920 from North Otago (Mineur & Richardson Citation1984). The very heavy socket ridges that fit into grooves beneath the teeth may be an adaptation to withstand being transported by currents or storms. The extra weight they provide may be an adaptation to keep the shell posterior down and the shell anterior up out of the sediment substrate for feeding and gas exchange. The sediment facies at other localities where Magadina is found include shallow-water limestones (Mount Brown beds, Canterbury, Wilson Citation1963; Balfour Quarry, Southland, Hyden Citation1979) and greensands (Kokoamu Greensand, North Otago, MacKinnon et al. Citation1993) suggesting the preferred habitat of Magadina species is likely to be shallow to mid-shelf depths rather than a shallow rocky shoreline. The material from these shallow to mid-shelf depth South Island localities is much less worn than most of the material from the Kawau Subgroup. This study suggests that some of the Kawau Subgroup material is reworked from older strata, some is coeval, and that specimens collected from rocky-shoreline facies and interbedded shelly limestones may be at the shallow end of their natural depth range.
Genus Pachymagas Ihering, 1903
‘Pachymagas’ sp.
?Magellania parki: Park (Citation1910)
?Terebratella oamarutica: Park (Citation1910)
?Magellania lenticularis: Park (Citation1910)
Pachymagas cf. uttleyi: Hayward (Citation1976b)
Pachymagas sp.: Hayward (Citation1976b)
Liothyrella sp.: Brook (Citation1979, Citation1983)
Thomson (Citation1915a) first placed New Zealand fossil brachiopods into the South American genus Pachymagas. Allan (Citation1949) stated that New Zealand Pachymagas were not closely related to the South American genus and Allan (Citation1957, Citation1960) introduced the term ‘Pachymagas’ for New Zealand fossil material. This study has not attempted to place this material into species because the genus ‘Pachymagas’ and the many named species need major revision. The presence of ‘Pachymagas’ at South Island localities has been mentioned in many publications (Dawson Citation1990) but (as far as the author is aware) only two publications mention North Island localities (Hayward Citation1976b, Waitakere Ranges; Nelson Citation1978, Waitanguru, Pakaumanu and Wilsons Quarry). Park (Citation1910) named three South Island brachiopod species from the Parnell Grit at Cheltenham Beach, his identifications are uncertain but included Magellania parki Hutton, Citation1904, a species that Thomson (Citation1915a) placed into Pachymagas. This study suggests Park may have found some morphologically variable specimens of ‘Pachymagas’.
Stratigraphic range
Duntroonian (Late Oligocene)–Clifdenian? (Middle Miocene) (Fryer Citation1999).
The material attributed to ‘Pachymagas’ in this study area probably ranges from Waitakian to Altonian.
Kawau, Manukau and Hukatere subgroup localities for ‘Pachymagas’
Kawau Subgroup published localities include Gibsons Beach (Hayward & Brook Citation1984) and (perhaps) Cheltenham Beach (Park Citation1910). Unpublished localities include Waiwera Beach and Kawau Island. The single published Manukau Subgroup locality is Walkers Bush (Hayward Citation1976b). One unpublished Hukatere Subgroup locality is Nihotetea Creek, western Hukatere Peninsula (Brook Citation1979, Citation1983) (, ).
Description
The two specimens from Walkers Bush show only the exterior. The outline of the complete (but worn) juvenile (A–B) is sub-oval, biconvex (C) with a sulcate anterior commissure (D) and a shallow dorsal sulcus and ventral fold. The deltidial plates of the juvenile are disjunct and those of the mid-size specimen are conjunct (A, E). The beaks of both specimens are straight and the foramina are submesothyrid. The adult specimens from Gibsons Beach are sub-oval, usually longer than wide (F–H). The beak is sub-erect with sharp beak edges, a mesothyrid foramen and conjunct deltidial plates. The anterior commissure is broadly sulcate, but the dorsal valve has no corresponding sulcus (I). The cardinalia of the mid-sized partial dorsal valve from Kawau Island has a small cardinal process, slim socket ridges angled steeply to form a septalium, a high septum and the beginning of posterior shell thickening (J). MicroCT scans of the mid-size partial specimen (E) also show a long septum, steeply angled septalium and a cardinal process that extends into the hinge trough (K–N). The cardinalia of a partial adult dorsal valve shows extensive thickening of the socket ridges, filling in the septalium, widening of the septum and heavy thickening on either side of the cardinalia, creating deep muscle scars and the cardinal process is enlarged but broken off (O).
Figure 5. A–N, ‘Pachymagas’. A–D, MA91060a. Juvenile specimen. A, Dorsal exterior. B, Ventral exterior. C, Lateral exterior. D, Anterior commissure. E, MA91060b. Mid-sized specimen. Dorsal exterior. F–I, B641. Adult specimen. F, Dorsal exterior. G, Ventral exterior. H, Lateral exterior. I, Sulcate anterior commissure. J, B642. Cardinalia of partial dorsal valve of mid-sized specimen. K–N, Images from CT scan of MA91060b. Figure K is the most dorsal, L–N are progressively more ventral. O, B643. Swollen cardinalia of adult specimen. P–R, OU54765a‐c. Terebratellidae indet. P, Whole juvenile specimen. Q, Partial ventral valve. R, Partial dorsal valve. All scale bars 5 mm unless indicated otherwise. cp, cardinal process; se, septum; slm, septalium; sr, socket ridge; thsr, thickened socket ridge. B = University of Auckland School of Environment, MA = Auckland Museum, OU = University of Otago Geology Museum, Dunedin.
Remarks
Specimens of ‘Pachymagas’ sp. are common in the Otaian Te Akau Member (Papakura Limestone Formation) at Gibsons Beach (). However, they often have a white matrix internally and are most likely reworked from underlying Waitakian age Te Kuiti Group strata (Hayward & Brook Citation1984). Specimens of ‘Pachymagas’ are rare in the Cape Rodney Formation, the only known locality is Kawau Island, where three partial valves, one including the cardinalia (J), have been collected. Specimens are rare in the ‘lateral equivalent’ of the Pakaurangi Formation, western Hukatere Peninsula, one small worn specimen and a single dorsal valve exterior (originally identified as Liothyrella in Brook Citation1979, Citation1983) are held by the University of Auckland (). Extant genus Neothyris is considered to be descended from Early Miocene ‘Pachymagas’ (Neall Citation1972).
