
ABSTRACT
The genus Palusphaera Lecal-Schlauder emend. R.E. Norris is a distinctive modern coccolithophore that accommodates monomorphic, monothecate coccospheres with one type of spine-bearing heterococcoliths. Having examined a set of scanning electron microscopy images from our collections, we were able to demonstrate that four distinct species of Palusphaera exist in the modern oceans, including the type species Palusphaera vandelii, an informally proposed form, Palusphaera sp. 1 (type robusta) that is herein taxonomically ratified as Palusphaera crosiae sp. nov., and a distinctive morphotype, which is formally described as Palusphaera bownii sp. nov. Biometric analyses and observations on the morphologies of numerous P. vandelii specimens revealed the existence of a further form, and its commonly distinguished coccolith morphotypes are therefore taxonomically separated from Palusphaera vandelii and established as a discrete species, Palusphaera probertii sp. nov.
INTRODUCTION
Coccolithophores are an abundant group of eukaryotic unicellular flagellate phytoplankton in the modern-day oceans, belonging to the phylum Haptophyta, class Prymnesiophyceae. Coccolithophores precipitate a multitude of elaborately shaped scales of calcium carbonate (coccoliths) to form a distinctive mineralized cell covering known as a coccosphere (Wallich Citation1877; Young et al. Citation1992). Although microscopic in size, coccolithophores exert a significant influence on both oceanic biogeochemistry and the global carbon cycle (Sigman & Boyle Citation2000; Hutchins Citation2011; Monteiro et al. Citation2016), and exhibit a vast array of coccolith morphologies as a result of a substantial variety of species-level calcite production (e.g. Monteiro et al. Citation2016).
Coccolithophores are one of the most comprehensively described and reliably identifiable groups of marine phytoplankton (Jordan & Green Citation1994; Jordan et al. Citation1995, Citation2004; Young & Bown Citation1997), and their taxonomy and taxonomic documentation continue to greatly advance, with approximately 280 morphospecies having already been discovered (Young et al. Citation2021). This has primarily been a matter of documenting new taxa from plankton assemblages (e.g. Young et al. Citation2003; Archontikis et al. Citation2020) and/or identifying cells during their life cycle transition (Cros et al. Citation2000; Archontikis & Young Citation2020) but, more recently, crystallographic properties of coccolith calcite and molecular genetics have been applied to calcareous nannoplankton (Young et al. Citation2014; Hagino et al. Citation2016) to reinforce the observations on the coccolith morphologies and correspondingly rectify important taxonomic problems. Yet, there are still several cases of enigmatic taxa with inconsistent generic use and thus their taxonomic treatment is becoming increasingly important to tackle.
The family Rhabdosphaeraceae Haeckel (Citation1894) is a diverse group of coccolithophores definitely including six extant genera (Rhabdosphaera Haeckel, Acanthoica Lohmann, Algirosphaera Schlauder emend. R.E. Norris, Cyrtosphaera Kleijne, Discosphaera Haeckel and Palusphaera Lecal-Schlauder emend. R.E. Norris), one exclusively fossil genus (Blackites W.W. Hay & Towe) and, more tentatively, an additional extant genus Solisphaera Bollmann, M.Y. Cortés, Kleijne, J.B. Østergaard & Jer.R. Young, which, however, is excluded from the subsequent discussion, because it differs in numerous ways from the other genera (see details in Bollmann et al. Citation2006). The family is characterized by the formation of planolith-type coccoliths, with a distinctive rim structure and a more variable central area (), often including the presence of spines (Aubry Citation1988; Varol Citation1989; Kleijne Citation1992; Young et al. Citation2003; Probert et al. Citation2007; van de Locht et al. Citation2014; Bown et al. Citation2017; ). The rim is almost always narrow and formed of two cycles of elements: a broad outer/upper cycle of V-units with subradial sutures and a narrow inner/lower cycle of R-units (e.g. ; see discussion in Bown et al. Citation2017) showing strong sinistral obliquity in proximal view. The central area, excluding the process, is typically formed of two cycles of units. First, a radial lath cycle, adjacent to the rim, composed of well-formed laths often with gaps between them, whose ends interdigitate regularly with the elements of the rim. Second, a lamellar cycle of less regular elements. The lamellar cycle is present in all genera, but the radial cycle is absent in Rhabdosphaera and Palusphaera. The central process is the most variable part of the structure, showing a wide range of shapes and morphologies, and is absent from some of the coccolith types on the coccospheres of Acanthoica and Rhabdosphaera.
Table 1. Description and diagnostic morphologies of all currently known extant Palusphaera species vs other spine-bearing taxa of the family Rhabdosphaeraceae. SEM images are not to the same scale
Figs 1–4. Ultrastructure of Rhabdosphaeraceae
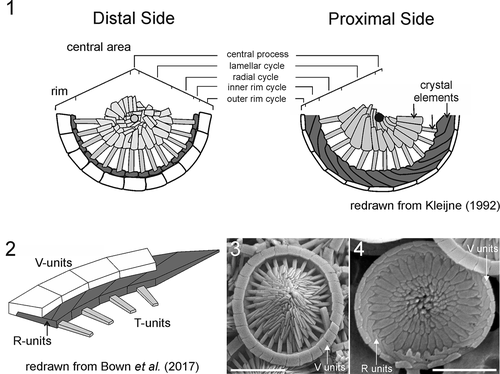
Within this group, Palusphaera is distinguished from the other genera by forming monomorphic coccospheres with body coccoliths, all bearing spines and lacking a radial cycle. Currently, there is only one formally described species in the genus, Palusphaera vandelii Lecal-Schlauder emend. R.E. Norris, but one other morphotype has been reported informally several times. Here, we also report on two further extant morphotypes of the genus and provide formal descriptions of all currently known forms.
MATERIAL AND METHODS
We have examined the scanning electron microscope (SEM) images of extant Palusphaera specimens from our collections. The material used in the present study comes from seawater samples collected in different marine basins () and from several sampling projects carried out over the last two decades. The methodology used here is analytically explained below in chronological order and according to the sample source. Information about all used samples is given in .
