
ABSTRACT
Fish otoliths are essential tools for reconstructing fossil teleost faunas. Here, we describe otoliths from estuarine to deep-water Late Oligocene localities in southern Zealandia. Thirty-one species are recognised including 14 new species and two new genera. New species are: Moringua waimumuensis n.sp., Heteroconger? mataura n.sp., Tonganago coplandi n.sp., Sardinops robinsoni n.sp., Lotella latidorsalis n.sp., Trachyrincus tewaewae n.sp., Eurypleuron debilis n.sp., Neobythites lindqvisti n.sp., Aotearichthys vestalis n.sp., Centroberyx worthyi n.sp., Krebsiella chattonensis n.sp., Lesueurina transoceana n.sp. and Micropercops pomahaka n.sp., a representative of a basal gobioid family (Odontobutidae) now restricted to fresh- and brackish waters of eastern Asia. The new otolith-based genera are: Aotearichthys n.gen. (Dinematichthyidae) and Tonganago n.gen. (Congridae). The diverse faunal association reflects the range of paleoenvironments from estuarine to deep marine allowing an unusually complete reconstruction of the fish ecosystems. We compare the fauna with the extant marine fauna of Zealandia and discuss its phylogenetic and biogeographic significance.
Introduction
Fish macrofossils are rare but otoliths have been observed regularly in Cenozoic marine sediments of New Zealand and have been the focus of several publications (Frost Citation1924, Citation1928, Citation1933; Stinton Citation1957; Schwarzhans Citation1980; Grenfell Citation1984; Grenfell and Schwarzhans Citation1999; Schwarzhans et al. Citation2012). A monographic review of fossil otoliths of New Zealand (Schwarzhans in prep) will result in the recognition of more than 300 otolith-based fish species. This paper describes otoliths from a suite of Late Oligocene (Duntroonian and early Waitakian = Chattian) sites in the Southland Basin that represents a range of habitats from brackish estuarine to deep offshore environments. It provides an overview of the fish biodiversity across the environmental range locally and a comparative basis for understanding the origins of the modern fish fauna of the southwest Pacific region.
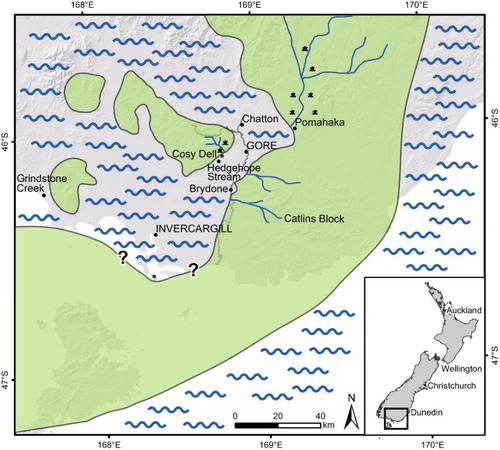
The otolith associations described here may seem relatively small with 32 recognised species, including four in open nomenclature and 14 new to science, but they cover an amazingly diverse range of paleoenvironments present in the Late Oligocene (Duntroonian and early Waitakian (27.3 Ma to c.23 Ma)) in southern New Zealand. Otoliths were collected from a brackish to estuarine setting at Pomahaka, a rocky shore and sandy beach setting at Cosy Dell, a fully marine, sheltered embayment at Brydone, from inner shelf environments at Hedgehope Stream and Chatton, and finally from a truly deep marine offshore environment with turbiditic influence at Grindstone Creek (). The otolith-based fish associations in each of these environmental settings proved to be highly divergent from each other. Individual locations commonly contain species not identified from any of the other localities and apparently stemming from fishes adapted to very specific habitats. The elucidation of these habitual dependencies is a main part of the results of our research. Other aspects discussed in the light of our findings are the nature and origin of endemic groups of fishes in the New Zealand region and interactions that may have occurred with neighbouring Australia.
Figure 1. Paleogeographic map showing the extent of the Late Oligocene sea overlaid on a modern map of southern New Zealand. Otoliths were collected for this study from Pomahaka, Chatton, Cosy Dell, Hedgehope Stream, Brydone and Grindstone Creek.
Localities, stratigraphy and regional geology
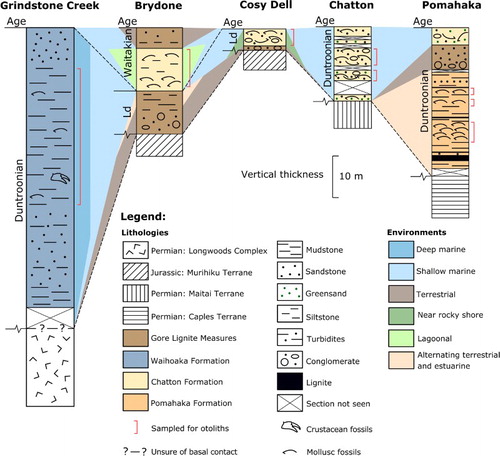
The otoliths were collected from six localities of Late Oligocene age from southern New Zealand (): Pomahaka, Brydone, Cosy Dell, Hedgehope, Chatton and Grindstone Creek. They represent three formations: Pomahaka, Chatton and Waihoaka which are summarised below ().
Figure 2. Stratigraphic and paleoenvironmental correlation chart of the studied locations from the Late Oligocene in the Southland Basin, South Island, New Zealand. The sections are modified from Suggate et al. (Citation1978), Isaac and Lindqvist (Citation1990), Lee et al. (Citation2014), Gard (Citation2016) and Lindqvist et al. (Citation2016). Horizontal distances are not to scale.
Pomahaka Formation
This is a 30–90 m thick estuarine deposit comprising interbedded mudstone, lignite, shell beds and thin sands with a distinctive fauna of estuarine and freshwater molluscs (Isaac and Lindqvist Citation1990; Lindqvist et al. Citation2016). The microtidal estuary extended over an area of at least 50 km2, and sediments record numerous cycles of transgressive flooding of forest swamps by fresh or estuarine waters. Pomahaka Formation rests unconformably on Caples Terrane basement of Permian age (Isaac and Lindqvist Citation1990) and is overlain by a thin bed of basal Gore Lignite Measures. The formation is dated as Duntroonian from palynological evidence (Pocknall Citation1982). Ten otoliths placed in two families were retrieved from Pomahaka Formation.
Chatton Formation
This widespread formation comprises fossiliferous shallow marine sandstones, sandy siltstones, greensands, grainstones and conglomerates exposed in thin and discontinuous outcrops over much of eastern Southland (Isaac and Lindqvist Citation1990; Gard Citation2016). Fossil autecology suggests water depths between intertidal and mid shelf. Dating of Chatton Formation using foraminifera and molluscs reveals that the formation is Duntroonian in the north and Waitakian in the south at Brydone (Isaac and Lindqvist Citation1990; Gard Citation2016). The Chatton Formation was deposited during a significant highstand in a shallow seaway that extended over the east Southland platform during the late Duntroonian and early Waitakian (Isaac and Lindqvist Citation1990).
Brydone
Two otoliths representing two families are present in shelly, muddy siltstone exposed in a river section near Brydone, a few metres above the contact with the underlying Murihiku Terrane basement of Jurassic age (Isaac and Lindqvist Citation1990; Gard Citation2016). The fauna comprises many undescribed species of molluscs and indicates a fully marine, sheltered bay with algal-covered reefs and intertidal mudflats nearby (Gard Citation2016).
‘Shell Gully’, Kelly’s Farm, Chatton
About 100 otoliths representing 16 species were collected from this site which is the type locality of Chatton Formation. The unconformable contact with the underlying Permian Maitai Terrane is not exposed, but pebbles of Atomodesma-bearing siltstone in the shelly horizons suggest Maitai Terrane basement was being eroded close by during deposition of Chatton Formation (Gard Citation2016). The diverse molluscan fauna from ‘Shell Gully’ has been investigated for more than a century (see Marwick Citation1929). The molluscan and benthic foraminiferan faunas indicate a shallow sublittoral setting with chitons and other taxa indicating a nearby rocky shore.
Cosy Dell
Several hundred otoliths representing 20 species and 16 families were collected from a small quarry on Cosy Dell farm where a 5 m thick exposure of Chatton Formation rests unconformably on Murihiku Terrane basement of Jurassic age. The fauna and flora of the site were discussed in detail in Lee et al. (Citation2014) and Conran et al. (Citation2014), respectively. The locality has yielded a remarkable array of diverse and exceptionally well-preserved fossils derived from rocky shore, estuarine, sandy beach and shallow subtidal habitats.
Hedgehope Stream
Eleven otoliths in three families were collected from a concretionary shell bed in Chatton Formation at Hedgehope Stream, c. 1 km south of the Cosy Dell farm quarry (Isaac and Lindqvist Citation1990, figs 18 and 19). The fauna comprises a moderately diverse range of molluscs and foraminifera that indicate an exposed shallow sublittoral setting (Gard Citation2016).
Waihoaka Formation
This formation crops out on the eastern margin of the Waiau basin south of the Longwood Range and comprises marine facies including conglomerate, fossiliferous sandstone, limestone and mudstone (Turnbull and Allibone Citation2003) that rest unconformably on Longwood Complex igneous basement of Permian age. It is dated as Duntroonian on the basis of foraminiferal assemblages (Wood Citation1969).
Grindstone Creek
Thirteen otoliths representing six species have been collected from a massive blue-grey siltstone on the coast c. 200 m northwest of mouth of Grindstone Creek, near Orepuki (Wood Citation1969), c. 80 km southwest of Cosy Dell. This locality contains a mixed faunule of unusual deep-water molluscs and shallow-water, large foraminiferans. The sediments here include turbidites and some fossils appear to have been redeposited from shallow environments into deep water.
Materials and methods
Most otoliths from Cosy Dell, Brydone and Hedgehope Stream, and some of those from Pomahaka and Chatton were collected during fieldwork carried out as part of MSc thesis research by HJLG in 2015–2016, and are held in the reference collections of the Geology Museum, University of Otago. Some Chatton specimens, and most of those from Grindstone Creek were collected by the late Philip Maxwell, and are held in the vertebrate collections of Te Papa Tongarewa, the Museum of New Zealand, Wellington, which also holds otoliths from Pomahaka collected by Bryce McKenzie (in 1981), and Bruce Marshall and Phillip Maxwell (in 2003). A few specimens from Grindstone Creek were collected by Dick Willett (in 1940) and are held at GNS Science, Lower Hutt.
All otoliths were collected as isolated specimens on the sediment surface in the field or picked from dried sieve concentrate under a microscope after washing and sieving bulk samples with a minimum mesh size of 0.5 mm. Individual otoliths were photographed either with a JEOL 6700 FESEM electron microscope at the Otago Centre for Electron Microscopy, University of Otago or a Canon EOS camera mounted on a Wild M400 photomacroscope remotely controlled from a computer at the home of WS. Optical photos were taken at different field of depth levels and stacked with the Helicon Focus software of Helicon Soft Ltd. Individual figures were optimised by Adobe Photoshop enhancing for contrast and retouching of minor inconsistencies. All otoliths are figured from the inner face, unless otherwise indicated, and are shown as right otoliths to better facilitate comparison. Left otoliths are reversed and are annotated accordingly.
The terminology of otoliths follows Koken (Citation1891), with amendments by Weiler (Citation1942) and Schwarzhans (Citation1978). Abbreviations used: CaL = cauda length, OH = otolith height, OL = otolith length, OsL = ostium length, OT = otolith thickness, SuH = sulcus height, SuL = sulcus length.
All localities have been given fossil record numbers and are logged in the New Zealand Fossil Record File (see http://www.fred.org.nz—FRED, Fossil Record Electronic Database of the Geosciences Society of New Zealand; accessed February 2017). Macro- and microfossil collections are held in the Geology Department, University of Otago (catalogue numbers prefixed with OU), in Te Papa Tongarewa, the Museum of New Zealand, Wellington, (catalogue numbers prefixed with NMNZ) and in the paleontological collections at GNS Science, Lower Hutt.
Systematic paleontology
Generic allocation of fossil otoliths depends very much on comparison with modern (Holocene) material, except for those instances where articulated skeletons are known with otoliths in situ (see Schwarzhans et al. Citation2017a, Citation2017b, Citation2017c). As a consequence, the accuracy of the attribution of fossil otolith-based species to extant genera decreases with geological age. This has led specialists in the field of otolith research to propose alternative open generic nomenclature schemes or to use names of extant genera in a very broad meaning (sensu lato) to avoid erecting a plethora of otolith-based fossil genera. Fortunately, the fauna covered here from the Late Oligocene of southern New Zealand exhibits a high degree of consistency with the Holocene New Zealand fish fauna and many of the fossil otoliths described can be contributed to extant genera with a high degree of certainty, sometimes, as stated above, in a sensu lato meaning (then accompanied by a ‘?’). We have refrained from the usage of open generic nomenclature, because none of the methods proposed to date are fully compliant with the ICZN rules (see Schwarzhans Citation2012 and Bratishko et al. Citation2015 for details). In a few instances, the morphology of certain otolith-based species clearly indicates the presence of extinct teleost groups. In these cases, we have made use of fossil otolith-based genera already established (denoted as † in ) or we have defined new genera when appropriate.
Table 1. Otolith species list of the studied localities from the Late Oligocene of the Southland Basin.
The higher classification of teleosts is in a highly dynamic state since molecular analyses have become common practice in phylogenetic studies. This molecular work is challenging many of the traditional arrangements in the higher classification of fishes, most notably in Percomorphi and Perciformes (Wiley and Johnson Citation2010; Betancur et al. Citation2013; Chen et al. Citation2014; Miya and Nishida Citation2014). We consider that these phylogenies are still too much in a state of flux to be applied to our study and therefore, have selected to follow the more ‘traditional’ classifications of Nelson (Citation2006), Roberts et al. (Citation2015) and the composition of Ophidiiformes following Møller et al. (Citation2016).
A complete monographic review of the fossil otoliths of New Zealand under preparation by WS will encompass documentation and definition of all species so far recognised, including the ones described here. We have therefore kept the systematic part as an annotated species list and provided full diagnoses and descriptions only for new taxa.
Order Anguilliformes
Family Moringuidae
Genus Moringua Grey 1831
Moringua waimumuensis n.sp.
(A)
Figure 3. Anguilliform otoliths. A, Moringua waimumuensis n.sp., holotype, OU22814, Cosy Dell, F45/f0396, Duntroonian (A1 = anterior view, A3 = ventral view). B,C, Heteroconger? mataura n.sp., Chatton, F45/f9668, Duntroonian, B = holotype, OU22830 (B2 = anterior view, B3 = ventral view); C = paratype (reversed), OU22831. D, Gnathophis ornatus (Frost Citation1928), OU22823, Cosy Dell, F45/f0396, Duntroonian (D2 = ventral view). E, Pseudophichthys sp., Chatton, F45/f9668, Duntroonian, NMNZ S.46910. F, Rhynchoconger otapiriensis (Frost Citation1933), OU22832, Cosy Dell, F45/f0396, Duntroonian (F2 = ventral view).