Terebratellidae indet.
Terebratellidae indet.: Hiller (2011)
Hiller (Citation2011) described and figured specimens as Terebratellidae indet. from the Hukatere Peninsula and five localities in Northland, and noted that this was the most common Otaian brachiopod in that study with ‘> 460’ specimens (mostly very small juveniles) having been collected from the Pakaurangi Formation (Waiteroa Member) at Hollands Point. The larger specimens (up to 8 mm) collected by Hiller (Citation2011, figure 8D) were just beginning to develop costae and Hiller (Citation2011, p. 83) noted that ‘the largest specimens bear a close resemblance to the Pleistocene–Holocene terebratellid Calloria inconspicua’. Jones (Citation1970, table 1) listed ‘cf. Waltonia’ [= Calloria] as being present in the Pakaurangi Formation. The author found four complete (but poorly preserved) juvenile specimens (the largest was 3.5 mm long) and five partial valves in the Pakaurangi Formation (Hollands Member) that appear to be the same as Hiller’s material. They are too small to have any clearly diagnostic features. As noted above, smooth specimens of Magasella neozelandica in the Kawau Subgroup resemble Holocene Calloria inconspicua. It may be that all these Waitakere Group Terebratellidae specimens are juvenile to young M. neozelandica or they may be a (perhaps) smaller and more delicate unknown species.
Description
The specimens are elongate oval, maximum width at about two-thirds of valve length. The dorsal valve exterior shows a slight sulcus (P). The foramen is wide with thin, disjunct deltidial plates, the teeth lack dental plates (Q). The dorsal valve interior has a transverse cardinal process, the inner hinge plates slope medially and attach to the septum forming a septalium and the dorsal septum is almost half of the valve length (R).
Paleotemperature and paleoecology
The presence of large reef corals in the Kawau Subgroup at localities Hays Stream (Eagle & Hayward Citation1992) and Motuketeke Island (Campbell et al. Citation2004) indicate subtropical to marginally tropical sea temperatures. Based on the presence of corals, Squires (Citation1962) suggested a mean temperature range of 19–28 °C and Hayward (Citation1977) suggested a range of 18–27 °C.
The Cape Rodney Formation includes rocky-shore, gravel and inner-shelf sand communities (Eagle et al. Citation1994). Eagle et al. (Citation1994) described fossil faunas from Kawau Island and listed taxa with extant relatives that live on rocky shorelines including Cellana (limpet), Sarmaturbo, Conus, Conilithes, Haliotis and Cookia (gastropods), Goniocidaris, Histocidaris and Phyllacanthus (echinoids), Alveopora, Leptastrea and Oculina (hermatypic corals), suggesting a high-energy, rocky-bottom, shallow-water environment, at an inferred depth of 0–20 m. In the Cape Rodney Formation, Notosaria antipoda and Magasella neozelandica are common to abundant, Magadina browni is common at some localities, Liothyrella gravida and ‘Pachymagas’ are rare; the latter three genera are shallow to mid-shelf dwellers. At three localities at Waiheke Island, N. antipoda and T. suessi are uncommon to rare in the shallow-water shelly gravels, coarse sands and muddy sands of the Tipakura Sandstone Formation. Eagle et al. (Citation1995) suggested paleodepths of 0–2, 0–10 and 10–20 m. The wide ranging lithologies and associated faunas of the various members of the Papakura Limestone Formation suggest that they formed at shallow to mid- to outer-shelf depths (Eagle & Hayward Citation1992; Campbell et al. 1994; Eagle et al. Citation1995). Notosaria antipoda and M. neozelandica may be common, whereas L. gravida and T. suessi are uncommon to rare. Jones (Citation1970) discussed the paleoecology and paleoenvironment of the rich and diverse Pakaurangi Point fossil assemblages (Pakaurangi Formation) in detail and concluded that the fossil assemblages were living at depths ranging from a few metres down to perhaps 250 m, and that surface temperatures were subtropical 20–27 °C. The only common brachiopod from these strata is the material identified as Terebratellidae indet. (Hiller Citation2011). Rare genera include Liothyrella, Thecidellina, Terebratulina, Argyrotheca and ‘Pachymagas’. The slumped volcaniclastic sandstones and gravels of the Nihotupu Formation contain faunas that lived at depths ranging from 10 to 100 m (Hayward Citation1976b); Liothyrella, (possibly) Magasella and ‘Pachymagas’ are rare.
Discussion
Faunal provinces
Beu (Citation1966, ) suggested that the New Zealand sea temperature peaked at marginally tropical in the Otaian (based on fossil molluscs). Beu and Maxwell (Citation1968, p. 73) suggested that the sea temperatures in Northland were ‘marginally tropical from at least Bortonian to Waiauan times’ (based on fossil molluscs). Hornibrook (Citation1968) suggested that Northland was probably marginally tropical from the Otaian to the Waiauan (based on foraminifera). Keyes (Citation1968, ) suggested that there was a short peak during the Otaian when sea temperatures reached tropical conditions (above 25 °C) based on scleractinian corals. Hornibrook (Citation1992) reviewed all the literature on the Cenozoic paleoclimate of New Zealand, including publications on foraminifera, Mollusca, the brachiopod genus Lingula Bruguière, Citation1791, bryozoans, corals, diatoms, the trees Nothofagus and Cocos, pollens and oxygen isotopes and stated that the temperatures in New Zealand reached ‘a peak of warmth in the upper Altonian and Clifdenian stages’ (p. 83) and that ‘A warm subtropical or possibly marginally tropical sea … .surrounded the whole country with the possible exception of the Canterbury Basin’ (p. 96) in the Early Miocene.