Table 2. List of all sampling localities, including physicochemical data, with Palusphaera specimens used in the present study
Fig. 5. Map showing the locations from which Palusphaera specimens were obtained
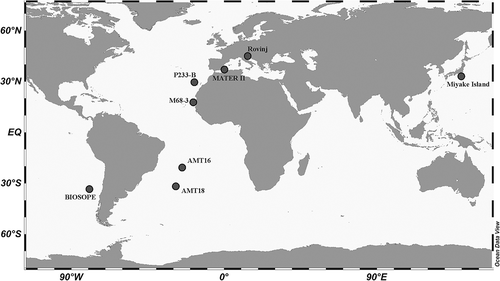
Sample location
ATLANTIC AND PACIFIC OCEANS
Water samples were collected aboard the R/V Poseidon during the P233 Expedition B (Knoll et al. Citation1998) in the Canary Islands, northeastern Atlantic Ocean in September 1997 and during the M68-03 Expedition in the Mauretania upwelling, northeastern Atlantic Ocean onboard the R/V Meteor in April–August 2006 (Koschinsky et al. Citation2009). Specimens of Palusphaera were also found in seawater samples that were obtained from the Miyake Island, western Pacific Ocean in November 1999 and from the eastern Pacific Ocean in October–December 2004 during the oceanographic cruise BIOSOPE aboard the R/V L’Atalante (Claustre & Sciandra Citation2004). Samples also originate from surface waters of the South Atlantic Ocean during the Atlantic Meridional Transect cruises AMT16 (Robinson et al. Citation2006) and AMT18 (Woodward Citation2009) aboard, respectively, the RRS Discovery in May–June 2005 and R/V James Clark Ross in October–November 2008.
WESTERN MEDITERRANEAN SEA
Samples have been obtained from the surface waters of the northwestern Mediterranean and Alboran Seas on board the R/V Hesperides during the oceanographic cruise MATER II in September–October 1999 (Font Citation1999).
ADRIATIC SEA
A time series sample used in the present work comes from the surface waters (5 m water depth) of a coastal station (45°04.8'N, 13°36.6'E) off Rovinj, Croatia, in the northeastern Adriatic Sea in March 2009 aboard the R/V Villa Velebita (as described in Godrijan et al. Citation2018).
Filtration and SEM
Seawater samples were obtained via 5-litre Niskin bottles that were attached to CTD rosette samplers, except for samples originating from the Miyake Island that were collected using a bucket. Approximately 1 litre of seawater was filtered through 0.8-μm pore size Whatman Nuclepore polycarbonate track-etched filters of 25 mm diameter apart from P233-B cruise, during which 4 litres of seawater were filtered onto 47-mm cellulose nitrate filters (Sartorius) of 0.45 μm pore size. A low-pressure vacuum pump was used during the filtration process to prevent mechanical damage of the coccosphere specimens. The filters were subsequently oven-dried at 40°C and stored in 47-mm Millipore plastic Petri dishes; a portion of each filter was cut out and fixed onto aluminium stubs to be gold-coated. Examination of the samples was then performed on a Phillips XL-30 FEG field emission SEM at the facilities of the Natural History Museum, London.
Terminology and morphology
Morphological descriptions are given based on the guidelines of Kleijne (Citation1992) and Young et al. (Citation1997). With regards to the terminology of Rhabdosphaeraceae cycles, we follow the scheme proposed by Kleijne (Citation1992), as also used by Young et al. (Citation2003) and Bown et al. (Citation2017). All morphological measurements presented in this study were obtained using the image analysis software programme ImageJ (Schneider et al. Citation2012).
RESULTS
Separation of P. probertii sp. nov. from P. vandelii
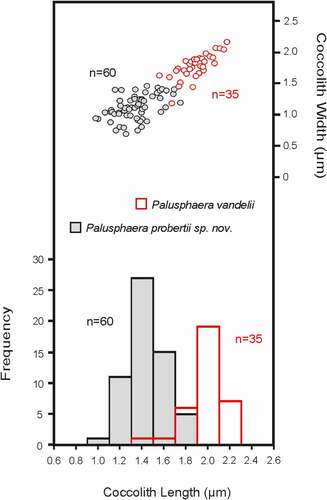
Our examination of SEM micrographs yielded in total 24 P. vandelii-like coccospheres (14 specimens of P. vandelii and 10 specimens of P. probertii sp. nov.; ). These forms, however, were seen to display considerable variations in their coccolith morphologies and diameters. In order to assess this variability, we subjectively separated specimens with typical P. vandelii characteristics (i.e. circular coccolith outline, larger coccolith size and spine structure with spirally oriented elements) vs specimens that bore coccoliths of noticeably more elliptical shape, smaller size and a spine structure with elements parallel aligned to the long axis. We subsequently performed morphometric analyses on several of these specimens and measured 95 (in total) well-preserved body coccoliths (BCs) in plan view; these allowed a straightforward biometric analysis and correspondingly yielded more objective data (Table S1).
There were 35 typical P. vandelii coccoliths from five coccospheres and 60 well-differentiated forms from four well-preserved coccosphere specimens. The morphometric frequency plots () indicated a clear bimodal distribution pattern of all studied specimens with an obvious clustering at 1.3 μm to 2.3 μm (mean 2.0 μm) for P. vandelii and a cluster group at 0.9 μm to 1.9 μm (mean 1.4 μm) for all other specimens. This well-defined separation pattern was further supported by our coccolith width data (), with specimens of P. vandelii demonstrating higher width values (>1.5 μm). Hence, two different coccolith size groups could be seen, essentially confirming our initial morphologic grouping, and we therefore taxonomically separated them and proposed them as two discrete taxa.
