Abstract
New Zealand (NZ) sea lions (Phocarctos hookeri) were relisted as ‘nationally critical’ by the New Zealand Government in 2010, owing to recent severe declines in pup production at the Auckland Islands. However, research on Campbell Island/Motu Ihupuku indicates that this population's pup production is increasing. Between December 2009 and February 2010, surveys were undertaken aiming to determine production and early mortality rates (up to eight weeks) of NZ sea lion pups on Campbell Island. In total, 402 pups were tagged and a minimum pup production of 681 estimated from direct count. This estimate is 17% higher than the 2008 estimate of 583 pups and equates to 27% of the total pup production of the NZ sea lions in 2009–10. However, early pup mortality (55%) continues to be the highest recorded for any NZ sea lion breeding area and it is hypothesised that this breeding area is unlikely to be able to maintain this pup production increase.
Introduction
The New Zealand (NZ) sea lion (Phocarctos hookeri) is New Zealand's only endemic pinniped. In 2010, the NZ sea lion was classified as ‘nationally critical’ (Baker et al. Citation2010), the highest threat classification before extinction, owing to severely declining pup production at the Auckland Islands (50% decrease in pup production between 1989 and 2009; ). The breeding range of the NZ sea lion is one of the most restricted of any otariid, and is centred on the New Zealand sub-Antarctic islands (between latitudes 48°S and 53°S). The largest breeding population occurs in the Auckland Islands, where previous estimates suggest that 79% of pups are born (Maloney et al. Citation2009). The other significant breeding population is at Campbell Island/Motu Ihupuku (hereafter referred to as Campbell Island), where 21% of pups are born (Maloney et al. Citation2009). The only other area where breeding occurs consistently is Otago Peninsula, where a small single matriarchal line of breeding has begun (McConkey, Heinrich et al. Citation2002; McConkey, McConnell et al. Citation2002).
Figure 1 Pup production estimates from Auckland Islands (1998–2010 [Chilvers 2011]) and completed pup production estimates Campbell Island/Motu Ihupuku (2003–10).
![Figure 1 Pup production estimates from Auckland Islands (1998–2010 [Chilvers 2011]) and completed pup production estimates Campbell Island/Motu Ihupuku (2003–10).](/cms/asset/25d3a820-3396-4420-b607-d6fea9bd05f0/tnzz_a_614265_o_f0001g.jpg)
The present sub-Antarctic distribution of NZ sea lions is thought to be a relict population at the extreme south of the natural range for this species (Childerhouse & Gales Citation1998). The largest remaining breeding population, in the Auckland Islands, has been monitored annually since 1995 (Gales & Fletcher Citation1999). The number of pups born within this population decreased by 50% between 1998 and 2009 (), with the lowest ever recorded pup production numbers in 2009 (1501 pups), which was a 30% decrease in pup production from 2008. The decrease in pup production has been directly related to philopatric females not returning to breeding areas (Chilvers Citation2010).
There have been infrequent attempts to monitor pup production and the NZ sea lion population on Campbell Island (Bailey & Sorenson Citation1962; Russ Citation1980; Moore & Moffatt Citation1990; Cawthorn Citation1993; McNally et al. Citation2001; Childerhouse et al. Citation2005), which appears to have an increasing pup production. Methodologies have varied between expeditions, so direct comparisons of pup production are difficult to make. The NZ sea lion pup production and population on Campbell Island was last surveyed in January and February 2008, at which time pup production was estimated to be 583 (Maloney et al. Citation2009). A 2009–10 expedition aimed to build on the 2008 research, using comparable methodology to see if a pup production trend could be established, further examine reasons for early pup mortality and investigate if the same 30% decline, as seen in the Auckland Island population between 2008 and 2009, occurred at Campbell Island.
Methods
Research was undertaken on Campbell Island (52°33′S, 169°09′E; ) between 17 December 2009 and 18 February 2010. The mean parturition date of NZ sea lions at the Auckland Islands is 26–27 December, with 69% of pups born one week either side of these dates (Chilvers, Robertson et al. 2007). Our survey was timed to start early in the pupping season than the 2008 survey (which started 4 January) to allow the best possible estimate of pup production and early mortality.