Hiller (Citation2011) described a fauna of micromorphic brachiopods from the marginally tropical Otaian of Northland, noted that most of the genera present (Novocrania Lee & Brunton, Citation2001, Lacazella Munier-Chalmas, Citation1880, Notozyga Cooper, Citation1977, Thecidellina, Argyrotheca and Megerlina) are not found in the warm–temperate Otaian brachiopod faunas of Canterbury and Otago and suggested that ‘The brachiopod faunas therefore appear to support the notion of distinct faunal provinces within the Early Miocene of the New Zealand region’ (p. 87). However, all the Waitemata and Waitakere group strata of Auckland and lower Northland discussed here that contain brachiopods, and those from Northland of Hiller (Citation2011), lie within the Cenozoic Northland Basin (Hayward Citation1993). Beu (Citation1966) stated of the molluscs from Pakaurangi Point that ‘At Pakaurangi occur the warmest known elements of our [New Zealand] fauna’ (p. 177) and that ‘it seems likely that the 70 °F [21.6 °C] isotherm, a convenient biological boundary for the tropical zone, lay across New Zealand in about the vicinity of Rotorua during the Otaian time’ (p. 183). This suggests that the brachiopod faunas of the Waitemata and Waitakere groups must be included in the marginally tropical faunal province of Early Miocene Northland and that the genera Notosaria, Liothyrella, Terebratulina, Magasella, Magadina and ‘Pachymagas’ occurred in both provinces at the same time. All six micromorphic genera from Northland in Hiller (Citation2011) listed above have extant species, but only Thecidellina is restricted to subtropical–tropical waters (Lee & Robinson Citation2003). This study suggests that brachiopods may not be a good indicator of faunal provinces in the Miocene of New Zealand.
Three of the New Zealand genera (Notosaria, Liothyrella and Magasella) that lived in marginally tropical conditions in the Otaian of Auckland–lower Northland now live in cool–temperate waters around southern New Zealand and Liothyrella is widespread in the cold waters around Antarctica. This study suggests that New Zealand brachiopod genera appear to be very adaptable to changes in water temperature and that, more generally, many brachiopod genera are not reliable indicators of paleotemperature. The exceptions, in New Zealand, may be the genera Thecidellina (Lee & Robinson Citation2003) and Lingula (Lee & Campbell Citation1987).
Rocky-shoreline faunas and South Island Early Miocene faunas
Fossil brachiopod faunas from shallow rocky-shore communities are uncommon in the fossil record because this environment is not conducive to the accumulation of fossiliferous deposits. Three publications list brachiopods from other New Zealand Late Oligocene–Early Miocene rocky-shore communities. Lee et al. (Citation2014) documented a very rich and diverse Late Duntroonian subtropical fauna from Cosy Dell farm, Southland, Lee et al. (Citation1983) briefly described a Duntroonian–Waitakian fauna from Mount Luxmore, Fiordland and Bosel and Coombs (Citation1984) briefly described a Waitakian–Otaian ‘warm-water’ fauna from Bluff. All three localities have elements in common with the fauna of the Cape Rodney Formation. Notosaria antipoda is present at all localities, and possibly Magasella, although only indeterminate juvenile valves (Lee et al. Citation2014, figure 7K) and fragments of larger ribbed valves (unpublished) occur at Cosy Dell. The Cosy Dell fauna includes micromorphic genera Discinisca and Megerlia and mostly fragmentary Novocrania not seen at the other localities (note that in Lee et al. Citation2014, figure 7L, some specimens of Megerlia from Cosy Dell were identified as Argyrotheca). However, this may be due to the coarse cemented facies of the Mount Luxmore, Bluff and Cape Rodney deposits (Cosy Dell has uncemented sandy horizons with excellent preservation of microfossils).
It is difficult to directly compare the Early Miocene brachiopod faunas of the Waitemata and Waitakere groups with South Island faunas of the same age because the facies are quite different. Whereas the brachiopod-rich strata of the Waitemata and Waitakere groups are generally coarse grained and the faunas were mostly living in water depths of 20 m or less, the Early Miocene South Island brachiopod faunas are found in finer grained shallow to mid-shelf sandy limestones and greensands. There are some similar associations of genera, Notosaria and Magasella occur together in the limestone at Otaian locality Blands Bluff, Canterbury (Marwick Citation1938). Magadina and ‘Pachymagas’ appear to have coexisted in the Cape Rodney Formation shelly limestones of Kawau Island, these genera coexist (along with Waiparia and Rhizothyris) in the Mount Brown limestone, Canterbury. A point of difference is that the most common brachiopods in the Waitemata Group strata are the attached genera Notosaria and Magasella, whereas the most common brachiopods at many Early Miocene South Island localities are genera that are free-living or potentially free-living; Aetheia Thomson, Citation1915c, Magadina, Waiparia, Rhizothyris and ‘Pachymagas’. A reflection of the water depth, turbulence and substrate of the various communities.
Conclusions
This study makes a contribution to the poorly documented brachiopod faunas of the North Island. Seven genera of brachiopods, from five families, are described and illustrated in detail for the first time from the Waitemata and Waitakere groups of Auckland and lower Northland: Notosaria, Thecidellina, Liothyrella, Terebratulina, Magasella, Magadina and ‘Pachymagas’. Many publications mention the presence of brachiopods in these strata and the large number of published synonymous names is listed here. For the first time, the type specimens of the species Liothyrella gravida and Magasella neozelandica are figured, Magadina browni and ‘Pachymagas’ are described and figured from North Island strata, and Terebratulina suessi is described and figured from the Waitemata and Waitakere groups. There are possibly many synonymous species names for New Zealand fossil Liothyrella and because L. gravida is the oldest valid name, its true stratigraphic and geographic range may be extensive. Magasella neozelandica is confirmed as a separate species based on morphology and paleoecology and its possible presence in two Late Oligocene to Early Miocene rocky-shoreline deposits in Southland and Fiordland is noted. The Otaian range of Magadina browni is extended northwards to Auckland, the stratigraphic range of this species may be Duntroonian–Waiauan, and the name M. waiparaensis may be a junior synonym. Many Cenozoic brachiopod genera in New Zealand are in need of taxonomic review and this study makes a contribution to the review process of Liothyrella, Magasella, Magadina and ‘Pachymagas’. The brachiopod fauna described here inhabited the margins of a subsiding marine basin in the Auckland and lower Northland area for c. 5 Ma in marginally tropical conditions in the Early Miocene. Six brachiopod genera occupied both the marginally tropical faunal province of Northland and the warm–temperate faunal province of Canterbury–Otago. Many of the brachiopods living in the Cenozoic Northland Basin during the Early Miocene have extant relatives; among these only Thecidellina is restricted to subtropical–tropical waters. Brachiopods may not be a reliable indicator of faunal provinces in the Miocene of New Zealand.