Fig. 6. Frequency and bivariate plots of the main morphological parameters (coccolith length and width) measured in specimens of Palusphaera vandelii (image codes 222-04, 289-62, 246-06a 262-20, 247-17 276-30, 280-14) and Palusphaera probertii sp. nov. (image codes 118-55, 188-03, 288-06, 297-77)
Taxonomy
Division Haptophyta D.J. Hibberd (Citation1972) ex Edvardsen & Eikrem in Edvardsen et al. (Citation2000)Class Prymnesiophyceae D.J. Hibberd (Citation1976) emend. Cavalier-Smith et al. (Citation1996)
Order Syracosphaerales W.W. Hay (Citation1977) emend.Jer.R. Young et al. (Citation2003)
Family Rhabdosphaeraceae Haeckel (Citation1894)
Genus Palusphaera Lecal-Schlauder emend. R.E. Norris
emended description
Coccosphere monomorphic and monothecate, with only spine-bearing BCs and no differentiated circum-flagellar coccoliths. BCs flat, with two rim cycles, an outer/upper cycle with near-radial sutures, an inner/lower cycle showing strong sinistral obliquity in proximal view. The radial lath cycle shown by many other Rhabdosphaeraceae is absent; instead, the lamellar cycle fills the central area. The lamellar cycle of tabular elements shows dextrogyral obliquity in distal view and laevogyral obliquity in proximal view. Central process is a spine, variable in shape but without a collar; proximal side with a central pore surrounded by several (usually three) angular nodes.
type Species
Palusphaera vandelii Lecal-Schlauder (Citation1966) emend. R.E. Norris (Citation1984). The original author published under the name Lecal.
Palusphaera vandelii Lecal-Schlauder emend. R.E. Norris
Figs 7–11. SEM micrographs of Palusphaera vandelii. Scale bar = 5 μm for Figs 7, 8, 9, 11
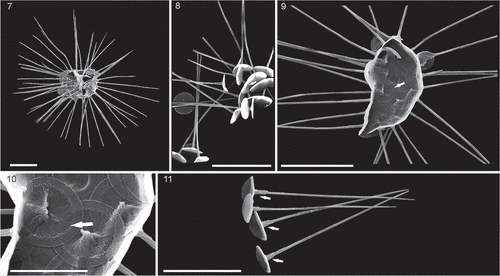
published illustrations of the species
Lecal (Citation1966, pp 68-69, text- fig. K, pl. 2, fig. 9); Lecal (Citation1967, pp 318-320, text-fig. 13, figs 19-20); Norris (Citation1984, p. 35, figs 1f, 9, 10); Kleijne (Citation1992, pp 38-39, pl. 8, fig. 1); Giraudeau & Bailey (Citation1995, p. 1833, pl. 3, fig. 3); Aubry (Citation1999, pp 298-300, figs 3-13); Yang et al. (Citation2001, pp 295-296, pl. III, fig. 8); Cros (Citation2002, p. 36, pl. 9, fig. 5); Cros & Fortuño (Citation2002, pp 24-25, fig. 22B); Young et al. (Citation2003, pp 56-57, pl. 25, fig. 9); Andruleit et al. (Citation2005, p. 12, pl. 2, fig. 4); Kahn & Aubry (Citation2006, pp 319, 334, text-fig. 2c and pl. 2, figs 1-3 non figs 4-6); Kahn (Citation2007, p. 38, pl. 2, figs 1-3 non figs 4-6); Gravalosa et al. (Citation2008, p. 21, pl. I, fig. 4); Malinverno et al. (Citation2008, p. 65, fig. 36); Wang et al. (Citation2012, p. 5, pl. 2, figs G, I); Guerreiro et al. (Citation2014, p. 355, fig. B.17); Malinverno et al. (Citation2015, p. 504, pl. 3, fig. 10); Chang & Northcote (Citation2016, p. 8, fig. 3b); Karatsolis et al. (Citation2017, p. 144, pl. 2, fig. 7); Chang (Citation2019, p. 51, figs 20C, D).
previous records under misapplied names
Acanthoica quattrospina Lohmann sensu Halldal & Markali (Citation1955, pp 15-16, fig. 3, non figs 1, 2, 4). Rhabdosphaera longistylis J. Schiller sensu Norris (Citation1971, p. 902, fig. 4); Kling (Citation1975, p. 6, pl. 3, figs 13-14); Conley (Citation1979, p. 30, pl. 3, fig. 18 and pl. 4, fig. 9); Reid (Citation1980, p. 157, pl. 4, fig. 2 non fig. 3). Halopappus cf. H. adriaticus J. Schiller sensu Winter et al. (Citation1979, p. 200, pl. III, fig. 5). Palusphaera vandelii var. vandelii sensu Dimiza (Citation2006, p. 61, pl. X, figs 3-4; unpublished PhD thesis).
emended description
Coccosphere monomorphic, spherical but usually seen collapsed, composed of c. 50 BCs. BCs planoliths with a subcircular to circular outline; rim thin with typical Rhabdosphaeraceae rim structure. Lamellar cycle fills central area, convex on distal side and concave on the proximal side. Lamellar cycle elements rod shaped, overlapping and displaying dextrogyral curvature on the distal side and laevogyral in proximal view (e.g. ). Central process with a central pore in proximal view, usually surrounded by two to three small nodes; distal side with a thin and long spine, formed of numerous spirally arranged elements at the base becoming less spiral upwards. Spine is styliform-shaped without a collar.
dimensions
Coccosphere diameter c. 5–10 μm excluding processes and c. 25–30 μm with processes. BCs c. 2.0 μm long and c. 1.7 μm wide; rim c. 0.2–0.5 μm. Spine c. 10 μm long.
Palusphaera probertii Archontikis & Jer.R. Young sp. nov.