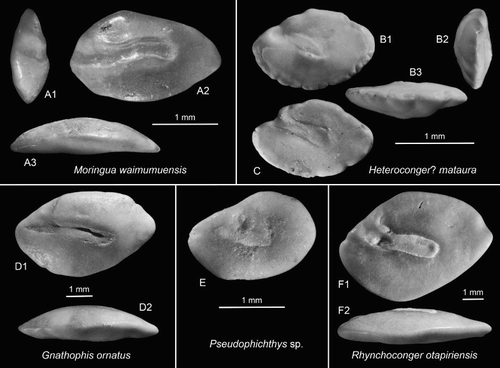
Holotype (and unique specimen): (A), OU22814, Cosy Dell, F45/f0396, Chatton Formation, Duntroonian, Late Oligocene.
Etymology: After the township of Waimumu located near the type locality.
Diagnosis: OL:OH = 1.65. Broad predorsal angle, flat postdorsal region. Posterior tip expanded. Inner face strongly convex. Sulcus sinuate, anteriorly open, without excisura and not deepened.
Description: The unique holotype is well preserved and 2.3 mm long. It is moderately elongate and thick (OL:OH = 1.65; OH:OT = 2.5). The dorsal rim is steep anteriorly and gently inclined, nearly straight posteriorly with a broad predorsal angle in between. The ventral rim is regularly curved and deepest just behind its middle. The anterior tip is broadly rounded without rostrum, antirostrum or excisura. The posterior tip is broad, somewhat expanded, dorsally rounded and ventrally flat. All rims are smooth.
The inner face is distinctly convex, with a narrow, somewhat deepened and slightly supramedian positioned, sinuate sulcus. The sulcus shows a vague distinction into approximately equally long ostium and cauda. The ostium is anteriorly open, slightly downward inclined and very slightly widened; its ventral margin is bent and its dorsal margin turns upwards anteriorly almost like an indication of an ostial channel. The cauda is markedly sinuate and distinctly upward directed with its tip swinging slightly downwards again. It terminates far from the posterior tip of the otolith (OL:SuL = 1.45). There is an indistinct dorsal depression and no ventral furrow. The outer face is flat and smooth.
Discussion: The Moringuidae are a small family of eel-like fishes living in burrows on sandy bottoms in shallow marine tropical seas, primarily the Indo-West Pacific. Their otoliths belong to a group in the Anguilliformes characterised by an anteriorly open and somewhat deepened sulcus. This group, which also comprises Anguillidae, Ophichthidae, Muraenidae and others, are best diagnosed by the moderate depth of the sulcus and the lack of an excisura. Moringua waimumuensis is the first fossil record for the family and it differs from the few described Holocene species in the sinuate shape of the sulcus (for figures of Holocene otoliths see Lin and Chang Citation2012 and Nolf Citation2013).
Family Congridae
Genus Gnathophis Kaup 1859
Gnathophis ornatus (Frost Citation1928)
(D)
1928 Ot. (Congridarum) ornatus – Frost: figure 4
1928 Congeris wharekuriensis – Frost: figure 1
1928 Congeris rectus - Frost: figure 2
1957 Heterenchelys crassus – Stinton: figure 6
1980 Gnathophis ornatus (Frost Citation1928) - Schwarzhans: figures 20–22
Material: Five specimens: four specimens, figured specimen OU22823, Cosy Dell, F45/f0396; one specimen, OU22824, Chatton, F45/f9668.
Discussion: Gnathophis ornatus is a widely distributed and common species throughout the Duntroonian to Altonian (27.3–15.9 Ma) of New Zealand (Schwarzhans Citation1980).
Genus Heteroconger Bleeker 1868
Heteroconger? mataura n.sp.
(B,C)
Holotype: (B), OU22830, Kelly’s Farm near Chatton, F45/f9668, Chatton Formation, Duntroonian, Late Oligocene.
Paratypes: Two specimens, OU22831, same data as holotype.
Etymology: Named after the Mataura River, the main drainage river of the Southland region with the type locality located close to it. Mataura is used as noun in apposition.
Diagnosis: OL:OH = 1.5–1.55. Symmetrical otolith with equally pointed anterior and posterior tips. Inner face flat except for slightly elevated sulcal area. Sulcus short, shallow, steeply inclined at 22–25° versus long otolith axis, colliculum reduced to rear part of sulcus.
Description: Small, oval otoliths reaching c. 1.55 mm in length (holotype). They are moderately thick (OH:OT = 2.0). The dorsal and ventral rims are gently curved and equally deep with their deepest point at about their respective middle. The ventral rim is more regularly curved than the dorsal rim and slightly crenulated while the dorsal rim is irregularly undulating. The anterior and posterior tips are moderately projecting, with a central position and are symmetrical in development.
The inner face is flat; though slightly elevated in the area of the cauda. The sulcus is short, steeply inclined at c. 25° and shallow. The opening of the sulcus towards the anterior-dorsal rim is fading in a way that it remains incomplete. There is no, or only a faint indication of, an ostial channel close to the anterior tip of the sulcus. The index OL:SuL is very high (2.0). The sulcus shows no clear distinction into ostium and cauda, but it appears that the small, oval colliculum is only developed in the rear part of it, probably equivalent to the cauda, whereas the anterior part (the ostium) bears no colliculum. There is a very indistinct dorsal depression and no ventral furrow. The outer face is distinctly convex, stronger than the inner face and more or less smooth.
Discussion: The genera Heteroconger and Gorgasia are known as garden eels and inhabit shallow tropical sandy bottom environments. Holocene otoliths are figured in Schwarzhans and Bratishko (Citation2011). The otoliths of Heteroconger? mataura differ from the known Holocene otoliths of this genus in that they have a more strongly inclined sulcus and an anteriorly reduced colliculum. It is therefore only tentatively associated with the genus Heteroconger. Juveniles of other, larger congrid species could account for the small size of these otoliths; however, the only other species identified so far from the Duntroonian of the area is Rhynchoconger otapiriensis (see below). Rhynchoconger otapiriensis does share many characters, but small specimens of this species always differ in the inner face, being more distinctly convex than the outer face, and in the ostial region of the sulcus, which shows no such reduction of the colliculum and the presence of a clear, albeit short ostial channel.
Genus Pseudophichthys Roule 1915
Pseudophichthys sp.
(E)
Material: One juvenile specimen of 1.95 mm length, NMNZ S.46910, Chatton, F45/f9668.
Discussion: Small otolith characterised by an oval shape (OL:OH = 1.45) with a depressed predorsal rim. The sulcus is rather short and unsegmented, with a single, oval colliculum and a distinct ostial channel positioned above the anterior third of the colliculum.
Genus Rhynchoconger Jordan & Hubbs 1925
Rhynchoconger otapiriensis ( Frost Citation1933 )
(F)
1933 Ot. (Congridarum) otapiriensis – Frost: pl. 22, figure 28
1980 Gnathophis otapiriensis (Frost Citation1933) – Schwarzhans Citation1980: figures 18–19
Material: 17 specimens: two specimens, figured specimen OU22832, Cosy Dell, F45/f0396; 15 specimens Chatton, F45/f9668.
Discussion: Rhynchoconger otapiriensis appears to be endemic to the Duntroonian of the South Island of New Zealand (Schwarzhans Citation1980).
Genus Tonganago n.gen.
Type species: ?Scalanago fastigatus Schwarzhans Citation1980
Etymology: A combination of the Māori word tonga (= southern) and anago, a word stem found in several congrid genera such as Scalanago, with which it compares. Tonganago refers to its common occurrence in the fossil record in the Southern Hemisphere, particularly in New Zealand.
Diagnosis: A fossil otolith-based genus of the family Congridae with the following combination of characters: inner face markedly convex; outer face flat to slightly convex. Sulcus long, deep, narrow, straight and reaching close to the anterior-dorsal rim of the otolith. Ostial channel indistinct, very short, close to anterior tip of ostium. Sulcus inclination angle moderate (3–10°). Posterior third of sulcus distinctly widened ventrally and tapering in a dorsally positioned tip. Dorsal depression absent or indistinct. Dorsal rim thickened.
Discussion: Otoliths of Tonganago resemble otoliths of several extant bathymyrine genera such as Scalanago, Congriscus, Paraconger and congrine genera such as Gnathophis in the convex inner face without a clear dorsal depression, and the deepening and overall shape of the sulcus. It differs from all these in two main characters: the thickened dorsal rim contrasting with the sharp ventral rim and the peculiar shape of the posterior third of the sulcus. In addition, the ostial channel is very short, and located in a more forward position than in most comparable Holocene genera.
Species: Four species from the Eocene to Miocene of New Zealand are here placed in Tonganago: Tonganago fastigatus (Schwarzhans Citation1980) (originally described as ?Scalanago fastigatus) from the Mangaorapan to Bortonian (52–39.1 Ma), Tonganago obliquus (Stinton Citation1957) (originally described as ?Mystriophis obliquum) from the Clifdenian, Tonganago sagittisulcatus (Schwarzhans Citation1980) (originally described in incertae sedis Congridae) from the Duntroonian and Waitakian and T. coplandi n.sp. from the Duntroonian to Altonian. A fifth undescribed species may be present in the Eocene. To date, Tonganago is known exclusively from New Zealand.
Tonganago coplandi n.sp.
(A–C)
Figure 4. Anguilliform and clupeiform otoliths. A–C, Tonganago coplandi n.gen. n.sp., Cosy Dell, F45/f0396, Duntroonian, A = holotype (reversed), OU22833 (A1 = anterior view, A3 = ventral view); B,C = paratypes (B reversed), OU22834 (B2 = ventral view, B3 = anterior view). D–F = Tonganago sagittisulcatus (Schwarzhans Citation1980), OU22835, Cosy Dell, F45/f0396, Duntroonian (D2 = anterior view, F2 = ventral view). G, Sardinops robinsoni n.sp., holotype, OU22806, Cosy Dell, F45/f0396, Duntroonian (G2 = ventral view).
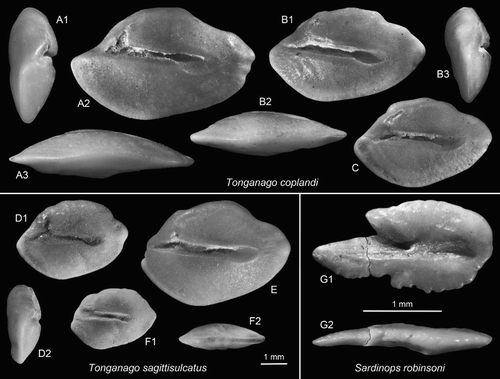
Holotype: (A), OU22833, Cosy Dell, F45/f0396, Chatton Formation, Duntroonian, Late Oligocene.
Paratypes: 30 specimens: 29 specimens, OU22834, same data as holotype; one specimen, OU22825, Hedgehope Stream, F45/f0410, Chatton Formation.
Etymology: In honour of Trevor Copland, the owner of the Cosy Dell farm, who has been extremely helpful and supportive in allowing collecting at this important fossil location.
Diagnosis: OL:OH = 1.65–1.8. Anterior rim with sharp, rostrum-like projection. Middorsal section expanded. Distinctive ventral widening at rear part of sulcus. Sulcus inclined at 4–7° versus long otolith axis. Outer face flat to slightly convex.
Description: Moderately large, oval otoliths reaching c. 8 mm length (holotype 7 mm), moderately thick (OH:OT = 1.8–2.3). The dorsal rim is irregularly curved, expanded at its middle section, slightly concave postdorsally and slightly concave to straight predorsally. The ventral rim is gently curved and deepest anterior of its middle. The anterior tip is projecting and pointed. The posterior tip is broadly projecting, rounded, slightly dorsally pronounced.
The inner face is strongly convex and smooth. The sulcus is long (OL:SuL = 1.5–1.6), moderately deepened and slightly inclined. Its rear tip is ventrally widened and dorsally tapering, which is typical for the genus. Anteriorly it closely approaches the anterior-dorsal rim and shows a short ostial channel. The colliculum is poorly defined due to the deepening of the sulcus. There is no, or a very indistinct, dorsal depression and no ventral furrow. The outer face is flat to slightly convex and smooth.
Discussion: Tonganago coplandi differs from the contemporaneous T. sagittisulcatus in the projecting and pointed anterior tip, the ratio OL:OH (1.65–1.8 vs 1.4–1.6) and the relatively strongly ventrally widened posterior third of the sulcus. Also, T. coplandi seems to grow to larger sizes than T. sagitticulcatus (max. length 8 mm vs 6 mm). Tonganago coplandi differs from T. fastigatus from the Eocene in the middorsally expanded region and the combination of a sharp anterior and a broad posterior tip (vs rounded anteriorly and a spine-like posterior tip). The Miocene T. obliquus is distinguished by a flat dorsal rim (versus middorsally expanded), rounded anterior and posterior tips and a very high ratio OL:OH (1.9–2.1 vs 1.65–1.8). Tonganago coplandi thus occupies a morphologically and stratigraphically intermediate position between T. fastigatus and T. obliquus.
Tonganago sagittisulcatus (Schwarzhans Citation1980)
(D–F)
1980 Congridarum sagittisulcatus – Schwarzhans: figure 37
Material: 47 specimens, figured specimens OU22835 Cosy Dell, F45/f0396.
Discussion: Tonganago sagittisulcatus has the most compressed otolith morphology in this genus (OL:OH = 1.4–1.6) characterised by a very reduced and blunt anterior tip and a relatively high dorsal rim. The species is stratigraphically restricted to the Duntroonian and Waitakian.
Order Clupeiformes
Family Clupeidae
Genus Sardinops Hubbs 1929
Sardinops robinsoni n.sp.
(G)
Holotype: (G), OU22806, Cosy Dell, F45/f0396, Chatton Formation, Duntroonian, Late Oligocene.
Paratypes: One incomplete specimen, OU22826, same data as holotype.
Etymology: In honour of Jeffrey Robinson, University of Otago in recognition of his support in many aspects of fossil collecting and processing from Cosy Dell and Pomahaka.
Diagnosis: OL:OH = 1.9. Rostrum 35% of OL; antirostrum 7% of OL. Rostrum slender, set off from rear part of ventral rim by a marked indention at c. 43% from anterior tip. Posterior rim broadly rounded, slightly dorsally pronounced. Dorsal rim shallow, slightly curved.