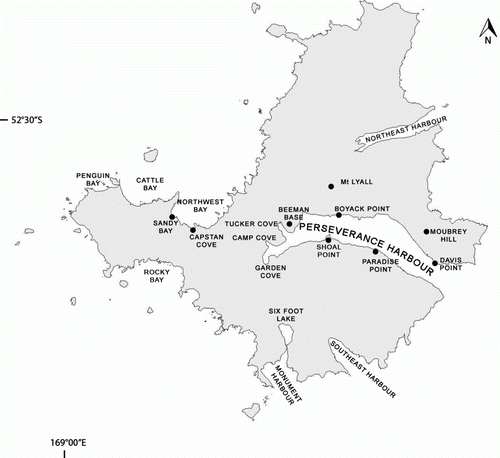
Figure 2 Campbell Island/Motu Ihupuku, showing all locations of pup and tagged New Zealand sea lion sightings from December 2009 to February 2010.
The research was divided into four sections:
| 1. | An investigation of the coastline to search for previously unknown NZ sea lion breeding areas from 17 to 19 December | ||||
| 2. | Direct counts, tagging and necropsies (gross examination only) at the main breeding colony, Davis Point, from 20 December to 14 January | ||||
| 3. | Searching for and tagging pups at the smaller Paradise Point breeding colony from 17 to 21 January | ||||
| 4. | Searching the rest of Campbell Island for pups born away from the main colonies from 22 January to 18 February. During the final period, return visits were also made to Davis and Paradise Points to check for further mortality and any untagged live pups. | ||||
During research period 1 (17–19 December), the research team conducted a visual survey of much of the accessible coastline of Campbell Island from the sailing vessel Tiama to search for previously unknown breeding colonies. Much of the coastline, particularly in the northern half of the island, is inaccessible to NZ sea lions (and humans) owing to sheer cliffs (McNally et al. Citation2001). The team searched the coast from Northeast Harbour in the northeast to Rock Bay on the southwest coast (). Landings were made where pups had been sighted during previous expeditions (McNally et al. Citation2001; Childerhouse et al. Citation2005), at the head of Northeast Harbour, at Monument Harbour/Six Foot Lake, and at Southeast Harbour (). A landing was also made at Paradise Point to make an early count of adult and pup numbers, as Paradise Point was otherwise not visited until later in the season.
During research period 2 (20 December to 14 January), research effort was concentrated on the largest breeding colony at Davis Point. Daily counts were made of females, sub-adult males, adult males and pups. Accessible pups were tagged on both pectoral flippers with uniquely numbered green, ‘coffin’-shaped Dalton Jumbotags (Dalton ID systems, Oxon, UK). Where possible, the tag numbers of adult and sub-adult animals with tags were recorded—NZ sea lion pups have previously been tagged at Campbell Island in 1998 (McNally et al. Citation2001), 2003 (Childerhouse et al. Citation2005) and 2008 (Maloney et al. Citation2009), and at the Auckland Islands annually since 1998–99 (Chilvers, Wilkinson et al. 2007).
Daily counts were also made of dead pups. Where practical, dead pups were removed from the colony area using an extendable pole and hook (and usually then thrown into the sea so they could not be recounted as the steep rocky area surrounding the breeding area would prevent any carcasses being washed back ashore). As it was not possible to remove all bodies, a map was drawn each day with the position of each carcass. In this way, the cumulative body count was made as accurately as possible. Necropsies (gross examinations only) were carried out on three to six carcasses a day (the size of the daily mortality precluded performing necropsies on all carcasses). Only fresh carcasses (those estimated to be two days old or less) were necropsied. The weights of dead pups (only) were measured to the nearest 0.1 kg using an electronic spring balance (Berkeley); and length, girth and blubber depth were measured using a tailor's tape measure. Notes were made of external and internal abnormalities, and contributing causes of death.
For research period 3 (17–21 January), the smaller breeding colony at Paradise Point was visited, with the intention of tagging as many pups as possible and counting dead pups to get the best possible estimate of breeding at this site. No attempt was made to perform necropsies at this colony owing to the logistics of getting equipment and personal to the site. The colony at Paradise Point was first visited and identified on 28 January 2008 (Maloney et al. Citation2009). This expedition started its survey 11 days earlier, to get a more accurate count of pup numbers, as pups disperse from the breeding area to surrounding thick bush around this time (pers. obs. A. Maloney, Veterinarian, 2008).