Acknowledgements
I would like to thank Daphne Lee, Norton Hiller, Bruce Hayward, Kathleen Campbell, Jack Grant-Mackie and Donald Macfarlan whose suggestions greatly improved this manuscript; Luke Easterbrook for technical support; Heidi Schlumpf and Severine Hannam of the Auckland Museum; Neville Hudson of the School of Environment, University of Auckland, and Bruce Marshall of Te Papa Tongarewa, the National Museum of New Zealand, Wellington, for their help in examining the collections and for the loan of materials; Louise Cotterall of the Department of Environment, University of Auckland, for help with photographing the material; Andreas Kroh and Alice Schumacher of the Naturhistorisches Museum Vienna, Austria, for providing images of the type specimens; Andrew McNaughton of the Otago Centre of Microscopy for the micro-CT scans; and Laurie Mahmood for taking me to and from Gibsons Beach. Associate Editor: Professor Kathy Campbell.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
References
- Allan RS. 1931. On a new recent Terebratella from northern New Zealand. NZ J Sci Tech (Wellington). 12(6):385–387.
- Allan RS. 1932. Tertiary Brachiopoda from the Chatham Islands. Trans Proc N Z Inst. 63(1):11–23.
- Allan RS. 1949. Notes on a comparison of the Tertiary and Recent Brachiopoda of New Zealand and South America. Trans Proc R Soc N Z. 77(5):288–289.
- Allan RS. 1957. Appendix 3: brachiopod faunas. In: Gage M, editor. Geology of the waitaki subdivision. New Zealand Geological Survey Bulletin 55. Dunedin: John Mcindoe Ltd.; p. 104–107.
- Allan RS. 1960. The succession of Tertiary brachiopod faunas in New Zealand. Rec Canterbury Mus. 7(3):233–268.
- Ballance PF. 1976. Stratigraphy and bibliography of the Waitemata Group of Auckland, New Zealand. N Z J Geol Geophys. 19(6):897–932. doi: 10.1080/00288306.1976.10420746
- Beu AG. 1966. Sea temperatures in New Zealand during the Cenozoic Era, as indicated by molluscs. Trans R Soc N Z. 4(9):177–187.
- Beu AG, Maxwell PA. 1968. Molluscan evidence for Tertiary sea temperatures in New Zealand: a reconsideration. Tuatara. 16(1):68–74.
- Beu AG, Nolden S, Darragh TA. 2012. Revision of New Zealand Cenozoic fossil Mollusca described by Zittel (1865) based on Hochstetter’s collections from the “Novara” Expedition. Mem Assoc Australas Palaeontologists. 43:1–69.
- Bitner MA. 2005. Recent brachiopods from the Fiji and Marquesas Islands, Southern Pacific, collected during the French cruises MUSORSTOM 9, 10 and BORDAU 1. In: Harper DAT, Long SL, McCorry M, editors. Fifth international brachiopod congress. Copenhagen: Abstracts; p. 35–36.
- Bitner MA. 2007. Recent brachiopods from the Austral Islands, French Polynesia, Couth-Central Pacific. Zoosystema. 29(3):491–502.
- Bitner MA. 2009. Recent Brachiopoda from the Norfolk Ridge, New Caledonia, with description of four new species. Zootaxa. 2235:1–39.
- Bitner MA. 2010. Biodiversity of shallow-water brachiopods from New Caledonia, SW Pacific, with description of a new species. Scientia Marina. 74(4):643–657. doi: 10.3989/scimar.2010.74n4643
- Bitner MA. 2014. Living brachiopods from French Polynesia, Central Pacific, with descriptions of two new species. Pac Sci. 68(2):245–265. doi: 10.2984/68.2.6
- Bosel CA, Coombs DS. 1984. Foveaux formation: a warm-water, strandline deposit of Landon-Pareora age at Bluff Hill, Southland, New Zealand. N Z J Geol Geophys. 27(2):221–223. doi: 10.1080/00288306.1984.10422529
- Brook FJ. 1979. Lower Miocene geology of western Hukatere Peninsula [Unpublished Honours thesis]. New Zealand: University of Auckland.
- Brook FJ. 1983. Lower Miocene geology of North Kaipara, Northern New Zealnd, with an emphasis on the Paleontology [Unpublished PhD thesis]. New Zealand: University of Auckland.
- Brook FJ. 1998. Stratigraphy and paleontology of Pleistocene submarine volcanic-sedimentary sequences at the northern Kermadec Islands. J R Soc N Z. 28(2):235–257. doi: 10.1080/03014223.1998.9517561
- Bruguière JG. 1791. Tableau Encyclopédique et Méthodique des trois Règnes de la Nature: vers, coquilles, mollusques et polypes divers. Paris, Panckoucke. Tome 1. Pp. 172–344.
- Buchanan J. 1870. On the Wanganui Beds (Upper Tertiary). Trans Proc R Soc N Z. 2:163–166.
- Campbell KA, Grant-Mackie JA, Buckeridge JS, Hudson N, Alfaro AC, Hoverd J, Morgan S, Horne N, Banfield A. 2004. Paleoecology of an early Miocene, rapidly submerging rocky shore, Motuketekete Island, Hauraki Gulf, New Zealand. N Z J Geol Geophys. 47:731–748. doi: 10.1080/00288306.2004.9515086
- Carter RM. 1972. Wanganui strata of Komako District, Pohangina Valley, Ruahine Range, Manawatu. J R Soc N Z. 2(3):293–324. doi: 10.1080/03036758.1972.10421819
- Clarke E. 1905. The Fossils of the Waitemata and Papakura Series. Trans Proc N Z Inst. 37:413–421.
