
ABSTRACT
The fossil record of the genus Carcharodon (Lamniformes: Lamnidae) dates to the late Oligocene and has a near global distribution. Today the genus is represented by a single species, the white shark Carcharodon carcharias. However, multiple extinct species are recognised in the Cenozoic including Carcharodon hastalis, Carcharodon hubbelli and Carcharodon planus comb. nov. Here we present the first occurrences of Carcharodon hubbelli and Carcharodon planus from New Zealand/Aotearoa, from the South Island/Te Waipounamu. Carcharodon hubbelli is previously reported from late Miocene deposits in the Pacific basin of North and South America, Asia, and Australia. Whereas Carcharodon planus has been reported from the Miocene of North America and Asia. One tooth from Carcharodon hubbelli is reported from the presumed late Miocene of Motunau Beach, North Canterbury, while one upper early Miocene tooth recovered from the Kakahu River, South Canterbury and an earliest Miocene tooth from the Big River mouth, Tasman District are referred to Carcharodon planus. In addition to being the first reported occurrences of these two species in New Zealand, the teeth of Carcharodon planus represent some of the earliest occurrences of the taxon as well as its southernmost occurrence.
Introduction
The extant white shark, Carcharodon carcharias (Linnaeus Citation1758), is an iconic cosmopolitan apex predator in today’s oceans. Its fossil record extends back to the Pliocene (Ehret et al. Citation2012). Other species of Carcharodon have been identified in the fossil record and there is still much to learn about their geologic ages, distributions, and relationships (Ehret et al. Citation2009, Citation2012; Kent Citation2018). The most recently described species is Carcharodon hubbelli Ehret et al. Citation2012 from late Miocene (up to 5–7 mya) Pacific deposits of North and South America, Asia, and Australia (Ehret et al. Citation2012). Carcharodon hubbelli is a transitional evolutionary stage between Carcharodon hastalis (Agassiz Citation1843) and C. carcharias (Ehret et al. Citation2012). In older literature, the genus Isurus was attributed to the species hastalis and planus. While, the generic assignment of these closely-related species remains in flux (see Purdy et al. Citation2001; Ward and Bonavia Citation2001; Ehret et al. Citation2012; Kriwet et al. Citation2015; Yun Citation2022), for the time being, we follow Ehret et al. (Citation2012) and Kent (Citation2018) by considering it most parsimonious to place these shark species in the genus Carcharodon. C. hastalis had a global distribution and occurred from the late Oligocene to the lower Pliocene (Purdy et al. Citation2001; Kent Citation2018) whereas Carcharodon planus comb. nov. is only known with certainty from the Miocene of the Pacific (Boessenecker Citation2011; Nazarkin Citation2013; Yun Citation2022). Kemp (Citation1991) reported Oligocene records of C. planus from Australia, though these records remain inconclusive because the teeth were not figured or described in any detail.
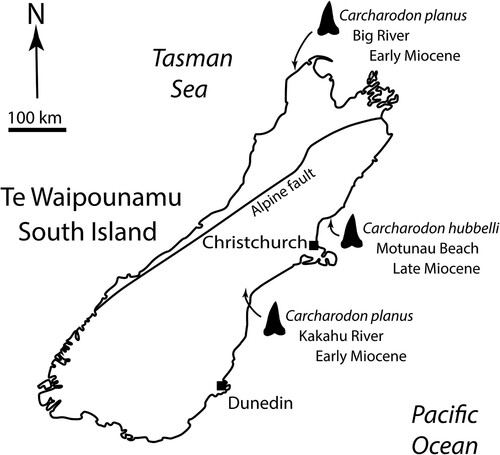
To date, two species of Carcharodon have been reported from the fossil record of New Zealand/Aotearoa: Carcharodon hastalis from the Cenozoic (King et al. Citation2009) and C. carcharias from the Pliocene-Pleistocene (Keyes Citation1972). In this paper we document the first records of two other shark species from this clade: Carcharodon planus and Carcharodon hubbelli, both from Miocene deposits in the South Island/Te Waipounamu ().
Figure 1. Localities for Carcharodon hubbelli and Carcharodon planus specimens from New Zealand.