Description: The holotype is a fragile, delicate otolith with a broken but retrieved rostrum, while the paratype lacks the rostrum. The length of the holotype is 2.3 mm, the thickness OH:OT is 3.2. The rostrum is long, with parallel dorsal and ventral margins for about half of its length except for the pointed tip. The excisura is deep and sharp; the antirostrum moderately long with an angular tip. The dorsal rim is shallow, curves gently and is slightly undulating. The ventral rim shows a distinct notch at the base of the rostrum and posteriorly, is only slightly curved and irregularly serrated. The posterior rim is broadly rounded, somewhat forward bent ventrally and slightly expanding dorsally.
The inner face is slightly bent anteriorly and nearly flat posteriorly. It carries a median, deep and widely opened sulcus that terminates close to the posterior rim. Ostium and cauda are not distinguishable along the nearly straight ventral sulcus margin, but through the more deepened ostium. The ratio OsL:CaL is 1.3. The dorsal sulcus margin turns upward above the ostium towards the tip of the antirostrum and is slightly bent above the cauda. The deepened ostial colliculum shows a well defined and straight ventral margin. The cauda is somewhat deepened in its posterior part. The dorsal depression is indistinct and there is also a narrow ventral depression on the rostrum below the ostium. The outer face is flat to slightly concave anteriorly and with a shallow umbo opposite of the ostial–caudal joint of the inner face.
Discussion: Sardinops robinsoni is easily recognised by the characteristic shape of the rostrum and the deep excisura and moderately large antirostrum. It resembles the Holocene Sardinops sagax (Jenyns 1842) from New Zealand and southern Australia (otoliths figured in Furlani et al. Citation2007), but differs in the shape of the ostium, the less pronounced dorsal portion of the posterior rim and the lack of a dorsal depression.
Order Myctophiformes
Family Myctophidae
Genus Diaphus Eigenmann & Eigenmann 1890
Diaphus curvatus Schwarzhans Citation1980
1980 Diaphus curvatus – Schwarzhans: figures 163–165
Material: One specimen, Grindstone Creek, D46/f0054.
Discussion: Diaphus curvatus was a common species in beds representing offshore environments of Duntroonian to Altonian age in New Zealand. There are also two myctophid specimens from Grindstone Creek and six further examples from Chatton, which are too poorly preserved for identification beyond family level.
Order Gadiformes
Family Moridae
Genus Lotella Kaup 1858
Lotella latidorsalis n.sp.
(A–C)
Figure 5. Gadiform otoliths. A–C, Lotella latidorsalis n.sp., A = holotype, OU22807, Cosy Dell, F45/f0396, Duntroonian (A2 = outer face, A3 = anterior view, A4 = dorsal view, A5 = ventral view, A6 = posterior view); B,C = paratypes (C reversed), NMNZ S.46911-12, Chatton, F45/f9668, Duntroonian (B2 = outer face, B3 = ventral view). D,E, Trachyrincus tewaewae n.sp. Grindstone Creek, D46/f0054, Duntroonian, D = holotype (reversed), NMNZ S.46913 (D2 = outer face, D3 = ventral view, D4 = anterior view); E = paratype, NMNZ S.46914. F, Bregmaceros sp., GNS F5957, Grindstone Creek, D46/f8497, Duntroonian (F2 = posterior view, F3 = ventral view).
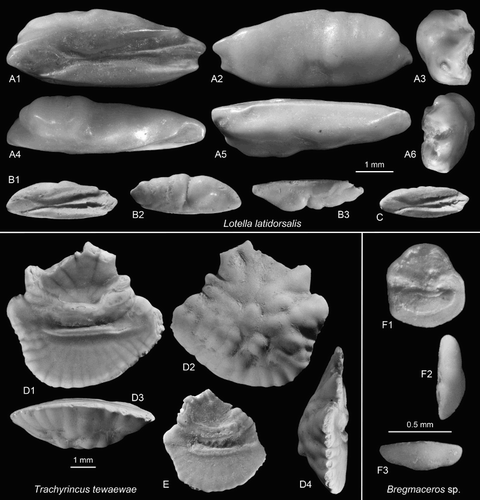
Holotype: (A), OU22807, Cosy Dell, F45/f0396, Chatton Formation, Duntroonian, Late Oligocene.
Paratypes: Four specimens: one specimen, OU22827, same data as holotype; one specimen, OU22828, Brydone, F46/f8492, Chatton Formation; two specimens, NMNZ S.46911-12, Chatton, F45/f9668, Chatton Formation.
Etymology: Combined from latus (Latin) = broad and dorsalis (Latin), referring to the broad dorsal rim extending well over the ostium.
Diagnosis: OL:OH = 2.65–2.85; OH:OT = 1.3. Dorsal rim shallow, expanding over ostium resulting in broad anterior rim. Ostial colliculum short; CaL:OsL = 2.2–2.4. Caudal colliculum projecting only slightly beyond ventral and dorsal cristae.
Description: Elongate, moderately thick, reaching a size of 5.3 mm length (holotype). The dorsal rim is shallow, almost flat and slightly undulating, somewhat expanding over the ostium and posteriorly reduced. The ventral rim is shallow, only slightly bent and smooth. It is anteriorly joined to the ventral margin of the ostium and terminates posteriorly in a slightly projecting tip of the crista inferior. The anterior tip is broad, rounded. The posterior tip is pointed at a short projection bearing the ridge like caudal colliculum; the ventral projection formed by the tip of the crista inferior is slightly shorter; the dorsal projection is indistinct, slightly shorter than the ventral projection and corresponding to the tip of the crista superior. The rear part of the crista superior including the dorsal projection is slightly bent outwards from the inner face (best seen in dorsal view; A4).
The inner face is nearly flat to slightly bent, somewhat stronger at the posterior dorsal field. The ostium is shallow, short, with oval colliculum. The cauda is deep, widening posteriorly and wide open, with a straight, highly elevated, thin central collicular ridge, which is anteriorly connected to the ostial colliculum and terminates posteriorly in the longest projection at the posterior tip. The ventral field is narrow and smooth. The dorsal field is small, short, and developed only above anterior part of the cauda, and shows an edge towards the overlapping outer face. The outer face is moderately convex in vertical direction (A3 and A6), relatively smooth with few crossing furrows, overlapping onto the inner face along the entire dorsal rim, and slightly thickened at the anterior-dorsal area.
Discussion: Lotella latidorsalis resembles Holocene otoliths of the genera Lotella and Eeyorius. The low dorsal rim without predorsal projection, the moderate thickness and the three tips at the posterior rim being relatively equal in length are typical for otoliths of these genera. Both are shallow-water morid genera known from the temperate seas of Australia, New Zealand, Japan and the Juan Fernandez Islands of the southeastern Pacific (see Schwarzhans et al. Citation2012 for figures of Holocene otoliths). Also, extant representatives are characterised by a relatively long ostial colliculum resulting in a ratio CaL:OsL of 1.5–1.8. Lotella latidorsalis differs from extant Lotella species in the shorter ostium (CaL:OsL = 2.2–2.4) and the anterior dorsal rim not being depressed. The short ostium resembles otoliths of many other morid genera typically residing in deeper water on continental slopes, such as Laemonema, Physiculus or Tripterophycis. Since all other diagnostic features (shape of dorsal rim, relation of tips at posterior rim, thickness) match with Lotella we assume that the short ostium may represent a plesiomorphic character state within the Lotella/Eeyorius lineage.
Family Bregmacerotidae
Genus Bregmaceros Thompson 1840
Bregmaceros sp.
(F)
Material: Two small and somewhat eroded specimens from Grindstone Creek, D46/f8497.
Discussion: These small otoliths of 0.75 mm length are remarkable for their regular outline without spines or major angles at the ventral rim, the straight, vertical anterior rim and the rounded dorsal rim. The index OL:OH is 0.95 and the index OH:OT is c. 3.0. They are further remarkable for their nearly flat inner face. However, the small size and moderate preservation preclude a specific identification.
Family Macrouridae
Genus Trachyrincus Giorna 1809
Trachyrincus tewaewae n.sp.
(D,E)
Holotype: (D), NMNZ S.46913, Grindstone Creek, D46/f0054, Waihoaka Formation, Duntroonian, Late Oligocene.
Paratypes: Two specimens, NMNZ S.46914, same data as holotype.
Etymology: Named after the Te Waewae Bay into which the Grindstone Creek opens. The Māori name is used as noun in apposition.
Diagnosis: OL:OH c. 1.0 (predorsal tip fragmented in holotype and posterior tip fragmented in paratype). Ventral rim deep; dorsal rim with massive and large predorsal lobe. Anterior-ventral rim concave. Sulcus narrow and short; SuL:SuH = 3.7–4.3; OL:SuL = 1.6–1.8. Dorsal and ventral fields with long radial furrows. Outer face with many central tubercles.
Description: Moderately large, high bodied otoliths reaching at least 6.3 mm length (holotype). The short, rounded anterior tip is located above the sulcus and set off dorsally by a deep and narrow concavity and ventrally by a broad and wide concavity. The dorsal rim is high, strongly sculptured (where preserved) and occupied by a broad and strongly projecting pre- to middorsal lobe. The posterior part of the dorsal rim is slightly concave and irregularly undulating. The posterior tip is rounded, rather short and positioned distinctly inferior to the anterior tip. The ventral rim is deeply curved, slightly undulating, with the deepest point distinctly anterior of its middle.
The inner face is straight in the vertical direction (D4) and slightly bent in the horizontal direction (D3). It carries a centrally positioned, rather short and narrow, and slightly deepened sulcus, which terminates at some distance from the anterior and posterior rims. It is filled by a merged colliculum, which is clearly bound ventrally, and anteriorly and posteriorly, but slightly confuse along its somewhat depressed dorsal margin. The anterior tip of the sulcus is connected via a furrow to the concave anterior-dorsal rim of the otoliths above its anterior tip. The inner face shows many fine radial furrows on the dorsal and ventral fields reaching close to the sulcus. There is a broad and deep dorsal depression, well-marked towards the sulcus. No ventral furrow is discernible. The outer face is strongly convex with radial furrows near the rims and many tubercles at its centre.
Discussion: Trachyrincus tewaewae shows all the hallmarks of a typical otolith of the genus including the outline of the otolith with the massive pre- to middorsal lobe, the thick outer face, the intense ornamentation and the characteristic shape of the sulcus with a single colliculum. It represents the oldest record of the genus and differs from the Holocene representatives (see Nolf Citation2013 for figures) in the short and narrow sulcus.
Genus Coelorinchus Giorna 1809
Coelorinchus sp.
Material: Two poorly preserved and eroded specimens from Grindstone Creek, D46/f0054.
Family Macruronidae
Genus Macruronus Günther 1873
Macruronus merlucciiformis Schwarzhans Citation1980
1980 Macruronus merlucciiformis – Schwarzhans: figure 228
Material: One fragmentary otolith from Cosy Dell, F45/f0396.
Order Ophidiiformes
Family Carapidae
Genus Eurypleuron Markle & Olney 1990
Eurypleuron debilis n.sp.
(A–E)
Figure 6. Ophidiiform otoliths. A–E, Eurypleuron debilis n.sp., A = holotype (reversed), OU22808, Cosy Dell, F45/f0396, Duntroonian (A1 = anterior view, A3 = ventral view, A4 = outer face); B = paratype (reversed), OU22809, Cosy Dell, F45/f0396, Duntroonian (B2 = ventral view); C–E = paratypes (E reversed), NMNZ S.46915-17, Chatton, F45/f9668, Duntroonian (C2 = ventral view). F,G, Neobythites lindqvisti n.sp., Grindstone Creek, D46/f0054, Duntroonian, G = holotype (reversed), NMNZ S.46918 (G2 = anterior view, G3 = ventral view); F = paratype (reversed), NMNZ S.46919 (F2 = ventral view). H, Aotearichthys vestalis n.gen. n.sp., holotype, OU22810, Brydone, F46/f8492, Waitakian (H2 = ventral view).
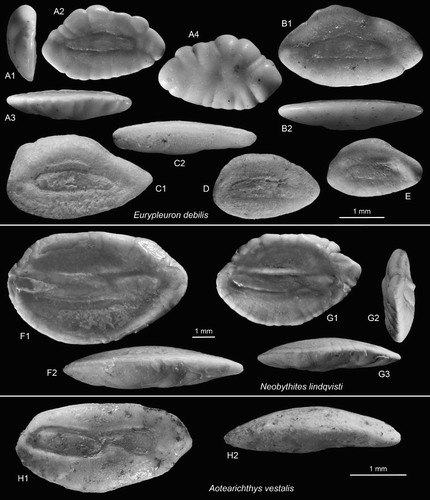
Holotype: (A), OU22808, Cosy Dell, F45/f0396, Chatton Formation, Duntroonian, Late Oligocene.
Paratypes: 14 specimens: four specimens, OU22809, same data as holotype; 10 specimens NMNZ S.46915-17 and OU22829, Chatton, F45/f9668, Chatton Formation.
Etymology: From debilis (Latin) = weak, feeble, referring to the thin appearance of the otoliths.
Diagnosis: OL:OH = 1.65–1.75; OH:OT = 2.5–2.7. Sulcus short and narrow; OL:SuL = 1.7–1.85; SuL = SuH = 3.8–4.5. Dorsal margin of sulcus slightly bent upwards middle of otolith and away from colliculum. Dorsal rim only slightly thickened. Sulcus terminating equally far from anterior and posterior tips of otolith.
Description: Delicate and rather thin otoliths for a carapid, reaching c. 3.25 mm length (holotype 2.75 mm). The dorsal rim is relatively high, irregularly crenelated or nearly smooth, not or slightly expanded anterior-dorsally. The ventral rim is more regularly curved, moderately deep and deepest anterior of the middle. It is smooth or finely crenelated. The anterior tip is more or less broadly rounded and slightly inferior. The posterior tip, likewise, is slightly inferior and rounded or slightly projecting and pointed.
Inner face nearly flat and only slightly bent in horizontal direction. The sulcus shows the typical shape of a carapid otolith, i.e. with a more strongly curved dorsal and a nearly straight ventral margin. However, the sulcus is comparatively short and narrow, and does not reach close to the anterior tip of the otolith. The single colliculum is clearly defined and shallow, level with the remainder of the smooth inner face. The central portion of its dorsal margin is slightly expanded, often leaving a widened space between sulcus margin and colliculum or a marked crista superior in that area. Dorsal and ventral fields are smooth, without clear dorsal depression or ventral furrow. The dorsal field shows no, or only a faintly outward bent portion of the inner face close to the otolith rim (A1). The otoliths are relatively thin including the dorsal portion and hence, the outer face is only slightly convex and smooth or with short radial furrows.