- Cohen BL, Bitner MA, Harper EM, Lee DE, Mutschke E, Sellanes J. 2011. Vicariance and convergence in Magellanic and New Zealand long-looped brachiopod clades (Pan-Brachiopoda: Terebratelloidea). Zool J Linn Soc. 162:631–645. doi: 10.1111/j.1096-3642.2010.00682.x
- Cooper GA. 1954. Recent Brachiopods, Bikini and nearby atolls, Marshall Islands. Geological Survey Professional Paper, Volume 260-G:315–318.
- Cooper GA. 1959. Genera of Tertiary and recent Rhynchonelloid Brachiopods. Smithsonian Miscellaneous Collections. 139 (5). Smithsonian Institution Press, Washington, DC. 90 p.
- Cooper GA. 1964. Brachiopods from Eniwetok and Bikini drill holes. Bikini and nearby atolls, Marshall Islands. Geological Survey Professional Paper 260-FF:1117–1120.
- Cooper GA. 1977. Brachiopods from the Caribbean Sea and adjacent waters. Studies in tropical oceanography 14. Miami: University of Miami Press. 212 p.
- Cooper GA. 1978. Tertiary and quarternary brachiopods from the Southwest Pacific. Smithsonian Contributions to Paleobiology. 43:1–93. doi: 10.5479/si.00810266.38.1
- Cooper GA, Doherty PJ. 1993. Calloria variegata, a new recent species of brachiopod (Articulata: Terebratulida) from northern New Zealand. J R Soc N Z. 23(3):271–281. doi: 10.1080/03036758.1993.10721225
- Cooper GA, Lee DE. 1993. Calloria, a replacement name for the recent brachiopod genus Waltonia from New Zealand. J R Soc N Z. 23(3):257–270. doi: 10.1080/03036758.1993.10721224
- Cox SH. 1881. Geology of the Rodney and Marsden Counties. Colonial Museum and Geological Survey of New Zealand. Rep Geol Explorations. 13:13–39.
- Cox SH. 1882. North Auckland district, including Thames, Coromandel, Island of Kawau, and Drury coal field. Colonial Museum and Geological Survey of New Zealand. Rep Geol Explorations. 14:17–41.
- Dall WH. 1870. A revision of the Terebratulidae and Lingulidae, with remarks on and descriptions of some Recent forms. A J Conchology. New York. 6(2):88–168.
- Dall WH. 1900. Some names which must be discarded. Nautilus (Havertown). 14:44–45.
- Davidson T. 1850. On the genus Waltonia. Annals and Magazine of Natural History, Series. 2. 5:474–476.
- Davidson T. 1852. Description of a few new species of Brachiopoda. Proc Zool Soc (London). 20:75–83.
- Dawson EW. 1990. The Cenozoic brachiopoda of New Zealand: A commentary, reference list and bibliography. Miscellaneous Publication 103. Wellington: New Zealand Oceanographic Institute. 99 p.
- Douville H. 1879. Note sur quelques genres de Brachiopodes (Terebratulidae et Waldheimidae). Bulletin de la Société géologique de France, Series 3. 7:251–277.
- Eagle MK, Hayward BW. 1992. Paleontology and paleoecology of Early Miocene sequences in Hays and Tipakuri Streams, northern Hunua Ranges, Auckland. Rec Auckland Inst Mus. 31:187–204.
- Eagle MK, Hayward BW, Carter G. 1994. Early Miocene rocky shore and coarse sediment fossil communities, Kawau Island, Auckland. Rec Auckland Inst Mus. 31:187–204.
- Eagle MK, Hayward BW, Grant-Mackie JA. 1995. Early Miocene beach, rocky shore, and enclosed bay fossil communities, Waiheke Island. Rec Auckland Inst Mus. 32:17–44.
- Eagle MK, Hayward BW, Grant-Mackie JA, Gregory MR. 1999. Fossil communities in an early Miocene transgressive sequence, Mathesons Bay, Leigh, Auckland. Tane. 37:43–67.
- Elliot GF. 1958. Classification of thecidean brachiopods. J Paleontology (Lawrence). 32(2):373–000.
- Firth CW. 1929. Geology of the north-west portion of Manukau County, Auckland. Trans Proc N Z Inst. 61:85–137.
- Flügel E. 1959. Statement concerning the types and figured originals from the collections of the Novara Expedition in the custody of Geological-Paleontological section of the Museum of Natural History, Austria. N Z J Geol Geophys. 2(5):826–845. doi: 10.1080/00288306.1959.10422752
- Fox CE. 1902. The volcanic beds of the Waitemata Series. Trans Proc N Z Inst. 34:452–493.
- Fryer S. 1999. Systematics and evolution of the brachiopods ‘Pachymagas’ and Waiparia in New Zealand. Unpublished MSc Thesis, University of Canterbury, New Zealand. 260 p.
- Gmelin JF. 1790. Systema Naturae, 13th ed. Leipzig: Vermes. 88 p.
- Grant-Mackie JA. 1965. Ancient Life in the Auckland Area. In: Kermode LO, editor. Science in Auckland. 11th New Zealand Science Congress Auckland. Royal Society of New Zealand Handbook; p. 20–27.
- Grant-Mackie JA, Chapman-Smith M. 1971. Paleontological notes on the Castlecliffian Te Piki Bed, with descriptions of new mollusc taxa. N Z J Geol Geophys. 14(4):655–704. doi: 10.1080/00288306.1971.10426328
- Grant-Mackie JA, Moore PR. 1970. Correlation of the Colville Formation (Letter to the Editor). N Z J Geol Geophys. 13(2):561–563. doi: 10.1080/00288306.1970.10428915
- Gray JE. 1840. Synopsis of the contents of the British Museum. 42nd ed. London: G. Woodfall. 151 p.
- Hayasaka I. 1938. A new neotreme genus of Brachiopoda from Japan. Venus. 8:9–13.