Methods
Measurements of teeth were taken following Keyes (Citation1972), Hubbell (Citation1996), Shimada (Citation2002a) and Ehret et al. (Citation2009). Crown height (CH) is the vertical distance between a line, drawn across the lowest reaches where the tooth enameloid touches the root and the apex of the crown. Crown width (CW) was measured as the maximum transverse measurement across the crown where the crown enameloid meets the root. Crown thickness (CT) was measured in a medial position on the crown where thickness is greatest.
NMNZ = Museum of New Zealand Te Papa Tongarewa, Wellington, New Zealand.
Results
Systematic paleontology
Class Chondrichthyes Huxley Citation1880
Order Lamniformes Berg Citation1958
Family Lamnidae Müller and Henle Citation1838
Genus Carcharodon Smith in Müller and Henle Citation1838
Carcharodon hubbelli Ehret et al. Citation2012 ((A,B)).
Figure 2. Carcharodon hubbelli (NMNZ S.48965) lingual (A), labial (B); Carcharodon planus (NMNZ S.44937) lingual (C), labial (D); Carcharodon planus (NMNZ S.49347) lingual (E), labial (F). Scale bar = 1 cm.
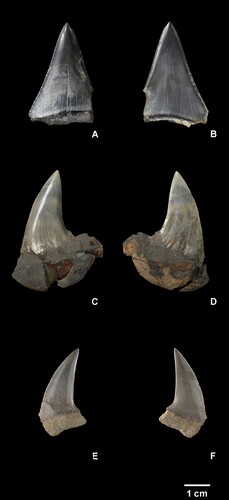
Description: NMNZ S.48965 is a large triangular tooth with weakly serrated cutting edges, corresponding to an upper first or second anterior tooth. This tooth is approximately symmetrical and labiolingually compressed. A dental band (ie. chevron or bourlette) is not present, and the base of the crown is nearly straight both labially and lingually. Only a small fragment of the root remains. Fine serrations (up to 0.3 mm) are present along the basal ¾ of the crown and are largest basally; they become finer apically and the apical ¼ of the crown has a smooth cutting edge. The serrations are irregular in size and shape, occasionally with several smaller serrations in between larger. No notches between serrations are present. NMNZ S.48965 has a crown height of 38.6 mm, a crown width of >26.5 mm and a crown thickness of 8.8 mm.
Identification: Large, triangular, labiolingually compressed and flattened teeth lacking labiolingual curvature are assignable to the genus Carcharodon. Four named species (extant and extinct) of Carcharodon are currently recognised and three are considered chronospecies: C. hastalis for unserrated teeth (extinct), C. hubbelli for weakly serrated teeth (extinct), and C. carcharias for coarsely serrated teeth (extant). NMNZ S.48965 has weak, irregular serrations lacking notches, closely resembling teeth of C. hubbelli. Serrations are lower in height and less well defined than in previously reported teeth of C. hubbelli including the holotype (Ehret et al. Citation2012) and a tooth from the Purisima Formation of California (Boessenecker Citation2011, Figure 4). NMNZ S.48965 matches the description of C. hubbelli most closely, owing to the presence of weak serrations.
Collection details: Motunau Beach, North Canterbury, −43.049985, 173.074399. Collected by Morne Wium, 6 February 2021. Proximal end of crown found protruding from a small concretion, prepared out with pneumatic airscribe by M. Wium. New Zealand Fossil Record File Database (www.fred.org.nz) fossil record number N34/f0180 ().
Geological Setting: The pholad-bored concretion of very fine sandstone that contained the tooth is referred to the Greta Formation. The fossiliferous concretions found on the wave cut platform beneath the sea cliffs near the Motunau River mouth derive from early Pleistocene (early Nukumaruan local stage) submarine canyon-head debris flow deposits that are transported, mixed and concentrated early Miocene to early Pleistocene (Otaian – Nukumaruan local stage; Aquitanian to Gelasian) fossiliferous clasts, exposed in the coastal cliffs (Lewis Citation1976; Beu and Maxwell Citation1990). The resulting issues of dating fossils found at Motunau beach are discussed by Fordyce (Citation1991). The Greta Formation represents an upper bathyl setting, though the tooth-bearing concretion may originally derive from a continental shelf setting before being transported down submarine channels by debris flow (Browne and Field Citation1985; Beu and Maxwell Citation1990).
