
ABSTRACT
Trilophosauridae are an extinct group of archosauromorphs from the Upper Triassic of western North America and Nova Scotia. Possible trilophosaurids Tricuspisaurus thomasi and Variodens inopinatus have also been reported from Upper Triassic fissure fills in the southwest of the United Kingdom, based on their multi-cuspid dentitions. Here, we provide an updated description of V. inopinatus and add to the description of T. thomasi, including re-examining CT scans of its holotype dentary. We confirm that the apparent gap on the dentary of T. thomasi is pathological, with the dentine of the teeth on either side of the gap asymmetrically thick, and that heavy vascularization of the anterior part of the dentary confirms it likely had a keratinous rhamphotheca. Our cladistic analyses, to which we add nine new characters, confirm that T. thomasi and V. inopinatus are closely related taxa and the youngest known trilophosaurids. Further, we support a previous suggestion that the relatively small size of these two European trilophosaurids results from insular dwarfism.
INTRODUCTION
Trilophosauridae are an extinct group of lizard-like, herbivorous reptiles included in the archosauromorph clade Allokotosauria, all known for their diverse dental morphologies (Nesbitt et al., Citation2015; Kligman et al., Citation2020). Trilophosaurids, characterized by their transversely broadened, bicuspid/tricuspid dentition, are known from the Upper Triassic of western North America and Nova Scotia (Gregory, Citation1945; Mueller and Parker, Citation2006; Spielmann et al., Citation2008; Kligman et al., Citation2020). Trilophosaurus was suggested as Carnian–Norian in age (e.g., Heckert et al., Citation2006), but radiometric and biochronological age estimations for the Chinle Formation and Dockum Group have been revised upwards, so that the genus Trilophosaurus is not recorded before the early Norian (Kligman et al., Citation2020). There are six currently accepted species of trilophosaurids, all from North America: Trilophosaurus buettneri, T. jacobsi, T. dornorum, T. phasmalophos, Teraterpeton hrynewichorum, and Spinosuchus caseanus (Murry, Citation1987; Heckert et al., Citation2006; Mueller and Parker, Citation2006; Kligman et al., Citation2020). Spielmann et al. (Citation2007, Citation2008, Citation2009) regard T. dornorum as conspecific with T. jacobsi, citing similarities of the teeth of the former to large teeth of the latter, but we follow the most recent analysis of Kligman et al. (Citation2020) who consider the two as separate species.
In Europe, two possible trilophosaurids, Tricuspisaurus thomasi and Variodens inopinatus, have been reported from Rhaetian, Upper Triassic fissure infill deposits of the southwestern United Kingdom (Robinson, Citation1957a; Skinner et al., Citation2020:pp. 549–551), based on their transversely broadened and multi-cuspid dentition and, as in T. thomasi, likely possession of a keratinous rhamphotheca (or beak). This makes them the youngest trilophosaurids, but their phylogenetic identity has been disputed (Robinson, Citation1957a; Fraser, Citation1986; Sues and Olsen, Citation1993; Skinner et al., Citation2020). The fissure fill rock containing Variodens inopinatus Robinson, Citation1957a was collected from Emborough Quarry in southwest England and the two specimens were prepared at University College London (UCL; Robinson, Citation1957a). The specimens that were later named T. thomasi were collected in 1947 or 1948 and recognized as probably trilophosaurid by Walter G. Kühne (1911–1991) in a note of sale to the NHMUK (Skinner et al., Citation2020). T. thomasi was first described by Robinson (Citation1957a) from jaw fragments and teeth from the Ruthin quarries, east of Bridgend, south Wales (Skinner et al., Citation2020), the material of which has been stored at the NHMUK ever since. The holotype of T. thomasi, and some additional material from the NHMUK and NMW, was then redescribed by Skinner et al. (Citation2020), whose CT scans we use here.
The Ruthin Tricuspisaurus thomasi and Emborough Variodens inopinatus were located on two different paleoislands in the Bristol archipelago (U.K. grid references SS 975795 and ST 623505 respectively; Fig. S1), a series of islands formed of uplifted Carboniferous limestone in shallow, tropical seas that existed from the Rhaetian to Sinemurian. The fissure infill deposits contain the fossils of many animals besides these two trilophosaurids, including procolophonids, rhynchocephalians, kuehneosaurids, crocodylomorphs, dinosauromorphs, and cynodonts (Robinson, Citation1971; Evans and Kermack, Citation1994; Whiteside et al., Citation2016; Skinner et al., Citation2020). These vertebrate faunas document the transition through the Late Triassic mass extinction events, during which several key clades, including trilophosaurids and procolophonids became extinct.
In this paper we examine the external and internal morphology of the holotype specimen of Tricuspisaurus thomasi using CT scanning and re-evaluate the holotype and paratype specimens of Variodens inopinatus. For the first time we run a cladistic analysis for both European trilophosaurids to evaluate their relationships among other archosauromorphs.
Institutional Abbreviations––NHMUK, Natural History Museum, London, United Kingdom; NMMNH, New Mexico Museum of Natural History and Science, Albuquerque, New Mexico, U.S.A.; NMW, National Museum of Wales, Cardiff, U.K.; TMM, Vertebrate Paleontology Collections, the University of Texas at Austin, Austin, Texas, U.S.A.; UMMP, University of Michigan Museum of Paleontology, Ann Arbor, Michigan, U.S.A.
MATERIALS AND METHODS
The key specimens of Tricuspisaurus thomasi, collected by Kühne, are shown in . The holotype left dentary of T. thomasi (NHMUK PV R6106, , ) was scanned by Skinner et al. (Citation2020), using a Nikon XTH 225ST X-ray computed tomography (CT) scanner at the University of Bristol, and we use these scans here. We digitally segmented the scans from scratch as the original surface files were not available, using Avizo Lite 9.0 (Visualization Sciences Group) to examine the internal and external morphology of the dentary. The two specimens of Variodens inopinatus described by Robinson (Citation1957a) are shown in (NHMUK PV R36850 and NHMUK PV R36849).
FIGURE 1 . Specimens of Tricuspisaurus thomasi from Ruthin Quarry. The holotype left dentary of Tricuspisaurus thomasi (NHMUK PV R6106) in A, lateral; B, occlusal; C, surface model of the occlusal views; and D, close up of the anterior occlusal region. E, NHMUK PV R6108, a probable maxilla in lateral view. A fragment of a probable right? maxilla (NHMUK PV R6107) in F, occlusal; G, lateral; and H, magnified occlusal (and slightly anterior) views. New character 230-1, bone of attachment above dorsal dentary margin, shown in A. Specimens digitally removed from matrix in A, B and D.
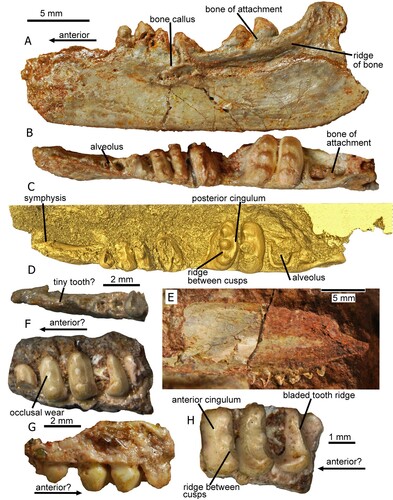
FIGURE 2. The holotype left dentary of Tricuspisaurus thomasi (NHMUK PV R6106); A, C, E, G, H, I, segmented in lateral, medial, occlusal, anterior, posterior, and ventral views, respectively; B, D, F, line diagrams in lateral, lingual, and occlusal views, respectively; J, sagittal cross-section of the anterior region of the dentary. K, coronal cross-section of the anterior region of the dentary. L, coronal cross-section of the penultimate tooth. M, sagittal cross-section of the mid-region of the dentary. New character 229-1, pits, foramina and rugosity suggestive of a beak, shown in A, B, and J. Abbreviations: bk, beak; br, broken; cap, pulp cavity; cpr, coronoid process; dent, dentine; fac, facet; por, porosity; ppr, posterior process; reg, region; rid, ridge; soc, socket; spl, splenial; sur, surangular; td, thickened bone; th, tooth.