- Hayward BW. 1976a. Lower Miocene stratigraphy and structure of the Waitakere Ranges and the Waitakere Group (new). N Z J Geol Geophys. 19(6):871–895. doi: 10.1080/00288306.1976.10420745
- Hayward BW. 1976b. Macropaleontology and paleoecology of the Waitakere Group (lower Miocene), Waitakere Hills, Auckland. Tane. 22:177–206.
- Hayward BW. 1977. Lower Miocene corals from the Waitakere Ranges, North Auckland, New Zealand. J R Soc N Z. 7(1):99–111. doi: 10.1080/03036758.1977.10419340
- Hayward BW. 1979. Eruptive history of the early to mid Miocene Waitakere volcanic arc, and palaeogeography of the Waitemata Basin, Northern New Zealand. J R Soc N Z. 9(3):297–320. doi: 10.1080/03036758.1979.10419410
- Hayward BW. 1993. The tempestuous 10 million year life of a double arc and intra-arc basin – New Zealand’s Northland Basin in the early Miocene. In: Balance PF, editor. South Pacific Sedimentary Basins. Sedimentary Basins of the World 2. Amsterdam: Elsevier Science Publishers; p. 113–142.
- Hayward BW, Brook FJ. 1984. Lithostratigraphy of the basal Waitemata Group, Kawau Subgroup (new), Auckland, New Zealand. N Z J Geol Geophys. 27:101–123. doi: 10.1080/00288306.1984.10422521
- Hedley C. 1899. Mollusca of Funafuti, Part 2, Pelecypoda and Brachiopoda. Memoirs of the Australian Museum. 3(80):508–510.
- Hiller N. 2000. Provincialism in New Zealand Early Miocene brachiopods? (Abstract). The Millenium Brachiopod Congress. London: The Natural History Museum. p. 43.
- Hiller N. 2011. Micromorphic brachiopods from the Early Miocene (Otaian) of Northland, New Zealand. N Z J Geol Geophys. 54(1):75–87. doi: 10.1080/00288306.2011.537608
- Hiller N, MacKinnon DI, Nielsen SN. 2008b. A review of the systematics, biogeography, and evolutionary relationships of Recent and fossil brachiopods of the Superfamily Kraussinoidea Dall, with descriptions of two new fossil species from New Zealand and Chile. Earth and Environmental Science Transactions of the Royal Society of Edinburgh. 98:379–390. doi: 10.1017/S1755691007078474
- Hiller N, Robinson JH, Lee DE. 2008a. The micromorphic brachiopod Argyrotheca (Terebratulida, Megathyridoidea) in Australia and New Zealand. Proc R Soc Victoria. 120(1):167–183.
- Hills DA. 1971. New species of Terebratulina (Brachiopoda) from the Lower Miocene, North Canterbury. N Z J Geol Geophys. 14(2):284–287. doi: 10.1080/00288306.1971.10421925
- von Hochstetter F. 1864. Geologie von Neu-Seeland. Beiträge zur Geologie der Provinzen Auckland und Nelson. Reise der Österreichischen Fregatte Novara um die Erde in den Jahren 1857, 1858, 1859 unter der Befehlen der Commodore B. von Wüllerstorf-Urbair. Geologischer Theil. 1(1), 1–273. [translation: Fleming 1959a].
- von Hochstetter F. 1867. New Zealand. Its physical geography, geology and natural history with special reference to the results of government expeditions in the provinces of Auckland and Nelson. Translated from the German original published in 1863 by Sauter E. Stuttgart, JG Cotta. 515 p.
- von Hochstetter F, Hörnes M, von Hauer F, editors. 1865. Paläontologie von Neu-Seeland. Beiträge zur Kenntniss der fossilien Flora und Fauna der Provinzen Auckland und Nelson, von Prof. Dr. F. Unger, Prof. Dr. Karl Zittel, Prof. E. Suess, Felix Karrer, Dr. Ferdinand Stoliczka, Dr. Guido Stache, Dr. Gustav Jaeger. Reise der Österreichischen Fregatte Novara um die Erde in den Jahren 1857, 1858, 1859 unter der Befehlen der Commodore B. von Wüllerstorf-Urbair, Geologischer Theil. 1(2), i-vii, 1–318, 26 pls [27 April 1865; Higgins 1963].
- Hoffmann J, Klann M, Matz F. 2009. Recent thecideide brachiopods from the northern Great Barrier Reef, Australia (SW Pacific Ocean). Zoosystematics and Evolution. 85(2):341–349. doi: 10.1002/zoos.200900010
- Hornibrook NdeB. 1968. Distribution of some warm water benthic Foraminifera in the New Zealand Tertiary. Tuatara. 16(1):11–15.
- Hornibrook NdeB. 1992. New Zealand Cenozoic marine paleoclimates: A review based on the distribution of some shallow water and terrestrial biota. In: Tsuchi R, Ingle JC, editor. Pacific Neogene: Environment, evolution and events. Tokyo: University of Tokyo Press; p. 83–106.
- Hutton FW. 1870. On the relative ages of the Waitemata Series and the Brown Coal Series of Drury and Waikato. Trans Proc N Z Inst. 3:244–251.
- Hutton FW. 1873. Catalogue of Tertiary Mollusca and Echinodermata of New Zealand in the collection of the Colonial Museum. Wellington: Government Printer. 48 p.
- Hutton FW. 1904. Revisions of the Tertiary Brachiopoda of New Zealand. Trans Proc N Z Inst. 37:474–481.
- Hyden FM. 1979. Mid-Tertiary temperate shelf bioclastic limestones, Southland, New Zealand. Unpublished PhD thesis. University of Otago, Dunedin, New Zealand. 487 p.
- Ihering H. 1903. Les Brachiopodes Tertiares de Patagonie. Anales del Museo Nacional de Buenos Aires. 9:32–394.
- Isaac MJ, Herzer RH, Brook FJ, Hayward BW. 1994. Cretaceous and Cenozoic sedimentary basins of Northland. Lower Hutt, New Zealand: Institute of Geological and Nuclear Sciences Monograph. 8. 203 p.
- Jones BG. 1969. The stratigraphy and structure of Pakaurangi Point, Kaipara, New Zealand. Trans R Soc N Z. 6(17):219–246.
