Abstract
In this paper we argue that moral behaviour is largely situation‐specific. Genetic make‐up, neurobiological factors, attachment security and rearing experiences have only limited influence on individual differences in moral performance. Moral behaviour does not develop in a linear and cumulative fashion and individual morality is not stable across time and situations. To illustrate our position we present two studies on children’s willingness to donate their money to a charity (UNICEF) as a prime example of pro‐social behaviour. In two samples of seven‐year‐old children we found no evidence for a role of attachment, temperament or parenting. Using a twin design we did not find any evidence for a genetic component either. The most striking finding in both studies was that very few children were inclined to donate any money after viewing a promotional UNICEF film about children suffering from poverty. Only after gentle probing by an experimenter were most children willing to donate some of their money. The situation appeared to be a much more powerful determinant of donating behaviour than any other factor. These findings are discussed in the broader context of evidence for situational canalisation of moral behaviour. We conclude that moral competence may be a universal human characteristic, but that it takes a situation with specific demand‐characteristics to translate this competence into actual prosocial performance.
Introduction
Most parents like to believe that infants and young children are innocent human beings and that morality develops in a cumulative fashion so that with increasing age children would act more empathically and prosocially. In the science of morality we have for a long time been attached to the idea that individuals steadily progress through a series of stages toward an ever more sophisticated moral stance and that stable differences exist in the degree to which individuals commit themselves to empathic or prosocial acts (Piaget, Citation1932; Kohlberg, Citation1984). It requires a major acrobatic feat, however, to combine this optimistic model of morality with the indifferent, immoral and anti‐social behaviour that most adults seem capable of when the situation triggers or even only leaves room for such behaviour. From Auschwitz (Sagi‐Schwartz et al., Citation2003) to Abu Ghraib (Zimbardo, Citation2007) recent history is filled with disturbing examples of the banality of evil (Arendt, Citation1963/2006). But history also shows almost universal empathic concern with victims of man‐made or natural disasters, such as the 2004 Indian Ocean earthquake or tsunami, stimulating millions of people around the globe to donate an estimated seven billion dollars for repairs and help for the survivors. Situational canalisation (to use the famous concept of the biologist C. H. Waddington [Citation1942]) of moral behaviour might also create ‘the banality of altruism’.
Noble newborns
Human infants seem to be endowed with an inborn capacity to be empathic to distress and to favour morally good individuals over morally bad persons. The empathic baby is an appealing idea. In a pioneering study on empathic distress, Sagi and Hoffman (Citation1976) exposed 58 one‐day‐old infants to a newborn cry, a synthetic cry or silence. Infants exposed to a real cry cried significantly more often than those exposed to a synthetic cry of the same intensity or to silence. Although mimicry may be a viable alternative explanation, the authors suggest that this selective cry response in newborns may provide some evidence for an inborn empathic distress reaction.
In a groundbreaking study Hamlin et al. (Citation2007) showed that 6‐ and 10‐month‐old infants are already able to evaluate an individual on basis of their hostile or helpful actions in computer‐animated events. Astonishingly, most preverbal infants prefer an individual who helps another to one who hinders another, prefer a helping individual to a neutral individual and prefer a neutral individual to a hindering individual. The authors argue that the capacity of very young children to derive moral evaluations from simple actions may serve as the foundation for moral thought and action and they suggest that its early emergence supports the view that morality is inborn.
The nature and nurture of donating
Generous genes?
Altruism and prosocial behaviour have been speculated to be evolutionary‐based universal competences of human beings and of a large variety of avian and mammalian species as well. From these competences the actual altruistic performance might follow, as well as individual and situational differences in altruistic behaviour. Evolutionary roots of altruism and prosocial behaviour have been discussed ever since Darwin’s (Citation1871/1982) observation that human beings might have become unusually prosocial because in‐group solidarity created elevated chances of survival in deadly struggles with out‐groups and predators (see also Krebs, Citation2008). Conflict would be ‘altruism’s midwife’ and ‘parochial altruism’ the rule (Bowles, Citation2008). On the level of individual organisms and genes, the inclusive fitness theory (Hamilton, Citation1964a, Citation1964b) also explains altruistic behaviour to relatives if it facilitates the reproduction of one’s (shared) genes into the next generations. For example, inclusive fitness nicely explains parental altruism (though not unlimited) to offspring (Trivers, Citation1974).
De Waal (Citation2008) argued that the basic ability for empathic concern may have developed in the context of parental care long before the human species evolved. He refers to the evolutionary‐based attachment behaviour of offspring (see Van IJzendoorn et al. [Citation2009] for infant chimpanzees’ human‐like attachments) and the probability that avian or mammalian parents who were sensitive to their offspring’s needs likely out‐reproduced those who remained indifferent. Recently, Hrdy (Citation2009) extended the attachment model and proposed a new hypothesis based on ‘cooperative breeding’. She notes that it takes around 13 million calories to rear a human from birth to nutritional independence. This is far more than a foraging mother could provide on her own and help from her mate and group members other than the parents, so‐called alloparents, appeared essential. Cooperative breeding was necessary for survival in an environment with high parent and child mortality and selection would favour those individuals best equipped to monitor the mental states of others and to contribute to cooperative breeding. Prosocial behaviour, empathic concern and sharing of scarce resources would be rooted in the basic need for cooperative child rearing.
Twin studies
Even if we assume an evolutionary basis for altruism and empathic concern, the question remains whether individual differences in actual moral performance are associated with differences in genetic make‐up or with specific dispositional or personality characteristics. To address the role of genetic factors in the explanation of individual differences in altruistic and prosocial behaviour, studies with monozygotic and dizygotic twins have been conducted. Simply put, if correlations of moral behaviour for monozygotic twins (100% genetically similar) are substantially higher than the correlations for dizygotic twins (on average 50% genetically similar) there is some evidence for a genetic basis. One of the first twin studies on altruism included 573 adult twin pairs who were asked to complete the Self‐Report Altruism Scale requiring respondents to report the frequency with which they had engaged in 20 specific moral behaviours such as ‘I have donated blood’ (Rushton et al., Citation1986). More than 50% of the variance in altruism was found to be genetically based, whereas an almost equal percentage was explained by the unique environment component including measurement error. A tiny 2% appeared to be due to common environmental factors that make individuals within a family similar to each other.