Variability and ontogeny: The available otoliths show some degree of variability expressed in the strength of the marginal crenulation and ornamentation of the outer face, and the expression of the posterior tip of the otolith and its predorsal rim (compare A and C). The short and narrow sulcus and the thin appearance, however, clearly indicate the specimens to represent the same species.
Carapid otoliths are also known for a remarkable ontogenetic allometry. Small specimens of sizes < 2 mm length tend to be more compressed and thinner than larger specimens. Holocene otoliths of the genus Eurypleuron achieve sizes of 3.5 mm, those of the genus Carapus 5–6 mm and of Onuxodon < 2 mm. We therefore conclude that the large specimens of E. debilis are diagnostically mature and represent adult specimens.
Discussion: Carapid otoliths are easily identified by the flat inner face, the uniform and shallow sulcus, the thickening of the dorsal rim and the overall shape of otolith and sulcus. However, differentiation of genera and species is less straightforward. Otoliths of Eurypleuron are characterised by the short sulcus terminating far from the anterior tip of the otolith and the thin appearance (Holocene specimen in collection of WS). These are the characters also found in E. debilis, which thus represents the first fossil record of the genus. In the Holocene, Eurypleuron contains two antitropical species, one off Japan and the other in the Southern Ocean off South Africa, Australia, New Zealand and Sala y Gomez.
Family Ophidiidae
Genus Neobythites Goode & Bean 1885
Neobythites lindqvisti n.sp.
(F,G)
Holotype: (G), NMNZ S.46918, Grindstone Creek, D46/f0054, Waihoaka Formation, Duntroonian, Late Oligocene.
Paratypes: One specimen, NMNZ S.46919, same data as holotype.
Etymology: Named in honour of Jon K. Lindqvist, University of Otago in recognition of his contributions to the better understanding of the depositional environments of the Southland region.
Diagnosis: OL:OH = 1.55–1.6. Predorsal and postdorsal angles equally strong; middorsal flat, horizontal, long, c. 50%–60% of OL. Sulcus narrow, with distinct and broad notch in ventral margin at ostial–caudal joint. OsL:CaL = 2.3.
Description: Compact, moderately elongate and moderately thick, reaching 8 mm length (holotype 6.4 mm). The dorsal rim is characterised by a long, straight and horizontal middorsal section. The anterior section is also straight, but steeply inclined at c. 45–50°; the posterior section is slightly longer, inclined at c. 35–40° and straight or slightly concave. The pre- and postdorsal angles are obtuse, rounded, and equally pronounced. The ventral rim is regularly curved and deep; deepest at about its middle. The anterior tip is broadly rounded, the posterior slightly more pointed; both positioned symmetrically and slightly superior. All rims are slightly crenulated; the ventral very finely and the dorsal more coarsely and irregularly.
The inner face is mildly convex and smooth. The sulcus is narrow, reaching close to the anterior tip of the otolith, and shows a straight or even slightly concave dorsal margin. OL:SuL = 1.25–1.35. Its ventral margin is slightly bent and shows a broad indentation at the level of the ostial–caudal joint. The ostium is more than twice as long as the cauda and not wider. Both are filled with clearly separated colliculi, which are elevated to the level of the remainder of the inner face. The dorsal depression is very large, but shallow and poorly defined except towards the sulcus. The ventral field is smooth and without ventral furrow. Fine radial furrows are ingressing from the marginal crenulation both dorsally and ventrally to various degrees. The outer face is moderately convex and shows some radial marginal furrows and flat tubercles at the centre.
Discussion: Neobythites is a species-rich genus with over 50 extant species. Otoliths are known from most of them (Schwarzhans Citation1994; Nielsen Citation1995, Citation1997, Citation1999, Citation2002), and are characterised by the general outline of otolith and sulcus, and in the sulcus being rather long and reaching close to the anterior tip of the otolith. Their differences are subtle and rely on details of the dorsal rim and proportions of the otolith and the sulcus. In addition, several species are known for sexual dimorphism in otoliths, a rare condition first described by Schwarzhans (Citation1994). In the event of sexual dimorphism, otoliths of males are generally morphologically altered and it is mandatory to define species by female otoliths. Fortunately, morphological differences are always similar, and if sexual dimorphism was at all developed it can be safely concluded that the two specimens described here were females. The combination of the narrow sulcus, the rather compressed shape and the proportions and shape of the dorsal rim distinguish N. lindqvisti well from all known Holocene and fossil species of the genus.
Family Dinematichthyidae
Genus Aotearichthys n.gen.
Type species: Aotearichthys vestalis n.sp.
Etymology: Deduced from Aotearoa (Māori) for New Zealand combined with ichthys (Greek) = fish.
Diagnosis: A fossil otolith-based genus in the family Dinematichthyidae with the following combination of characters. Inner face markedly convex; outer face flat, anteriorly thickened. Dorsal margin anteriorly depressed, without predorsal lobe. Posterior tip broadly projecting and slightly dorsally pronounced. Sulcus long, clearly distinguished in a shallow ostium and a slightly deepened cauda. OL:SuL = 1.35; OsL:CaL = 1.35. Ostium slightly upwards directed; ventral margin of cauda with broad and deep indentation behind ostium. Ventral furrow absent.
Discussion: Aotearichthys is a typical dinematichthyid otolith, but with a remarkable long cauda (low index OsL:CaL) and a similarly remarkably depressed pre-dorsal rim. Both characters distinguish it from all extant dinematichthyid genera except Porocephalichthys, a monospecific endemic genus off southwestern Australia (Møller and Schwarzhans Citation2008). The otoliths of Aotearichthys vestalis differ from Porocephalichthys dasyrhynchus in the shorter sulcus (OL:SuL = 1.35 vs 1.2), the broad indentation at the ventral sulcus margin between ostium and cauda (vs reduced small notch), and in the caudal colliculum being slightly deepened (vs shallow, like the ostial colliculum). No doubt, both genera are closely related and it appears that Aotearichthys represents the link between the highly advanced otolith morphology of Porocephalichthys to such genera as Alionematichthys or Dinematichthys.
Species: Aotearichthys is monospecific with A. vestalis from the Late Oligocene of southern New Zealand.
Aotearichthys vestalis n.sp.
(H)
Holotype (and unique specimen): (H), OU22810, Brydone, F46/f8492, Chatton Formation, Waitakian, Late Oligocene.
Etymology: From vestalis (Latin) = vestal, referring to the less developed morphology in comparison to the derived one of the genus Porocephalichthys.
Diagnosis: Monospecific genus; see diagnosis of genus.
Description: Moderately elongate and well preserved, 3.1 mm in length, with some dark mineral incrustations on the surface. OH:OT = 1.95; OH:OT = 1.95. The dorsal rim is shallow, anteriorly depressed and without pre- or postdorsal angles. The ventral rim is deeply curved, deepest at about its middle. The anterior tip is bluntly rounded, the posterior tip somewhat expanded, broad and dorsally pronounced. All rims are smooth.
The sulcus is positioned slightly supramedian and is clearly distinguished into a shallow ostium. Cauda is slightly deepened. The ostium is slightly upward inclined towards the anterior and reaches close to the anterior tip of the otolith. An ostial furrow connecting ostium and anterior rim of the otolith is not visible. The cauda likewise is slightly turned upward towards the posterior. It is separated from the ostium by a broad, abrupt indentation of the ventral sulcus margin behind the ostium. The caudal termination is broadly rounded and far from the posterior tip of the otolith. There is only a faint and narrow indication of a dorsal depression above the sulcus and no indication of a ventral furrow. The outer face is smooth, slightly convex anteriorly and slightly concave posteriorly.
Discussion: Like many other dinematichthyids, extant specimens of Porocephalichthys are mainly collected over rocky shores or associated with reefs in shallow water down to c. 20 m. This adaptation makes fossil occurrences rare and patchy in nature. Aotearichthys vestalis has only been found at Brydone in what was a very shallow environment with nearby algal encrusted reefs.
Order Beryciformes
Family Berycidae
Genus Centroberyx Gill 1862
Centroberyx worthyi n.sp.
(A–G)
Figure 7. Beryciform otoliths. A–G, Centroberyx worthyi n.sp., B = holotype (reversed), NMNZ S.46920, Grindstone Creek, D46/f0054, Duntroonian (B1 = ventral view, B3 = anterior view); A, C–G = paratypes (A, D–G reversed), OU22811, Cosy Dell, F45/f0396, Duntroonian (C2 = ventral view, D2 = outer face, D3 = ventral view). H–J, Optivus moko n.sp., Cosy Dell, F45/f0396, Duntroonian, H = holotype, OU22812 (H1 = anterior view, H3 = outer face, H4 = ventral view); I,J = paratypes (J reversed), OU22813 (I2 = ventral view).

Holotype: (B), NMNZ S.46920, Grindstone Creek, D46/f0054, Waihoaka Formation, Duntroonian, Late Oligocene.
Paratypes: Seven specimens, OU22811, Cosy Dell, F45/f0396, Chatton Formation.
Etymology: Named in honour of Trevor Worthy, Flinders University, Adelaide, in recognition of his many contributions to the knowledge of fossil vertebrates in New Zealand.
Diagnosis: OL:OH = 1.25–1.3. Inner face distinctly convex. Sulcus with wide and large ostium and tapering cauda. CaL:OsL = 0.8–0.95.
Description: Large otoliths up to 11.6 mm (holotype). Specimens below 5 mm length are not diagnostically mature and hence the following description is based on large specimens only. OH:OT = 4.0. The dorsal rim is somewhat variably expressed, but usually shows more or less well developed pre- and postdorsal angles. In one specimen, the predorsal rim is depressed and the postdorsal angle pointed as a result (A). The ventral rim is more deeply curved than the dorsal rim and more regular, with rounded and closely spaced pre- and postventral angles; the preventral angle is usually the deepest. The anterior tip shows a moderately projecting and broadly rounded rostrum. Antirostrum and excisura are minute or absent. The posterior tip is moderately projecting and slightly more pointed than the rostrum (damaged in the holotype). It is usually positioned higher than the rostrum.
The inner face is markedly convex and shows a distinctly supramedian sulcus, which is anteriorly open and posteriorly terminates very close to the posterior tip of the otolith. The ostium is very wide; about twice as wide as, and distinctly longer than, the cauda. It is filled by a well defined, shallow colliculum. The cauda is slightly deepened and posteriorly tapering, and also filled with a well defined colliculum. The dorsal depression is wide and large, but only well marked towards the sulcus by a crista superior. There is no or only a faint indication of a ventral furrow. The outer face is concave and almost smooth in large specimens and convex and intensely ornamented in smaller ones.
Variability and ontogeny: There is some variation in the expression of the dorsal rim, which usually shows a distinct predorsal angle but is depressed in one large specimen (A). Also, the degree of irregular undulation of the dorsal rim is variable.
Centroberyx otoliths are known for a large degree of ontogenetic alteration as first described by Schwarzhans (Citation1980). Unfortunately, some of the observed ontogenetic allometry then discussed was based on incorrectly identified Holocene specimens and subsequently became adjusted in Schwarzhans (Citation2012). Nevertheless, in the case of C. worthy, one can observe the following ontogenetic changes. Small specimens are intensely crenulated along the rims and show a strong ornamentation on the outer face, while large specimens are much more smooth. Small specimens are thicker than large ones (OH:OT = 2.7 vs 4.0) with a convex outer face, which becomes concave in large specimens. Finally, the width of the ostium increases distinctly with size. All in all, specimens below a length of 5 mm may not be identifiable to the species level if found isolated without associated large specimens.
Discussion: Centroberyx otoliths are well known since the Late Cretaceous and species-rich during the Paleogene (Schwarzhans Citation2010, Citation2012, Schwarzhans and Bratishko Citation2011). During the Neogene, their abundance decreases significantly and only few otolith-based data are recorded (Nolf Citation2013). In modern seas, the genus Centroberyx comprises seven species (Froese and Pauly Citation2016), four in temperate Australia and one in New Zealand. The closest to C. worthyi is C. affinis (Günther 1859) (figured in Schwarzhans Citation1980), which is also the only living species occurring in New Zealand waters. Centroberyx worthyi differs from C. affinis in the slightly more compressed shape (OL:OH = 1.25–1.3 vs 1.35–1.6, increasing with size) and the absence of a ventral furrow. Both show a similar degree of variation of the dorsal rim.
Family Trachichthyidae
Genus Optivus Waite, 1899
Optivus moko n.sp.
(H–J)
Holotype: (H), OU22812, Cosy Dell, F45/f0396, Chatton Formation, Duntroonian, Late Oligocene.
Paratypes: Two specimens, OU22813, same data as holotype.
Etymology: From moko (Māori) = tattoo; referring to the unusual preservation of the holotype that shows clear-cut growth rings on the outer face facilitating in promptu recognition of allometric ontogenetic growth. The name is used as noun in apposition.
Diagnosis: OL:OH = 1.5–1.7 (increasing with size). Dorsal rim shallow, long, reaching forward to about centre of ostium. Tip of rostrum sharp. Cauda slightly bent upwards, terminating close to posterior rim of otolith. CaL:OsL = 0.95–1.05.
Description: Elongate, thin and reach a length of 5.2 mm (holotype). OH:OT = 3.8–4.0. The dorsal rim is shallow with well developed pre- and postdorsal angles and reaches anteriorly to a short antirostrum positioned above the middle of the ostium. The ventral rim is more deeply curved than the dorsal rim and shows distinct and widely spaced pre- and postventral angles. The preventral angle is deepest. The anterior tip shows a strongly projecting, broad rostrum with a sharply pointed tip. Antirostrum and excisura are minute. The posterior tip is moderately projecting, angular and positioned higher than the rostrum. All rims are slightly and irregularly undulating, particularly the dorsal rim.
The inner face is slightly convex and shows a slightly supramedian sulcus, which is anteriorly open and posteriorly terminates close to the posterior tip of the otolith. The ostium is distinctly wider than the cauda; its ventral margin is strongly turned upwards anteriorly towards the rostral tip. It is filled by a well defined, slightly deepened colliculum. The cauda is also slightly deepened, slightly turned upwards posteriorly and with a slightly widened tip. It is also filled with a well defined colliculum. The dorsal depression is narrow and developed only above the anterior part of the cauda. The ventral furrow is distinct in the holotype and runs at considerable distance from the ventral rim of the otolith, but is poorly visible in the two paratypes. The outer face is flat and rather smooth.
Ontogeny: The holotype of O. moko offers the rare occasion to study the ontogenetic growth in the same species by growth rings clearly exhibited on the outer face. It shows that an otolith of 3.4 mm length is more compressed than the 5.2 mm long holotype (OL:OH 1.5 vs 1.7), shows more intense marginal crenulation (vs gentle undulation) and exhibits a shorter and blunter rostrum. It also shows that the holotype was derived from a specimen of 5–6 years in age.