- Jones BG. 1970. Paleontology and paleoecology of Pakaurangi Point, Kaipara, New Zealand. Trans R Soc N Z Earth Sci. 7:137–176.
- Jones TR. 1860. Notes on the fossils; notes on the plates XII and XIII. In: Heaphy C. On the volcanic country of Auckland, New Zealand. Geol Soc Lond Q J. 16:251–252. doi: 10.1144/GSL.QJEG.1983.016.03.12
- Kear D, Schofield JC. 1959. Te Kuiti group. N Z J Geol Geophys. 2(4):685–717. doi: 10.1080/00288306.1959.10422764
- Keyes IW. 1968. Cenozoic marine temperatures indicated by the scleractinian coral fauna of New Zealand. Tuatara. 16(1):21–25.
- King W. 1850. A Monograph of the Permian fossils of England. Palaeontographical Soc (London), Monograph. 3(1):1–258.
- Kuhn. 1949. Lerbuch der Palaeozoologie. Schweizerbart'sche Verlagsbuchhandlung. Stuttgart. 326 p.
- Laurin B. 1997. Brachiopodes récoltés dans les eaux de la Nouvelle-Calédonie et des Îles Loyauté, Matthew et Chesterfield. In: Crosnier A, editor. Résultats des Campagnes MUSORSTOM 18. Mémoires du Muséum national d’Histoire naturelle. 176:411–471.
- Laws CR. 1931. Geology of the Papakura-Hunua District, Franklin County, Auckland. Trans Proc N Z Inst. 62:37–66.
- Leach WE. 1814. The Zoological Miscellany. Volume 1 and Atlas. London: McMillan. 144 p.
- Lee DE. 1978. Aspects of the ecology and paleoecology of Notosaria nigricans (Sowerby). J R Soc N Z. 8(4):395–417. doi: 10.1080/03036758.1978.10423321
- Lee DE. 1980. Cenozoic and Recent Rhynchonellide brachiopods of New Zealand: Systematics and variation in the genus Tegulorhynchia. J R Soc N Z. 10(3):223–245. doi: 10.1080/03036758.1980.10415330
- Lee DE, Brunton CHC. 2001. Novocrania, new name for the genus Neocrania Lee & Brunton, 1986 (Brachiopoda, Craniida); preoccupied by Neocrania Davis, 1978 (Insecta, Lepidoptera). Bulletin of the Natural History Museum (Geology Series). 57(1):5.
- Lee DE, Campbell JD. 1987. Cenozoic records of the genus Lingula (Brachiopoda: Inarticulata) in New Zealand. J R Soc N Z. 17(1):17–30. doi: 10.1080/03036758.1987.10421705
- Lee DE, Carter RM, King RP, Cooper AF. 1983. An Oligocene rocky shore community from Mt Luxmore, Fiordland (Note). N Z J Geol Geophys. 26:123–126. doi: 10.1080/00288306.1983.10421529
- Lee DE, Lindqvist JK, Beu AG, Robinson JH, Ayress MA, Morgans HEG, Stein JK. 2014. Geological setting and diverse fauna of a Late Oligocene rocky shore ecosystem, Cosy Dell, Southland. N Z J Geol Geophys. 57(2):195–208. doi: 10.1080/00288306.2014.898666
- Lee DE, Robinson JH. 2003. Kakanuiella (gen. nov.) and Thecidellina: Cenozoic and recent thecideide brachiopods from New Zealand. J R Soc N Z. 33:341–361. doi: 10.1080/03014223.2003.9517734
- Lee DE, Wilson JB. 1979. Cenozoic and recent Rhychonellide brachiopods of New Zealand: Systematics and variation in the genus Notosaria. J R Soc N Z. 9(4):437–463. doi: 10.1080/03036758.1979.10421831
- MacKay A. 1884. On the relations of the Tertiary and Cretaceo-Tertiary strata on the coastline between Auckland and Mahurangi. Colonial Museum and Geological Survey of New Zealand. Rep Geol Explorations. 16:101–106.
- MacKinnon DI, Beus SS, Lee DE. 1993. Brachiopod fauna of the Kokoamu Greensand (Oligocene), New Zealand. N Z J Geol Geophys. 36(3):327–347. doi: 10.1080/00288306.1993.9514579
- Mancenido MO, Owen EF. 2002. Notosaridae. In: Kaesler RL, editor. Treatise on Invertebrate Paleontology, Part H, Brachiopoda Revised, Volume 4: Rhynchonelliformea (part). Boulder, Colorado: Geological Society of America & University of Kansas Press; p. 1367.
- Marwick J. 1938. Paleontology. Lists of fossils – brachiopods. In: Speight R, editor. The geology of the Mount Somers district. Wellington: Department of Scientific and Industrial Research 3; p. 60–63.
- Mineur RJ, Richardson JR. 1984. Free and mobile brachiopods from New Zealand Oligocene deposits and Australian waters. Alcheringa: An Australasian Journal of Palaeontology. 8(4):327–334. doi: 10.1080/03115518408618954
- Munier-Chalmas EP. 1880. Note sommaire sur les genres de la famille des Thecideidae. Bulletin de la Société Géologique de France, Series 3. 8:279–280.
- Neall VE. 1970. Notes on the ecology and paleoecology of Neothyris, an endemic New Zealand brachiopod. New Zealand Journal of Marine and Freshwater Research. 4(2):117–125. doi: 10.1080/00288330.1970.9515332
- Neall VE. 1972. Systematics of the endemic New Zealand brachiopod Neothyris. J R Soc N Z. 2(2):229–247. doi: 10.1080/03036758.1972.10429376
- Nelson CS. 1978. Stratigraphy and paleontology of the Oligocene Te Kuiti group, Waitomo County, South Auckland, New Zealand. N Z J Geol Geophys. 21(5):553–594. doi: 10.1080/00288306.1978.10424086
- Newman J, Bradshaw JD. 1981. Oligocene-Miocene rocks of the Brechin Burn outlier, Waimakariri Valley, Canterbury, New Zealand. J Geol Geophys. 24(4):469–476. doi: 10.1080/00288306.1981.10422738
- Orbigny A. 1847. Considerations zoologiques et geologiques sur les brachiopodes ou palliobranches. Comptes Rendus Hebdomadaires des Seances de l'Academie des Sciences (Paris). 25(5):193–195, 25(7):266–269; Annales des Sciences Naturelles, Series 3. 8:241–270.