Only two twin studies have been published on children’s prosocial behaviour, in particular empathic helping behaviour and concern for another person’s pain and distress. Volbrecht et al. (Citation2007) observed more than 200 twin pairs in the second year of life reacting to their primary caregiver who pretended to have pinched her finger in a clipboard and feigned pain for a brief period. Helping behaviour and empathic concern to distress did not show any genetic influence and both shared and unique environment explained the variation in these prosocial behaviours. In the second study, Knafo et al. (Citation2008) observed more than 400 twin pairs longitudinally from 14 to 36 months of age in a similar empathic distress procedure, but also including a stranger simulating pain. At 14 months, no genetic effect was found on a composite measure of empathy, based on mother and experimenter simulations. By 24 months, a genetic effect accounting for about a quarter of the variance emerged and this effect remained stable toward 36 months. With age, genetic effects on prosocial behaviour and empathic concern appeared to increase and shared environmental effects to decrease, which, according to the authors, might explain the considerable influence of genetics on altruism found in the adult twin study (Rushton et al., Citation1986).
Prosocial parents?
Some 25 years ago, Hoffman (Citation1984) suggested a role for parents in promoting or hampering the development of prosocial behaviour. He contended that if parents create a warm, sensitive atmosphere and, at the same time, consistently discipline child behaviour that is damaging to others, they pave the way for feelings of empathy in their children. Non‐empathic, authoritarian control combined with frequent threats and love withdrawal would, on the other hand, lead to compulsive compliance and a lack of moral internalisation (Richters & Waters, Citation1992; Van IJzendoorn, Citation1997).
Sensitive parents who respond promptly and adequately to the distress and anxiety of their children stimulate a secure bond and at the same time model the use of empathy and altruism (Spiecker, Citation1991; De Waal, Citation2008; Hrdy, Citation2009). Following this line of reasoning, we have hypothesised that children with an insecure attachment relationship with their primary caregiver may be less inclined to act prosocially than children with secure relationships (Van IJzendoorn, Citation1997). Mikulincer and Shaver (Citation2008) present some experimental evidence for the role of attachment in adult helping behaviour. They primed young adults with security‐enhancing figures such as their best friend and observed more compassion with a woman in distress and more willingness to take over her disagreeable tasks in the primed subjects as opposed to participants in control conditions.
Temperamental fearfulness may moderate the effect of parenting on moral development (Kagan & Lamb, Citation1990). Kochanska et al. (Citation1995, Citation2007) found evidence for diverse pathways to moral internalisation and prosocial behaviour of children with different temperaments. For temperamentally fearful children, gentle maternal discipline predicted optimal moral internalisation. For fearless children, however, security of attachment was associated with internalisation and gentle discipline appeared less important. Attachment security and temperament emerged as complementary factors in the development of morality. The moderating influences of temperament on the impact of parenting appeared to be restricted to assessments in the second year of life predicting preschoolers’ prosocial behaviour (Kochanska et al., Citation2007). It should be noted that temperament may also play a role in the process of translating empathic distress into prosocial action. Fearful or emotionally reactive children might be more upset by another individual’s pain and distress and then incapacitated to take action.
Considering the available evidence, the following optimistic hypothetical model has led our Leiden research program on the early roots of morality in the past two decades (see Figure ). The model assumes that the interplay between parenting and genetics or temperament is crucial for our understanding of children’s altruistic behaviour like donating. Sensitive parents facilitate secure attachments in their children, which makes the youngsters more willing to share scarce resources with individuals in need. At the same time parental sensitivity is the first and foremost display of altruism to a dependent human being and a role model for the developing child. Sensitivity is empathic concern for the children’s needs in everyday life, which might be mirrored by the children’s empathy for others. However, individual differences in prosocial performance might be related to genetic differences since empathic concern seems to become more genetically determined with growing age. Also, genetically‐based temperamental differences in emotion regulation may hamper or enable individuals to translate their empathic distress into prosocial actions or may make them less or more susceptible to parenting influences (Belsky et al., Citation2007).
Figure 1 Attachment‐based developmental model of moral behaviour
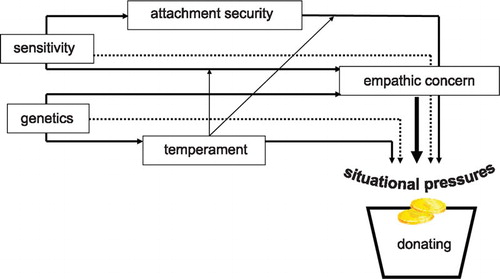
Situational morality
Noble newborns revisited
If the idea of the empathic baby were valid, somewhere during the early years this moral capacity must be pruned quite drastically or, alternatively, kept in check by the social context. Lamb and Zakhireh (Citation1997), for example, documented a remarkable lack of empathic behaviour in 45 toddlers (18 months of age) in the natural setting of a day care centre. These researchers observed 345 distress incidents during 20 hours of video‐recording in the (average quality) centres and registered how children in the group responded to this distress. Only unambiguous instances of prosocial responses to distress were coded, such as offering a toy or patting a crying child, but not simple approaches or concerned looks. Of the 345 incidents a meagre 11 incidents were followed by a prosocial action from one of the peers. Prosocial response (whether or not the child had ever responded prosocially) was not related to age or gender. Of course, this study does not prove that young children would not have the disposition to be empathic; it demonstrates that in the day care setting they display indifferent behaviour. The children may have the competence to act empathically but their prosocial performance does not emerge in the specific situation.