Age: The age presented for the tooth is inferred to be upper Tongaporutuan to Kapitean (NZ local stage; upper Tortonian – Messinian, Late Miocene). NMNZ S.48965 independently supports a late Miocene age for some of the reworked concretions in the early Pleistocene debris flow deposits of the Greta Formation, which is the source for the loose beach boulder lag at Motunau Beach. Feldmann and Keyes (Citation1992) proposed that the numerous crustacean-containing concretions derived from the Greta Formation on Motunau beach were of upper middle to lower late Miocene age (Waiauan – Tongaporutuan local stage; Serravalian – Tortonian) due to the presence of Tumidocarcinus giganteus coupled with the constrains the depositional ages of debris flow clast sources presented by Lewis (Citation1976). This information together with the reported age range for C. hubbelli elsewhere suggests the most plausible age range of initial deposition of NMNZ S.48965 to be upper Tongaporutuan to Kapitean (∼5.33–8.96 Ma; Raine et al. Citation2015). See more discussion below.
Carcharodon planus (Agassiz Citation1856)
Description: NMNZ S.44937 ((C, D)) and S.49347 ((E, F)) are large teeth with distally inclined crowns. The crowns are flattened and not labiolingually curved. In NMNZ S.44937 the mesial edge is sinuous, being mesially convex apically and concave basally; in NMNZ S.49347, the mesial edge is continuously convex, and despite breakage, appears to have been slightly mesially concave basally. The distal edge in NMNZ S.44937 is sinuous, being slightly convex apically and strongly concave basally; in NMNZ S.49347 the distal edge is nearly straight. In both teeth, the apex of the crown is positioned distally to the distal crown heel. The crown is labiolingually compressed and narrow, with a flat labial face; in NMNZ S.44937 the labial face exhibits several longitudinal wrinkles along the basal 5.0 mm of the crown. The root is broken in NMNZ S.49347 and in NMNZ S.44937 it is obscured by a nodule. In NMNZ S.49347 damage precludes determination of the shape, but the root seems to have been apicobasally shallow and likely had rectangular proportions – though whether the lobes were rounded is unclear. NMNZ S.44937 has a crown height 0f 30.2 mm, a crown width of ∼22 mm and a crown thickness of 8.3 mm. NMNZ S.49347 has a crown height of 26.9 mm, a crown width of >16.3 mm and a crown thickness of 7.0 mm.
Identification: NMNZ S.44937 and S.49347 are clearly large lamnid shark teeth owing to the triangular shape, roughly rectangular root, and labiolingual compression. The strong distal inclination, strongly convex (rather than straight) to even sinuous mesial edge of the crown is shared only with Carcharodon planus. Upper lateral teeth of ancestral C. hastalis can be distally inclined, however these teeth are typically always triangular with approximately straight mesial and distal cutting edges. The specimens discussed here have a much more pronounced distal inclination than any teeth of C. hastalis seen by any of the authors. Although the roots are obscured or broken in both specimens, we feel confident in the identifications due to the shape of the crowns. Previous authors have suggested that Carcharodon planus exhibits a triangular root with rounded lobes, which is shared with other closely related species and is not itself a diagnostic characteristic (Kuga Citation1985; Karasawa Citation1989; Nazarkin Citation2013; Yun Citation2022). NMNZ S.44937 was initially identified as Carcharodon hastalis, then as Parotodus in the lab, but we note that this specimen differs considerably from Parotodus by being linguo-labially flattened, bearing sinuous mesial and distal margins, and by lacking the distinctive dental chevron (or bourlette) on the lingual side of the tooth that distinguishes all Otodontidae.