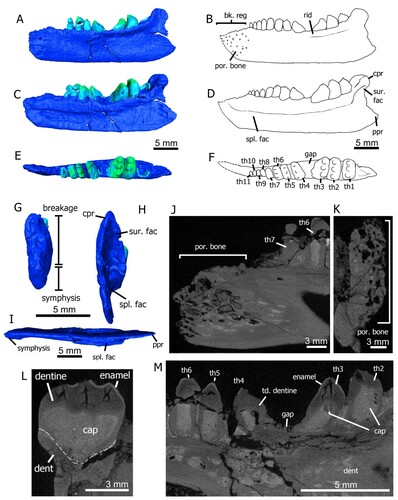
FIGURE 3. Specimens of Variodens inopinatus. The holotype left dentary of Variodens inopinatus (NHMUK PV R36850), in A, lateral (and slightly occlusal); B, occlusal (and slightly lateral); and C, medial views. The numbers above the teeth in A and G are a guide to the tooth positions (using Robinson’s [Citation1957a] nomenclature), mentioned in the text. The paratype specimen (NHMUK PV R36849), an anterior fragment of a left dentary, in D, medial; E, lateral; and F, occlusal views. Reconstruction of the dentary by P.L. Robinson (Citation1957a) in G, occlusal; and H, lateral views. New character 229-0, pits, foramina but no suggestion of a beak, shown in E. New character 230-1, bone of attachment above dorsal dentary margin, shown in A, C and D.
![FIGURE 3. Specimens of Variodens inopinatus. The holotype left dentary of Variodens inopinatus (NHMUK PV R36850), in A, lateral (and slightly occlusal); B, occlusal (and slightly lateral); and C, medial views. The numbers above the teeth in A and G are a guide to the tooth positions (using Robinson’s [Citation1957a] nomenclature), mentioned in the text. The paratype specimen (NHMUK PV R36849), an anterior fragment of a left dentary, in D, medial; E, lateral; and F, occlusal views. Reconstruction of the dentary by P.L. Robinson (Citation1957a) in G, occlusal; and H, lateral views. New character 229-0, pits, foramina but no suggestion of a beak, shown in E. New character 230-1, bone of attachment above dorsal dentary margin, shown in A, C and D.](/cms/asset/5ad2f0de-189c-41c6-ae50-c98f73c0a5a7/ujvp_a_1999250_f0003_oc.jpg)
Our phylogenetic analysis is based on the recent archosauromorph dataset containing Allokotosauria matrix last modified by Kligman et al. (Citation2020), and prior to that by Nesbitt et al. (Citation2015), from Pritchard et al. (Citation2015). We further modified this matrix by adding nine characters, and the two operational taxonomic units (OTUs) Tricuspisaurus thomasi and Variodens inopinatus. We also identified 25 parsimony-uninformative characters that we also removed. The new matrix (see Supplemental Material) consists of 33 OTUs and 231 characters and was rooted using the Late Carboniferous diapsid Petrolacosaurus kansensis. We ran a parsimony analysis in PAUP* 4.0a (Swofford, Citation2002) using a heuristic search and tree-bisection-reconnection (TBR) branch swapping, in addition to calculating Bremer values, and consistency, retention, and rescaled consistency indices (CI, RI, RC, respectively). Ten of the characters were ordered (Chs. 1, 4, 9, 10, 19, 31, 47, 62, 181, 189) following the steps of previous authors (e.g., Nesbitt et al., Citation2015; Kligman et al., Citation2020). All recovered MPTs were then summarized in a strict consensus tree. We also sought out rogue taxa using the iterPCR protocol in TNT V.1.5 (Goloboff and Catalano, Citation2016), but the analysis did not identify any. In addition, we ran a Bayesian analysis, using MrBayes v. 3.2 (Huelsenbeck and Ronquist, Citation2001; Ronquist et al., Citation2012) for 107 generations, sampling parameters every 1000 generations, and the first 25% of sampled trees were discarded as burn-in.
SYSTEMATIC PALEONTOLOGY
ARCHOSAUROMORPHA Huene, Citation1946
TRILOPHOSAURIDAE Gregory, Citation1945
TRICUSPISAURUS Robinson, Citation1957a
TRICUSPISAURUS THOMASI Robinson, Citation1957a
Holotype––A near complete dentary, NHMUK PV R6106.
Referred Specimens––A left maxilla, NHMUK P.L.R. 500; a probable right maxilla, NHMUK PV R6107; a probable maxilla, NHMUK PV R6108; an isolated tricuspid tooth, NMW 88.12 G.73; left dentary fragment with teeth, NMW 88.12 G.36.
Locality and Horizon––Upper Triassic, Rhaetian locality of Ruthin, South Wales, United Kingdom (Robinson, Citation1957a; Skinner et al., Citation2020). All definite material for Tricuspisaurus thomasi is known from the Ruthin quarries (Robinson, Citation1957a; Skinner et al., Citation2020:fig. 1), located east of Bridgend, South Wales. In total there are four quarries at Ruthin, and the fossils came from the small northern quarry (see fig. 1 in Skinner et al., Citation2020). The high abundance of reptilian remains in this deposit was first reported by Thomas (Citation1952), but much of the material was collected earlier by Kühne including the holotype T. thomasi dentary examined in this paper. This infill accumulated in a network of fissures within the uplifted Carboniferous limestone islands. The infill is dominated by red marl and calcitic sandstone rich in vertebrate fossils, but also contains a mixture of non-fossiliferous yellow-gray limestone and brown shale (Thomas, Citation1952; Skinner et al., Citation2020). The quarries at Ruthin are believed to contain the oldest fissure infill in the Triassic–Jurassic Bristol Archipelago (Skinner et al., Citation2020), and the terrestrial fauna is dominated by procolophonids, but also includes archosauromorphs and early diverging rhynchocephalians such as Diphydontosaurus sp. and Planocephalosaurus sp.
DESCRIPTION
The holotype, NHMUK PV R6106 (), is a near-complete left dentary with some damage to its anterodorsal tip, though part of the symphysis is preserved. Several of the teeth are missing, the sockets of which are partially obscured anteriorly beneath lacquer. The rest of the teeth have varying levels of damage but are, for the most part, complete. Although Skinner et al. (Citation2020) suggested there were nine teeth and alveoli, there is evidence for 10 or 11 recognizable tooth implantations, but there could have been more in the anteriormost area () that is generally regarded as edentulous (e.g., Robinson, Citation1957a). Skinner et al. (Citation2020) reported about five cusps and possible alveoli in the anterior region, but this area is substantially obscured by the lacquer and not visible in the current resolution of the CT scan. The teeth sit in shallow sockets and are protothecodont (Skinner et al., Citation2020; = ankylothecodont of Heckert et al., Citation2006) (). Protothecodonty describes shallow-rooted, ankylosed, socketed teeth and ankylothecodonty a more deep-rooted dentition (see discussions in Small, Citation1997; Cabreira and Cisneros, Citation2009); the teeth of T. thomasi are shallow rooted and of the protothecodont type. The teeth are ankylosed to the dentary alveoli by outer fibrous bone of attachment (Robinson, Citation1957a), which is prominent on the larger teeth and the distalmost alveolus (). The most distinctive feature of the teeth is the three cusps that are positioned labiolingually, with the distances between the labial to mid cusp and mid to lingual cusp approximately equidistant (). The labial and lingual cusps have slightly larger bases than the mid cusp and they tend to be higher, but this is not always the case. The tricuspid teeth are remarkable as they have well developed distal and mesial cingula on the crown (see Fig. S2). It is clear from the surface model () that a sharp transverse ridge runs between the labial and mid cusp and another between the mid and lingual cusp. The mid and distal teeth sit on a laterally and medially expanded dorsal region of the dentary (). There is a gap in the tooth row that we consider to be a pathological abnormality that we discuss later.