In contrast to a trait‐like and cumulative stage‐wise interpretation of morality, unselfish or immoral behaviour might better be considered as shaped by the demand‐characteristics of the specific situation (Hartshorne & May, Citation1928/1932; Zimbardo, Citation2007). Prosocial behaviour may be situation‐specific rather than a disposition or personality trait. Exploring the limits of the genetic, dispositional and socialisation views on moral development, we now turn to the influence of situational pressures on moral and immoral behaviour.
Situational immorality
The hypothetical model of morality emphasises the potential roles of genes and parenting. Some disturbing empirical evidence suggests, however, that moral behaviour may be mainly situation‐specific. Several widely known but little digested social‐psychological experiments vividly demonstrate this possibility. A powerful example of situational preponderance over dispositional differences was the Zimbardo et al. (Citation2000) Stanford prison experiment in which psychologically healthy Stanford students were induced to play the roles of prisoners or guards. After a few days the students identified themselves so completely with their roles as torturous guards that the experiment had to be aborted to avoid real casualties.
Abu Ghraib
How close to reality this experiment is became poignantly evident from the Abu Ghraib prison’s abuses and tortures of Iraqi prisoners by regular American (male and female) soldiers. In 2004, a detailed report on the American military prison located near Baghdad was written by the American Major General Taguba who described numerous instances of ‘sadistic, blatant, and wanton criminal abuses’ of Iraqis by American soldiers. There was convincing evidence of systematic abuse, including ‘detailed witness statements and the discovery of extremely graphic photographic evidence’, sexual‐sadistic pictures taken by American soldiers during the abuse (see also Zimbardo [Citation2007, p. 324 ff] for a detailed psychological analysis of Abu Ghraib). In his book on the Lucifer effect Zimbardo (Citation2007) thus argues for more emphasis on situational determinants of prosocial and antisocial behaviour, in addition to the currently prevailing emphasis on dispositional factors.
Milgram experiments
In the (in)famous Milgram (Citation1974) experiments most healthy and balanced adult participants were easily stimulated into the inhumane act of electrocuting another human being only on the authority of the experimenter and the demand‐characteristics of the scientific setting. Situational pressures seemed to override any genetic, dispositional or personality differences in determining moral choices (Blass, Citation2000).
How is this succumbing to situational pressures related to moral reasoning? It might be argued that in the Milgram experiments subjects are tested for their empathic responses (to the experimenter or to the victim) instead of a morally justified reaction. In contrast, we suggest that Milgram examined moral behaviour in vivo, for which empathic concern as well as moral reasoning might be important but not decisive. Whatever moral competence the subjects in the Milgram experiments might have had, most of them performed immorally. Kohlberg was acutely aware of the importance of these experiments for his theory and investigated their association with his moral stages. Kohlberg (Citation1984, pp. 546–547) argued that participants’ obedience to the authority of an experimenter in the original Milgram paradigm was not completely context‐dependent but also predictable by their responses to the Moral Judgment Interview (MJI). He reported MJI data on 27 students who completed the Milgram experiment, either quitting against the will of the experimenter (n = 8) or not (n = 19). At first glance the data seem to support his hypothesis that more advanced moral reasoning was associated with quitting the torturing of the victim, as five out of eight quitters were classified at the highest moral level (level 4) and only one student on this level did not quit. However, half of the students reasoning on the lowest level (level 3) did quit as well.
We tested the association between moral level and quitting using the exact Fisher Chi‐square test and we found a significant overall effect, χ2 (2, N = 27) = 13.69, p < .01, with significant adjusted standardised residuals for level 3/4 (sr = −3.5: fewer subjects at this level than expected did quit) and for level 4 (sr = 3.3: more subjects at this level than expected did quit), but the lowest level did not support the prediction of more subjects at that level complying with the experimenter. Only the post‐hoc re‐coding of the Substage A and Substage B moral reasoning (Kohlberg, Citation1984, p. 546) led to confirmation of the prediction that Substage B was indeed associated with quitting and Substage A with non‐quitting, χ2 (1, N = 27) = 14.26, p < .01. We derived as an effect size the correlation between Substage and quitting, which amounted to r = .73. This figure is implausibly high considering the reliability of both the MJI classification and the outcome of the Milgram paradigm (see below for a discussion of maximum correlations). Replication of this inflated finding is as urgent as it is impossible, because the original Milgram experiments have become ethically unacceptable (but see Meeus and Raaijmakers [Citation1995] for a cleverly designed alternative administrative obedience paradigm and Burger [Citation2009] for an ethically less controversial procedure that demonstrate the validity of Milgram’s findings even half a century later).
Milgram with children
To our surprise we discovered that in the 1970s Milgram’s original test of obedience had been replicated with children. Although serious ethical doubts might be raised about the conduct of the highly intrusive experiment with adults, let alone with children, the virtual neglect of the findings from this stunning study cannot be legitimised by such ethical concerns about the experiment itself. Shanab and Yahya (Citation1977) tested 192 Jordanian children (6–8, 10–12 and 14–16 years) in two kinds of punishment instructions. Half of the children received instructions similar to the original Milgram experiment, namely to administer electric shocks to learners (who were of course so‐called confederates or assistants of the experimenter) each time the latter made a mistake in a paired‐associate learning task and to increase the shock level with each additional mistake. The other half of the children were given a free choice of delivering or not delivering shocks each time the learner made a mistake.
Pressed by the experimenter almost three‐quarters of the children continued to deliver shocks to maximum voltage. Only 16% of the children with a free choice went that far. The situational pressures were much more important than other factors: obedience rates were not associated with age or with gender. The authors rated the emotional responses of the subjects during the Milgram experiment, recording tense behaviour like loud nervous laughter, lip biting and trembling and if they showed 11 or more of these emotional behaviours they were considered to display intense tension. Disturbingly, the number of children in the intense tension category significantly decreased with age. Of those who expressed intense tension, 44%, 25%, and 16% were in the age groups 6–8, 10–12 and 14–16, respectively.