Figs 12–16. SEM micrographs of Palusphaera probertii sp. nov
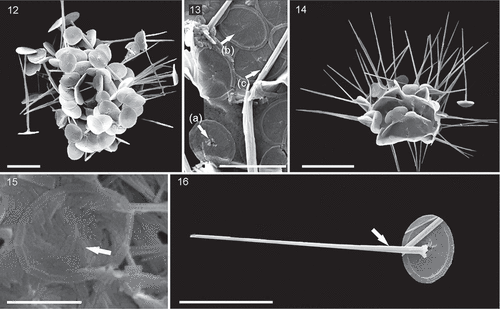
Figs 17–21. SEM micrographs of Palusphaera crosiae sp. nov. Scale bar = 5 μm for Figs 17, 18, 19
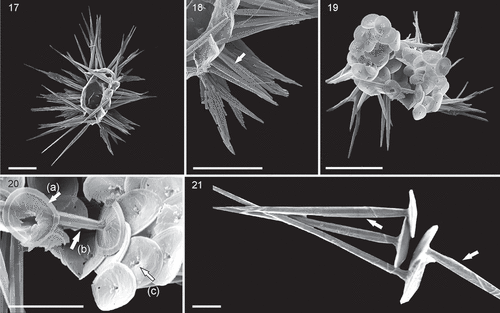
Figs 22–24. SEM micrographs of Palusphaera bownii sp. nov. Scale bar = 2 μm
previous records under misapplied names
Rhabdosphaera longistylis J. Schiller sensu Reid (Citation1980, p. 157, pl. 4, fig. 3). Palusphaera vandelii sensu Cros (Citation2002, p. 36, pl. 9, fig. 3); Cros & Fortuño (Citation2002, pp 24-25, fig. 22A); Yang et al. (Citation2003, p. 38, pl. IV, fig. 1); Young et al. (Citation2003, pp 56-57, pl. 25, fig. 8); Kahn & Aubry (Citation2006, p. 334, pl. 2, figs 4-6); Andruleit (Citation2007, p. 43, fig. 6d); Kahn (Citation2007, p. 38, pl. 2, figs. 4-6); Wang et al. (Citation2012, p. 5, pl. 2, fig. H).
description
Coccosphere, monomorphic and spherically shaped, but usually seen collapsed, composed of c. 45–60 BCs. BCs broadly elliptical to subcircular in plan view, with narrow rims and nearly flat bases. Rim with typical Rhabdosphaeraceae rim structure. Central area filled by a lamellar cycle of numerous rod-shaped, slightly overlapping crystal segments; no radial cycle is observed. Central process bears a thin, hollow, styliform spine with no collar; spine structure composed predominantly of numerous elongate elements arranged parallel to the axis of the spine. The spine is remarkably long compared to the coccosphere diameter. The proximal planolith side has three robust angular nodes around the central pore.
dimensions
Coccosphere diameter c. 7–8 μm excluding processes and c. 13–18 μm with processes. BCs c. 1.5 μm × c. 1.1 μm; rim c. 0.2 μm in length and width. Spine is typically c. 5–8 μm long.
holotype
118-55 (specimen illustrated in ), stub no. 257/0, deposited at the collections of the Natural History Museum, London under the designation PM NF 4591 118-55.
paratypes
212-09 (stub no. 511/3, designation PM NF 4875 212-09); 188-03 (stub no. 459/2, designation PM NF 4814 188-03); 297-77 (stub no. 729/0, designation PM NF 4928 297-77); 212-19 (stub no. 511/3, designation PM NF 4875 212-19). Specimens of these stubs are illustrated by .
type locality
North-eastern Atlantic Ocean (29°45.7'N, 17°55.8'W, depth 25 m, 24 September 1997, P233B Expedition, Station P233b-2).
distribution
Subtropical waters.
number of specimens studied
Ten.
etymology
After Dr. Ian Probert (Station Biologique de Roscoff) in recognition of his many contributions to extant coccolithophore biology and physiology.
remarks
The species is similar to P. vandelii but with noticeably smaller BCs that present a more broadly elliptical shape. The outer rim of the BCs on the distal side is usually narrower than that of P. vandelii, and the styliform central process is hollow and predominantly composed of a single set of very long elements aligned parallel to the long axis. This clearly opposes the spine structure of P. vandelii, which is formed by numerous shorter (c. 1 μm) elements in a spiral arrangement.
Palusphaera crosiae Archontikis & Jer.R. Young sp. nov.
previous records under misapplied names
sp. aff. Palusphaera sensu Kleijne (Citation1992, p. 38, described under remarks on Palusphaera but not illustrated). Palusphaera sp. 1 (type robusta) sensu Cros (Citation2002, p. 36, pl. 9, figs 4, 6); Cros & Fortuño (Citation2002, p. 84, figs 22C, 22D); Young et al. (Citation2003, pp 56–57, pl. 25, figs 10, 11); Malinverno et al. (Citation2008, p. 66, fig. 37). Palusphaera vandelii var. crassa sensu Dimiza (Citation2006, pp 61–62, pl. X, figs 5–6 and pl. XI, figs 1–6, unpublished PhD thesis, invalid).
description
Coccosphere, monomorphic and generally seen collapsed but probably spherical in shape, with c. 45–60 BCs. BC bases broadly elliptical in outline and slightly concavo-convex. Rim shows typical Rhabdosphaeraceae rim structure. Central area filled by the lamellar cycle; no radial cycle is present. Central process is a long, tapering spindle-shaped spine. Maximum thickness is at one-third to one-quarter of the height and it tapers to a fine point. The spine is formed from robust laths, c. 1.0 μm × 0.15 μm at the base, becoming smaller towards the tip; they abutt neatly to form a 6- to 8-μm sided smooth hollow structure. Adjacent laths are offset by about one-third of their length. BC proximal side with a central pore surrounded by a few angular nodes.
dimensions
Coccosphere diameter c. 5–9 μm without processes and c. 15–25 μm including processes. BCs c. 1.2–2.0 μm × c. 1.0 μm; rim is c. 0.2 μm. Spine c. 6 μm long.
holotype
200-07 (specimen illustrated in ), stub no. 470/3, deposited at the collections of the Natural History Museum, London under the designation PM NF 4917 200-07.
paratypes
211-21 (stub no. 487/0, designation PM NF 4855 211-21); 211-22 (stub no. 487/0, designation PM NF 4855 211-22); 177-56 (stub no. 302/2, designation PM NF 4663 177-56). Specimens of these stubs are illustrated by .