- Park J. 1886a. Waitemata, Eden and Manukau Counties. Colonial Museum and Geological Survey of New Zealand. Rep Geol Explorations. 17:147–164.
- Park J. 1886b. Waikato and Raglan Counties. Colonial Museum and Geological Survey of New Zealand. Rep Geol Explorations. 17:136–147.
- Park J. 1889. On the conformable relations of the different members of the Waitemata Series. Trans Proc N Z Inst. 22:391–399.
- Park J. 1910. The Geology of New Zealand. Wellington: Whitcombe & Tombs Ltd. 488 p.
- Raine JI, Beu AG, Boyes AF, Campbell HJ, Cooper RA, Crampton JS, Crundwell MP, Hollis CJ, Morgans HEG. 2015. Revised calibration of the New Zealand Geological Timescale: NZGT2015/1. GNS Science Report 2012/39. 53 p.
- Richardson JR. 1981. Distribution and orientation of six articulate brachiopod species from New Zealand. N Z J Zool. 8:189–196. doi: 10.1080/03014223.1981.10427961
- Richardson JR. 1987. Brachiopods from the carbonate sands of the Australian shelf. Proc R Soc Victoria. 99(1):37–50.
- Ricketts BD, Ballance PF, Hayward BW, Mayer W. 1989. Basal Waitemata group lithofacies: rapid subsidence in an Early Miocene interarc basin, New Zealand. Sedimentology. 36:559–580. doi: 10.1111/j.1365-3091.1989.tb02086.x
- Robinson JH, Donald KM, Brandt AJ, Lee DE. 2016. Magasella sanguinea (Leach, 1814) and Magasella haurakiensis (Allan, 1931): resolving the taxonomic placement of these endemic New Zealand brachiopods using morphological and molecular traits. J R Soc N Z. 46(2):139–163. doi: 10.1080/03036758.2016.1182930
- Scott GH. 1965. Utility of Haeuslerella Parr (Foraminifera) in New Zealand Middle tertiary stratigraphy. New Zealand Paleontological Bulletin 38. Wellington: Government Printer. 48 p.
- Skinner DNB. 1969. Colville Formation – a new formation possibly correlative with the Waitemata Group. N Z J Geol Geophys. 12:349–360. doi: 10.1080/00288306.1969.10420286
- Sowerby GB. 1846. Descriptions of thirteen new species of brachiopods. Proc Zool Soc Lond. 14:285–314.
- Squires DF. 1962. A scleractinian coral fauna from Cape Rodney. N Z J Geol Geophys. 5:508–514. doi: 10.1080/00288306.1962.10420104
- Suess E. 1864. Brachiopoden. In: Zittel KA. Fossilen Mollusken und Echinodermen aus Neu-Seeland. “Reise der Österreichischen Fregatte “Novara” um die Erde in Jahren 1857, 1858, 1859”. Geologischer Theil. 1(2):56–61.
- Thomson JA. 1915a. Brachiopod genera: the position of shells with magaselliform loops, and of shells with bouchardiform beak characters. Trans Proc N Z Inst. 47:392–403.
- Thomson JA. 1915b. A new genus and species of the Thecidiinae. The Geological Magazine, Decade. 6, 2:461–464.
- Thomson JA. 1915c. The genera of Recent and Tertiary Rhynchonellids. The Geological Magazine, Decade 6. 2:387–392.
- Thomson JA. 1916. Additions to the knowledge of the Recent and Tertiary Brachiopoda of New Zealand and Australia. Trans Proc N Z Inst. 48:41–47.
- Thomson JA. 1918a. Appendix. In: Park J. The Geology of the Oamaru District, North Otago (eastern Otago Division). New Zealand Department of Mines, Geological Survey Branch, Bulletin 20; p. 117–119.
- Thomson JA. 1918b. Australasian Antarctic Expedition, 1911–1914, under the leadership of Sir Douglas Mawson, D. Sc., B.E., Brachiopoda. Scientific reports (Sydney), Series C, 4(3):1–76.
- Thomson JA. 1920. The Notocene geology of the Middle Waipara and Weka Pass district, North Canterbury, New Zealand. Trans Proc N Z Inst. 52:322–415.
- Thomson JA. 1926. A revision of the subfamilies of the Terebratulidae (Brachiopoda). Ann Mag Nat Hist (London), Series 9. 18(107):523–530. doi: 10.1080/00222932608633548
- Waagen WH. 1883. Salt Range Fossils. I. Productus Limestone Fossils. Geological Survey of India, memoirs. Palaeontologia Indica (Series 13). 492:391–546.
- Waterhouse BC. 1974. Notes from the New Zealand Geological Survey-8. N Z J Geol Geophys. 17(2):487–490. doi: 10.1080/00288306.1974.10430406
- Wilson DD. 1963. Geology of Waipara Subdivision. New Zealand Geological Survey Bulletin 64. 122 p.
- von Zittel KA. 1864. Fossilen Mollusken und Echinodermen aus Neu-Seeland. Nebst Beiträgen von den Herren Bergrath Franz Ritter v. Hauer und Professor Eduard Suess. 15–68, pls 9–15. In: Hochstetter F von, Hörnes M, Hauer FR von eds. Paläontologie von Neu-Seeland. Beiträge zur Kenntniss der fossilien Flora und Fauna der Provinzen Auckland und Nelson, von Prof. Dr. F. Unger, Prof. Dr. Karl Zittel, Prof. E. Suess, Felix Karrer, Dr. Ferdinand Stoliczka, Dr. Guido Stache, Dr. Gustav Jaeger. Reise der Österreichischen Fregatte Novara um die Erde in den Jahren 1857, 1858, 1859 unter der Befehlen der Commodore B. von Wüllerstorf-Urbair, Geologischer Theil. 1(2).