The benevolent brain?
The idea of dispositional morality may seem to be supported by recent brain research (Hauser, Citation2007). Situational morality seems to run counter to growing evidence that empathic and altruistic behaviour and attitudes can be located in specific areas of the brain, showing that individual differences in empathy are associated with individual differences in blood oxygenation level dependent (BOLD) signals registered by functional magnetic resonance imaging (fMRI) during an empathy or altruism task. Blood oxygenation level dependent signals are based on the elevated need for energy, and thus for oxygen, in those areas of the brain that become more active because of some stimulus or task.
Startlingly high correlations have been found between BOLD responses in the scanner and behavioural or attitudinal measures of individual differences in empathic concern or altruism. For example, Singer et al. (Citation2004) reported that pain‐related empathic concern for a loved one was visible in higher activation of the insula and the anterior cingulate cortex, not only when participants (16 couples) experienced physical pain themselves but also when they witnessed similar pain induction to their partner. Most surprisingly the BOLD signals in these areas were strongly correlated with scores on empathy questionnaires completed outside the scanner. The BOLD signals in the insula, for example, correlated r = .72 with the Mehrabian Balanced Emotional Empathy Scale (Singer et al., Citation2004).
Moll et al. (Citation2006) observed charitable donation behaviour in the scanner. Nineteen participants were asked to choose to endorse or oppose societal causes by donating or refraining from donating to real charitable organisations, with missions such as promoting children’s rights or fighting the death penalty. Participants would receive $128 if they only made egoistic choices. The amount of money decreased depending on how often they made altruistic choices. Again, the results were striking. Not only the mesolimbic reward system seemed to be engaged in donations (suggesting the self‐serving nature of donating behaviour, ‘feeling good’) but also the orbitofrontal cortex that is implicated in attachments (as speculated by Schore, Citation2000). Most importantly, anterior parts of the prefrontal cortex (where higher cognitive and executive functions are located) were more active in costly choices between altruistic and selfish interests. It would not be difficult to reconstruct a neuroscientific defence of our hypothetical model of morality from these findings.
The correlation of the BOLD signals in the prefrontal area during the task and self‐reported engagement in real‐life voluntary activities amounted to r = .87 (Moll et al., Citation2006). Individual differences in prefrontal cortex activity seem to completely determine voluntarism. But correlations as high as .72 or .87 for any brain‐behaviour association are too strong to be true. In fact, this is a case of over‐determination. Analysing 55 reports in social neuroscience using fMRI, Vul et al. (Citation2009) showed that many published correlations are much higher than can be expected given the limited reliability of both fMRI and personality measures. Blood oxygenation level dependent signals in social neuroscience show test‐retest reliabilities of on average about .70, and for most empathy‐related or altruism questionnaires an optimistic reliability estimate would be .80. The maximum correlation between two measures is the square root of the product of the reliability figures of the two measures, in this case .74 (Nunnally, Citation1970; Vul et al., Citation2009).
The logical implication is that the Singer et al. (Citation2004) study almost hit the ceiling. Clearly, Moll et al. (Citation2006) even rose above this ceiling and explained more variance than in fact available if we would take errors of measurement seriously. As was the case with many other studies in the Vul et al. (Citation2009) review, a non‐independence problem might be the reason for the implausible correlation. The scanner registers BOLD signals in so‐called voxels (volumetric pixels) that have a size of 1∼125 cubic mm. Per scan 40.000∼500.000 voxels are usually assessed and the crucial question is how to select from this enormous number of signals the data to be included in the statistical analyses. Vul et al. (Citation2009) show that in many cases researchers first select the voxels (maybe in some specific regions of interest) with highly significant associations with the outcome variable, combine these signals into one BOLD score and correlate this score with the same outcome variable, for example a measure of empathy or voluntarism, with highly inflated correlations as a result.
Whatever the exact cause of the strong correlations, they are implausible on psychometric grounds. The scanner is a powerful method to gain insight into brain‐behaviour connections, but we should be cautious of some of the overstatements emerging in the field of the social neuroscience of morality. The brain is a relay station between mind and body but not a homunculus that is the ultimate causative agent of our moral behaviour. If we see elevated brain activity in moral choice settings this is not necessarily causative for pro‐ or antisocial behaviour; it may as well be a consequence or a concomitant of such behaviour. Across the life‐span brain activity can be affected by environmental input influencing epigenetic programming (McGowan et al., Citation2009) and recovery from extreme circumstances has been shown to permanently change electrical activity in the brain (Marshall et al., Citation2008). Brain activity may therefore be the explanandum as well as the explanans, that is, the end product as well as the starting point of moral behaviour.
Empathy and donating in the Leiden Attachment Research Program
Our focus is donating behaviour as an example of altruistic, prosocial behaviour. More specifically, we address the question of what is most important in predicting children’s donation of hard‐won money to the cause of a well‐known charity (UNICEF): genetic differences, differences in disposition (attachment and temperament) or situational differences. We define altruism in a somewhat loose way as the individual’s inclination to spend resources without the expectation of personal gain. Donating to a charity is an example of prosocial behaviour that realises the altruistic inclination and empathic concern might be considered a motivational prerequisite of altruistic behaviour like donating. Donating behaviour is, of course, just one example of moral behaviour and various types of moral behaviour should be observed in a range of situations. Here we only want to examine whether donating behaviour is primarily determined by disposition, genes or situation. If it turns out to be situationally determined it may be considered falsifying evidence (in the Popperian sense) for the dispositional or genetic viewpoint. It would also show that it is problematic to consider moral behaviour to be stable across situations as minor changes in settings might canalise behaviour in different directions (see Hartshorne & May, Citation1928/1932).