Taxonomic Note: We provisionally refer the species Cosmopolitodus planus (or alternatively Isurus planus) to the genus Carcharodon and recombine it as Carcharodon planus. Carcharodon planus is distinctive in possessing laterally recurved teeth and rounded root lobes but otherwise is quite similar to Carcharodon hastalis (Jordan and Hannibal Citation1923): broad, labiolingually flattened crown, unserrated cutting edges, and labiolingually flattened and subrectangular root. Amongst Neogene and extant species of lamnid sharks, it shares these features in common only with Carcharodon hastalis, and aside from serrations, Carcharodon hubbelli and Carcharodon carcharias as well. Similarities with Carcharodon hastalis led to initial assignment as Isurus and Cosmopolitodus when C. hastalis was formerly assigned to these genera (Purdy et al. Citation2001; Nazarkin Citation2013; Yun Citation2022). We consider that C. planus is congeneric with C. hastalis, the latter of which was assigned to Carcharodon by Ehret et al. (Citation2012). We follow Ehret et al. (Citation2012) here but note that others have assigned both to Cosmopolitodus (Yun Citation2022). Despite being common in Pacific rim Miocene marine strata, Carcharodon planus is poorly studied, and we recommend that future investigations utilise morphometric or other quantitative methods to assess the taxonomy of this species (e.g. Nyberg et al. Citation2006; Türtscher et al. Citation2021).
Referred Specimen NMNZ S.44937 ((C,D))
Collection Details: Kakahu River, 2.5 km west of Hilton, South Canterbury, −44.137334, 171.136506. Collected by Graeme M. Mason, 1972 (field number FM7), in situ from siltstone. M. Richards’ field observations (5 April 2021) suggest that the specimen was collected from the base of the riverbank cliffs on the true left bank ∼70 m upstream of the provided coordinates in the New Zealand Fossil Record File Database (www.fred.org.nz) fossil record number J38/f0002.
Geological Setting: Collected from a massive blue-grey siltstone of the Mount Harris Formation ( = Tokama Siltstone of Field and Browne Citation1986). Chemically weathered outcrops are buff yellow. The Mount Harris Formation was deposited on a wide continental shelf of probable outer shelf depth (Beu and Maxwell Citation1990) ().
Age: Inferred to be upper early Miocene (Altonian local stage; 15.9-18.7 Ma) via stratigraphic placement high in the formation by GM Mason. A fossil dolphin collected locally from the same formation, though possibly a lower part of the sequence (J38/f0113), yielded an Altonian assemblage of foraminiferans (Aguirre-Fernández Citation2013).
Referred Specimen NMNZ S.49347 ((E,F))
Collection Details: At prominent rock outcrop ∼120 m elevation on hilltop of eastern side of Big River outlet, Tai Tapu Station, Northwest Nelson area, Tasman District, −40.766079, 172.264060. Collected in situ by shepherd Leonard Mason, March 2021, donated to NMNZ by Piers Maclaren at the request of landowners Dave and Elva Harwood. Field photos by L. Mason show that the tooth root was not preserved in the rock, possibly being bioeroded during deposition. New Zealand Fossil Record File Database (www.fred.org.nz) fossil record number L25/f0019 ().
Geological Setting: Collected from the Kaipuke Siltstone, a grey indurated massive calcareous siltstone that was deposited in a mid-shelf setting in water temperatures ranging between ∼12-17°C (Aguirre-Fernández and Fordyce Citation2014).
Age: Inferred to be earliest Miocene (upper Waitakain local stage; Aquitanaian). Fossil odontocete Papahu taitapu (OU 22066; M25/f0057) was collected from the Kaipuke Siltstone at a shore platform ∼15 km northeast of NMNZ S.49347. The cetacean was stratigraphically ∼10 m upsequence from the conformable contact with the underlying Takaka Limestone (Aguirre-Fernández and Fordyce Citation2014), and was originally dated by a foraminiferal fauna sampled along strike (M25/f0059) to early Miocene (upper Waitakian – Otaian; Aquitanian – mid Burdigalian). New strontium isotope data by Marx et al. (Citationin press) from Lentipecten shells associated with OU 22066 produce a weighted mean Sr ratio of 0.708271 ± 0.000027 (2se) giving an age of 22.73 ± 0.53 Ma. This refines the lower Kaipuke Siltstone to the upper Waitakian (Aquitanian). The shark tooth locality is inferred to be of similar age due to its stratigraphic placement close to the underlying Takaka Limestone.