Lastly, during research period 4 (28 January to 18 February—the end of this expedition), other NZ sea lion accessible parts of the island were searched on foot. Particular attention was paid to places where pups had been found in 2008 (Maloney et al. Citation2009), and areas not visited during research period 1 of this expedition, including Northwest Bay, Cattle Bay and Penguin Bay (). An additional visit was also made to Southeast Harbour. Regular checks were made of the coastline at the head of Perseverance Harbour, from Tuckers Cove to Shoal Point ().
A return visit to Davis Point was made on 26 January and to Paradise Point on 10 February to count, search for untagged pups and remove tags from new pup carcasses (pups that were tagged were counted when alive and were not recounted when tags removed as carcasses). During this phase of the expedition, all untagged pups were caught and tagged (with the exception of one inaccessible pup at Paradise Point), and tag numbers of previously tagged animals were recorded where possible.
Results
During the 2009–10 survey, 402 live pups were tagged, 1 live pup was unable to be tagged on last day of expedition, and 278 untagged dead pups were counted (some tagged pups also died, but these are not included in the dead pups to avoid counting the same individual twice). This gives a minimum estimate of pup production of 681. The number of pups at each location is shown in . The number of pups seen and not tagged (and, therefore, not counted to avoid double counting) was only two. The sex ratio of the tagged pups was 0.82:1 (female:male). In total, 375 dead pups (tagged and untagged) were counted on the island, giving a known pup mortality rate on 18 February (the expedition end) of 55%. Of those pups tagged, 45 male pups died (20% of male pups tagged) and 52 female pups died (29% of female pups tagged).
Table 1 Location and number of pups directly counted at various locations around Campbell Island (see ), 18 December 2009 to 16 February 2010.
Davis Point
The Davis Point colony occupied the same area as in 2008—an exposed basalt platform on the north side of Perseverance Harbour (Maloney et al. Citation2009). As in 2008, the majority of known births at Campbell Island occurred here in 2009–10, with a minimum pup production estimate of 503 (74% of the island total). The cumulative dead pup count to 26 January was 319, giving an early mortality rate of 63%.
Paradise Point
The Paradise Point colony appears to be less geographically stable than that at Davis Point. Approximately one-third of the animals pupped at the original site discovered in 2008—an exposed slope jutting out into the south side of Perseverance Harbour (hereafter referred to as Paradise Point 1); the remaining animals were found to have moved approximately 300 m to the west (hereafter referred to as Paradise Point 2), where harems initially formed on the narrow rocky shore and the adjacent slopes (a more sheltered habitat consisting of Blechnum fern and Dracophyllum forest). The minimum total pup production at Paradise Point (Paradise Point 1 and Paradise Point 2 combined) was 168 (25% of the island total). The cumulative dead pup count to 10 February was 54, giving a mortality rate of 32%. Of this, Paradise Point 1 had a pup production of 54, which included 33 dead pups—a mortality rate of 61%; and Paradise Point 2 had a pup production of 114, including 21 dead pups—a mortality rate of just 18%.
Other parts of Campbell Island
A total of 10 pups were found away from the main colonies, of which 2 were dead and the remaining 8 were tagged. This is just 1% of total known pup production. Pups were found from sea level to 250 m above sea level, in habitat ranging from seashore to fellfield on high ridges, over 2 km from the sea. Some were suspected to be close to their place of birth, while others (including the three found in the post-breeding harems at Tucker Cove) were more likely to have been moved there by their mothers from other parts of the island. Of all pups tagged at either Davies or Paradise Point, seven pups were later re-sighted with their mothers (by 10 February 2011) away from their tagging sites (from Davies Point to Paradise Point, or Davies or Paradise Points to Beeman Base). No other breeding aggregations were found away from the Davies and Paradise colonies.
Necropsies
Forty-nine pups were necropsied at the Davis Point colony between 21 December 2009 and 13 February 2010. shows factors likely to have contributed to pup death as determined by gross examination. These factors add up to more than 49 as many pups had more than one pathology considered to have contributed to mortality. For example, several pups had traumatic bite wounds that subsequently became infected, and concurrent hookworm infection.