The Leiden Longitudinal Empathy Study (LLES)
We observed the development of empathy from infancy to middle childhood and examined its relation to donating. Eighty‐seven middle‐class Dutch mothers with a firstborn female infant were seen at 15, 18 and 89 months of age. Our sample included only girls, as a counterweight to Kohlberg’s longitudinal sample on only male subjects, and—more pragmatically—to enhance the power of statistical analyses. Prosocial and empathic behaviour are characteristics usually ascribed more and earlier to girls than to boys (Eisenberg & Fabes, Citation1998).
The development of empathy
Simulations of pain and sadness were used at 18 and 24 months (see Van der Mark et al., Citation2002) and again at 89 months (Pannebakker, Citation2007). The experimenter pretended to hurt her finger or her knee for about 20–30 seconds. After about five minutes the mother was asked to pretend to hurt her knee or her finger. Mothers were also asked to cough as if they choked, several minutes after the other simulations. Mothers did not look at the child during these simulations, in order to avoid extra stimulation of the child’s reactions. Simulations were done during ongoing activities, to avoid awareness of the pretend actions in the child. The simulations were coded for their credibility; differences in credibility were not associated with children’s behaviour. We adapted the empathy coding systems used by Zahn‐Waxler et al. (Citation1992) and by Hastings and colleagues (Citation2000) (for older children). A global score for empathy was assigned on the basis of shown empathic concern and prosocial behaviour, which was unrelated to the persuasiveness of the mothers’ simulations.
A repeated measures analysis of variance with victim (mother or experimenter) and time (18, 24 and 89 months) as within‐subjects factors for empathic concern showed a significant effect of victim (F[1, 86] = 81.39, p < .01), time (F[2, 85] = 24.72, p < .01) and the interaction between victim and time (F[2, 85] = 28.58, p < .01). However, empathic concern did not develop in a linearly progressive way. Empathic concern towards the mother decreased from 24 to 89 months, after an increase from 18 to 24 months. Empathic concern towards the experimenter decreased from 18 to 89 months. Contrary to the developmental model of Hoffman (Citation2000) and others, growing older did not mean becoming more empathic. In line with evolutionary psychology, children showed more empathic concern towards their mother than towards an unrelated person from 18 to 89 months. Similar results were found in studies showing that individuals, especially women, are generally more inclined to help relatives than to help non‐relatives (Eagly & Crowley, Citation1986; Burnstein et al., Citation1994). The associations across time were significant but disappointingly low and they did not point to strong early childhood determination of a developmental pathway for empathic concern. A lack of associations between empathy to the mother and to the experimenter also throws doubt on the concept of general empathy independent of the target. In fact, the (biological) relationship to or familiarity with the target might affect empathic concern rather substantially.
Situational donating?
High‐cost donating behaviour was measured by the amount of money (the number of €0.20 coins) the child donated in response to a videotaped call for donation to UNICEF. After an hour of performing various tasks, the children received 10 coins of 0.20 for their cooperation, in the absence of their mother. They were then left alone and shown a two‐minute UNICEF promotional film of a child in a ‘resource‐limited’, developing country. During the last fragments of the promotion the voice‐over asked to donate money; a moneybox had been positioned next to the video screen. The moneybox was filled with several euros in order to enhance credibility. After one minute the experimenter came back into the room and asked in a standardised way if the child would want to donate any money.
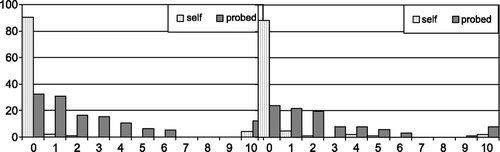
Left alone, only a very small minority (11%) of the children donated any money. Being gently probed by the experimenter, the percentage of children who donated steeply increased to 76% (see Figure ). The within‐subject effect of time was significant, F(1, 86) = 70.93, p < .01 and the effect size was very strong, η 2 = .45, also because errors of measurement were minimised. Donating behaviour was not associated with empathic concern, attachment security, temperament or any other predictor in this study. We tortured the data (sensu Sue Goldberg) but failed to find predictors of donating, except for the situation. Donating money to UNICEF or keeping the money for personal use appeared to be an easy, selfish choice for many children, but a choice that was also readily influenced by a change in the setting.
Figure 2 Proportions of the children donating one, two, three etc. coins after the UNICEF promotional film (self) and after the experimenter’s prompt (probed) in the two Leiden studies: LLES (left) and LLTS (right)
The Leiden Longitudinal Twin Study (LLTS)
We used the same donating paradigm in a study of 91, seven‐year‐old, same‐sex twin children (30 monozygotic, 61 dizygotic; 43 male, 48 female pairs) to examine whether individual differences in donating after being probed by the experimenter are genetically based or whether at this age the (shared and unique) environment also affects donating behaviour. As in the previous study, the percentage of children that donated without being probed was too small to allow genetic modelling.
Is donating heritable?
The correlation for the MZ twin pairs was r = .45 (p = .01) and the correlation for the DZ twin pairs was r = .44 (p < .01). These correlations are almost equal and therefore do not leave much room for genetic influences. To formally test the contributions of genetic, shared and unique environmental differences, we conducted a behavioural genetic analysis. In behavioural genetic analyses, the similarity of pairs of twins is broken down into similarity due to additive genetic factors (A) and similarity due to shared environmental experiences (C); dissimilarity is accounted for by unique or non‐shared environmental influences and measurement error (E). The analysis was performed with the structural equation modelling program Mx (Neale et al., Citation1999).
The CE model fitted the data as good as the ACE model and should be preferred to the ACE model, as the difference in fit was not significant and the CE model is more parsimonious. The AE model fitted the data not significantly worse than the ACE model either, although the difference in fit was almost significant (p = .07). Further restriction of the model with both A and C fixed to zero (the E model) reduced the fit significantly. These results indicate the CE model as the most plausible model (see Table ). In this model, 45% of the variance in donating was explained by shared environmental influences (C, 95% confidence interval 25–72%) and 55% of the variance by unique environment and measurement error (E, 95% confidence interval 42–75%). Of course, the absence of a main genetic effect is not incompatible with indirect effects through gene by environment.