The ventral part of the dentary is mediolaterally narrow and dorsoventrally tall and bears a huge facet for the splenial (), similar to the condition in Trilophosaurus buettneri (Spielmann et al., Citation2008:fig. 28C), indicating the jaw would have been more robust in life. Other bone facets are less defined but one for the surangular appears to be visible on the dorsal postero-lateral surface of the coronoid process (). The anterior part of the dentary, either edentulous or bearing very tiny teeth, has porous, highly vascularized bone visible both externally on the lateral surface of the dentary () and internally in the CT scan (), evidence further supporting the former presence of a trilophosaurid-like beak (Gregory, Citation1945; Robinson, Citation1957a; Spielmann et al., Citation2008; Skinner et al., Citation2020:549–551). As here, the presence of a keratinous rhamphotheca is inferred in other Triassic groups (e.g., aetosaurs, dicynodonts, ornithischians, shuvosaurids, silesaurids) from an edentulous anterior portion of the dentary and numerous neurovascular foramina (Wynd et al., Citation2020), all of which are present in T. thomasi. The coronoid process has been described as human ‘ear-shaped’ by Robinson (Citation1957a) (, ) and is more prominent, with a greater dorsal extent, than in Trilophosaurus buettneri (see e.g., Gregory, Citation1945:fig. 4).
The symphyseal region of Tricuspisaurus thomasi is mediolaterally narrower than in Trilophosaurus buettneri (Robinson, Citation1957a) but is also formed by an interdigitation of the left and right bones (see the dentary of Trilophosaurus buettneri, Fig. S3A; Spielmann et al., Citation2008:fig. 27e), which we infer in Tricuspisaurus thomasi (Fig. S2J; Skinner et al., Citation2020:fig. 10N) and Trilophosaurus jacobsi (Fig. S3B) and is likely present in Variodens inopinatus (see the medial view of the symphyseal region, ).
Tricuspisaurus thomasi and Trilophosaurus species including T. buettneri and T. jacobsi are distinguished further by the large distal dentary teeth lying in variable positions from 3rd–6th most distal in the Trilophosaurus dentition (see Spielmann et al., Citation2008:figs. 27a, 93k, l: Gregory, Citation1945:fig. 6) but in the penultimate distal position in Tricuspisaurus ().
Two other Tricuspisaurus thomasi specimens collected by Kühne are NHMUK PV R6107 and NHMUK PV R6108 which we identify tentatively as posterior portions of a maxilla. The slightly convex shape of the base of the dentition of NHMUK PV R6108 () indicates that this specimen is a very poorly preserved fragment of maxilla. Skinner et al. (Citation2020) regarded NHMUK PV R6107 () as a possible partial maxilla. The teeth on the specimen have a slightly elongated ‘neck’ near the base of the crown that resemble the distal maxillary teeth of Trilophosaurus buettneri in Spielmann et al. (Citation2008:fig. 20A). The penultimate tooth is a curious L-shape blade with a sharp ridge running transversely across the apex of the cusp (). One interpretation (suggested independently by Edwards [Citation2001] and Skinner et al. [2020]) is that the tooth is fractured, possibly has an exposed pulp cavity, is worn and would have been tricuspid in life, whereas Robinson (Citation1957a) argued that the tooth is complete. Robinson cited similar isolated teeth in the NMW as support for her suggestion and it is clear from that there is no indication of a possible mid cusp, so we cannot resolve this debate. The close-up of the two largest teeth in also indicates a second slight vertical ridge placed to one side of the central cusp. There are also smaller teeth distally on the right and left maxilla of Trilophosaurus buettneri TMM 31025-140 (Spielmann et al., Citation2008:figs. 18d, 19d) and anteriorly on the maxillae of specimens TMM 31025-207 and TMM 31025-143 (Spielmann et al., Citation2008:figs. 20c, 21d). Of these examples the mesial tooth of the right maxilla of TMM 31025-207 bears most similarity to the bladed tooth of Tricuspisaurus thomasi NHMUK PV R6108.
Robinson (Citation1957a) referred to a specimen ‘PLR 500’ from her own collection as a maxilla, but this could not be located in the NHMUK. The specimen is shown in Robinson (Citation1957a:fig. 3) and Skinner et al. (Citation2020:fig. 10a) but its appearance is akin to the holotype dentary NHMUK PV R6106 and bears little similarity to the Trilophosaurus buettneri maxilla depicted by Spielmann et al. (Citation2008:fig. 20a, b). The bone below the tooth row of ‘PLR 500’ appears to have the ridge developed in the holotype dentary () and the base of the dentition follows the dentigerous surface of the bone in the concave manner of NHMUK PV R6106, rather than the straight or slightly convex line of the maxilla of T. buettneri. Unless ‘PLR 500’ is rediscovered it is not possible to confirm its identification.
Pathology of Tricuspisaurus thomasi
On the dentary of NHMUK PV R6106, there appears to be a small gap between the 3rd and 4th tooth (), and similar gaps in Trilophosaurus buettneri have been explained as an artifact of preparation or as tooth replacement (Gregory, Citation1945; Demar and Bolt, Citation1981; Spielmann et al., Citation2008). There has been some debate whether this triangular gap in Tricuspisaurus thomasi is an autapomorphy or the result of injury (Robinson, Citation1957a; Skinner et al., Citation2020). Our scans provide further evidence to suggest that the latter is the case. In sagittal cross-section () the dentine of the teeth on either side of the gap is asymmetrically thickened, and the pulp cavity of these teeth is distorted in shape. The teeth either side of the gap also appear to be growing at an angle rather than vertically, which differs from the rest of the teeth in the tooth row.
There is a bony callus () on the upper part of the lateral side of the dentary just below and slightly anterior to the gap. This probably represents a healed fracture, and it was this damage, experienced some time before death, that might have generated the edentulous gap and the deformed teeth. Alternatively, the callus might have been caused by an abscess representing infection, and the bone and dentition has become re-modeled. Overall, the evidence supports the view that the gap is likely pathological and is the reason for the empty tooth alveolus.
We have considered the dentary carefully in our phylogenetic analysis and have left out the triangular gap as a character but assess the main features of the mesial and distal teeth (implantation, relative width, cusps, cingula, and arêtes) as sufficiently characteristic to be scored.
VARIODENS INOPINATUS Robinson, Citation1957a
Holotype––Mostly complete left dentary, NHMUK PV R36850.
Referred Specimens––Paratype, NHMUK PV R36849.
Locality and Horizon––Upper Triassic, Rhaetian fissure locality in Emborough Quarry, southwest England, United Kingdom (Robinson, Citation1957a, Citationb; Whiteside et al., Citation2016). The type specimens are from a fissure (UK grid reference ST 623505; see Fig. S1) figured by Robinson (Citation1957b:pls. 2–4) and described by Savage (Citation1977:89–90). In addition, there is also a fragment of a probable Variodens sp. from a Rhaetian fissure in Holwell ‘Microlestes’ Quarry, collected by Charles Moore in the 1850s and described by Whiteside and Duffin (Citation2017).
The Emborough Quarry Triassic fissure fill is best known as the type locality for the gliding lepidosauromorph Kuehneosaurus latus (Robinson, Citation1962), found in huge numbers of mostly disarticulated but associated bones in a red conglomeratic matrix, the main fossiliferous lithology in the fissure. We were unable to determine who discovered the Variodens inopinatus specimens. However, reptile-bearing Emborough rocks were first collected by Kühne in 1946 (then of UCL; Robinson, Citation1957b) but Pamela L. Robinson of UCL and Tom Fry, a collector employed by the University of Bristol, also made substantial collections from the site. In addition to K. latus and V. inopinatus, the fauna collected includes small archosauromorphs and lepidosaurs.