TYPE LOCALITY
North-western Mediterranean and Alboran Seas (37°25.98'N, 0°25.3'W, depth 70 m, October 1999, MATER II Expedition, Station 69-08).
distribution
Subtropical waters.
number of specimens studied
Nine, plus two published in Cros & Fortuño (Citation2002).
etymology
After Dr. Lluïsa Cros (Institut de Ciències del Mar, CSIC), who first illustrated the taxon, and in recognition of her many contributions to extant coccolithophore taxonomy.
remarks
The species differs from the other taxa in possessing a styliform spine with robust and thick lath-like crystal segments, markedly thicker at the one-third to one-quarter height from the base of the central area. The distal side outer rim cycle is distinctly narrower compared to that of P. vandelii.
Palusphaera bownii Archontikis & Jer.R. Young sp. nov.
description
Coccosphere, monomorphic of subspherical to spherical shape, usually found collapsed, with c. 80–120 BCs. BCs broadly elliptical to circular in outline with slightly convex planolith bases. Rim shows the typical Rhabdosphaeraceae rim structure. Central area, filled by lamellar cycle, with overlapping tabular elements showing clockwise curvature in distal view. The radial cycle is absent. Central process is a trumpet-shaped spine without a collar, formed of vertically elongate elements arranged in spiralling series; the distal end is formed of blockier elements, but these appear to have developed from the spiral elements of the spine. Trumpet aperture wider than the basal spine width. On the proximal side of the coccolith there is a central pore surrounded by two to three nodes.
dimensions
Coccosphere diameter c. 10–15 μm. BCs 1.0–1.5 μm long and wide; rim c. 0.2 μm in width. Spine c. 1.5 μm long and trumpet aperture c. 0.5 μm wide, although few spines tend to bear slightly closer apertures at their outermost part.
holotype
302-04 (specimen illustrated in ), stub no. 756/1, deposited at the collections of the Natural History Museum, London under the designation PM NF 4930 302-04.
paratype
308-031 (stub no. 768/0, designation PM NF 4932 308-031), 308-032 (stub no. 768/0, designation PM NF 4932 308-032). One specimen of this stub is illustrated by , .
type locality
Subtropical waters of the South Atlantic Ocean (8°49.52'S, 24°59.72'W, depth 0 m, 28 October 2008, AMT18 Expedition, Station CTD063).
number of specimens studied
Two.
etymology
After Professor Paul R. Bown (University College London), in recognition of his many contributions to Jurassic, Cretaceous and Paleogene coccolithophore taxonomy.
remarks
We have only found two coccospheres of this species and to our knowledge no other specimens have been published. Nonetheless, this is a distinctive form and clearly separate from any other coccolithophore, so we are confident it is a discrete species. The species shows possible affinities to several genera. The trumpet-like spines resemble those of Discosphaera, but the base lacks a radial cycle, and the spine shows a structure that is not seen in Discosphaera. The spine structure is closer to that of Rhabdosphaera, but in Rhabdosphaera coccospheres are dimorphic with both spine-bearing and non-spine-bearing BCs. Finally, the species resembles Palusphaera in being monomorphic and lacking a radial cycle. In addition, the rows of the laminar cycle elements show the same curvature (clockwise in distal view) as that of Palusphaera coccoliths and the opposite of that shown by Rhabdosphaera coccoliths. Therefore, we place the species in Palusphaera.
DISCUSSION
The genus Palusphaera is a distinctive extant coccolithophore, known only from the heterococcolith-bearing phase, and based on our observations it now comprises four discrete taxa (), namely, P. vandelii, P. probertii sp. nov., P. crosiae sp. nov. and P. bownii sp. nov., which show a range of shared morphological characteristics. All of these species possess monomorphic and monothecate coccospheres composed of spine-bearing BCs, without differentiated circum-flagellar coccoliths. The BCs consist of solid circular to elliptical planolith bases with a rim and an imbricate central area (see ; ). The rim, as in all typical Rhabdosphaeraceae (e.g. ; Kleijne Citation1992; Bown et al. Citation2017) is formed of two cycles: an outer and upper cycle with radial sutures and an inner and lower cycle with strongly oblique sutures, showing sinistral obliquity in proximal view. Unlike Acanthoica, Algirosphaera, Cyrtosphaera and Discosphaera, there is no radial lath cycle present. Instead, the central area is closed by the lamellar cycle of subrectangular, slightly overlapping crystallites. In proximal view, these can be seen to be arranged in rows showing laevogyral curvature; i.e. they appear bent to the left as they run from the centre to the rim; the proximal surface of the central process typically shows a central pore that is surrounded by a limited number of nodes, usually two to three (e.g. ). In distal view, lamellar cycle segments show dextrogyral curvature but this is less obvious due to overlap of elements. The central elements of the lamellar cycle extend outwards to form an imbricate spine in distal view. The spines are formed of butting segments that disintegrate into separate elements (e.g. , , ), and they do not bear collars at their bases.
Fig. 25. Schematic representation of the coccolith morphology and ultrastructure of all currently known morphospecies in the genus Palusphaera
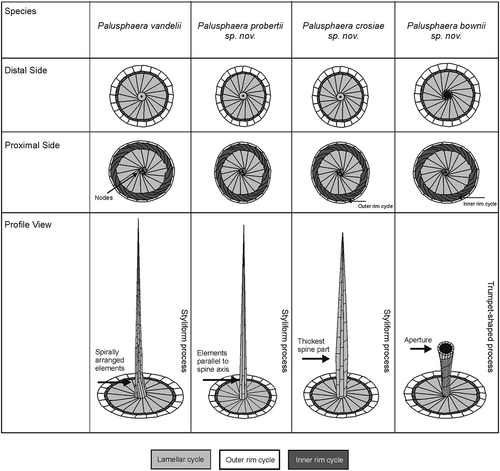
Palusphaera is most obviously separated from Rhabdosphaera by being monomorphic; all of the BCs have spines, whereas in Rhabdosphaera there is an outer layer of non-spine-bearing coccoliths. In addition, however, our observations suggest that two other features consistently separate the four Palusphaera species from the two extant Rhabdosphaera species. First, spines in Rhabdosphaera species are robust solid structures formed of intergrown elements, whereas those of Palusphaera are delicate hollow structures formed of directly abutting elements. Second, the lamellar cycle elements in Rhabdosphaera show the opposite sense of curvature to those in Palusphaera; i.e. they show laevogyral curvature on the distal side and dextrogyral on the proximal side.