Table 1. Behaviour genetic modelling of donating behaviour
Our twin study does not confirm previous findings of a genetic component affecting altruistic attitudes and behaviours in an adult twin sample (Rushton et al., Citation1986) and in one of the two studies with twin children (Knafo et al., Citation2008). In fact, our study supports Volbrecht et al.’s (Citation2007) finding of no genetic component in explaining differences in empathic concern. It should be noted that the genetic component in the Knafo et al. (Citation2008) study amounted to only about 25% of explained variance, which might go undetected in our sample because of lack of statistical power.
Donating behaviour, attachment and temperament
In our search for predictors of donating we conducted a multivariate analysis on donating behaviour of the first‐born children of each twin pair with attachment and temperament as factors and maternal educational level and child age as covariates. The model was significant, F (5, 85) = 2.88, p = .02, η 2 = .15. But only child age (p < .01) and maternal educational level (p = .02) were significantly associated with the amount of money that the children donated after being probed; older children and children from higher socioeconomic status (SES) families donated more. The significant shared environment component we found in the behavioural genetic analysis might be due to maternal educational level as a proxy for SES of the family. Kagan (Citation2009) argued that SES is still the strongest determinant or correlate of almost any facet of child development, despite the neuroscientific revolution in psychology and the educational sciences.
Again, we did not find main or interaction effects for attachment and temperament. The absence of associations between attachment and donating converges with the surprising outcome of a study on donating involving maltreated children, with a past of bad parenting and insecure attachments. Koenig et al. (Citation2004) used a donating task comparing physically maltreated, neglected and typical five‐year‐old children from low socioeconomic backgrounds. The children were asked to donate (some of) their earnings during the experiments to a poor family whose house had burnt down. Donation was anonymous and out of sight of the experimenter. About a quarter of the children donated some money, but children without maltreatment experiences were not inclined to donate more to the suffering family than maltreated children, although the rearing experiences of the two groups differed dramatically.
Situational morality: donating behaviour across two settings
Examining the number of coins donated after the promotional film and then after the experimenter’s probe, we conducted a repeated measures analysis. The within‐subject effect of time was significant, F(1, 90) = 52.49, p < .01, and again the effect size was large, η 2 = .37. Only a very small minority (9%) were inclined to donate any money spontaneously. After being urged by an experimenter to donate, the percentage of children who donated some or all of their money rather steeply increased to 67% (see Figure ). These findings are remarkably similar to those of the LLES with female participants only. In the twin study girls donated somewhat more than boys after viewing the promotional film, t (48.52) = 2.08, p = .04 (unequal variances) but the gender difference disappeared after the experimenter’s prompt, t(89) = 0.58, p = .56.
We must conclude that seven‐year‐olds are not inclined to spontaneously donate their money to a charity and only after an adult’s probe are ready to share part of their money with less privileged children elsewhere in the world. It should be noted that we examined only donating behaviour of seven‐year‐old children and we should be careful about making generalisations to other age cohorts. In their study on 3–16‐year‐old children, Grunberg et al. (Citation1985) found that donating to a charity reaches a dip around seven years. Other studies discussed in this paper, however, document situational morality in a wide range of ages, so our findings seem not restricted to this specific age. Our behaviour‐genetic finding of a large shared environment component is not incompatible with the large influence of the situation on donating behaviour. The effect of the situation is observed at the group level, whereas the influence of the shared environment is located at the level of individual differences in donating, on top of the group effect.
Converging evidence for situational morality
The situational nature of prosocial donating behaviour seems to mirror the situation‐specificity of antisocial behaviour in the Milgram (Citation1974) and Zimbardo (Citation2007) experiments. The crucial question is whether situations can also be created to channel participants’ behaviour into a prosocial direction. Fortunately, some independent experimental studies suggest that this is possible. Freeman et al. (Citation2009) tested the influence of witnessing or reading about moral exemplars on donating to a charity. Students watching a video clip that documented moral excellence or reading a story about an extraordinary moral act or person donated more money to a charity that promoted goals somewhat antithetic to their political views. The experiments were based on Haidt’s (Citation2007) theory of moral elevation leading to more intense moral emotions and moral acts. Witnessing an act of moral excellence would stimulate thoughts, emotions and a motivational state that encourages people to show more empathic concern and caring behaviour.
Van Rompay et al. (Citation2009) showed that small situational changes such as installing a security camera implying the presence of an audience resulted in similar enhancement of prosocial behaviour in a student sample. Students provided more help in collecting and sorting a pile of questionnaires accidentally fallen on the floor in the presence of a camera, although the camera did not affect the participants’ reported donations to charitable organisations. The dispositional trait of need for social approval also explained some variance in helping behaviour, on top of the situational characteristics.
Kallgren et al. (Citation2000) tested the influence of the presence of a confederate picking up a crumpled fast‐food bag from the floor of a parking garage. Participants were visitors to a public urban hospital who were returning to their cars. When participants reached their cars, they encountered handbills attached to their windshield. Throwing the handbills on the floor was the observed outcome. The simple witnessing of the confederate picking up a piece of litter decreased littering behaviour from 43% to a mere 9.3%. With a simple manipulation of the situation (setting an example, focusing on the prosocial norm) prosocial behaviour was strongly stimulated.
In our study the situational difference between watching a promotional video clip alone and with the probe of an experimenter explained 37% of the variance in donating behaviour, whereas only half as much (19%) of the variance in donating after the experimenter’s probe was explained by background variables (age, maternal educational level). Dispositional (attachment) and constitutional (temperament) differences seemed irrelevant. Increasing the situational pressures might affect donating behaviour until the point is reached that almost all children donate almost all their money to the charity when social pressures are maximised, mirroring the maltreating behaviour of most participants in the Milgram and Zimbardo experiments. What type of manipulations with children would be acceptable from an ethical point of view, even when they are meant to provoke prosocial behaviour, is food for thought. Modelling or encouraging the child to donate money to a charity may constitute one end of the continuum, with making starvation of a videotaped undernourished infant the responsibility of the child being the other end. Depending on the age of the child and debriefing possibilities experimental manipulations should move along this continuum in order not to violate the ethical balance between means and aims.