Discussion: The otoliths of the trachichthyid genera Aulotrachichthys (considered a subgenus of Paratrachichthys in Koltyar Citation1996), Optivus, Paratrachichthys, Parinoberyx (figure in Kotlyar Citation1996) and Sorosichthys all look rather similar and it is difficult to identify diagnostic features at the genus level. Those of Paratrachichthys and Aulotrachichthys are more elongate and appear to be characterised by an anteriorly reduced and/or depressed dorsal rim starting behind the middle of the ostium (otoliths of P. trailli (Hutton 1875) figured in Schwarzhans Citation1980, of Aulotrachichthys sajademalensis (Kotlyar 1979) in Kotlyar Citation1996, and of Aulotrachichthys prosthemius (Jordan & Fowler 1902) in Ohe Citation1985. Those of Parinoberyx and Sorosichthys are more compressed. Optivus contains three species along southern Australia and New Zealand, with O. elongatus (Günther 1859) (figured in Schwarzhans Citation1980) endemic to New Zealand. It is characterised by a somewhat deepened ostial colliculum, a feature also found in O. moko. This, and the overall similarity in the otolith outline and the cauda reaching relatively close to the posterior rim of the otolith all match with O. moko and hence support its allocation as the first fossil record of the genus. Optivus moko differs from O. elongatus in the shallower dorsal rim, the less reduced ostial colliculum and the slightly more elongate shape in otoliths above 5 mm length (OL:OH = 1.7 vs 1.5).
Order Scorpaeniformes
Family Platycephalidae
Genus Platycephalus Bloch 1795
Platycephalus maxwelli Schwarzhans Citation1980
1980 Platycephalus maxwelli – Schwarzhans: figures 431–433
Material: Three specimens; one specimen Cosy Dell, F45/f0396; two specimens Chatton, F45/f9668; all Chatton Formation.
Order Perciformes
Family Serranidae
Material: One unidentifiable juvenile specimen from Cosy Dell, F45/f0396, Chatton Formation.
Family Sillaginidae
Genus Sillago Cuvier 1816
Sillago recta Schwarzhans Citation1980
(A–C)
Figure 8. Perciform otoliths: Sillaginidae, Sparidae, Haemulidae. A–C, Sillago recta Schwarzhans Citation1980, NMNZ S. 047166; NMNZ S. 047169, Pomahaka G45/f0099, Duntroonian (B reversed, C2 = ventral view). D, Dentex sp., OU22815, Cosy Dell, F45/f0396, Duntroonian (D2 = outer face, D3 = ventral view). E–K, Pomadasys? longicaudatus (Schwarzhans Citation1980), OU22816, Cosy Dell, F45/f0396, Duntroonian (E–F reversed, E2 = ventral view, F2 = anterior view, F3 = ventral view, F4 = outer face, J2 = ventral view).
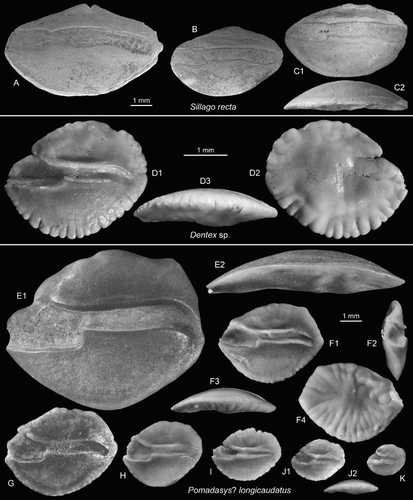
1980 Sillago recta – Schwarzhans: figure 558
Material: Eight specimens from Pomahaka, figured specimens NMNZ S. 047166; NMNZ S. 047169, G45/f0099, Pomahaka Formation.
Discussion: Otoliths of Sillago recta are characterised by a rather narrow sulcus including an only slightly widened ostium and rear part of the cauda. The holotype figured in Schwarzhans (Citation1980) is a relatively large specimen with a low dorsal rim. However, newly collected and yet unpublished material from the same stratigraphical interval (Altonian) show that the dorsal rim is usually somewhat higher and with an obtuse middorsal angle.
The fishes in the family Sillaginidae are typical of very shallow-water environments in the vicinity of fluviatile discharge into the sea and in estuaries. They have not been recorded from any other location in this study other than Pomahaka, which matches their preferred habitat.
Family Sparidae
Genus Dentex Cuvier 1814ß
Dentex sp.
(D)
Material: Five specimens; four specimens, figured specimen OU22815, Cosy Dell, F45/f0396; one specimen Hedgehope Stream, F45/f0410; all Chatton Formation.
Discussion: All five specimens are rather small, up to 3.3 mm length, and have not developed pertinent diagnostic characters. They are considered to stem from juveniles, as also indicated by the very fine crenulation of the ventral rim. Their compressed, high bodied shape and the mildly swung cauda are typical for species of Dentex.
Family Haemulidae
Genus Pomadasys Lacepède 1802
Pomadasys? longicaudatus ( Schwarzhans Citation1980 )
(E–K)
1980 Pomadasyidarum longicaudatus – Schwarzhans: figures 570–572
Material: 504 specimens; 491 specimens, figured specimens OU22816, Cosy Dell, F45/f0396; nine specimens Hedgehope Stream, F45/f0410; four specimens Chatton, F45/f9668; all Chatton Formation.
Description: Compact otoliths reaching sizes of 9.2 mm. OL:OH = 1.15–1.3; OH:OT = 3.0–3.7, increasing with size. The dorsal rim shows a mostly rounded predorsal and a more square-cut postdorsal angle. The ventral rim is deep and regularly curved. The rostrum is moderately long and with a rounded tip and often damaged. Antirostrum and excisura are usually absent or very feeble. The posterior tip is angular, positioned at about the level of the rostrum. All rims are smooth in large specimens and variably undulating or crenulated in smaller specimens.
The inner face is markedly convex and smooth with a slightly supramedian and deepened sulcus. The ostium is ventrally widened with a distinct edge towards the ostial–caudal joint and about twice as wide as the cauda. CaL:OsL = 1.5. The cauda is narrow with a slightly bent tip which closely approaches the posterior tip of the otolith. The dorsal depression is wide and extensive, but with indistinct margins except towards the sulcus. The ventral furrow is mostly feeble and runs close to the ventral rim of the otolith. The outer face is more or less flat, rather smooth in large specimens and with many long radial furrows in smaller specimens.
Ontogeny: The abundance of this species at Cosy Dell has furnished a complete sequence of sizes exemplifying the entire allometric ontogenetic growth. Very small specimens below 2 mm length exhibit more thickset otoliths with a less strongly bent inner face and a rather short and straight cauda. These also show little ornamentation. These otoliths are considered to represent very young fishes and, when found in isolation, could easily be taken for a different species. From c. 2 to 2.5 mm onwards, the otoliths of P.? longicaudatus already exhibit the diagnostic sulcus shape and otolith proportions, but the dorsal rim is more gently curved and all rims and the outer face are strongly ornamented. The dorsal rim gets its angular pre- and postdorsal shape in otoliths of c. 4.5–5 mm length and the slight swinging of the caudal tip increases. Specimens of 7–7.5 mm onward show no more fine crenulation on rims or the outer face and a somewhat more undulating dorsal rim with less clearly discernible pre- and postdorsal angles.
Discussion: Pomamdasys? longicaudatus has rarely been observed in the Late Oligocene to Early Miocene of New Zealand, but at Cosy Dell it represents by far the dominant species. This strongly indicates a neritic faunal assemblage near a rocky shore at that location. Most haemulid otoliths show a rather strongly downward bent of the caudal tip, but this is less so in some genera, for instance Brachydeuterus, Diagramma or Pomadasys, which has led us to tentatively assign this species to the latter genus.
Family Pinguipedidae
Genus Parapercis Bleeker 1863
Parapercis finlayi Frost Citation1924
1924 Parapercis finlayi – Frost: figure 20
1928 Cottus otiakensis – Frost: figure 12
1980 Parapercis finlayi Frost Citation1924 – Schwarzhans: figures 437–441
Material: Four specimens Chatton, F45/f9668, Chatton Formation.
Parapercis fatuus Schwarzhans Citation1980
1980 Parapercis fatuus – Schwarzhans: figures 447–448
Material: Two specimens Chatton, F45/f9668, Chatton Formation.
Family Percophidae
Genus Hemerocoetes Valenciennes 1837
Hemerocoetes minutus Schwarzhans Citation1980
(A–C)
Figure 9. Perciform otoliths: Hemerocoetidae. A–C, Hemerocoetes minutus Schwarzhans Citation1980, A = OU22817 (reversed), Cosy Dell, F45/f0396, Duntroonian (A2 = ventral view); B,C = OU22818, Chatton, F45/f9668, Duntroonian. D–F, Krebsiella chattonensis n.sp., Chatton, F45/f9668, Duntroonian, F = holotype, NMNZ S.46921 (F2 = anterior view, F3 = ventral view); D,E = paratypes (reversed), NMNZ S.46922-23 (D2 = ventral view). G–I, Waitakia robusta Schwarzhans Citation1980, G = OU22819 (reversed), Cosy Dell, F45/f0396, Duntroonian; H,I = NMNZ S.46924 (H reversed), Chatton, F45/f9668, Duntroonian.
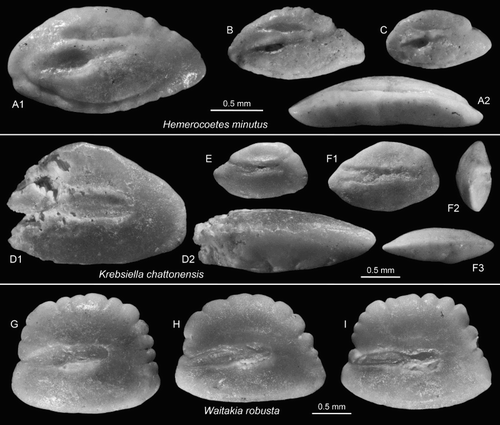
1980 Hemerocoetes minutus – Schwarzhans: figures 471–475
Material: 17 specimens; four specimens, figured specimen OU22817, Cosy Dell, F45/f0396; 13 specimens, figured specimens OU22818, Chatton, F45/f9668; all Chatton Formation.
Discussion: Hemerocoetes minutus was a common species in shallow-water environments of New Zealand in the Late Oligocene and Early Miocene. The otoliths show a number of typical characters of the genus: outline with the dorsal rim being more strongly bent than the ventral rim and both without angles, the distinct and deep ventral furrow curving upwards and away from the ventral otolith rim distally, and the ostium being much longer and somewhat wider than the cauda; showing as a sharply bounded deepening of part of the cauda.
Genus Krebsiella Schwarzhans Citation1980
Krebsiella chattonensis n.sp.
(D–F)
Holotype: (F), NMNZ S.46921, Chatton, F45/f9668, Chatton Formation, Duntroonian, Late Oligocene.
Paratypes: Four specimens, NMNZ S.46922-23, same data as holotype.
Etymology: Named after the type-location and the type-formation – Chatton.
Diagnosis: OL:OH = 1.6–1.7; OH:OT = 1.9–2.0. Inner and outer faces equally strongly convex. Sulcus positioned horizontally, not inclined.
Description: Small, compact and thick. The largest specimen is damaged anteriorly (D) and may have had a length of c. 2.6 mm; the holotype is 1.4 mm long. The dorsal rim is moderately high with variably strong pre- and postdorsal angles. The ventral rim is nearly as deeply curved as the dorsal rim and is more regular. The rostrum is massive, moderately long and with a blunt tip; antirostrum and excisura are not developed. The posterior tip is blunt and clearly inferior with its tip close to the ventral margin.
The inner and the outer faces are similarly and markedly convex. The sulcus is shallow, straight, not inclined, anteriorly open and rather short (OL:SuL = 1.3–1.4). The ostium is slightly wider and longer than the cauda and filled by a clearly defined colliculum. OsL:CaL = 1.2–1.4. The cauda is slightly deeper than the ostium and filled with a less well defined colliculum. The dorsal depression is small and indistinct; the ventral furrow is rather feeble and runs very close to the ventral margin of the otolith except distally, where it turns upwards and away from it. The convex outer face is smooth.
Discussion: Krebsiella chattonensis differs from the other two known species of this fossil otolith-based genus primarily in thickness, the markedly convex outer face, and the horizontally positioned sulcus.
Genus Waitakia Schwarzhans Citation1980
Waitakia robusta Schwarzhans Citation1980
(G–I)
1980 Waitakia robusta – Schwarzhans: figures 483–485
Material: 25 specimens; three specimens, figured specimen OU22819, Cosy Dell, F45/f0396; 22 specimens, NMNZ S.46294, Chatton, F45/f9668; all Chatton Formation.
Discussion: Waitakia robusta is an easily recognisable species by its otolith outline and sulcus morphology, which is widely distributed in the Late Oligocene and Early Miocene of New Zealand.
Family Leptoscopidae
Genus Lesueurina Fowler 1908
Lesueurina transoceana n.sp.
(A–H)
Figure 10. Perciform otoliths: Leptoscopidae, Uranoscopidae. A–H, Lesueurina transoceana n.sp., Cosy Dell, F45/f0396, Duntroonian, B = holotype, OU22820 (reversed) (B2 = ventral view, B3 = anterior view); A, C–H = paratypes, OU22821 (A, D, E, G reversed) (C1 = ventral view, F2 = ventral view). I,J, Uranoscopus sp., OU22822 (I reversed), Cosy Dell, F45/f0396, Duntroonian (I2 = ventral view).
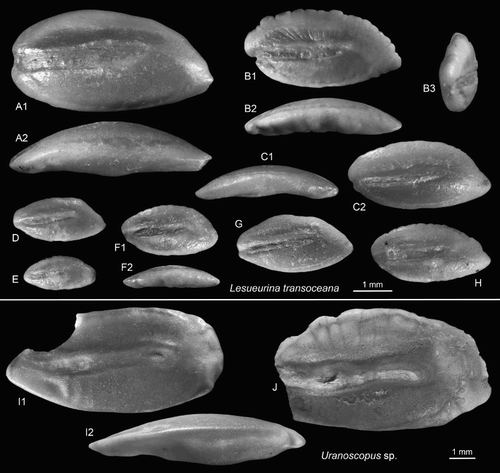
Holotype: (B), OU22820, Cosy Dell, F45/f0396, Chatton Formation, Duntroonian, Late Oligocene.
Paratypes: 30 specimens, OU22821, same data as holotype.
Etymology: From trans (Latin) = across and oceanus (ocean) referring to the occurrence of the fossil species in New Zealand compared to the only extant species being restricted to Australia ‘across the ocean’.