Table 2 Factors likely to have contributed to New Zealand sea lion pup deaths. Diagnoses were based on necropsies undertaken between 21 December 2009 and 13 January 2010 at Davies Point, Campbell Island.
Only a small proportion of the dead pups were necropsied, owing to the very high death rate. The causes of death recorded here do not, therefore, accurately represent the true percentages of these pathologies across the entire season's dead pups. For example, on 2 January, after a night of extreme weather (2 °C at 0800 h, gale-force winds and rain), 57 fresh dead pups were found. Owing to time limitations, only six of these were necropsied, of which five had no abnormalities detected and were considered to have perished from exposure. It is likely that the majority of the other dead pups that day also perished from exposure.
Of the pups necropsied, the most common diagnosis was trauma (22 pups, 44%), with presentations including cranial bruising and liver rupture (associated with being trampled by adults), and bite wounds. Infection was diagnosed as the primary cause of death for eight pups (16%), but this is likely to be an underestimate, as this figure only includes animals with gross pathology (including abscesses, pneumonia, pleurisy and pericarditis). Malnutrition was diagnosed as the primary cause of death for 18 pups (36%), and was defined as pups with no visible fat in the blubber layer. Hookworm was also common, with 14 pups affected (28%), although this average figure does not reflect the fact that hookworm was becoming near-ubiquitous towards the end of the necropsy study. Five pups died from exposure, but this is also likely to have been a contributing factor in the deaths of many other pups, particularly the drowning and malnutrition diagnoses.
An extremely common, non-lethal pathology found in necropsied pups was ulceration of the carpus on the fore flippers, and of various parts of the hind flippers—19 pups (38%; ). While most of these were relatively minor, some may have limited pup mobility and contributed to death by trauma, or allowed access for infection. One pup had such severe ulceration to both carpi that the joint capsules were open, and the bones and cartilage were partially worn away.
Figure 3 New Zealand sea lion pup at Davis Point showing ulceration of the carpus on the fore flippers. Photo: C. Muller.

Re-sightings
There were 87 re-sights of previously tagged animals aged between 2 and 12 years during the entire survey period. Of these, 29 were identified as males and 32 as females. All females were tagged as pups at Campbell Island, while 23 of the re-sighted males were tagged as pups at Campbell Island and 6 were tagged as pups from the Auckland Island group.
Discussion
This survey provided a pup production estimate of 681 for NZ sea lions at Campbell Island, which is 17% higher than the largest and most recent previous estimate of 583 (; Maloney et al. Citation2009). It also provided the highest early pup mortality rate recorded in NZ sea lions by late January (55%)—higher than the 40% mortality recorded during the 2008 expedition (Maloney et al. Citation2009), and the average mortality (not including epidemic years) of 7% at Enderby Island, Auckland Islands, up to one month after the mean pupping date (Chilvers, Wilkinson et al. 2007). Indeed, this value is well in excess of the highest recorded mortality rate ever recorded at Enderby Island (42%) during the mass mortality event of 1997–98 (Baker Citation1999).
Despite the very high mortality rate, pup production at the main colonies appears to have increased between the 2008 and 2009–10 seasons. The higher pup production figures for both Davis Point and Paradise Point between 2008 and 2010 may in part be inflated owing to an earlier arrival of the research team for the 2009–10 season and hence a more accurate cumulative pup count. This result is in contrast to the decline in pup production at the Auckland Islands during the same period (; Chilvers Citation2011).
Campbell Island pup mortality
Campbell Island is at the extreme southern end of the natural range of NZ sea lions (Worthy Citation1994; Gill Citation1998; Childerhouse & Gales Citation1998). During the NZ austral summer (December–January, when pups are born and most vulnerable), mean and minimum daily temperatures at Campbell Island are 6.7 °C and 9.2 °C, respectively, compared with 7.7 °C and 10 °C for Enderby Island, or 10.8 °C and 13.7 °C for Dunedin, New Zealand (the areas where other NZ sea lion pups are born). Similarly, rainfall during the same period averages 110 mL/month with rain falling on average 18 days per month, compared with 93 mL/month and 15 days for Enderby Island or 57 mL/month and only 6 days in Dunedin (NIWA National Climate Database, http://cliflo.niwa.co.nz/). Young pups, with their small body size and little blubber, are therefore at greater risk from the negative effects of low temperatures and extreme weather at Campbell Island relative to their previous natural distribution across all of New Zealand (Childerhouse & Gales Citation1998).