The dating of the Emborough Quarry fissure deposit is debated. Robinson (Citation1957b) considered it to be older than the Rhaetian transgression and, therefore, Norian, whereas Whiteside et al. (Citation2016) argued that the entrance to the fissure was not inundated by the sea until after the deposition of the base of the Westbury Formation in the early Rhaetian. This early Rhaetian age has been confirmed by detailed mapping studies reported by Lovegrove et al. (Citation2021).
DESCRIPTION
The holotype specimen, NHMUK PV R36850 (), is a mostly complete left dentary, nearly 20 mm long, but lacking the most anterior region and the posterior part of the coronoid process. Although broken, the coronoid process appears to be more similar to that in the reconstruction of Trilophosaurus buettneri (Spielman et al., Citation2008:fig. 22b, c) than to the much higher feature in Tricuspisaurus thomasi. A splenial is probably present on the medial side () but the bone is too fractured to discern its contacts with the dentary. The paratype specimen NHMUK PV R36849 () is a much smaller fragment of the anterior region of a left dentary, including a part of the symphysis, but the combination of the two specimens enabled Robinson (Citation1957a) to provide a thorough description and reconstruction () of the dentary. Her account is paraphrased here together with new information. Robinson (Citation1957a:284) combined the evidence from the two dentaries to demonstrate that there would have been 18–19 teeth on a complete dentary with a length of about 22 mm. From the mesialmost position, the smaller dentary has tooth positions 1–9 and the larger dentary alveoli from position 5–18 (using Robinson’s Citation1957a nomenclature). The clearest comparative positions are 8–9 in the paratype and 3–4 in the holotype; in both cases the teeth are readily recognized as tricuspid (compare ). Considering a composite dentary composed of the two specimens, the holotype would have tooth positions 5–18.
The teeth are protothecodont and sit in shallow alveoli (); this includes the mesial conical teeth of NHMUK PV R36849 deemed pleurodont by Robinson (Citation1957a). There is prominent fibrous or spongy (Robinson, Citation1957a) bone of attachment () that ankyloses the base of each tooth crown to the rim of the dental alveoli; this bone of attachment extends up the ‘neck’ region reaching the base of the upper crown. There is a probable resorption pit (), recognized by Robinson (Citation1957a) as evidence that the teeth were actively replaced. The 14 teeth of NHMUK PV R36850 are all multi-cusped except the most mesial and distal. However even in the most mesial tooth there is evidence of raised areas positioned labially and lingually of the conspicuous central cusp; the tooth lying immediately distal has a rudimentary development of two minor cusps flanking the central apex (tooth 2, ). Distal to these two teeth, the characteristic transversely wide tricuspid morphology is present in all but the distalmost erupting tooth, which has a simple small conical shape with its apex directed mesially. The most mesial tooth position of NHMUK PV R36849 consists of a remnant base and partial alveolus but the following four mesial teeth are all simple conical forms. Distally to these teeth, the next two have rudimentary very small, raised cusps which Robinson (Citation1957a:table, p. 284) regarded as ‘transitional to the 3-cusped type,’ and the final two teeth (numbers 8 and 9) are tricuspid. The three cusps on Variodens inopinatus dentary teeth are all similar in height except on the largest teeth 12 and 13 of NHMUK PV R36850 (, S2D) where the central cusp is noticeably highest. Although mesial and distal cingula are present on teeth 7–13, they are less defined than those of Tricuspisaurus thomasi (Robinson Citation1957a; cf. Fig. S2C, F).
In occlusal view, teeth 6–12 are cassinoid-shaped (Fig. S2B), meaning possessing two rounded ends and a central constriction; this is a unique feature of Variodens inopinatus among the trilophosaurids. In contrast, the large tooth 13 is sub-circular in occlusal view. Teeth 12 and 13 have the most complex crown surface morphology, with accessory cusps and additional cingula (Robinson, Citation1957a; Fig. S2D). The most prominent accessory cusps are on the distal sides of the lingual and labial cusps of tooth 12 and behind the labial cusp of tooth 13 (, S2D). However, both teeth also have raised areas or ‘shoulders’ on the distal side of the labial cusp. In lingual view, a ‘shoulder’ is present on the distal side of the apex of the lingual cusp on teeth 12 and 13 (). As a result of the cusps and accessory cusps, basins (Fig. S2D) are developed between the labial and central cusp and the lingual and central cusp on both these large teeth. In addition to mesial and distal cingula more cingula are present between cusps particularly on tooth 13 (Fig. S2D).
The lateral sides of both dentaries have a profusion of large foramina that are especially numerous at the anterior end of NHMUK PV R36849 (), indicating that this was a highly vascularized region. The number of foramina is much greater than observed in Tricuspisaurus thomasi but unlike that taxon there is no evidence of a rugose anterior region that could indicate the presence of a keratinous rhamphotheca. In common with T. thomasi, there is a prominent, laterally projecting, longitudinal ridge of bone on the lateral side of the large dentary below the tooth row ().
Remarks
There are many features, particularly in the dentition, common to both Tricuspisaurus thomasi and Variodens inopinatus. Most notably the transversely wide tricuspid nature of the larger teeth and type of implantation, protothecodont, is similar with the teeth ankylosed in shallow alveoli by fibrous bone of attachment (compare with ) which originates from the rim of the alveolus. This bone of attachment reaches the base of the expanded upper crown region in both taxa. There are, however, significant differences, with T. thomasi teeth displaying more prominent cingula and a more typical trilophosaurid sub-ovoid shape in occlusal view (cf., Heckert et al., Citation2006:figs. 3A, 7D, F; Spielmann et al., Citation2008:figs. 8, 9) with relatively straight mesial and distal sides. This shape is depicted best in the mid tooth region (teeth 4 and 5) of the T. thomasi holotype in front of the gap in and two of the teeth in ; also see Skinner et al. (Citation2020:fig. 10G, J, M). By contrast, the cassinoid outline of the Variodens inopinatus tricuspid teeth (, S2B) differs from all other trilophosaurids. Further, and unlike any other trilophosaurid, V. inopinatus has many accessory cusps on the largest distal teeth and the penultimate tooth is a unique sub-circular shape in occlusal view (Fig. S2B, D).
The similar size and relatively equal spacing of the three cusps on T. thomasi tricuspid teeth are reminiscent of Trilophosaurus buettneri teeth (e.g., Heckert et al., Citation2006:fig. 7B, D) rather than the larger central cusp and slight asymmetrical placement of cusps in Trilophosaurus jacobsi (e.g., Heckert et al., Citation2006:fig. 7A, C). The pronounced cingula of Tricuspisaurus thomasi teeth are more similar to those of Trilophosaurus jacobsi but also resemble the teeth of juvenile T. buettneri (see Demar and Bolt Citation1981:fig. 6). V. inopinatus has similarly sized cusps in most tricuspid teeth but, of the three major cusps, the central one of each of the two largest distal teeth is noticeably highest. V. inopinatus (Fig. S2D) therefore differs from T. thomasi and Trilophosaurus species in the unique cassinoid shape (in occlusal view) of most of its tricuspid teeth, but also in the circular shape of the penultimate tooth and the greater complexity of the crown surface in the largest distal teeth (see Fig. S2D).