Discussion
We document records of two poorly known large mackerel shark species from the New Zealand fossil record. Carcharodon planus was previously known from the lower through upper Miocene rocks (Burdigalian through Tortonian) of the Pacific Rim, particularly from the west coast of the United States (Jordan and Hannibal Citation1923; Boessenecker Citation2011) and Mexico (Deméré Citation1984), Australia (Kemp Citation1991), the Korean Peninsula (Yun Citation2022), Sakhalin Island (Nazarkin Citation2013), and Japan (Kuga Citation1985; Karasawa Citation1989). The New Zealand records extend the range of this species further southwest in the Pacific basin however this is unsurprising given the Australian occurrences. At 44° S, this new record of Carcharodon planus from New Zealand is the highest latitude occurrence of this taxon in the Pacific basin, paralleling the record in the western North Pacific (47° N, Sakhalin; Nazarkin Citation2013), and further indicating a wide latitudinal range for this species. Curiously, teeth of C. planus have not been reported from the densely sampled and rich fossil beds of Peru and Chile (e.g. Chávez-Hoffmeister and Villafaña Citation2023), suggesting a circum-North Pacific and western South Pacific distribution.
NMNZ S.44937 and NMNZ S.49347 represent early Miocene records of Carcharodon planus. A similar-aged record of Carcharodon planus from the Astoria Formation of Oregon (USA) was listed (but not figured) by Welton (Citation1972); this unit dates to 17.3-16.6 Ma according to paleomagnetism (Prothero et al. Citation2001). The youngest known occurrence of Carcharodon planus appears to be the Santa Margarita Sandstone of central California (Perry Citation1993), estimated to be 10–9 Ma in age (Repenning and Tedford Citation1977). Accordingly, the species is common and conspicuous in this stratum and the Sharktooth Hill Bonebed in the Langhian Round Mountain Silt of California (Boessenecker, pers. obs.) but absent from well-sampled (albeit poorly documented) marine vertebrate assemblages from the ‘late’ Tortonian upper Monterey Formation (Barnes et al. Citation1985: Table 1) and the lower San Mateo Formation (Barnes et al. Citation1981) of southern California. Notably, teeth of Carcharodon hastalis and Isurus oxyrinchus are common in these assemblages, suggesting an extinction likely during the Tortonian, perhaps 8–10 Ma.
Carcharodon hubbelli also appears to be a Pacific basin endemic species, being reported from Peru (Muizon and DeVries Citation1985; Ehret et al. Citation2009, Citation2012), California (Boessenecker Citation2011), Japan (as Carcharodon sp.; Yabe Citation2000), and Australia (Kemp Citation1991, Fig. 19 h). The typical range of dates for specimens of this taxon from well-dated deposits is latest Miocene, approximately 5–7 Ma (Boessenecker Citation2011; Ehret et al. Citation2012; Ochoa et al. Citation2022). The New Zealand record therefore also extends the range of this species further to the southwest in the Pacific. The age of fossils from the boulder lag of concretions found on Motunau Beach, North Canterbury, is difficult to determine, however C. hubbelli can be used as a high-resolution biostratigraphic indicator due to its well constrained age range (Ehret et al. Citation2012). A latest Miocene age (Kapitean local stage; Messinian) for NMNZ S.48965 is likely as the Kapitean (5.33-7.2 Ma; Raine et al. Citation2015) overlaps nicely with the ages of overseas records. An upper Tongaporutuan age (∼7.2–8.96 Ma; Raine et al. Citation2015), at the maximum end of C. hubbelli's known age range, is possible for NMNZ S.48965 because it has the least-developed serrations yet recorded for this transitional species. Also, a Kapitean age has yet to be reported for any fossiliferous concretionary material at Motunau whilst some macrofossils previously described at the site have proposed Tongaporutuan or older ages (Lewis, Citation1976; Feldmann and Keyes Citation1992). However, macrofossils such as mollusca can be hard at times to distinguish between these two New Zealand stages (Beu & Maxwell Citation1990), so a conservative approach of late Miocene (upper Tongaporutuan – Kapitean; upper Tortonian – Messinian) is preferred. Further work is required to date the fossiliferous concretion assemblage exposed on the Motunau coastline, though temporally constrained specimens like C. hubbelli aid in refining the ages for some of the ancient debris-flows’ source stratigraphy.