Research indicates that exposure is an issue at these southern latitudes, with an overall early pup mortality of 55%. However, there were also large differences between breeding sites—the highest mortality being 63% at Davis Point, 61% at Paradise Point 1 and 18% at Paradise Point 2. This is considered to be because of Davis and Paradise Point's harsh habitat (see photos in Maloney et al. 2009), with step slopes and basalt headlands jutting into Perseverance Harbour being extremely exposed to wind, rain and swells (), and Davis Point in particular having many rock pools and peat mires in and around the breeding area which trap and drown pups. In contrast, the Paradise Point 2 breeding area is not a headland, has a gentle slope and is well vegetated.
The basalt habitat of Davis Point is likely to be a reason for the high trauma rate (44%) of necropsied pups owing to its unyielding nature. The rock is also highly abrasive, leading to ulcerations of the carpi (on the fore flippers) and/or various parts of the hind flippers in 38% of necropsied pups (). Lesions like these were not observed in any pups at Paradise Point or anywhere at the Auckland Islands and have never previously been reported for any other sea lion species. Flipper lesions may allow infection (such as hookworm) to enter the body (Castinel, Duignan, Lyons et al. 2007; Castinel, Duignan, Pomroy et al. 2007) and, in some cases, were so severe that they are likely to have restricted movement, thus increasing the pups’ likelihood of death or injury by trauma.
Despite these habitat challenges for NZ sea lion pups, death owing to exposure appears to be the major cause of high mortality at Davis Point. Acute exposure was a significant cause of death during a storm on 2 January 2010, with 57 pups dying overnight (18% of Davis Point's early mortality in one night). However, chronic exposure is a more significant factor overall. Of all necropsied pups, 36% were diagnosed with malnutrition (blubber layer contained no visible fat); similarly 44% of pups were diagnosed with malnutrition during the 2008 season (Maloney et al. Citation2009).
Davis Point colony
Given the consistently high pup mortality seen at Davis Point for over a decade (44% [McNally et al. 2001]; 36% [Childerhouse et al. 2005]; 44% [Maloney et al. 2009], 63% [present study]) how is it that the number of animals breeding at Davis Point appears to be increasing?
We know that the Davis Point breeding area is not increasing owing to emigration of female NZ sea lions from the Auckland Islands, because female NZ sea lions show high philopatry (Chilvers & Wilkinson Citation2008) and no tagged female NZ sea lions from Campbell or Auckland Islands have been sighted at the other island (Childerhouse et al. Citation2005; Chilvers & Wilkinson Citation2008; Maloney et al. Citation2009).
The most likely reason for an increase in pup production at Davis Point is that this area is a population sink, maintaining its size from the immigration of females who have been born or have previously bred elsewhere on Campbell Island, similar to what has been observed for South American sea lions (Otaria flavescens) (Cassini & Fernández-Juricic Citation2003). Davis Point is a relatively recent breeding site for NZ sea lions, first identified in November 1991 (pers. comm., J. Amey, Department of Conservation, 1991). Numbers have increased since then, at a rate faster than could have occurred as a result of a natural increase (births–deaths) from the small numbers (55 females) observed in 1991. For sea lions, it is thought that when populations are low, it is more advantageous for females not to breed colonially, but rather to pup alone in secluded locations to avoid excessive attention from males (Cassini & Fernández-Juricic 2003; Chilvers et al. 2005). However, as populations grow it is more advantageous for females to breed colonially, seeking protection in numbers rather than seclusion, as is documented for South American sea lions (Cassini & Fernández-Juricic 2003).