The occlusal wear on the cusps of the multicuspid teeth is similar in T. thomasi and V. inopinatus, forming blunted apices (, S2C) that suggests an orthal motion of the jaws with the upper and lower cusps meeting. Cusp wear of this type is common in Trilophosaurus buettneri (Demar and Bolt, Citation1981). However, there is additionally mesial wear on the central cusp of tooth 12 of V. inopinatus (Fig. S2D) indicating that the upper and lower cusps are slightly offset in that region; Robinson (Citation1957b:284) noted this wear facet and two further concave wear facets on the same tooth. Demar and Bolt (Citation1981:fig. 5) reported six regions of mesial and distal wear facets in some T. buettneri teeth. The V. inopinatus dentary is dorsoventrally more gracile compared with T. thomasi and Trilophosaurus species. Further, the anterior region on the dentary of V. inopinatus is toothed, rather than edentulous, and lacks any evidence of a beak. Variodens inopinatus is the smallest trilophosaurid known from a toothed bone with an estimated length, based on T. buettneri of Spielmann et al. (Citation2008) of a body length of about 0.45 m.
Tricuspisaurus thomasi is therefore a more ‘typical’ trilophosaurid, albeit with a noticeably greater prominence of the mesial and distal cingula on the dentary teeth compared with adult Trilophosaurus buttneri (e.g., the holotype UMMP 2338 in Spielman et al., Citation2008:fig. 9i) but with a similar eminence to those of Trilophosaurus jacobsi and skeletally immature T. buettneri. The similarity to Trilophosaurus was recognized by Kühne in his sale of the specimens to the NHMUK in 1948 (Skinner et al., Citation2020). A major problem with the description of T. thomasi is that it is based on a holotype dentary with a pathological abnormality. However other specimens, particularly with the characteristic trilophosaurid teeth clearly demonstrate its affinities.
Cladistic Analysis
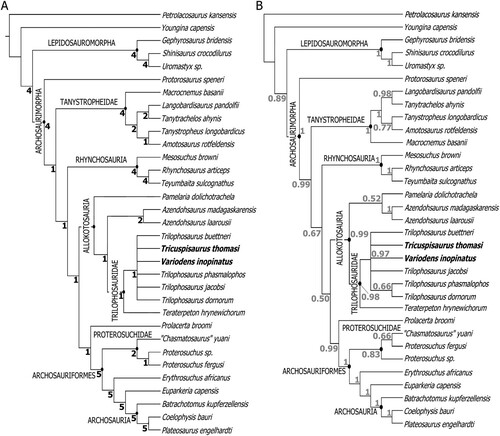
The maximum parsimony analysis () retained 68 trees of length 616 steps (consistency index, CI = 0.409; retention index, RI = 0.644; rescaled consistency index, RC = 0.263), in total there were 616,458 rearrangements tried. The Bayesian analysis result () closely resembles the parsimony analysis but with lower resolution within Trilophosauridae. Both analyses retrieve Allokotosauria as monophyletic, with Pamelaria dolichotrachela and the azendohsaurids being the earliest diverging groups. In the maximum parsimony analysis Pamelaria dolichotrachela is the earliest diverging member of Allokotosauria, whereas in the Bayesian analysis P. dolichotachela is the sister group to the genus Azendohsaurus (see also Ezcurra, Citation2016). The earliest diverging member of Trilophosauridae is Teraterpeton hrynewichorum, which agrees with previous analyses (Nesbitt et al., Citation2015; Kligman et al., Citation2020). The relationships within Trilophosaurus are poorly supported with many collapsing to form a polytomy, as in previous analyses (Kligman et al., Citation2020), but the majority consensus tree for the Bayesian analysis suggests that T. dornorum and T. jacobsi are sister taxa, with the same relationship suggested by the 50% majority-consensus tree maximum parsimony analysis (Fig. S4). The 50% majority-consensus maximum parsimony tree also tentatively suggests a group comprising T. phasmalophus, Spinosuchus caseanus, T. dornorum, and T. jacobsi.
FIGURE 4. Phylogeny of Neodiapsida, demonstrating the relationships within Trilophosauridae, and with other related clades. A, strict consensus tree obtained from maximum parsimony analysis. B, 50% majority rule consensus tree obtained from Bayesian inference analysis. Bremer values are in black, clade credibility values as decimals are in gray. Bremer values that fall below 1 collapse to form a polytomy in the parsimonious strict consensus tree. CI = 0.409, RI = 0.644, RC = 0.263.
DISCUSSION
Phylogenetic Identity of the European Trilophosaurids
Our phylogenetic analysis suggests that Tricuspisaurus thomasi and Variodens inopinatus are both trilophosaurids. In previous discussions, some authors suggested Tricuspisaurus thomasi was a procolophonid (Fraser, Citation1986; Sues and Olsen, Citation1993; Edwards, Citation1998; Spielmann et al., Citation2008), or a trilophosaurid (Robinson, Citation1957a; Edwards and Evans, Citation2006; Skinner et al., Citation2020). We demonstrate that both T. thomasi and Variodens inopinatus are indeed trilophosaurids based on the following evidence:
The teeth of Tricuspisaurus thomasi and Variodens inopinatus are attached by fibrous bone to the alveolar rim in a protothecodont (ankylothecodont of Heckert et al. [Citation2006] and Spielmann et al. [Citation2008]) implantation rather than the acrodonty of procolophonids (Cabreira and Cisneros, Citation2009), and the transversely positioned tricuspid nature of the teeth is in common with Trilophosaurus species. Similar tooth implantation is recorded in other allokotosaurians such as Teraterpeton hrynewichorum, Pamelaria dolichotrachela, Shringasaurus indicus, and Azendohsaurus madagaskarensis (Sues, Citation2003; Nesbitt et al., Citation2015; Ezcurra, Citation2016; Sengupta et al., Citation2017).
There is evidence that Tricuspisaurus thomasi had a keratinous rhamphotheca, including highly vascularized bone in the anterior part of the dentary, as in trilophosaurids but not procolophonids. Variodens inopinatus also had highly vascularized bone on the anterior part of the dentary although there is no evidence of a beak.
The tricuspid tooth-bearing region of Trilophosaurus buettneri, Trilophosaurus jacobsi, Tricuspisaurus thomasi, and Variodens inopinatus dentaries consist of many emplacements or sockets for a battery of teeth. The number of these teeth is similar in the four taxa: about 13 (or 14 if the ultimate tooth is included) in Variodens inopinatus, 12–15 in Trilophosaurus buettneri (Demar and Bolt, Citation1981; mean about 12–13, Gregory, Citation1945), 11 in T. jacobsi (Spielman et al., Citation2008), but fewer, eight (perhaps as many as ten), in Tricuspisaurus thomasi. Therefore, despite having considerably smaller dentaries, Tricuspisaurus thomasi and Variodens inopinatus fall within or close to the tooth number range of Trilophosaurus species. In comparison, the multicuspid teeth in the dentary of contemporaneous Late Triassic procolophonids are far fewer, Leptopleuron lacertinum has 5–6 (Säilä, Citation2010) whereas Hypsognathus fenneri, has five (Colbert, Citation1946).
The shape of the multicuspid tooth row in Trilophosaurus species and Variodens inopinatus, in occlusal view of the dentary, is an elongated ellipse and the largest teeth are in the distal half. In comparison, large teeth of the contemporaneous procolophonid dentaries are positioned mesially (such as the ‘massive’ incisiform of Hypsognathus, Sues et al., Citation2000:p. 282), as well as distally (Säilä, Citation2010). The tooth row of the procolophonid Leptopleuron lacertinum is more rectangular in occlusal view (Säilä, Citation2010:fig. 8h); an occlusal view of the Hypsognathus fenneri dentary is not reported. The multicuspid tooth row of Tricuspisaurus thomasi is more triangular and much wider towards the posterior than ellipsoidal in occlusal view (), which is likely due to the pathological damage, but as in Variodens inopinatus, the largest teeth are in the penultimate and antepenultimate (3rd from the most distal) positions. The largest teeth vary in position in Trilophosaurus species and can be 3rd to 5th from the ultimate distal position (see e.g., T. buettneri specimen TMM 31025-125 in Spielman et al. [Citation2008:fig. 27a] and T. jacobsi specimen NMMNH P-41400 in Spielman et al. [Citation2008:fig. 93k, l]). Gregory (Citation1945:fig. 6) reconstructs the dentary of T. buettneri with the largest teeth in the 5th or 6th position from the most distal.