Carcharodon planus seems to have been more modest in size relative to extant Carcharodon carcharias. Large teeth of C. planus from the Round Mountain Silt of California on display at CCNHM (Mace Brown Museum of Natural History, Charleston, South Carolina, USA) measure up to 31 mm in crown height, corresponding to a body length of 3.2 metres (based on tooth:body length regressions of Isurus oxyrinchus; Shimada Citation2002b) to 3.6 metres (based on tooth:body length regressions of Carcharodon carcharias; Shimada Citation2002a). Whereas Ehret et al. (Citation2009) and Ehret et al. (Citation2012) showed that C. hubbelli grew at a slower rate than modern C. carcharias but reached similar sizes. The addition of these two sharks to the New Zealand Miocene fauna demonstrates that the Cenozoic seas of New Zealand had a more diverse fauna of apex-predators than previously recorded.
Acknowledgments
We thank the collectors for generously donating their specimens to public collections; and Jean-Claude Stahl (NMNZ) for taking the photos used in this publication. We thank Dave & Nikki Stanton for granting access to the Kakahu River locality and for their hospitality, and Macgregor Willis, Dave Pease and Benjamin Richards for their assistance to M. Richards on a fieldtrip to this site. Thank you to landowners Dave & Elva Harwood for facilitating the collecting in Northwest Nelson. Thank you to Patrick Jambura and two anonymous reviewers for their helpful comments.
Disclosure statement
No potential conflict of interest was reported by the author(s).
References
- Agassiz LJR. 1843. Recherches sur les poissons fossiles. [Research of fossil fishes]. Tome III (livr. 15-16). Neuchâtel: Imprimerie de Petitpierre, p. 157–390.
- Agassiz LJR. 1856. Notice on the fossil fishes found in California by WP Blake. American Journal of Science and Arts. 21(Series. 2 ):272–275.
- Aguirre-Fernández G. 2013. Neogene dolphins from New Zealand: implications for the evolution of the family Kentriodontidae [dissertation]. Dunedin: University of Otago.
- Aguirre-Fernández G, Fordyce, RE. 2014. Papahu taitapu, gen. et sp. nov., an early Miocene stem odontocete (Cetacea) from New Zealand. Journal of Vertebrate Paleontology. 34(1):195–210. doi:10.1080/02724634.2013.799069.
- Barnes LG, Howard H, Hutchison JH, Welton BJ. 1981. The vertebrate fossils of the marine Cenozoic San Mateo Formation at Oceanside, California. In: Abbott PL, O'Dunn S, editors. Geologic investigations of the coastal plain. San Diego: San Diego Association of Geologists; p. 53–70.
- Barnes LG, Raschke RE, McLeod SA. 1985. A late Miocene marine vertebrate assemblage from Southern California. National Geographic Society Research Reports. 21:13–20.
- Berg LS. 1958. System der rezenten und fossilen Fischartigen und Fische. Berlin: Deutsche Verlag Wissenschaften.
- Beu AG, Maxwell PA. 1990. Cenozoic mollusca of New Zealand. New Zealand Geological Survey Paleontological Bulletin. 58:1–518.
- Boessenecker RW. 2011. A new marine vertebrate assemblage from the late Neogene Purisima Formation in central California, part I: fossil sharks, bony fish, birds, and implications for the age of the Purisima Formation west of the San Gregorio Fault. PalArch's Journal of Vertebrate Paleontology. 8(4):1–30.
- Browne GH, Field BD. 1985. The lithostratigraphy of late Cretaceous to early Pleistocene rocks of northern Canterbury, New Zealand. NZ Geological Survey Record. 6:1–63.
- Chávez-Hoffmeister MF, Villafaña JA. 2023. The Neogene record of cartilaginous fishes (chondrichthyes: holocephali, elasmobranchii) from northern Chile: a review and identification guide. Journal of South American Earth Sciences. 124:104230. doi:10.1016/j.jsames.2023.104230.
- Deméré TA. 1984. Paleontology of the middle Miocene Los Indios Member of the Rosarito Beach Formation, northwestern Baja California, Mexico. Pacific Section A.A.P.G. 54:47–56.