Consequently, the observations reported in our study conclusively supported a proposal to emend the description of both Palusphaera and Palusphaera vandelii. In parallel, based on the well-differentiated subset of taxa within the Rhabdosphaeraceae, our results provided the evidence needed to formally maintain Palusphaera separable from its sister taxa and consider it a clearly diagnosable and taxonomically valid genus.
Supplemental Material
Download MS Word (24.4 KB)Acknowledgements
We thank the captains and the crews of R/V Poseidon, R/V James Clark Ross, RRS Discovery, R/V Meteor, R/V L’Atalante, R/V Hesperides and R/V Villa Velebita for their valuable assistance in seawater sampling in the Atlantic and Pacific Oceans and in the Mediterranean and Adriatic Seas. We warmly acknowledge Markus Geisen, Alexandra Broerse, Andy Howard, Claudia Sprengel, Daniel Vaulot and Masanobu Kawachi for their assistance in obtaining seawater samples and/or with the scanning electron microscope analyses. We gratefully acknowledge Jelena Godrijan for providing the sample from the Adriatic Sea used in this study. Special thanks are due to Giles Miller for his assistance in cataloguing our material at the Natural History Museum London, UK. We also extend our gratitude to the editor and the two anonymous reviewers for their insightful suggestions that greatly improved the quality of the article. The work contains data supplied by the Natural Environment Research Council.
SUPPLEMENTARY MATERIAL
Supplemental data for this article can be accessed on the publisher’s website.
Correction Statement
This article has been republished with minor changes. These changes do not impact the academic content of the article.
Additional information
Funding
REFERENCES
- Andruleit H. 2007. Status of the Java upwelling area (Indian Ocean) during the oligotrophic northern hemisphere winter monsoon season as revealed by coccolithophores. Marine Micropaleontology 64: 36–51. DOI: https://doi.org/10.1016/j.marmicro.2007.02.001.
- Andruleit H., Rogalla U. & Staeger S. 2005. Living coccolithophores recorded during the onset of upwelling conditions off Oman in the western Arabian Sea. Journal of Nannoplankton Research 27: 1–14.
- Archontikis O.A. & Young J.R. 2020. Extant Ceratolithus cristatus life-cycle observations and taxonomic simplification. Journal of Nannoplankton Research 38: 27–39.
- Archontikis O.A., Young J.R. & Cros L. 2020. Taxonomic revision and classification of extant holococcolithophores previously placed in the genus Anthosphaera Kamptner emend. Kleijne 1991. Acta Protozoologica 59: 121–139. DOI: https://doi.org/10.4467/16890027AP.20.010.13265.
- Aubry M.-P. 1988. Handbook of Cenozoic calcareous nannoplankton. Book 2: Ortholithae (Catinasters, Ceratholiths, Rhabdoliths). Micropaleontology Press, American Museum of Natural History, New York, New York, USA.
- Aubry M.-P. 1999. Handbook of Cenozoic calcareous nannoplankton. Book 5: Heliolithae (Zygoliths and Rhabdoliths). Micropaleontology Press, American Museum of Natural History, New York, New York, USA.
- Bollmann J., Cortes M.Y., Kleijne A., Østergaard J.B. & Young J.R. 2006. Solisphaera gen. nov. (Prymnesiophyceae), a new coccolithophore genus from the lower photic zone. Phycologia 45: 465–477. DOI: https://doi.org/10.2216/05-14.1.
- Bown P.R., Lees J.A. & Young J.R. 2017. On the Cretaceous origin of the Order Syracosphaerales and the genus Syracosphaera. Journal of Micropalaeontology 36: 153–165. DOI: https://doi.org/10.1144/jmpaleo2016-001.
- Cavalier-Smith T., Allsopp M., Häuber M., Gothe G., Chao E., Couch J. & Maier U.-G. 1996. Chromobiote phylogeny: the enigmatic alga Reticulosphaera japonensis is an aberrant haptophyte, not a heterokont. European Journal of Phycology 31: 255–263. DOI: https://doi.org/10.1080/09670269600651461.
- Chang F.H. 2019. The Marine Biota of New Zealand. Catalogue and description of the coccolithophores (Haptophyta, Coccolithophyceae), calcareous, scale-bearing microalgae, in New Zealand waters. NIWA Biodiversity Memoir 131: 1–128.
- Chang F.H. & Northcote L. 2016. Species composition of extant coccolithophores including twenty six new records from the southwest Pacific near New Zealand. Marine Biodiversity Records 9: Article 75. DOI: https://doi.org/10.1186/s41200-016-0077-7.
- Claustre H. & Sciandra A. 2004. BIOSOPE cruise, RV L’Atalante. DOI:https://doi.org/10.17600/4010100.
- Conley S.M. 1979. Recent coccolithophores from the great barrier reef-coral sea region. Micropaleontology 25: 20–43. DOI: https://doi.org/10.2307/1485208.
- Cros L. 2002. Planktic coccolithophores of the NW Mediterranean. PhD thesis. Universitat de Barcelona, Barcelona. 181 pp.
- Cros L. & Fortuño J.-M. 2002. Atlas of northwestern Mediterranean coccolithophores. Scientia Marina 66 (Suppl. 1): 7–182. DOI: https://doi.org/10.3989/scimar.2002.66s11.
- Cros L., Kleijne A., Zeltner A., Billard C. & Young J.R. 2000. New examples of holococcolith-heterococcolith combination coccospheres and their implications for coccolithophorid biology. Marine Micropaleontology 39: 1–34. DOI: https://doi.org/10.1016/S0377-8398(0000010-4).
- Dimiza M. 2006. The study of recent coccolithophore biocommunities in the coastal environments of Andros island related to the late quaternary assemblages from central Aegean Sea. PhD thesis. National and Kapodistrian University of Athens, Athens. 338 pp.
- Edvardsen B., Eikrem W., Green J.C., Andersen R.A., Moon-Van der Staay S.Y. & Medlin L.K. 2000. Phylogenetic reconstructions of the Haptophyta inferred from 18S ribosomal DNA sequences and available morphological data. Phycologia 39: 19–35. DOI: https://doi.org/10.2216/i0031-8884-39-1-19.1.