Diagnosis: OL:OH = 1.9–2.0. Inner faces strongly convex. Rounded anterior tip usually shifted dorsally above sulcus. Ventral furrow close to ventral rim of otolith for the central part.
Description: Elongate and moderately thick, reaching 5.1 mm length (holotype 4 mm long). The dorsal rim is shallow, without pre- and postdorsal angles, posteriorly gently inclined and anteriorly projecting forward into a distinctly supramedian, rounded anterior tip positioned above the sulcus. It is variably ornamented, either smooth or very finely crenulated. The ventral rim is more regularly curved, anteriorly reduced below the ostial opening and posteriorly much projecting into a distinctly inferior and variably pointed tip.
The inner face is distinctly convex. The sulcus is shallow, more or less straight, and distinctly inclined upwards towards posterior at an angle of 5–10°. The index OL:SuL is 1.5–1.6. The ostium opens anteriorly, but usually in an indistinct manner with the ostial colliculum clearly separated from the anterior rim of the otolith. It is distinctly longer but not wider than the cauda and equally shallow. The distinction of the ostial and caudal colliculi is often poorly discernible. The index OsL:CaL is very variable, ranging from 1.5 to 2.2, probably owing to the poor distinction of ostium and cauda and its respective colliculi. The dorsal depression is narrow and long. The ventral furrow is mostly well developed; for most of its course it runs close to the ventral margin of the otolith except near the anterior tip, where it bends up slightly, and distally, where it turns upwards more strongly and curves forward towards the caudal tip. The flat to slightly concave outer face is mostly smooth.
Discussion: Today, Lesueurina is a monotypic genus with L. platycephala Fowler 1908 living on turbulent sandy beach zones of warm to temperate Australia (Froese and Pauly Citation2016). The occurrence of L. transoceana from the Late Oligocene of New Zealand is confined so far to the Cosy Dell site, where it is the third most common species after Pomadasys? longicaudatus and the two species of Tonganago. This similarly indicates a very specialised environmental adaptation. Lesueurina transoceana otoliths differ from L. platycephala in the less elongate shape (OL:OH = 1.9–2.0 vs 2.2), the anterior tip projecting above the sulcus and the main part of the ventral furrow running very close to the ventral rim of the otolith.
Family Uranoscopidae
Genus Uranoscopus Linnaeus 1758
Uranoscopus sp.
(I,J)
Material: Two large but incomplete specimens (8.3 mm and 8.2+ mm long) from Cosy Dell, OU22822, F45/f0396, Chatton Formation.
Description: Both large specimens are missing part of the anterior region of the otolith. The shape is elongate, moderately pointed anteriorly and with a broadly rounded or blunt posterior tip. OL:OH = 2.1; OH:OT = 2.0. The dorsal rim is gently curving without angles and somewhat irregularly undulating. The ventral rim is less strongly curved and smooth.
The inner face is moderately convex with a moderately long, sinuate, slightly posteriorly upward inclined and moderately deep sulcus. The ostium is considerably longer than the cauda, anteriorly narrowed and apparently closed or only indistinctly opened. It is slightly widened towards the ostial–caudal joint. OL:SuL = 1.4; OsL:CaL = 1.4. The cauda shows a rounded termination at some distance from the posterior tip of the otolith. The dorsal depression is long, narrow and indistinct. The ventral furrow is feeble, runs close to the ventral rim of the otolith and turns upwards and away from it only below the tip of the cauda. The outer face is flat to moderately convex and smooth.
Discussion: This is a typical uranoscopid otolith and represents an undescribed fossil species. There are, however, better specimens available that will be described in the forthcoming review of the fossil marine otoliths of New Zealand being prepared by WS.
Family Odontobutidae
Genus Micropercops Fowler & Bean 1920
Micropercops pomahaka n.sp.
(A,B)
Figure 11. Perciform otoliths: Odontobutidae. A,B, Micropercops pomahaka n.sp., Pomahaka G45/f0099, Duntroonian, A = holotype, NMNZ S.047166a (A2 = anterior view); B = paratype, NMNZ S.047166b. C,D, Micropercops cinctus (Dabry de Thiersant 1872), USNM 336883, extant from Peking, China, 40°19′N, 116°37′E (C2 = ventral view, C3 = anterior view). E, Odontobutis potamophila (Günther 1861), NSMF 2481, extant from China. F, Odontobutis obscura (Temminck & Schlegel 1845), extant from Japan (F1 = anterior view, F3 = ventral view). G, Perccottus glenii Dybowski 1877, BMNH 99.7.25.9, extant from Russia, Amur River.

Holotype: (A), NMNZ S.047166a, Pomahaka, G45/f0099, Pomahaka Formation, Duntroonian, Late Oligocene.
Paratype: 1 specimen, NMNZ S.047166b, same data as holotype.
Etymology: Referring to the type locality Pomahaka; noun used in apposition.
Diagnosis: Subtrapezoidal outline with projecting preventral and strongly postventral tips. OL:OH = 1.1–1.2. Inner face flat. Sulcus wide, with long ostium and short cauda, inclined at c. 5°. Ostium closely approaching anterior rim. Cauda without distinct subcaudal iugum.
Description: These are small, high bodied otoliths of 1.5 mm length (holotype 1.15 mm). The dorsal rim is much shorter than the ventral rim resulting in a subtrapezoidal outline of the otolith. The pre- and postdorsal angles are well developed and slightly expanded. The middorsal rim is nearly straight or slightly undulating and subhorizontal. The anterior rim is inclined towards a well-developed preventral projection at c. 80–85°. The holotype shows a broad concave anterior rim, which is deepest at the level of the ostium. The posterior rim is inclined at a shallower angle of 65–70° and shows a well projecting postventral tip. It is concave in the portion above the cauda.
The inner face is nearly flat with a centrally positioned, wide and shallow sulcus. The sulcus shows the typical sole-shaped gobioid outline where ostium and cauda are not clearly defined and filled by a fused colliculum. The ostium closely approaches the anterior rim of the otolith. The ostial lobe is rounded and shallow. There is no distinct subcaudal iugum, although both specimens show a broad area below the cauda that appears to be separated from the ventral field by an indistinct furrow. The dorsal depression is wide, but rather indistinct. Likewise, the ventral furrow is feeble. The outer face is slightly convex and smooth.
Discussion: The subtrapezoidal outline of the otolith with the much projected postventral tip in combination with the sulcus showing the low ostial lobe and being positioned close to the anterior margin are very typical for odontobutid otoliths. For comparison reasons, otoliths are shown of the Holocene species Micropercops cinctus (Dabry de Thiersant 1872) (C,D), Odontobutis obscura (Temminck & Schlegel 1845) (F), Odontobutis potamophila (Günther 1861) (E) and Perccottus glenii Dybowski 1877 (G). However, most odontobutid otoliths show similarities with gobiids in that they have a broad subcaudal iugum (C1, F2 and G), which varies in delimitation and usually is less distinct. Many otoliths of the family Eleotridae also have a similar otolith outline and show a sulcus which closely approaches the anterior rim of the otolith (see Schwarzhans et al. Citation2012 for figures), but they have no subcaudal iugum, a sulcus, which is not or only slightly inclined and rather narrow. Otoliths of certain eleotrid genera, such as Gobiomorphus from New Zealand and southern Australia (see Schwarzhans et al. Citation2012 for figures) are similar in outline at times, but differ in the sulcus not closely approaching the anterior rim of the otolith and in the absence of a subcaudal iugum. Otoliths of the New Zealand family Thalasseleotridae (unpublished material in the collection of Schwarzhans) also show a (small) sulcus located far from the anterior rim of the otolith. The Odontobutidae are a group of ‘primitive’, early-diverged gobioid fishes that are essentially demersal fishes of the freshwater and brackish water habitats of Southeast Asia from Thailand through China to the Amur River scheme, Japan and the Sea of Okhotsk. Their occurrence in the Late Oligocene of New Zealand is certainly unexpected, but indicates that they have been more widely distributed in the geological past. The discovery of a New Zealand species of Micropercops is of particular interest because of the great geographical separation of the fossil and Holocene species.
Order Pleuronectiformes
Family Bothidae
Genus Arnoglossus Bleeker 1862
Arnoglossus longus Schwarzhans Citation1980
1980 Arnoglossus longus – Schwarzhans: figures 577–578
Material: 10 specimens; two specimens Cosy Dell, F45/f0396; eight specimens Chatton, F45/f9668; all Chatton Formation.
Results and discussion
Composition of the fish fauna ()
The seas around Zealandia today are populated by a rich fish fauna with 1262 currently recognised species whose distribution reflects the high climatic and ecological diversity of the region (Roberts et al. Citation2015). New Zealand today extends across 2800 km from subtropical seas in the north around the Kermadecs (29°14′S) to cool temperate and subantarctic seas in the south around Campbell Island (52°37′S). It is separated from Australia, the nearest continent, to the west by the Tasman Sea which varies in width between 1500 and 2000 km.
During the Late Oligocene, the Southland Basin was positioned an estimated 2–3° further north than its current position, occupying a warmer climatic belt than it does today (King Citation2000). Plant fossil evidence indicated subtropical rainforest vegetation near Cosy Dell (Conran et al. Citation2014) and an abundance of warm-water species were identified in the marine environment (Lee et al. Citation2014). This could point to the influence of a warm-water eddy from the west as assumed similarly for Middle Miocene times (Field et al. Citation2002). The composition of the otolith-based fish fauna supports the assumption of a subtropical climate at the time. Fishes of the families Moringuidae and Haemulidae, and the genera Heteroconger (garden eels), Neobythites (Ophidiidae) and Dentex (Sparidae) are all warm-water groups no longer present in New Zealand waters. Conversely, cool-water environment species are largely missing. In particular, fishes of the Lotidae, Gadidae, certain scorpaeniform groups and Notothenioidei have not been found in the Oligocene/Early Miocene of Zealandia. A notable exception however is Macruronus merlucciiformis, a temperate Southern Ocean mesopelagic fish.
The largest number of otoliths were obtained from Cosy Dell (c. 640) followed by Chatton (100) with all other localities having contributed fewer than 20 specimens each. Cosy Dell yielded 20 species, and Chatton 16, representing a remarkably high degree of diversity compared with the sample size. However, the diversification index (the sum of the most common species up to 90% of the total association) is low at Cosy Dell with only four species, of which Pomadasys? longicaudatus alone makes up for 76%. The diversification index is considerably higher at Chatton with 10 species.
Late Oligocene endemism of the Zealandian marine fish fauna
The geographic isolation of Zealandia from other continental landmasses is expressed in a very high level of endemism (21.7% of the fish fauna), particularly in shallow marine zones down to 100 m and among freshwater fishes (c. 92%) (Roberts et al. Citation2015). The Late Oligocene fish fauna from the Southland Basin is also already rich in groups that are linked to extant species. Fifteen of 32 species (47%) belong to genera that are still present in New Zealand waters. One genus is endemic to New Zealand (Hemerocoetes), one to southern Australia and New Zealand (Optivus) and a third is now endemic to southern Australia (Lesueurina). Four more species belong to genera that today have an anti-tropical distribution pattern and are represented in the southern Pacific in New Zealand and Australia, and the northern Pacific in Japan or are related to an endemic group within a genus, i.e. Lotella latidorsalis, Trachyrincus tewaewae, Eurypleuron debilis and Centroberyx worthyi. Notably, seven species from the Late Oligocene of the Southland Basin relate to endemic genera or species of present-day Zealandia. Finally, there are four extinct otolith-based genera (Tonganago, Aotearichthys, Krebsiella and Waitakia) with five species, none of which are known from outside Zealandia. There is good reason to assume that these genera also represent (fossil) Zealandian endemics. We assume that 12 species of those identified in the Late Oligocene of the Southland Basin represent endemics in one way or another. Discounting the species in open nomenclature (i.e. 27 taxa identified to species level), this translates into a remarkable 44% of the fish fauna with clear links to the modern Zealandian fauna.
Outsiders to the present day New Zealand fish fauna
Nine species belong to genera not represented in New Zealand waters today. Some are warm-water groups common in the tropical Indo-West Pacific and their absence today is most likely primarily climatic. These are: Moringua waimumuensis, Heteroconger? mataura, Neobythites lindqvisti, Dentex sp. and Pomadasys? longicaudatus. The remaining four warrant some more specific discussion.
Aotearichthys vestalis represents a fossil genus of the Dinematichthyidae (for definition of the family see Møller et al. Citation2016). There is only one extant dinematichthyid known from New Zealand, Dermatopsis joergennielseni Møller and Schwarzhans Citation2006, which probably was a fairly recent immigrant and is known from the fossil record since the Pleistocene (Grenfell and Schwarzhans Citation1999, Møller and Schwarzhans Citation2006). The otolith of Aotearichthys is characterised by its long cauda (and low index OsL:CaL of 1.35), the depressed anterior-dorsal rim and the shape of the deep and wide incision of the ventral sulcus margin behind the ostial–caudal joint. Similar features are only found in the reef-dwelling Porocephalichthys dasyrhynchus (Cohen & Hutchins 1982), which is endemic to southwestern Australia. We conclude that Aotearichthys represents the link between the highly advanced otolith morphology of Porocephalichthys to such genera as Alionematichthys or Dinematichthys. It thus indicates that an endemic evolution had taken place within dinematichthyids in southern Australia and New Zealand since at least Oligocene times, of which Porocephalichthys dasyrhynchus is the only survivor.
Sillago recta is one of several fossil sillaginid otolith-based species from New Zealand known since Eocene times and persisting until the Early Miocene at least (Schwarzhans Citation1980). Nowadays, this family is not represented in New Zealand, but many species are known from Australia (McKay Citation1985, Citation1992). Thus, Sillago represents another example of a fish with a former trans-Tasman Sea distribution which now is confined to Australia.
Lesueurina is another case of a temperate Australian endemic which in the past had a trans-Tasman Sea distribution. The family Leptoscopidae contains three genera with five extant species, three of which are confined to New Zealand, and two to southern Australia. Lesueurina is the only genus of the three that is no longer present in New Zealand although the fossil record suggests that in the past the distribution of the Leptoscopidae was more continuous across Australia and New Zealand.