During breeding, females are known to be attracted to the sight of other females on land (Augé & Chilvers Citation2010). As the population at the very visible Davis Point increases, young females may be attracted to colonies rather than breeding in isolated locations. However, eventually this source of locally emigrating animals will naturally decrease and most animals will be colonial breeders as seen on the Auckland Islands (Chilvers, Wilkinson et al. 2007). At that stage, the high early pup mortality and other known impacts such as fisheries-related mortality and naturally occurring epizootic events known to affect NZ sea lion populations (Wilkinson et al. Citation2003, Citation2006; Robertson & Chilvers Citation2011) will result in a decrease in pup production numbers and a decline in the population.
NZ sea lions at Campbell Island versus Auckland Islands
Even with high pup mortality levels, pup production on Campbell Island, and presumably the population of NZ sea lions, is increasing (; Childerhouse et al. Citation2005; Maloney et al. Citation2009). It is apparent from this research that the pup production at Campbell Island did not have a 30% decrease in pup production between 2008 and 2010, as seen at the Auckland Islands (; Chilvers Citation2011). Overall, there has been a large decline in pup production at the Auckland Islands (Chilvers Citation2010; Chilvers & Mackenzie Citation2010). The differences in pup production () are unlikely to be a result of disease epizootics that have affected NZ sea lions in the past (Baker Citation1999; Wilkinson et al. Citation2003, Citation2006) as it is thought to have affected both island populations, although the severity is unknown at Campbell Island. The largest known and documented difference in mortality (particularly adult female mortality) between the two island groups is the significantly higher fisheries by-catch mortality of NZ sea lions around the Auckland Islands compared with Campbell Island (Robertson & Chilvers Citation2011). It is estimated that up to 140 NZ sea lions are killed annually in interactions with the arrow squid (Nototodarus sloanii) and scampi (Metanephrops challengeri) trawl fisheries around the Auckland Islands, with a female bias in deaths in the fisheries (Chilvers Citation2008; Thompson et al. Citation2010; Roberston & Chilvers 2011). In comparison, an average of 11 animals per year are estimated to be killed in the southern blue whiting (Micromesistius autralis) fishery around Campbell Island, which is predominantly biased toward juvenile male NZ sea lions (Thompson et al. Citation2010).
Conclusions
The pup production of NZ sea lions on Campbell Island has been increasing, with this survey recording the highest minimum pup production figure yet of 681 pups, equating to 27% of total NZ sea lion pup production for the 2009–10 breeding season. The absolute pup production is likely to be higher owing to this expedition's limited access to Paradise Point during peak pupping and the unknown number of non-colonial breeding females, which could be scattered across the island—a breeding pattern not known in the Auckland Islands. To estimate the number of these non-colonial bred pups, a mark–recapture survey of tagged (at colony) versus untagged pups could be carried out, with the re-visit occurring later in the breeding season (March–April) when pups have dispersed around the coast of Campbell Island, similar to the survey undertaken in 2003 (Childerhouse et al. Citation2005). The Davis Point colony is likely to have grown as a result of the population sink effect, with an initial small number of females attracting others to breed here, despite Davis Point being poor habitat for young pups. Without immigration of breeding age females from other parts of Campbell Island, it is unlikely that the colony will maintain its size in the long term. When comparing the Campbell Island NZ sea lion pup production to that of the Auckland Islands, there is an obvious opposite trend in pup production (), including no apparent large decrease seen in pup production in the 2009 season at Campbell Island (unlike Auckland Islands [Chilvers 2011]). These differences in pup production underscore the need for NZ sea lion populations and anthropogenic impacts (particularly the direct and indirect impacts of fishing) to be consistently monitored on both islands to aid effective and appropriate decision-making around the future management of this nationally critical species.
Acknowledgements
The work was funded and conducted under permit from the New Zealand Department of Conservation (DOC). Approval for all work was obtained from the DOC Animal Ethics Committee (Approval AEC199, 2 Nov 2009). We thank Henk Haazen for providing support for the exploration of the Campbell Island coastline, and the crews of both the Tiama and Evhoe for transport to and from the island. Thanks to T. Lind-Jackson for graphics help, and S. Cooper, A. Todd and two anonymous reviewers for providing helpful, critical reviews of the manuscript.
Related Research Data
References
- Augé , AA and Chilvers , BL . 2010 . Behavioural responses and attraction of New Zealand sea lions to on-land female decoys . New Zealand Journal of Zoology , 37 : 1 – 13 .