- Font J. 1999. MTPII–MATER/HESP SEP99 cruise, RV Hesperides.
- Giraudeau J. & Bailey G.W. 1995. Spatial dynamics of coccolithophore communities during an upwelling event in the Southern Benguela system. Continental Shelf Research 15: 1825–1852. DOI:https://doi.org/10.1016/0278-4343(94)00095-5.
- Godrijan J., Young J.R., Marić Pfannkuchen D., Precali R. & Pfannkuchen M. 2018. Coastal zones as important habitats of coccolithophores: a study of species diversity, succession, and life-cycle phases. Limnology and Oceanography 63: 1692–1710. DOI: https://doi.org/10.1002/lno.10801.
- Gravalosa J.M., Flores J.-A., Sierro F.J. & Gersonde R. 2008. Sea surface distribution of coccolithophores in the eastern Pacific sector of the Southern Ocean (Bellingshausen and Amundsen Seas) during the late austral summer of 2001. Marine Micropaleontology 69: 16–25. DOI: https://doi.org/10.1016/j.marmicro.2007.11.006.
- Guerreiro C., Sá C., de Stigter H., Oliveira A., Cachão M., Cros L., Borges C., Quaresma L., Santos A.I. & Fortuño J.-M. 2014. Influence of the Nazaré Canyon, central Portuguese margin, on late winter coccolithophore assemblages. Deep Sea Research Part II: Topical Studies in Oceanography 104: 335–358. DOI: https://doi.org/10.1016/j.dsr2.2013.09.011.
- Haeckel E. 1894. Systematische Phylogenie der Protisten und Pflanzen. Erster Theil des Entwurfs einer systematischen Stammesgeschichte. Georg Reimer, Berlin, Germany. XV + 400 pp.
- Hagino K., Tomioka N., Young J.R., Takano Y., Onuma R. & Horiguchi T. 2016. Extracellular calcification of Braarudosphaera bigelowii deduced from electron microscopic observations of cell surface structure and elemental composition of pentaliths. Marine Micropaleontology 125: 85–94. DOI: https://doi.org/10.1016/j.marmicro.2016.04.002.
- Halldal P. & Markali J. 1955. Electron microscope studies on coccolithophorids from the Norwegian Sea, the Gulf Stream and the Mediterranean. Avhandlinger Utgitt Av Det Norske Videnskapsakademi I Oslo Mat- Naturvid Klasse 1: 1–30.
- Hay W.W. 1977. Calcareous nannofossils. In: Oceanic micropaleontology (Ed. by A.T.S. Ramsay), pp 1055–1200. Academic Press, London, UK.
- Hibberd D.J. 1972. Chrysophyta: definition and interpretation. British Phycological Journal 7: 281.
- Hibberd D.J. 1976. The ultrastructure and taxonomy of the Chrysophyceae and Prymnesiophyceae (Haptophyceae): a survey with some new observations on the ultrastructure of the Chrysophyceae. Botanical Journal of the Linnean Society 72: 55–80. DOI: https://doi.org/10.1111/j.1095-8339.1976.tb01352.x.
- Hutchins D.A. 2011. Forecasting the rain ratio. Nature 476: 41–42. DOI: https://doi.org/10.1038/476041a.
- Jordan R.W. & Green J.C. 1994. A check-list of the extant Haptophyta of the world. Journal of the Marine Biological Association of the United Kingdom 74: 149–174. DOI: https://doi.org/10.1017/S0025315400035736.
- Jordan R.W., Kleijne A., Heimdal B.R. & Green J.C. 1995. A glossary of the extant Haptophyta of the world. Journal of the Marine Biological Association of the United Kingdom 75: 769–814. DOI: https://doi.org/10.1017/S0025315400038169.
- Jordan R.W., Cros L. & Young J.R. 2004. A revised classification scheme for living Haptophytes. Micropaleontology 50: 55–79. DOI: https://doi.org/10.2113/50.Suppl_1.55.
- Kahn A.C.M. 2007. Morphometric variability in the extant coccolithophores: implications for the fossil record. PhD thesis. The State University of New Jersey, Rutgers. 250 pp.
- Kahn A. & Aubry M.-P. 2006. Intraspecific morphotypic variability in the family Rhabdosphaeraceae. Micropaleontology 52: 317–342. DOI: https://doi.org/10.2113/gsmicropal.52.4.317.
- Karatsolis B.T., Triantaphyllou M.V., Dimiza M.D., Malinverno E., Lagaria A., Mara P., Archontikis O. & Psarra S. 2017. Coccolithophore assemblage response to Black Sea water inflow into the north Aegean Sea (NE Mediterranean). Continental Shelf Research 149: 138–150. DOI: https://doi.org/10.1016/j.csr.2016.12.005.
- Kleijne A. 1992. Extant Rhabdosphaeraceae (coccolithophorids, class Prymnesiophyceae) from the Indian Ocean, Red Sea, Mediterranean Sea and North Atlantic Ocean. Scripta Geologica 100: 1–63.
- Kling S.A. 1975. A lagoonal coccolithophore flora from Belize (British Honduras). Micropaleontology 21: 1–13. DOI:https://doi.org/10.2307/1485152.
- Knoll M., Mueller T.J. & Siedler G. 1998. ESTOC/CANIGO cruises with FS Poseidon cruise 202/1, 212, 233, 237/3. Berichte aus dem Institut fuer Meereskunde an der Christian-Albrechts - Universitaet Kiel 302: 78.
- Koschinsky A., Brandt P. & Körtzinger A. 2009. Tropical Atlantic 2006 METEOR Cruise No. 68 April 26–August 7, 2006 Bridgetown–Recife–Mindelo–Las Palmas.
- Lecal J. 1966. A propos des modalités d’élaboration des formations épineuses des Coccolithophoridés. Protistologica 1: 63–70.
- Lecal J. 1967. Le nannoplancton des Côtes d’Israel. Hydrobiologia 29: 305–387. DOI: https://doi.org/10.1007/BF00189902.