There is a longitudinal bony ridge on the posterior lateral side of the dentary just below the dentigerous region in T. thomasi and V. inopinatus, as in Trilophosaurus buettneri (Spielmann et al., Citation2008:fig. 27f, h, specimen TMM 31025-125). No such dentary ridge is reported or figured in the Late Triassic procolophonids, e.g., Acadiella psalidodon, Haligonia bolodon, or Scoloparia glyphanodon by Sues and Baird (Citation1998). The dentary bony ridge is absent in Libognathus sheddi (Small, Citation1997), Hypsognathus fenneri (Colbert, Citation1946), Leptopleuron lacertinum (Säilä, Citation2010), or the leptopleuronine from Ruthin Quarry described by Skinner et al. (Citation2020:fig. 6k).
The tricuspid teeth of Tricuspisaurus thomasi and Variodens inopinatus (excepting the penultimate tooth of that species) are labiolingually wider than mesiodistally long at the crown base. This is a feature common to all Trilophosaurus species. Tricuspid teeth are found in some procolophonids such as the unnamed taxon from Cromhall Fraser (Citation1986:fig. 6). However, in that specimen and another from Tytherington mistakenly assigned to Tricuspisaurus by Whiteside and Marshall (Citation2008:fig. 5oo, pp), the middle cusp is distinctively mesially positioned, not in a transverse row, and the acrodont teeth lack the cingula and arêtes of the trilophosaurids. It was these unnamed procolophonid multicuspid teeth that led Fraser (Citation1986), Sues and Olsen (Citation1993), Heckert et al. (Citation2006) and Spielmann et al. (Citation2008) to consider that Tricuspisaurus thomasi and/or Variodens inopinatus might be procolophonids rather than trilophosaurs. In comparison, Skinner et al. (Citation2020) and earlier, Edwards (Citation2001), recognized that the procolophonids of the U.K. fissure deposits were quite different to Tricuspisaurus thomasi and Variodens inopinatus and correctly identified those taxa as trilophosaurids.
Both Trilophosaurus jacobsi and Trilophosaurus buettneri as well as Tricuspisaurus thomasi () possess sharp edges (arêtes) between the cusps of their teeth (Heckert et al., Citation2006:fig. 3; Spielmann et al., Citation2008:figs. 4, 8, 9; Skinner et al., Citation2020:fig. 10). Transverse cutting ridges form between the cusps of leptopleuronine procolophonids but where recorded they are curved towards the anterior end of the jaw, e.g., Libognathus sheddi (Small, Citation1997) and the leptopleuronine teeth figured by Skinner et al. (Citation2020:fig. 6e–h). In contrast the cutting edges between cusps in Trilophosaurus species (Spielman et al., Citation2008:fig. 9iI) and in Tricuspisaurus thomasi () are straight.
In Trilophosaurus species, where jawbones are complete or near complete (from figures in Spielmann et al., Citation2008), there is a digitated anterior in medial view (Fig. S3). In Trilophosaurus buettneri the anteriormost region of the dentary is digitated so that an interlocked symphysis is formed between the left and right lower jawbones (Fig. S3A). This digitation is present in Trilophosaurus jacobsi (Fig. S3B) and a CT sagittal slice of the anterior dentary of Tricuspisaurus thomasi (Fig. S2J) indicates that it also has a similar digitation. In fact, the digitation is formed from a sequence of sub-longitudinal ridges and troughs so that in a CT slice a gap between ‘digits’ is observed (see Fig. S2J). Variodens inopinatus also displays the features of an interdigitating dentary symphysis in medial view (); it probably had an interlocking symphysis but as the anteriormost end is damaged we cannot be certain. The symphysis is known to have interdigitated in Tanystropheus (Spiekman et al., Citation2020) and rugose areas and small ridges are formed in the lower part of the symphysis in the rhynchosaurs Rhynchosaurus (Benton Citation1990:fig. 7b) and Teyumbaita sulcognathus (Montefeltro et al., Citation2010:fig. 8). In other taxa we researched, the symphysis appears straight in anterior view indicating smooth opposing surfaces such as in early taxa, e.g., Prolacerta Gow (Citation1975), in the lepidosaurs such as Gephyrosaurus Evans (Citation1980), and in dinosaurs such as Plateosaurus (Lallensack et al., Citation2021:fig. 9g2). In some of these cases the surfaces might be rugose, but in any event, there is no suggestion that they are deeply interlocking as in the trilophosaurids. However, we have not been able to extensively study all taxa in the cladistic matrix for this character and without CT scanning images such as that for Tanystropheus (Spiekman et al., Citation2020) we cannot be certain, so it was not included in our phylogenetic analysis. However, a digitated anterior dentary and therefore an interdigitating symphysis is lacking in all the descriptions and figures of procolophonids we researched in the Triassic. These include the Late Triassic Acadiella psalidodon, Haligonia bolodon, or Scoloparia glyphanodon in Sues and Baird (Citation1998) and a digitated symphysis is absent in Libognathus sheddi (Small, Citation1997), Hypsognathus fenneri (Colbert, Citation1946), and Leptopleuron lacertinum (Säilä, Citation2010). It is also lacking in procolophonids from the Early and Middle Triassic such as Procolina teresae (Borsuk–Bialynicka and Lubka, Citation2009:fig. 9), Kapes bentoni (Zaher et al., Citation2019:fig. 5), and Eomurruna yurrgensis (Hamley et al., Citation2021:fig. 7b).
From our phylogenetic analysis () and supplementary characters and matrix we assign Tricuspisaurus thomasi and Variodens inopinatus to the Trilophosauridae as they have the following synapomorphies in common with the genus Trilophosaurus: non-serrated marginal dentition (Nesbitt et al., Citation2015:Character and state 90-0; here 80-0); morphology of crown base of the marginal teeth a flattened platform with mesiodistally arranged cusps (93-2; here 83-2); tooth shape at crown base of the marginal teeth labiolingually wider than mesiodistally long (98-2; here 87-2).
From our nine new characters, Tricuspisaurus thomasi and Variodens inopinatus have seven synapomorphies with Trilophosaurus. There are two distal and/or mesial cingula only on the mid dentary teeth, i.e., not multi-cingulate on those teeth, (Ch. 223-2; shared only with Trilophosaurus buettneri). There is one mesial and one distal cingulum on all dentary teeth (224-2; shared only with T. buettneri) excepting that Variodens inopinatus has more cingula on the antepenultimate and penultimate teeth. The three cusps are equally or sub-equally spaced (Ch. 225-2; shared with T. buettneri and T. dornorum). The relative cusps are of equal or sub-equal size in a multiple transversely cusped taxon (226-1; shared with T. buettneri and T. phasmalophos) except the antepenultimate and penultimate teeth of Variodens inopinatus where the middle cusp is the highest; the middle cusp is also highest in T. jacobsi teeth and one flanking cusp is noticeably lower in T. dornorum (Mueller and Parker, Citation2006:fig. 4). The dentary is mediolaterally thin but dorsally is abruptly greatly expanded (greater than 2× mid dentary) forming a ridge both laterally and medially behind the mid-region (227-1; known in T. buettneri and Teraterpeton hrynewichorum only, data are lacking in other trilophosaurids). In lateral view, the observed distalmost dentary tooth base is noticeably more dorsally positioned than the penultimate tooth, placed about half the height of the penultimate tooth cusp on the anterior of coronoid process (228-1; known in T. buettneri only, data lacking in other trilophosaurids). On the dentary teeth, the bone of attachment reaches a pronounced height above the dentary dorsal margin (230-1, known in T. buettneri and T. hrynewichorum; data are lacking in other trilophosaurids), but also present in Azendohsaurus madagaskarensis.