Micropercops pomahaka is the most exotic and geographically, the most distantly related of all species identified from the Late Oligocene of the Southland Basin. The family Odontobutidae represents one of the most ‘primitive’ basal groups in the large suborder Gobioidei and their recent representatives are all confined to the fresh and brackish waters of Asia bordering the northwest Pacific. Their otolith morphology also indicates some basal character status with the ostium showing a rather uniform dorsal margin without lobe and its close approach to the anterior rim of the otolith. However, these are also highly diagnostic characters of the family. The occurrence of a species of Micropercops in the Oligocene of New Zealand shows that the Odontobutidae were not always restricted to their current geographical realms. This is surprising as today only two other rather basal gobioid families are known from New Zealand: Eleotridae and Thalassoeleotridae. The Eleotridae are represented by several freshwater species of the genus Gobiomorphus with many endemic species in New Zealand and two in Australia. It was concluded that all the extant Gobiomorphus species in New Zealand were descendants of a single lineage that separated from those two temperate Australian species (Stevens and Hicks Citation2009). However, Schwarzhans et al. (Citation2012) documented that earlier migrants were present in Zealandia when they described the fossil genus Mataichthys, which was established based on an articulated skeleton with in situ otoliths from lake sediments of the Early Miocene Bannockburn Formation. The family Thalassoeleotridae is a small family with two genera and three species which are all exclusive to the near-shore waters of New Zealand and temperate Australia (Gill and Mooi Citation2012). Gill and Mooi consider the Thalassoeleotridae to represent the sister group of the Gobiidae as newly diagnosed by them. There appear to be fossil otoliths pertaining to this family in the Early Miocene of New Zealand (project in progress by WS). Now with Micropercops pomahaka there is yet another group of gobioids documented from New Zealand as the earliest record of the group, demonstrating that the history of these fishes in Zealandia is even more complex than the extant record suggests.
Our findings indicate that marine faunal exchange between New Zealand and Australia in the past may have been more common than the present-day level of endemism in New Zealand would suggest. New Zealand and (southern) Australia may have represented one interchanging faunal province with a highly endemic evolution. How much of the endemic evolution might have been confined to New Zealand or Australia and how much has been interactive cannot be assessed due to the paucity of comparative fossil data from Australia.
The ecological diversity of the fish fauna in the Late Oligocene Southland Basin (; )
Following progressive submergence of the Zealandia landmass after rifting from Gondwana, much of Zealandia became submerged (Landis et al. Citation2008). A transgressive sedimentary system tract was deposited over a large part of Southland, mostly unconformably overlying eroded and peneplaned Mesozoic and older strata (Isaac and Lindqvist Citation1990). How much land may have persisted during this transgressive period remains the subject of ongoing investigations and scientific debate (see Landis et al. Citation2008; Lee et al. Citation2014; Lindqvist et al. Citation2016). The rocky shore reefal environments at Brydone and Cosy Dell (Lee et al. Citation2014) and the fluvial/estuarine environments of the Pomahaka Formation, (Lindqvist et al. Citation2016, Gard and Fordyce Citation2017) support the concept of a land area in central Otago and eastern Southland separating the Southland Basin from the Canterbury Basin (see Lee et al. Citation2014).
Figure 12. Block diagram schematic reconstruction of southern New Zealand during the Late Oligocene and typical fishes per locality/paleoenvironment. Fish icons are drawn based on extant relatives. Individual icons not to scale. The fossil otolith-based genus Waitakia (4) is represented with the same icon as the genus Hemerocoetes.
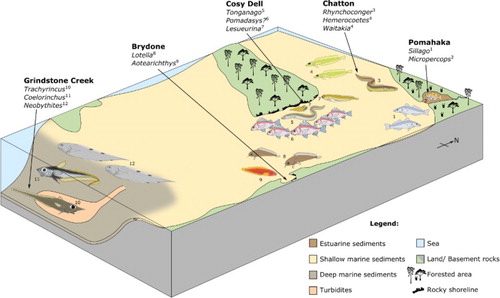
Table 2 Ecologic and habitat characterization of the closest extant relatives of the otolith-based species recognized in the Late Oligocene of the Southland Basin. Fossil otolith-based genera are omitted or referred at family level. Data compiled from Allen & Erdmann (2012), Froese & Pauly (2016), Nielsen et al. (1999), Roberts et al. (2015) and Scott et al. (1974).
The otoliths studied here originate from a variety of paleoenvironments in coastal, basinal and offshore settings ().
Pomahaka represents the most restricted marine environment of all locations studied and is interpreted to represent a paleoestuary at the mouth of a meandering river system draining a land area immediately to the north (Lindqvist et al. Citation2016). The fauna comprises c. 30 species of freshwater and estuarine bivalves and gastropods (including detritus feeders that inhabited intertidal mudflats), foraminifera (suggesting a sheltered, brackish estuarine environment) (Lindqvist et al. Citation2016), a barnacle representative of hard substrates in sheltered intertidal waters (Buckeridge Citation1984, Buckeridge and Newman Citation2010), small crustaceans, a cheloniid turtle (Gard and Fordyce Citation2017) and shark teeth of the family Odontaspididae (Gard Citation2014). Otoliths are rare and have only yielded two species, neither of which have been observed in any of the other localities studied here. The Sillaginidae (Sillago recta) is a characteristic fish of estuarine environments. The other, an odontobutid (Micropercops pomahaka) is remarkable in many ways as noted above. First it is the only record of the ‘primitive’ gobioid family in the Southern Hemisphere, and the only fish studied here that shows clear freshwater to brackish water affinities.
Brydone is stratigraphically slightly younger than the other locations, i.e. Waitakian. The depositional environment is interpreted to be a fully marine, but shallow sheltered bay with intertidal mudflats and reefal areas nearby (Gard Citation2016). The fauna comprises many undescribed species of molluscs including numerous chitons, bivalves and gastropods, including limpets, turbinids, rissooids (Beu and Maxwell Citation1990) and the algal grazer Scutus (Gard Citation2016). Only two otoliths have been found at this location, Lotella latidorsalis and Aotearichthys vestalis. The latter has only been found at Brydone and is of particular interest in understanding the endemic evolution of a branch of southern dinematichthyids (Ophidiiformes) that led to the extant Porocephalichthys dasyrhynchus of southern Western Australia. The Dinematichthyidae live only in very shallow water at depths from 0 to 30 m mostly in tropical to warm temperate climate zones associated with reefs or in sheltered lagoonal environments.
Cosy Dell has been the subject of several publications dealing with the rarely preserved rocky shore setting and the diversity and richness of its fossil biota (Buckeridge et al. Citation2014; Conran et al. Citation2014; Lee et al. Citation2014; Ayress et al. Citation2017). They represent one of the few rocky shore ecosystems preserved in the fossil record (another Late Oligocene otolith-bearing rocky shore environment has been described from Germany by Schwarzhans Citation2008). Cosy Dell has yielded a remarkable array of diverse and exceptionally well-preserved fossils derived from rocky shore, estuarine, sandy beach and shallow subtidal habitats. The fauna is remarkable for its taxonomic richness and diversity, particularly of molluscs; it includes robust, thick-shelled bivalves and large gastropods, oysters in life position on boulders of basement rock, an abundance of juveniles and micromolluscs, and aragonitic molluscs with nacreous shell and colour patterns preserved. More than 350 species of molluscs are present, including 10 chitons, 90 bivalves and 250 gastropods. Other notable components of the biota include nine genera of shallow-water barnacles (Buckeridge et al. Citation2014), foraminifera, brachiopods, bryozoans, echinoderms including Fibularia, solitary and hermatypic corals and a few penguin bones (Lee et al. Citation2014). Juvenile Haliotis and many trochids provide evidence for shallow-water algal grazers, and elongate limpets indicate seagrass meadows (Conran et al. Citation2014).
Otoliths are common and species-rich (20 species), but dominated by a single haemulid species (Pomadasys? longicaudatus) at 76%. This species has not been found in these numbers at any other locality studied. Many haemulids occupy the shallow neritic zone of warm and clear water over rocky and sandy bottom and elevated energy. Other remarkable elements of the fish fauna are the diverse composition and abundance of eel-like fishes and the occurrence of a leptoscopid (Lesueurina transoceana) that has not been found in any other locality. Species of Lesueurina today live on turbulent sandy beach zones of temperate southern Australia (Froese and Pauly Citation2016). Among eel-like fishes, a rare spaghetti-eel (Moringua waimumuensis) is also indicative of reefs or rocky shores (Allen and Erdmann Citation2012) The majority of eels belong to two species of the extinct genus Tonganago. Most of the other otolith-based fish taxa would also have been expected in near-shore shallow-water environments, but are less specialised, or have a wide depth range such as Eurypleuron, Macruronus, Optivus or Centroberyx.
Further from shore, but still in the shallow part of the Late Oligocene Southland Basin is Chatton, the type locality for the Chatton Formation. The diverse molluscan fauna from ‘Shell Gully’ (= Chatton) has been investigated for nearly a century (Marwick Citation1929), and more than 100 species have been described from the shell beds exposed there (Gard Citation2016). The molluscan and benthic foraminiferal faunas indicate a shallow sublittoral setting with chitons and other taxa indicating the presence of a nearby rocky shore. Otoliths collected from Chatton show a generally good comparison with Cosy Dell, but some of the most remarkable high-energy near-shore fishes are either very rare or missing, notably Tonganago, Pomadasys? longicaudatus and Lesueurina. In eels, the occurrence of a putative garden eel (Heteroconger? mataura) is notable. Garden eels have a predilection for open sandy terrain in shallow and sheltered environments (Allen and Erdmann Citation2012). In congrids, the absence of Tonganago is made up by Rhynchoconger otapiriensis. Flatfishes increase slightly in abundance (Arnoglossus longus) again indicating a low-energy shallow-water environment. A remarkable contrast to Cosy Dell is the shift in composition of trachinoid fishes. At Cosy Dell, Lesueurina transoceana is the dominant species in that group, with others (Hemerocoetes minutus and Waitakia robusta) relatively rare. At Chatton, Lesueurina is missing and Hemerocoetes and Waitakia much more common, with a third hemerocoetid, Krebsiella chattonensis. Schwarzhans (Citation1980) noted the sedentary nature and adaption to specific bottoms for several fossil species of this benthic group of fishes, but much more detailed studies are required to fully comprehend the mechanism that is responsible for the observed species replacements. The presence of rare small and unidentifiable myctophid otoliths at Chatton also indicate access to the open sea.
Grindstone Creek is more distant from the localities discussed above, i.e. c. 80 km to the southwest. The sediments of the time-equivalent Waihoaka Formation are thought to have been deposited in a deep marine offshore environment, possibly with influence of turbidites that have redeposited shallow-water sediments and fossils into deep water. The faunal association is remarkable for a mixed assemblage of unusual deep-water molluscs (including Myrtea, Uberella, Gemmula, Cryptodaphne and Waipaoa?) (see Beu and Maxwell Citation1990) and also one of the few New Zealand Late Oligocene occurrences of the shallow-water, large foraminiferans Lepidocyclina and Amphistegina. Only a few otoliths have been retrieved from this location, but their composition is in remarkable contrast to all other localities discussed above. Only one species is shared with any of the taxa recorded from Cosy Dell or Chatton, Centroberyx worthyi, a genus with a wide depth range. All other fish taxa are mesopelagic (Diaphus and Bregmaceros) or bathybenthic (Trachyrincus, Coelorinchus and Neobythites), usually found below 200 m water depth off the shelf break. There is no indication in the otoliths for redeposited shallow-water taxa.
Our study clearly documents that the amazing diversity of the otolith associations observed in these localities is primarily a function of a wide spectrum of paleoenvironments, which seem to represent all major habitats one would expect in the sea that covered the Southland Basin during Late Oligocene. It comprises three different near shore environments, one in an estuarine system (Pomahaka), one in a sheltered bay (Brydone) and one in a rocky shore ecosystem (Cosy Dell). Each of these near-shore environments has its own fish community with certain exclusive adaptations. The database is completed with faunules from more typical shelf environments (Hedgehope, Chatton) and a deep-water offshore location (Grindstone Creek), again each with their own typical fish fauna composition.
Conclusions
The otolith-based fish fauna studied here from the Late Oligocene of southern New Zealand is remarkable for its diversity in composition as a result of the variations of the associated paleoecological environments.
Sampling of otoliths from near-shore sheltered bay, estuarine, rocky shore, low-energy shallow shelf and deep-water environments of the Late Oligocene Southland Basin allows for an unusually complete reconstruction of paleoecosystems and the composition of their fish faunas.
The Late Oligocene fish fauna shows considerable continuity with the extant marine fish fauna of New Zealand. Nearly 50% of the species identified belong to genera also present in New Zealand waters today.
The level of endemism in the 27–23 million year old fish fauna was already high. Including putative extinct endemics, it amounts to nearly 45%. However, the degree of endemic separation to Australia is uncertain.
Certain disjunct extant distribution patterns in the northeast and southeast Pacific date back in Zealandia to the Late Oligocene.
Micropercops pomahaka represents the first Southern Hemisphere odontobutid, a family of ‘primitive’ east Asian freshwater Gobioidei.
The Cenozoic of New Zealand already ranks as one of the most prolific provinces worldwide in the knowledge of fossil otoliths. This study shows that there is still potential for more otolith paleobiodiversity. Fourteen of 31 species described herein are new, despite Late Oligocene to Early Miocene strata having been particularly well searched for otoliths.
Acknowledgements
We thank many landowners including for allowing access across their land, in particular Trevor Copland, and Maree and Bruce Wallace for allowing us to collect fossils from their respective farms. DEL would like to acknowledge helpful discussions about fossil fish and otoliths with Tom Trinski and Laith Jawad, Auckland War Memorial Museum. We thank Dianne Nyhof and Jeffrey Robinson for sorting otoliths and taking photographs respectively. Many people provided help in the field including Uwe Kaulfuss, John Conran, Marcus Richards, Ewan Fordyce, Jon Lindqvist and numerous students. Jon Lindqvist and Bill Lee made helpful comments on earlier drafts of the MS. Luke Easterbrook assisted with diagrams and Dee Roben with formatting of tables. Bruce Marshall, Alan Tennyson and Thomas Schultz enabled us to borrow otoliths from the Museum of New Zealand Te Papa Tongarewa, Wellington. The Department of Geology, University of Otago, Dunedin and GNS Science, Lower Hutt are thanked for the provision of resources to undertake this research.
We wish to acknowledge the great work of the late Phillip Maxwell who collected otoliths from Grindstone Creek and Chatton. We are thankful to colleagues from many institutions having supported one of us (WS) in building of an adequate comparative collection of Holocene otoliths relevant to this study, e.g. M. McGrouther and J. Paxton (AMS), O. Crimmen and J. Maclaine (BMNH), H. Endo and N. Nakayama (BSKU), D. Catania (CAS), R. Feeney and C. Thacker (LACM), K. Hartel (MCZ), P. Béarez (MNHN), S. Andrew (NMNZ), G. Shinohara (NSMT), R. McKay (QM), J. Williams (USNM), G. Allen and S. Morrison (WAM), E. Vasilieva (ZMMGU), J. Nielsen and P. Møller (ZMUC).