- Malinverno E., Dimiza M.D., Triantaphyllou M., Dermitzakis M. & Corselli C. 2008. Coccolithophores of the Eastern Mediterranean Sea: a look into the marine microworld. Ion Publishing Group, Peristeri, Greece. 188 pp.
- Malinverno E., Triantaphyllou M.V. & Dimiza M.D. 2015. Coccolithophore assemblage distribution along a temperate to polar gradient in the West Pacific sector of the Southern Ocean (January 2005). Micropaleontology 61: 489–506.
- Monteiro F., Bach L.T., Brownlee C., Bown P.R., Rickaby R.E.M., Poulton A.J., Tyrrell T., Beaufort L., Dutkiewicz S., Gibbs S.J. et al. 2016. Why marine phytoplankton calcify. Science Advances 2016: 1–14. DOI: https://doi.org/10.1126/sciadv.1501822.
- Norris R.E. 1971. Extant calcareous nannoplankton from the Indian Ocean. In: Proceedings of the second planktonic conference, Roma (Ed. by A. Farinacci), pp 899–910. Edizioni Tecnoscienza, Rome, Italy.
- Norris R.E. 1984. Indian Ocean nanoplankton. I. Rhabdosphaeraceae (Prymnesiophyceae) with a review of extant taxa. Journal of Phycology 20: 27–41. DOI: https://doi.org/10.1111/j.0022-3646.1984.00027.x.
- Probert I., Fresnel J., Billard C., Geisen M. & Young J.R. 2007. Light and electron microscope observations of Algirosphaera robusta (Prymnesiophyceae). Journal of Phycology 43: 319–332. DOI: https://doi.org/10.1111/j.1529-8817.2007.00324.x.
- Reid F.M.H. 1980. Coccolithophorids of the North Pacific Central Gyre with notes on their vertical and seasonal distribution. Micropaleontology 26: 151–176. DOI: https://doi.org/10.2307/1485436.
- Robinson C., Poulton A.J., Holligan P.M., Baker A.R., Forster G., Gist N., Jickells T.D., Malin G., Upstill-Goddard R., Williams R.G. et al. 2006. The Atlantic Meridional transect (AMT) programme: a contextual view 1995–2005. Deep-Sea Research Part 2, Topical Studies in Oceanography 53: 1485–1515. DOI: https://doi.org/10.1016/j.dsr2.2006.05.015.
- Schneider C.A., Rasband W.S. & Eliceiri K.W. 2012. NIH Image to ImageJ: 25 years of image analysis. Nature Methods 9: 671–675. DOI: https://doi.org/10.1038/nmeth.2089.
- Sigman D.M. & Boyle E.A. 2000. Glacial/interglacial variations in atmospheric carbon dioxide. Nature 407: 859–869. DOI: https://doi.org/10.1038/35038000.
- van de Locht R., Slater T.J.A., Verch A., Young J.R., Haigh S.J. & Kröger R. 2014. Ultrastructure and crystallography of nanoscale calcite building blocks in Rhabdosphaera clavigera coccolith spines. Crystal Growth & Design 14: 1710–1718. DOI: https://doi.org/10.1021/cg4018486.
- Varol O. 1989. Eocene calcareous nannofossils from Sile (northwest Turkey). Revista Española de Micropaleontología 21: 273–320.
- Wang J., Luan Q., Zuo T, Chen R., & Sun J. 2012. Taxonomic composition of marine-living coccolithophores in the Yellow Sea and East China Sea—new records and a species list. Marine Biodiversity Records 5: 1–8. DOI: https://doi.org/10.1017/S1755267212001005.
- Wallich G.C. 1877. Observations on the coccosphere. Annals and Magazine of Natural History 19: 342–350. DOI: https://doi.org/10.1080/00222937708682153.
- Winter A., Reiss Z. & Luz B. 1979. Distribution of living coccolithophore assemblages in the Gulf of Elat (‘Aqaba). Marine Micropaleontology 4: 197–223. DOI: https://doi.org/10.1016/0377-8398(79)90017-3.
- Woodward M. 2009. AMT-18 cruise report. Plymouth Marine Laboratory. https://www.bodc.ac.uk/resources/inventories/cruise_inventory/reports/jr218.pdf
- Yang T., Wei K. & Gwo-Ching G. 2001. Distribution of coccolithophorids and coccoliths in surface ocean off northeastern Taiwan. Botanical Bulletin of the Academia Sinica 42: 287–302.
- Yang T.-N., Wei K.-Y. & Chen L.-L. 2003. Occurrence of coccolithophorids in the northeastern and central South China Sea. Taiwania 48: 29–45. DOI: https://doi.org/10.6165/tai.2003.48(1).29.
- Young J.R. & Bown P.R. 1997. Higher classification of calcareous nannofossils. Journal of Nannoplankton Research 19: 15–20.
- Young J.R., Didymus J.M., Bown P.R., Prins B. & Mann S. 1992. Crystal assembly and phylogenetic evolution in heterococcoliths. Nature 356: 516–518. DOI: https://doi.org/10.1038/356516a0.
- Young J.R., Bergen J.A., Bown P.R., Burnett J.A., Fiorentino A., Jordan R.W., Kleijne A., van Niel B.E., Romein A.J.T. & von Salis K. 1997. Guidelines for coccolith and calcareous nannofossil terminology. Palaeontology 40: 875–912.
- Young J.R., Geisen M., Cros L., Kleijne A., Probert I. & Ostergaard J.B. 2003. A guide to extant coccolithophore taxonomy. Journal of Nannoplankton Research, Special Issue 1: 1–132.
- Young J.R., Hagino K. & Poulton A. 2014. Gladiolithus adeyi sp. nov. a new deep photic coccolithophore species and new molecular genetic and crystallographic observations on. Gladiolithus flabellatus. Journal of Nannoplankton Research 34: 61–68.
- Young J.R., Bown P.R. & Lees J.A. 2021. Nannotax3. http://www.mikrotax.org/Nannotax3;http://ina.tmsoc.org/Nannotax3. [ accessed 24 January 2021].