Additionally, Tricuspisaurus thomasi, but not Variodens inopinatus, shares one apomorphy with all Trilophosaurus species: a lengthy transverse cutting ridge (arête) between all cusps on at least some dentary teeth. Tricuspisaurus thomasi also shares the apomorphy (229-1) with Trilophosaurus buettneri and Trilophosaurus jacobsi: there is prominent anterior vascularization of the dentary marked by many pits and foramina with a rugosity suggesting a keratinous rhamphotheca, but V. inopinatus shows no indication of such a beak. Rhynchosaurus, Tricuspisaurus thomasi, and V. inopinatus are the only taxa to share the synapomorphy (231-1): the largest dentary tooth is in the penultimate or antepenultimate position.
Insular Dwarfism
Compared with other trilophosaurids, Tricuspisaurus thomasi and Variodens inopinatus are small animals, their dentaries measuring ∼2–3 cm in length. Trilophosaurus buettneri dentaries exceed more than twice this length; the dentary of T. jacobsi is about 60% longer. Skinner et al. (Citation2020:559) suggested that T. thomasi was much smaller than its close relative Trilophosaurus, ∼0.6–1 m, compared with 2–2.5 m for the latter, based on relative jaw sizes. Variodens inopinatus is smaller still, with an estimated body length of about 0.45 m. Our findings therefore agree with the hypothesis that insular dwarfism was a common feature of many tetrapods from the Bristol paleoislands (Klein et al., Citation2015; Whiteside et al., Citation2016; Skinner et al., Citation2020).
CONCLUSIONS
We provide evidence that Tricuspisaurus thomasi and Variodens inopinatus are trilophosaurids and are the youngest currently known members of Trilophosauridae, being the only examples with a probable Rhaetian age. There is an unconfirmed report of a worn trilophosaurid tooth from the Rhaetian Owl Rock Member of the Chinle Formation (U.S.A.) by Kirby (Citation1991) but this identification is generally queried (e.g., Kligman et al., Citation2020). We confirm that the gap in the tooth row in the holotype of T. thomasi is pathological and not an autapomorphy. The two European taxa are relatively small compared with other described trilophosaurids, in accord with the notion that they are insular dwarfs.
Supplemental Material
Download Zip (35.8 MB)ACKNOWLEDGMENTS
We thank T. Davies for having created and processed the original CT-scan dataset of NHMUK PV R6106, used here. The CT-scan dataset is the property of the NHMUK and permission to use it should be sought from that institution. We also acknowledge the Tratman Scholarship for funding SAVC-T’s PhD, and NERC grant NE/I027630/1 to M.J.B. We thank M. Day and S. Chapman for their great help and permission to study the collections of the NHMUK. We are grateful to F. Feneru for guidance in the use of the macro photography equipment at the Angela Marmont Centre for UK Biodiversity (NHMUK). We also thank Martin Ezcurra, Adam Marsh and an anonymous referee, as well as the JVP editors, for their very thorough and helpful comments.
Related Research Data
LITERATURE CITED
- Benton, M. J. 1990. The species of Rhynchosaurus, a rhynchosaur (Reptilia, Diapsida) from the Middle Triassic of England. Philosophical Transactions of the Royal Society of London B 328:213–306.
- Borsuk–Bialynicka, M., and M. Lubka. 2009. Procolophonids from the Early Triassic of Poland. Palaeontologia Polonica 65:107–144.
- Cabreira, S.F. and J.C. Cisneros. 2009. Tooth histology of the parareptile Soturnia caliodon from the Upper Triassic of Rio Grande do Sul. Brazil. Acta Palaeontologica Polonica 54:743–748.
- Colbert, E. H. 1946. Hypsognathus, a Triassic reptile from New Jersey. Bulletin of the American Museum of Natural History 86, 225–74.
- Demar, R and J. R. Bolt. 1981. Dentitional organization and function in a Triassic reptile. Journal of Paleontology 55:967–984
- Edwards, B. T. 1998. An unusual Upper Triassic microvertebrate assemblage from South West Britain. Journal of Vertebrate Palaeontology 18 (3):39A.
- Edwards, B. T. 2001. The Upper Triassic Microvertebrate Assemblage of Ruthin Quarry, South Wales. Unpublished Ph. D dissertation, University College London.
- Edwards, B. T., and S. E. Evans. 2006. A Late Triassic microvertebrate assemblage from Ruthin Quarry, Wales: pp. 33–35 in P. M. Barrett, and S. E. Evans (eds.). Ninth International Symposium on Mesozoic Terrestrial Ecosystems and Biota, Abstracts and Proceedings. Natural History Museum, London.
- Evans, S. E., 1980. The skull of a new eosuchian reptile from the Lower Jurassic of South Wales. Zoological Journal of the Linnean Society 70:203–264.
- Evans, S. E., and K. A. Kermack. 1994. Assemblages of small tetrapods from the early Jurassic of Britain: pp. 271–282 in N. C. Fraser, and H.-D. Sues (eds). In the Shadow of the Dinosaurs. Cambridge University Press, New York.
- Ezcurra, M. D. 2016. The phylogenetic relationships of basal archosauromorphs, with an emphasis on the systematics of proterosuchian archosauriforms. PeerJ 4:e1778 https://doi.org/https://doi.org/10.7717/peerj.1778
- Fraser, N. C. 1986. Terrestrial vertebrates at the Triassic-Jurassic boundary in Southwest Britain. Modern Geology 9:273–300.
- Goloboff, P. A. and S. A. Catalano. 2016. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics 32:221–238.
- Gow, C.E. 1975. The morphology and relationships of Youngina capensis Broom and Prolacerta broomi Parrington. Palaeontologia Africana 18:89–131.
- Gregory, J. T. 1945. Osteology and relationships of Trilophosaurus. University of Texas 4401:273–359.
- Hamley, T., J. C. Cisneros, and R. Damiani. 2021. A procolophonid reptile from the Lower Triassic of Australia. Zoological Journal of the Linnean Society 192:554–609.
- Heckert, A. B., S. G. Lucas, L. R. Rinehart, J. A. Spielmann, A. P. Hunt, and R. Kahle. 2006. Revision of the archosauromorph reptile Trilophosaurus, with a description of the first skull of Trilophosaurus jacobsi, from the upper Triassic Chinle Group, West Texas, USA. Palaeontology 49:621–640.
- Huelsenbeck, J. P., and F. Ronquist. 2001. MrBayes: bayesian inference of phylogenetic trees. Bioinformatics 17:754–755. doi: https://doi.org/10.1093/bioinformatics/17.8.754
- Huene, F. 1946. Die grossen Stämme der Tetrapoden in den geologischen Zeiten. Biologisches Zentralblatt 65:268–275.
- Kirby, R. E. 1991. A vertebrate fauna from the Upper Triassic Owl Rock Member of the Chinle Formation of Northern Arizona. Unpublished M.Sc. thesis, Northern Arizona University. 476 pp.
- Klein, C. G., D. I. Whiteside, V. Selles de Lucas, P. A. Viegas, and M. J. Benton. 2015. A distinctive Late Triassic microvertebrate fissure fauna and a new species of Clevosaurus (Lepidosauria: Rhynchocephalia) from Woodleaze Quarry, Gloucestershire, UK. Proceedings of the Geologists’ Association 126:402–416.
- Kligman, B. T., A. D. Marsh, S. J. Nesbitt, W. G. Parker, and M. R. Stocker. 2020. New trilophosaurid species demonstrates a decline in allokotosaur diversity across the Adamanian-Revueltian boundary in the Late Triassic of western North America. Palaeodiversity 13:25–37.