- Ehret DJ, Hubbell G, MacFadden BJ. 2009. Exceptional preservation of the white shark Carcharodon (Lamniformes, Lamnidae) from the early Pliocene of Peru. Journal of Vertebrate Paleontology. 29:1–13. doi:10.1671/039.029.0113.
- Ehret DJ, MacFadden BJ, Jones DS, DeVries TJ, Foster DA, Salas-Gismondi R. 2012. Origin of the white shark Carcharodon (Lamniformes: Lamnidae) based on recalibration of the upper Neogene Pisco Formation of Peru. Palaeontology. 55:1139–1153. doi:10.1111/j.1475-4983.2012.01201.x.
- Feldmann RM, Keyes IW. 1992. Systematic and stratigraphic review with catalogue and locality index of the Mesozoic and Cenozoic decapod Crustacea of New Zealand. New Zealand Geological Survey Record. 45:1–73.
- Field BD, Browne GH. 1986. Lithostratigraphy of Cretaceous and Tertiary rocks, southern Canterbury, New Zealand. New Zealand Geological Survey Record. 14:1–55.
- Fordyce RE. 1991. A new look at the fossil vertebrate record of New Zealand. In: Vickers-Rich P, Monaghan JM, Baird RF, Rich TH, editors. Vertebrate palaeontology of Australasia. Melbourne: Pioneer Design Studio and Monash University; p. 1191–1316.
- Hubbell G. 1996. Using tooth structure to determine the evolutionary history of the white shark. In: Kimley A, Ainley D, editors. Great white sharks: The biology of Carcharodon carcharias. San Diego, California: Academic Press; p. 9–18.
- Huxley TH. 1880. On the application of the laws of evolution to the arrangement of the Vertebrata, and more particularly of the Mammalia. Proceedings of the Zoological Society, London. 43:649–662.
- Jordan DS, Hannibal H. 1923. Fossil sharks and rays of the Pacific slope of North America. Bulletin of the Southern California Academy of Science. 22:27–63.
- Karasawa H. 1989. Late Cenozoic elasmobranchs from the Hokuriku district, central Japan. The Science Reports of the Kanazawa University. 34:1–57.
- Kemp NR. 1991. Chondrichthyans in the Cretaceous and Tertiary of Australia. In: Vickers-Rich P, Monaghan JM, Baird RF, Rich TH, editors. Vertebrate palaeontology of Australasia. Melbourne: Pioneer Design Studio and Monash University; p. 497–568.
- Kent BW. 2018. The cartilaginous fishes (chimaeras, sharks and rays) of the Calvert Cliffs, Maryland, USA. In: Godfrey SJ, editor. The geology and vertebrate paleontology of Calvert Cliffs, Maryland: Smithsonian contributions to paleobiology, vol. 100: p. 45–156.
- Keyes IW. 1972. New records of the Elasmobranch C. megalodon (Agassiz) and a review of the genus Carcharodonin the New Zealand fossil record. New Zealand Journal of Geology and Geophysics. 15(2):228–242. doi:10.1080/00288306.1972.10421956.
- King CM, Roberts CD, Bell BD, Fordyce RE, Nicoll RS, Worthy TH, Paulin CD, Hitchmough RA, Keyes IW, Baker AN, et al. 2009. Phylum chordata: lancelets, fishes, amphibians, reptiles, birds, mammals. In: Gordon DP, editor. The New Zealand inventory of biodiversity. 1. Kingdom Animalia. Christchurch: Canterbury University Press, p. 431–527.
- Kriwet J, Mewis H, Hampe O. 2015. A partial skeleton of a new lamniform mackerel shark from the Miocene of Europe. Acta Paleontologica Polonica. 60:857–875.
- Kuga N. 1985. Revision of Neogene mackerel shark of genus Isurus from Japan. Memoirs of the faculty of science, Kyoto university. Series of Geology and Mineralogy. 51(1-2):1–20.
- Lewis DW. 1976. Subaqueous debris flows of Early Pleistocene age at Motunau, North Canterbury, New Zealand. New Zealand Journal of Geology and Geophysics. 19(5):535–567. doi:10.1080/00288306.1976.10426308.
- Linnaeus C. 1758. Systema naturae, Tenth Edition. Stockholm.: Larentii Salvii.