Educational implications: situational morality in the just community?
What kind of educational intervention is compatible with situational morality? If the demand characteristics of the situation have most impact on prosocial and antisocial behaviour, the logical implication is to monitor and change the environment in which children are growing up and are being educated. De Waal (Citation1996) conducted an amazing educational experiment mixing easy‐going, tolerant but also physically larger stump‐tail macaques with smaller rhesus monkeys who are known for their high level of aggression. The groups were kept together day and night for five months. Because of their dominant physical status the macaques acted as tutors and models of peaceful interactions for the excitable rhesus monkeys. After some time the rhesus monkeys began to interact in a similar fashion as the friendly macaques, not only to their dominant co‐habitants but also to their own peers and the effects remained visible after the co‐habiting experiment was ended. With clever control conditions alternative interpretations were excluded, such as mere mimicry of any macaque behaviour by rhesus monkeys. It should be noted that five months of 24/7 co‐habitation is equal to more than two years in a human child’s life.
The just‐community approach to moral education, originated by Kohlberg (Power, Citation1979; Kohlberg, Citation1985; Oser et al., Citation2008) aimed at the immersion of students in an environment imbued with just and fair role models, rules and interactions and with concrete behavioural norms (to be on time, to abstain from fighting). Most importantly, the peers in the just community embodied and sanctioned the sociomoral norms that canalised individual students’ behaviour. Kohlberg (Citation1985) stressed that “the good” as altruism is cultivated by a “sense of community”, by a feeling of group cohesion and solidarity’ (p. 84). Essential ingredients were: the community meeting for democratic decision‐making; the discipline committee for confronting individuals with their misbehaviour and punishing and forgiving them; and moral dilemma discussions in the classroom to enhance the level of moral reasoning (Oser et al., Citation2008).
The just‐community approach was a short‐term success elevating the moral judgement level of the students, but as Kohlberg stated: ‘While the intervention operation was a success, the patient died’ (Kohlberg, Citation1985, p. 80). One year after the conclusion of the experiment not a single teacher continued to do moral discussions. Practising moral behaviour in the just community teaches children how to be moral, but generalisation across time and settings has not yet been proved in a randomised control trial. In fact, it is the just‐community setting that incorporates morality for some time, not the individual’s brain or mind, a premier illustration of situational morality. The successful failure of this just‐community experiment may have at least two educational implications. First, implementation of a just community should be prolonged and sustained across several years to effectively change the participants’ moral behaviour. De Waal’s intervention with macaque and rhesus monkeys, which appeared to lead to persistently positive changes in the rhesus monkeys’ interactions within their own group after closure of the experiment, lasted about two years on a human time‐scale. Second, part of moral education may consist of making students aware of the power of situations in determining their moral choices in experimental and natural settings. Teaching students the experimental evidence of situational morality, such as the Stanford prison experiment or the Milgram experiments, may lead to self‐defeating prophecies and create opportunities to make moral choices that deviate from the situational pressures to act in a specific (immoral) way. Discussing with students some real examples of situational immorality, such as Auschwitz or Abu Ghraib, as well as examples of canalisation of altruistic behaviour, such as donating after the tsunami, may add to their understanding of the determinants of their own moral behaviour.
Conclusion: one step forward, two steps back…
In sum, we have argued that moral behaviour is largely situation‐specific, much more so than we, in our previous work, have assumed. Genetic make‐up, neurobiological factors, attachment security and rearing experiences have only limited influence on individual differences in moral performance. Here we deliberately focused on observed moral behaviour, as most self‐report measures are doomed to be as invalid as they are reliable (Van IJzendoorn, Citation1984). Whether cognitive moral judgements play a role in children’s decisions to act morally, for example to donate to a charity, remains to be seen. The association between cognitive moral judgements and moral actions is complex (Blasi, Citation1980), may go both ways or may not exist at all. With growing age children might gain more empathic feelings and they also might respond to moral dilemmas in an ever more morally justified way, but we have argued here that moral behaviour might still be mainly situationally determined. And it is moral behaviour that really counts as the ancient proverb suggests: the proof of the pudding is in the eating. In general, a rather large gap has been detected between thought and action, attitudes and behaviour, in various domains of functioning (Deutscher, Citation1973). It would be gratifying, but hard to believe, if the same were not true for moral reasoning and action.
We may have to go beyond Kohlberg’s (Citation1984) cognitive‐developmental view on morality by going back to ‘…Hartshorne and May (who) found, that adolescents (and by extension adults) are not divided into groups, the conscientiously honest and the dishonest. They find instead that situational factors independent of conscience appear to be the determinants of honest behavior’ (p. 3). As much as moral competence is a universal human characteristic, it takes a situation with specific demand‐characteristics to translate this competence into actual prosocial performance.
Acknowledgements
We are grateful to Renske Gilissen and Mariëlle Beijersbergen for their contributions to data‐collection and coding. We also thank the parents and children who generously donated their time to our study, as well as the students who assisted in various research phases. Marinus H. van IJzendoorn and Marian J. Bakermans‐Kranenburg were supported by awards from the Netherlands Organization for Scientific Research (MHvIJ: SPINOZA prize; MJBK: VIDI grant no. 452‐04‐306). We thank the Editor and the reviewers of JME for their constructive comments on an earlier version of this paper.
Notes
This is the 22nd Lawrence Kohlberg Memorial Lecture presented at the 35th Annual Conference of the Association for Moral Education, University of Utrecht, The Netherlands, 3 July 2009.
Related Research Data
References
- Arendt , H. 1963/2006 . Eichmann in Jerusalem: a report on the banality of evil , New York : Penguin Books .