- Bailey , AM and Sorenson , JH . 1962 . Subantarctic Campbell Island . Proceedings of the Denver Museum of Natural History , 10 : 52 – 56 .
- Baker A 1999 . Unusual mortality of the New Zealand sea lion, Phocarctos hookeri, Auckland Islands, January–February 1998 . Wellington, Department of Conservation. Pp. 1–28 .
- Baker , CS , Chilvers , BL , Constantine , R , DuFresne , S , Mattlin , R , van Helden , A and Hichmough , R . 2010 . Conservation status of New Zealand Marine Mammals (suborders Cetacea and Pinnipedia), 2009 . New Zealand Journal of Marine and Freshwater Research , 44 : 1 – 15 .
- Cassini , MH and Fernández-Juricic , E . 2003 . Costs and benefits of joining South American sea lion breeding groups: testing the assumptions of a model of female breeding dispersion . Canadian Journal of Zoology , 81 : 1154 – 1160 .
- Castinel , A , Duignan , PJ , Lyons , ET , Pomroy , WE , Gibbs , N , Lopez-Villalobos , N , Chilvers , BL and Wilkinson , IS . 2007 . Epidemiology of hookworm (Uncinaria spp.) infection in New Zealand (Hooker's) sea lion (Phocarctos hookeri) pups on Enderby Island, Auckland Islands (New Zealand) during the breeding seasons from 1999/2000 to 2004/2005 . Parasitological Research , 101 : 53 – 62 .
- Castinel , A , Duignan , PJ , Pomroy , WE , López-Villalobos , N , Gibbs , NJ , Chilvers , BL and Wilkinson , IS . 2007 . A survey of neonatal mortality in New Zealand sea lions, Phocarctos hookeri at Sandy Bay Beach on Enderby Island from 1998 to 2005 . Journal of Wildlife Diseases , 43 : 461 – 474 .
- Cawthorn M 1993 . Census and population estimation of Hooker's sea lion at the Auckland Islands, December 1992–February 1993 . DOC Technical Series 2. Wellington, Department of Conservation. 34 p .
- Childerhouse , S and Gales , N . 1998 . Historical and modern distribution and abundance of the New Zealand sea lion Phocarctos hookeri . New Zealand Journal of Zoology , 25 : 1 – 16 .
- Childerhouse , S , Gibbs , N , McAlister , G , McConkey , S , McConnell , H , McNally , N and Sutherland , D . 2005 . Distribution, abundance and growth of New Zealand sea lion Phocarctos hookeri pups on Campbell Island . New Zealand Journal of Marine and Freshwater Research , 39 : 889 – 898 .
- Chilvers , BL . 2008 . New Zealand sea lions (Phocarctos hookeri) and squid trawl fisheries: bycatch problems and management options . Endangered Species Research , 5 : 193 – 204 .
- Chilvers BL 2010 . Research to assess the demographic parameters and at sea distribution of New Zealand sea lions, Auckland Islands . http://www.doc.govt.nz/conservation/marine-and-coastal/commercial-fishing/marine-conservation-services/meetings-and-project-updates/29-october-2010/ (accessed 26 July 2011)
- Chilvers BL 2011 . Research to assess the demographic parameters and at sea distribution of New Zealand sea lions, Auckland islands . http://www.doc.govt.nz/publications/conservation/marine-and-coastal/marine-conservation-services/csp-reports/assessing-the-demographic-parameters-and-at-sea-distribution-of-nz-sea-lions/ (accessed 26 July 2011)
- Chilvers , BL and Mackenzie , D . 2010 . Age and sex specific survival estimates incorporating tag loss for New Zealand sea lions, Phocarctos hookeri . Journal of Mammology , 91 : 758 – 767 .
- Chilvers , BL , Robertson , BC , Wilkinson , IS and Duignan , PJ . 2007 . Growth and survival of New Zealand sea lions, Phocarctos hookeri: birth to 3 months . Polar Biology , 30 : 459 – 469 .
- Chilvers , BL , Robertson , BC , Wilkinson , IS and Gemmell , NJ . 2005 . Harassment reduction strategies by female New Zealand sea lions, Phocarctos hookeri, during arrival on land and departure to sea . Canadian Journal of Zoology , 83 : 642 – 648 .