- Lallensack, J. N., E. M. Teschner, B. Pabst, and P. M. Sander. 2021. New skulls of the basal sauropodomorph Plateosaurus trossingensis from Frick, Switzerland: Is there more than one species? Acta Palaeontologica Polonica 66:1–28.
- Lovegrove, J., A. J. Newell, D. I. Whiteside, and M. J. Benton. 2021. Testing the relationship between marine transgression and evolving island palaeogeography using 3D GIS: an example from the Late Triassic of SW England. Journal of the Geological Society 178:jgs2020–158.
- Montefeltro, F. C., M. C. Langer, and C. L. Schultz. 2010. Cranial anatomy of a new genus of hyperodapedontine rhynchosaur (Diapsida, Archosauromorpha) from the Upper Triassic of southern Brazil. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 101:27–52.
- Mueller, B. D., and W. G. Parker. 2006. A new species of Trilophosaurus (Diapsida: Archosauromorpha) from the Sonsela Member (Chinle Formation) of Petrified Forest National Park, Arizona. Museum of Northern Arizona Bulletin 62:119–125.
- Murry, P. A. 1987. New reptiles from the Upper Triassic Chinle Formation of Arizona. Journal of Paleontology 61:773–786
- Nesbitt, S. J., J. J. Flynn, A. C. Pritchard, J. M. Parrish, L. Ranivoharimanana, and A. R. Wyss. 2015. Postcranial osteology of Azendohsaurus madagaskarensis (? Middle to Upper Triassic, Isalo Group, Madagascar) and its systematic position among stem archosaur reptiles. Bulletin of the American Museum of Natural History 398:1–126.
- Pritchard, A. C., A. H. Turner, S. J. Nesbitt, R. B. Irmis, and N. D. Smith. 2015. Late Triassic tanystropheids (Reptilia, Archosauromorpha) from northern New Mexico (Petrified Forest Member, Chinle Formation) and the biogeography, functional morphology, and evolution of Tanystropheidae. Journal of Vertebrate Paleontology 35:e911186.
- Robinson, P. L. 1957a. An unusual sauropsid dentition. Zoological Journal of the Linnean Society 43:283–293.
- Robinson, P. L. 1957b. The Mesozoic fissures of the Bristol Channel area and their vertebrate faunas Journal of the Linnean Society (Zoology) 43:260–282.
- Robinson, P. L. 1962. Gliding lizards from the Upper Keuper of Great Britain. Proceedings of the Geological Society of London 1601:137–146.
- Robinson, P. L. 1971. A problem of faunal replacement on Permo-Triassic continents. Palaeontology 14:131–153.
- Ronquist, F., M. Teslenko, P. Mark, D. Ayres, A. Darling, S. Hohna, B. Larget, L. Lui, M. Suchard, and J. Huelsenbeck. 2012. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Systematic Biology 61:539–542.
- Säilä, L.K., 2010. Osteology of Leptopleuron lacertinum Owen, a procolophonoid parareptile from the Upper Triassic of Scotland, with remarks on ontogeny, ecology and affinities. Earth and Environmental Science Transactions of the Royal Society of Edinburgh 101:1–25.
- Savage, R. J. G. 1977. The Mesozoic strata of the Mendip Hills: pp. 85–100 in R. J. G. Savage (ed.). Geological Excursions in the Bristol District. University of Bristol, Bristol.
- Sengupta, S., M. D. Ezcurra, and S. Bandyopadhyay. 2017. A new horned and long-necked herbivorous stem-archosaur from the Middle Triassic of India. Scientific Reports 7(1):8366.
- Skinner, M., D. I. Whiteside, and M. J. Benton. 2020. Late Triassic island dwarfs? Terrestrial tetrapods of the Ruthin fissure (South Wales, UK) including a new genus of procolophonid. Proceedings of the Geological Association 131:535–561.
- Small, B. J. 1997. A new procolophonid from the Upper Triassic of Texas, with a description of tooth replacement and implantation. Journal of Vertebrate Paleontology 17:674–678.
- Spiekman, S. N. F, J. M, Neenan, N. C. Fraser, V. Fernandez, O. Rieppel, S. Nosotti, and T. M. Scheyer. 2020. The cranial morphology of Tanystropheus hydroides (Tanystropheidae, Archosauromorpha) as revealed by synchrotron microtomography. PeerJ 8:e10299 DOI https://doi.org/10.7717/peerj.10299
- Spielmann, J. A., S. G. Lucas, L. F. Rinehart, and A. B. Heckert. 2008. The Late Triassic archosauromorph Trilophosaurus. Bulletin of the New Mexico Museum of Natural History and Science 43:1–177.
- Spielmann, J. A., S. G. Lucas, A. B. Heckert, L. F. Rinehart and A. P. Hunt. 2007. Taxonomy and biostratigraphy of the Late Triassic archosauromorph Trilophosaurus. Bulletin of the New Mexico Museum of Natural History and Science 40:231–240.
- Spielmann, J. A., S. G. Lucas, A. B. Heckert, L. F. Rinehart, and H. R. Richards. 2009. Redescription of Spinosuchus caseanus (Archosauromorpha: Trilophosauridae) from the Upper Triassic of North America. Palaeodiversity 2:283–313.
- Sues, H.-D. 2003. An unusual new archosauromorph reptile from the Upper Triassic Wolfville Formation of Nova Scotia. Canadian Journal of Earth Sciences 40:635–649.
- Sues, H.-D. and D. Baird. 1998. Procolophonidae (Reptilia: Parareptilia) from the Upper Triassic Wolfville formation of Nova Scotia. Journal of Vertebrate Paleontology 18:525–32.
- Sues, H.-D., and P. E. Olsen. 1993. A new procolophonid and a new tetrapod of uncertain, possibly procolophonian affinities from the Upper Triassic of Virginia. Journal of Vertebrate Paleontology 13:282–286.
- Sues, H.-D., P. E. Olsen, D. M. Scott, and P. S. Spencer. 2000. Cranial osteology of Hypsognathus fenneri, a latest Triassic procolophonid reptile from the Newark Supergroup of eastern North America. Journal of Vertebrate Paleontology 20:275–84.
- Swofford, D. L. 2002. PAUP: Phylogenetic Analysis Using Parsimony (and Other Methods), Version 4.0 Beta 10. Sinauer Associates, Sunderland.
- Thomas, T. M. 1952. Notes on the structure of some minor outlying occurrences of littoral Trias in the Vale of Glamorgan. Geological Magazine 89:153–162.
- Whiteside, D. I., and C. J. Duffin. 2017. Late Triassic terrestrial microvertebrates from Charles Moore’s ‘Microlestes’ quarry, Holwell, Somerset, UK. Zoological Journal of the Linnean Society 179:677–705.
- Whiteside, D. I., and J. E. A. Marshall. 2008. The age, fauna and palaeoenvironment of the Late Triassic fissure deposits of Tytherington, South Gloucestershire, UK. Geological Magazine 145:105–147.
- Whiteside, D. I., C. J. Duffin, P. G. Gill, J. E. A. Marshall, and M. J. Benton. 2016. The Late Triassic and Early Jurassic fissure faunas from Bristol and South Wales: Stratigraphy and setting. Palaeontologia Polonica 67:257–287.
- Wynd, B. M., R. N. Martínez, C. Colombi, and O. Alcober, 2020. A review of vertebrate beak morphologies in the Triassic; a framework to characterize an enigmatic beak from the Ischigualasto Formation, San Juan, Argentina. Ameghiniana 57:370–387.
- Zaher, M., R. A. Coram, and M. J. Benton. 2019. The Middle Triassic procolophonid Kapes bentoni: computed tomography of the skull and skeleton. Papers in Palaeontology 5:111–138.