- Marx FG, Coste A, Richards MD, Palin JM, Fordyce RE. in press. Strontium isotopes reveal a globally unique assemblage of early miocene baleen whales. Journal of the Royal Society of New Zealand.
- Muizon Cd, DeVries TJ. 1985. Geology and paleontology of late Cenozoic marine deposits in the Sacaco area (Peru). Geologische Rundschau. 74:547–563. doi:10.1007/BF01821211.
- Müller J, Henle FGJ. 1838. On the generic characters of cartilaginous fishes. Magazine of Natural History. 2:88–91.
- Nazarkin MV. 2013. Hooked mako Isurus planus (agassiz, 1856) from the Miocene of Sakhalin. Zoosystematica Rossica. 22:311–314. doi:10.31610/zsr/2013.22.2.311.
- Nyberg KG, Ciampaglio CN, Wray GA. 2006. Tracing the ancestry of the great white shark, Carcharodon carcharias, using morphometric analyses of fossil teeth. Journal of Vertebrate Paleontology. 26:806–814. doi:10.1671/0272-4634(2006)26[806:TTAOTG]2.0.CO;2.
- Ochoa D, DeVries TJ, Quispe K, Barbosa-Espitia A, Salas-Gismondi R, Foster DA, Gonzales R, Revillon S, Berrospi R, Pairazamán L, et al. 2022. Age and provenance of the Mio-Pleistocene sediments from the Sacaco area, Peruvian continental margin. Journal of South American Earth Sciences. 116:103799. doi:10.1016/j.jsames.2022.103799.
- Perry FA. 1993. Fossil sharks and rays of the southern Santa Cruz Mountains, California. Santa Cruz, California: Santa Cruz Museum Association; p. 1–48.
- Prothero DR, Bitboul CZ, Moore GW, Moore EJ. 2001. Magnetic stratigraphy of the lower and middle Miocene Astoria Formation, Lincoln county, Oregon. Pacific Section SEPM Special Publication. 91:272–283.
- Purdy RW, Schneider VP, Applegate SP, McLellan JH, Meyer RL, Slaughter BH. 2001. The neogene sharks, rays, and bony fishes from Lee Creek Mine, Aurora, North Carolina. Smithsonian Contributions to Paleobiology. 90:71–202. doi:10.5479/si.00810266.90.71.
- Raine JI, Beu AG, Boyes AF, Campbell HJ, Cooper RA, Crampton JS, Crundwell MP, Hollis CJ, Morgans HEG. 2015. Revised calibration of the New Zealand geological timescale: NZGT2015/1. GNS Science Report. 2012/39:1–53.
- Repenning CA, Tedford RH. 1977. Otarioid seals of the Neogene. US Geological Survey Professional Paper. 992:1–87.
- Shimada K. 2002a. The relationship between the tooth size and total body length in the white shark, Carcharodon carcharias (Lamniformes: Lamnidae). Journal of Fossil Research. 35:28–33.
- Shimada K. 2002b. The relationship between the tooth size and total body length in the shortfin mako, Isurus oxyrinchus (Lamniformes: Lamnidae). Journal of Fossil Research. 35:6–9.
- Türtscher J, López-Romero FA, Jambura PL, Kindlimann R, Ward DJ, Kriwet J. 2021. Evolution, diversity, and disparity of the tiger shark lineage Galeocerdo in deep time. Paleobiology. 47:574–590. doi:10.1017/pab.2021.6.
- Ward DJ, Bonavia CG. 2001. Additions to, and a review of, the Miocene shark and ray fauna of Malta. Central Mediterranean Naturalist. 3:131–146.
- Welton BJ. 1972. Fossil sharks of Oregon. The Ore Bin. 34(10):161–172.
- Yabe H. 2000. Teeth of an extinct great white shark, Carcharodon sp., from the Neogene Senhata Formation, Miura Group, Chiba prefecture, Japan. Tertiary Research. 20:95–105.
- Yun C. 2022. A tooth of the extinct lamnid shark, Cosmopolitodus planus comb nov. (Chondrichthyes: Elasmobranchii) from the Miocene of Pohang City, South Korea. Acta Paleontologica Romaniae. 18:9–16.