Disclosure statement
No potential conflict of interest was reported by the authors.
ORCID
Werner Schwarzhans http://orcid.org/0000-0003-4842-7989
Additional information
Funding
Related Research Data
References
- Allen GR, Erdmann MV. 2012. Reef fishes of the East Indies. University of Hawai’i Press. 1292 pp.
- Ayress M, Robinson JH, Lee DE. 2017. Mid-Cenozoic ostracod biostratigraphic range extensions and taxonomic notes on selected species from a new Oligocene (Duntroonian–Waitakian) fauna from southern New Zealand. Alcheringa: An Australasian Journal of Palaeontology 17:1–12. doi:10.1080/03115518.2017.1297483.
- Betancur R, Broughton RE, Wiley EO, Carpenter K, López JA, Li C, Holcroft NI, Arcila D, Sanciangco M, Cureton JC, et al. 2013. The tree of life and a new classification of bony fishes. PLOS Currents Tree of Life. 45.
- Beu AG, Maxwell PA. 1990. Cenozoic Mollusca of New Zealand: New Zealand Geological Survey Paleontological Bulletin 58: 518 pp.
- Bratishko A, Schwarzhans W, Reichenbacher B, Vernyhorova Y, Coric S. 2015. Fish otoliths from the Konkian (Miocene, early Serravallian) of Mangyshlak (Kazakhstan): testimony to an early endemic evolution in the Eastern Paratethys. Paläontologische Zeitschrift. 89:839–889. doi:10.1007/s12542-015-0274-4.
- Buckeridge JS. 1984. A new species of Elminius from Pomahaka River, Southland, New Zealand. New Zealand Journal of Geology and Geophysics. 27:217–219. doi: 10.1080/00288306.1984.10422528
- Buckeridge JS, Lee DE, Robinson JH. 2014. A diverse shallow-water barnacle assemblage (Cirripedia: Sessilia) from the Oligocene of Southland, New Zealand. New Zealand Journal of Geology and Geophysics. 57:253–263. doi: 10.1080/00288306.2013.873472
- Buckeridge JS, Newman WA. 2010. A review of the subfamily Elminiinae (Cirripedia: Thoracica: Austrobalanidae), including a new genus, Protelminius nov. from the Oligocene of New Zealand. Zootaxa. 2349:39–54.
- Chen W-J, Santini F, Carnevale G, Chen J-N, Liu S-H, Lavoué S, Mayden RL. 2014. New insights on early evolution of spiny-rayed fishes (Teleostei: Acanthomorpha). Frontiers in Marine Science 1: article 53: 17 pp.
- Conran JG, Mildenhall DC, Lee DE, Lindqvist JK, Shepherd C, Beu AG, Bannister JM, Stein JK. 2014. Subtropical rainforest vegetation from Cosy Dell, Southland: plant evidence for Late Oligocene terrestrial ecosystems. New Zealand Journal of Geology and Geophysics. 47:731–748.
- Field BD, Crundwell MP, Kennett JP, King PR, Jones CM, Scott GH. 2002. The early Middle Miocene paleoenvironmental setting of New Zealand. Revista Mexicana de Ciencias Geológicas. 19:242–251.
- Froese R, Pauly D, editors. 2016. FishBase. World Wide Web electronic publication. www.fishbase.org, version (10/2016).
- Frost E. 1924. Otoliths of fishes from the Tertiary formation of New Zealand. Trans. Proc. N.Z. Inst. 55:605–616.
- Frost E. 1928. Otoliths of fishes from the Tertiary formations of New Zealand and from Balcombe Bay, Victoria. Trans. Proc. N.Z. Inst. 59:91–97.
- Frost E. 1933. Otoliths from the Tertiary formations of New Zealand. Trans. Proc. N.Z. Inst. 63:133–142.
- Furlani D, Gales R, Pemberton D. 2007. Otoliths of common Australian temperate fish. CSIRO Publishing. 208 pp.
- Gard HJL. 2014. The paleoecology of the estuarine Pomahaka Formation, West Otago [Unpublished BSc HonsThesis]. University of Otago. 97 pp.
- Gard HJL. 2016. Sedimentology, paleoecology and paleogeography of the shallow marine Chatton Formation, Southern New Zealand [Unpublished MSc Thesis]. University of Otago. 199 pp.
- Gard HJL, Fordyce RE. 2017. A fossil sea turtle (Testudines: Pan-Cheloniidae) from the upper Oligocene Pomahaka Formation, New Zealand. Alcheringa: An Australasian Journal of Palaeontology. 41:134–140. doi:10.1080/03115518.2016.1206319.
- Gill AC, Mooi RD. 2012. Thalasseleotridae, new family of marine gobioid fishes from New Zealand and temperate Australia, with a revised definition of its sister taxon, the Gobiidae (Teleostei: Acanthomorpha). Zootaxa. 3266:41–52.
- Grenfell HR. 1984. Early Miocene teleost otoliths from Parengarenga Harbour, New Zealand. NZ Journal of Geology and Geophysics. 27:51–96. doi: 10.1080/00288306.1984.10422291
- Grenfell HR, Schwarzhans W. 1999. The fish otolith fauna of the Te Piki Member. Proceedings Taupaki Malacological Society. 2:12–14.
- Isaac MJ, Lindqvist JK. 1990. Geology and lignite resources of East Southland Group, New Zealand. New Zealand Geological Survey Bulletin. 101:202.
- King PR. 2000. Tectonic reconstructions of New Zealand: 40 Ma to the present. New Zealand Journal of Geology and Geophysics. 43:611–638. doi: 10.1080/00288306.2000.9514913
- Koken E. 1891. Neue Untersuchungen an Tertären Fischotolithen II. Zeitschrift der deutschen Geologischen Gesellschaft. 43:500–565.
- Kotlyar AN. 1996. Beryciform fishes of the world oceans. Moscow: VNIRO Publishing: 368 pp.
- Landis CA, Campbell HJ, Begg JG, Mildenhall DC, Paterson AM, Trewick SA. 2008. The Waipounamu Erosion Surface: questioning the antiquity of the New Zealand land surface and terrestrial fauna and flora. Geological Magazine. 145:173–197. doi: 10.1017/S0016756807004268
- Lee DE, Lindqvist JK, Beu AG, Robinson JH, Ayress MA, Morgans HEG, Stein JK. 2014. Geological setting and diverse fauna of a Late Oligocene rocky shore ecosystem, Cosy Dell, Southland. New Zealand Journal of Geology and Geophysics. 57:195–208. doi: 10.1080/00288306.2014.898666
- Lin C-H, Chang C-W. 2012. Otolith atlas of Taiwan fishes. NMMBA Atlas Series 12: 415 pp.
- Lindqvist JK, Gard HJL, Lee DE. 2016. Geological setting, sedimentology and biota of the estuarine late Oligocene Pomahaka Formation, New Zealand. New Zealand Journal of Geology and Geophysics. 59:352–365. doi:10.1080/00288306.2016.1150862.
- Marwick J. 1929. Tertiary molluscan fauna of Chatton, Southland. Transactions of the New Zealand Institute. 59:903–934.
- McKay RJ. 1985. A revision of the fishes of the family Sillaginidae. Memoirs of the Queensland Museum. 22:1–73.
- McKay RJ. 1992. Sillaginid fishes of the world. FAO species catalogue. FAO Synopsis. 125(14):1–87.
- Miya M, Nishida M. 2014. The mitogenomic contributions to molecular phylogenetics and evolution of fishes: a 15-year retrospect. Ichthyological Research 62:29–71. doi:10.1007/s10228-014-0440-9.
- Møller PR, Knudsen SW, Schwarzhans W, Nielsen JG. 2016. A new classification of viviparous brotulas (Bythitidae) - with the establishment of a new family Dinematichthyidae – based on molecular, morphological and fossil data. Molecular Phylogenetics and Evolution. 100:391–408. doi: 10.1016/j.ympev.2016.04.008
- Møller PR, Schwarzhans W. 2006. Review of the Dinematichthyini (Teleostei: Bythitidae) of the Indo-West Pacific. Part II. Dermatopsis, Dermatopsoides and Dipulus with description of six new species. The Beagle, Records of the Museums and Art Galleries of the Northern Territory. 22:39–76.
- Møller PR, Schwarzhans W. 2008. Review of the Dinematichthyini (Teleostei: Bythitidae) of the Indo-West Pacific. Part IV. Dinematichthys and two new genera with description of nine new species. The Beagle, Records of the Museums and Art Galleries of the Northern Territory. 24:87–146.
- Nelson JS. 2006. Fishes of the world, 4th ed. John Wiley & Sons. 601 pp.
- Nielsen JG. 1995. A review of the species of the genus Neobythites (Pisces, Ophidiidae) from the western Indian Ocean with descriptions of seven new species. Ichthyological Bulletin. 62:1–19.
- Nielsen JG. 1997. Deepwater ophidiiform fishes from off New Caledonia with six new species. Mémoirs de Musée National de Histoire Naturelle. 174:51–82.
- Nielsen JG. 1999. Atlantic occurrence of the genus Neobythites (Pisces, Ophidiidae) with three new species. Bulletin Maritime Sciences. 64:335–372.
- Nielsen JG. 2002. Revision of the Indo-Pacific species of Neobythites (Teleostei, Ophidiidae) with 15 new species. Galathea Report 19: 107 pp.
- Nielsen JG, Cohen DM, Markle DF, Robins CR. 1999. Ophidiiform fishes of the world (Order Ophidiiformes). FAP Species Catalogue 18: 178 pp.
- Nolf D. 2013. The diversity of fish otoliths, past and present. Brussels: Royal Belgian Institute of Natural Sciences. 222 pp.
- Ohe F. 1985. Marine fish-otoliths of Japan. Special volume of Bulletin (Earth-Science), the Senior High School attached to the Aichi University of Education: 184 pp.
- Pocknall DT. 1982. Palynology of late Oligocene Pomahaka estuarine bed sediments, Waikoikoi, Southland, New Zealand. New Zealand Journal of Botany. 20:263–287. doi: 10.1080/0028825X.1982.10428495
- Roberts CD, Stewart AL, Struthers CD. 2015. The fishes of New Zealand. Wellington: Te Papa Press. 1748 pp. (4 volumes).
- Schwarzhans W. 1978. Otolith-morphology and its usage for higher systematical units, with special reference to the Myctophiformes s.l. Mededelingen Werkgroep Tertiaire en Kwartaire Geologie. 15:167–185.
- Schwarzhans W. 1980. Die tertiäre Teleosteer-Fauna Neuseelands, rekonstruiert anhand von Otolithen. Brliner Geowissenschaftliche Abhandlungen A26: 211 pp. [English translation published in 1984: Fish otoliths from the New Zealand Tertiary. New Zealand Geological Survey Report 113: 269 pp.].
- Schwarzhans W. 1994. Sexual and ontogenetic dimorphism in otoliths of the family Ophidiidae. Cybium. 18:71–98.
- Schwarzhans W. 2008. Otoliths from upper oligocene near shore deposits of the lower rhine embayment (Northern Germany). Neues Jahrbuch Geologie und Paläontologie Abhandlungen. 248:11–44. doi: 10.1127/0077-7749/2008/0248-0011
- Schwarzhans W. 2010. The otoliths from the Miocene of the North Sea Basin. Weikersheim/Leiden: Backhuys/Margraf, 352 pp.
- Schwarzhans W. 2012. Fish otoliths from the Paleocene of Kressenberg, Bavaria, and Kroisbach, Austria. Palaeo Ichthyologica. 12:88.
- Schwarzhans W, Ahnelt H, Carnevale G, Japundžić S, Bradić K, Bratishko A. 2017a. Otoliths in situ from Sarmatian (Middle Miocene) fishes of the Paratethys. Part III: tales from the cradle of the Ponto-Caspian gobies. Swiss Journal of Palaeontology 136:45–92. doi:10.1007/s13358-016-0120-7.
- Schwarzhans W, Bratishko A. 2011. The otoliths from the middle Paleocene of Luzanivka (Cherkasy district, Ukraine). Neues Jahrbuch Geologie und Paläontologie Abhandlungen. 261:83–110. doi: 10.1127/0077-7749/2011/0154
- Schwarzhans W, Carnevale G, Bannikov AF, Japundžić S, Bradić K. 2017b. Otoliths in situ from Sarmatian (Middle Miocene) fishes of the Paratethys. Part I: Atherina suchovi Switchenska, 1973. Swiss Journal of Palaeontology 136:7–17. doi:10.1007/s13358-016-0114-5
- Schwarzhans W, Carnevale G, Bratishko A, Japundžić S, Bradić K. 2017c. Otoliths in situ from Sarmatian (Middle Miocene) fishes of the Paratethys. Part II: Gadidae and Lotidae. Swiss Journal of Palaeontology 136:19–43. doi:10.1007/s13358-016-0114-5.
- Schwarzhans W, Scofield RP, Tennyson AJD, Worthy JP, Worthy TH. 2012. Fish remains, mostly otoliths, from the non-marine early Miocene of Otago, New Zealand. Acta Palaeontologica Polonica. 57:319–350. doi: 10.4202/app.2010.0127
- Scott TD, Glover CJM, Southcott RV. 1974. The marine and freshwater fishes of South Australia. Handbook of the Flora and Fauna of South Australia, South Australian Government: 392 pp.
- Stevens MI, Hicks BJ. 2009. Mitochondrial DNA reveals monophyly of New Zealand’s Gobiomorphus (Teleostei: Eleotridae) amongst a morphological complex. Evolutionary Ecology Research. 11:109–123.
- Stinton F. 1957. Teleostean otoliths from the Tertiary of New Zealand. Transactions of the Royal Society of New Zealand. 84:513–517.
- Suggate RP, Stevens GR, Te Punga MT, editors. 1978. The geology of New Zealand. Wellington: Government Printer. 2 vols, 820pp.
- Turnbull IM, Allibone AH, compilers. 2003. Geology of the Murihiku area. Institute of Geological & Nuclear Sciences 1:250 000 geological map 20. 1 sheet and 74 pp. Lower Hutt, New Zealand. Institute of Geological & Nuclear Sciences Limited.
- Weiler W. 1942. Die Otolithen des rheinischen und nordwestdeutschen Tertiärs. Abhandlungen des Reichsmats für Bodenforschung, Neue Folge 206: 140 pp.
- Wiley EO, Johnson GD. 2010. A teleost classification based on monophyletic groups. In: Origin and phylogenetic interrelationships of teleosts. München: Verlag Dr. Friedrich Pfeil; p. 123–182.
- Wood BL. 1969. Geology of Tuatapere Subdivision, Western Southland. New Zealand Geological Survey Bulletin. 79:1–161.