- Chilvers , BL and Wilkinson , IS . 2008 . Philopatry and site fidelity of New Zealand sea lions, Phocarctos hookeri . Wildlife Research , 35 : 463 – 470 .
- Chilvers , BL , Wilkinson , IS and Childerhouse , S . 2007 . New Zealand sea lion, Phocarctos hookeri, pup production—1995 to 2005 . New Zealand Journal of Marine and Freshwater Research , 41 : 205 – 213 .
- Gales , NJ and Fletcher , D . 1999 . Abundance, distribution and status of the New Zealand sea lion Phocarctos hookeri . Wildlife Research , 26 : 35 – 52 .
- Gill , BJ . 1998 . Prehistoric breeding sites of New Zealand sea lions (Phocarctos hookeri, Carnivora: Otariidae) at North Cape . Records of the Auckland Museum , 35 : 55 – 64 .
- Maloney , A , Chilvers , BL , Haley , M , Muller , CG , Roe , W and Debski , I . 2009 . Distribution, pup production and mortality of New Zealand sea lion Phocarctos hookeri on Campbell Island, 2008 . New Zealand Journal of Ecology , 33 : 97 – 105 .
- McConkey , SD , Heinrich , S , Lalas , C , McConnell , H and McNally , N . 2002 . Pattern of immigration of New Zealand sea lion, Phocarctos hookeri to Otago, New Zealand: implications for management . Australian Mammalogy , 24 : 107 – 116 .
- McConkey , SD , McConnell , H , Lalas , C , Heinrich , S , Ludmerer , A , McNally , N , Parker , E , Borofsky , C , Schimanski , K and McIntosh , G . 2002 . A northward spread in the breeding distribution of the New Zealand sea lion, Phocarctos hookeri . Australian Mammalogy , 24 : 97 – 106 .
- McNally , N , Heinrich , S and Childerhouse , S . 2001 . Distribution and breeding of New Zealand sea lions Phocarctos hookeri on Campbell Island . New Zealand Journal of Zoology , 28 : 79 – 87 .
- Moore PJ , Moffatt RD 1990 . Research and management projects on Campbell Island 1987–88. Science and Research Internal Report Series 57 . Wellington, Department of Conservation. 101 p .
- Robertson BC , Chilvers BL 2011 . The population decline of New Zealand sea lions Phocarctos hookeri: a review of possible causes . Mammal Review: http://onlinelibrary.wiley.com/doi/10.1111/j.1365-2907.2011.00186.x/pdf (accessed 26 July 2011)
- Russ R 1980 . New Zealand sea lion and fur seal census and habitat survey, Campbell Island group . In: Preliminary reports of Campbell Island expedition 1975–76. Reserves Series No.7. Wellington, Department of Lands and Survey .
- Thompson FN, Oliver MD, Abraham ER 2010. Estimation of the capture of New Zealand sea lions (Phocarctos hookeri) in trawl fisheries from 1995–96 to 2007–08. New Zealand Aquatic Environment and Biodiversity Report No. 52. http://fs.fish.govt.nz/Page.aspx?pk = 113&dk=22271 (accessed 26 July 2011)
- Wilkinson IS , Burgess J , Cawthorn MW 2003 . New Zealand sea lions and squid—managing fisheries impacts on a threatened marine mammal . In: Gales N, Hindell M, Kirkwood R eds. Marine mammals: fisheries, tourism and management issues. Melbourne, CSIRO Publishing. Pp. 192–207 .
- Wilkinson IS , Duignan PJ , Grinberg A , Chilvers BL , Robertson BC 2006 . Klebsiella pneumoniae epidemics: possible impact on New Zealand sea lion recruitment . In: Trites AW, DeMaster DP, Fritz LW, Gelatt LD, Rea LD, Wynne KM eds. Sea lions of the world. Alaska, Lowell Wakefield Fisheries Symposium. Pp. 455–471 .
- Worthy , TH . 1994 . Holocene breeding of sea lions (Phocarctos hookeri) at Delaware Bay, Nelson, South Island, New Zealand . New Zealand Sciences , 21 : 61 – 70 .