- Belsky , J. , Bakermans‐Kranenburg , M. J. and Van IJzendoorn , M. H. 2007 . For better and for worse: differential susceptibility to environmental influences . Current Directions in Psychological Science , 16 (6) : 300 – 304 .
- Blasi , A. 1980 . Bridging moral cognition and moral action: a critical review of the literature . Psychological Bulletin , 88 (1) : 1 – 45 .
- Blass , T. 2000 . Obedience to authority: current perspectives on the Milgram paradigm , Mahwah , NJ : Lawrence Erlbaum .
- Bowles , S. 2008 . Being human: conflict: altruism’s midwife . Nature , 456 (7220) : 326 – 327 .
- Burger , J. M. 2009 . Replicating Milgram: would people still obey today? . American Psychologist , 64 (1) : 1 – 11 .
- Burnstein , E. , Crandall , C. and Kitayama , S. 1994 . Some neo‐Darwinian decision rules for altruism: weighing cues for inclusive fitness as a function of the biological importance of the decision . Journal of Personality and Social Psychology , 67 (5) : 773 – 789 .
- Darwin , C. 1871/1982 . The descent of man, and selection in relation to sex , Princeton , NJ : Princeton University Press .
- Deutscher , I. 1973 . What we say/what we do: sentiments and acts , Glenview , IL : Scott Foresman .
- De Waal , F. B. M. 1996 . Good natured: the origins of right and wrong in humans and other animals , Cambridge , MA : Harvard University Press .
- De Waal , F. B. M. 2008 . Putting the altruism back into altruism: the evolution of empathy . Annual Review of Psychology , 59 : 279 – 300 .
- Eagly , A. H. and Crowley , M. 1986 . Gender and helping‐behavior: a meta‐analytic review of the social psychological literature . Psychological Bulletin , 100 (3) : 283 – 308 .
- Eisenberg , N. and Fabes , R. A. 1998 . “ Prosocial development ” . In Handbook of child psychology. Vol. 3: Social, emotional and personality development , (5th edn) , Edited by: Damon , W. and Eisenberg , N. 701 – 778 . New York : Wiley .
- Freeman , D. , Aquino , K. and McFerran , B. 2009 . Overcoming beneficiary race as an impediment to charitable donations: social dominance orientation, the experience of moral elevation, and donation behavior . Personality and Social Psychology Bulletin , 35 (1) : 72 – 84 .
- Grunberg , N. E. , Maycock , V. A. and Anthony , B. J. 1985 . Material altruism in children . Basic and Applied Social Psychology , 6 (1) : 1 – 11 .
- Haidt , J. 2007 . The new synthesis in moral psychology . Science , 316 (5827) : 998 – 1002 .
- Hamilton , W. D. 1964a . Genetical evolution of social behaviour I . Journal of Theoretical Biology , 7 (1) : 1 – 16 .
- Hamilton , W. D. 1964b . Genetical evolution of social behaviour II . Journal of Theoretical Biology , 7 (1) : 17 – 52 .
- Hamlin , J. K. , Wynn , K. and Bloom , P. 2007 . Social evaluation by preverbal infants . Nature , 450 (7169) : 557 – 559 .
- Hartshorne , H. and May , M. 1928‐1932 . Studies in the nature of character , New York : Macmillian .
- Hastings , P. D. , Zahn‐Waxler , C. , Robinson , J. , Usher , B. and Bridges , D. 2000 . The development of concern for others in children with behavior problems . Developmental Psychology , 36 (5) : 531 – 546 .
- Hauser , M. D. 2007 . Moral minds: how nature designed our universal sense of right and wrong , Cambridge , MA : Harvard University Press .
- Hoffman , M. L. 1984 . “ Interaction of affect and cognition in empathy ” . In Emotions, cognitions and behaviour , Edited by: Izard , C. E. , Kagan , J. and Zajone , R. B. 103 – 131 . Cambridge : Cambridge University Press .
- Hoffman , M. L. 2000 . Empathy and moral development: implications for caring and justice , Cambridge : Cambridge University Press .
- Hrdy , S. B. 2009 . Mothers and others: the evolutionary origins of mutual understanding , Cambridge , MA : Harvard University Press .
- Kagan , J. 2009 . Two is better than one . Perspectives on Psychological Science , 4 (1) : 22 – 23 .
- Kagan , J. and Lamb , S. 1990 . The emergence of morality in young children , Chicago , IL : University of Chicago Press .
- Kallgren , C. A. , Reno , R. R. and Cialdini , R. B. 2000 . A focus theory of normative conduct: when norms do and do not affect behavior . Personality and Social Psychology Bulletin , 26 (8) : 1002 – 1012 .
- Knafo , A. , Zahn‐Waxler , C. , Van Hulle , C. , Robinson , J. L. and Rhee , S. H. 2008 . The developmental origins of a disposition toward empathy: genetic and environmental contributions . Emotion , 8 (6) : 737 – 752 .
- Kochanska , G. , Aksan , N. and Joy , M. E. 2007 . Children’s fearfulness as a moderator of parenting in early socialization: two longitudinal studies . Developmental Psychology , 43 (1) : 222 – 237 .
- Kochanska , G. , Aksan , N. and Koenig , A. L. 1995 . A longitudinal study of the roots of preschoolers’ conscience: committed compliance and emerging internalization . Child Development , 66 (6) : 1752 – 1769 .
- Koenig , A. L. , Cicchetti , D. and Rogosch , F. A. 2004 . Moral development: the association between maltreatment and young children’s prosocial behaviors and moral transgressions . Social Development , 13 (1) : 87 – 106 .
- Kohlberg , L. 1984 . Essays on moral development. Volume 2: The psychology of moral development , San Franscisco : Harper & Row .
- Kohlberg , L. 1985 . “ Resolving moral conflicts within the just community ” . In Moral dilemmas , Edited by: Harding , C. 71 – 98 . Chicago , IL : Precedent Publishing .
- Krebs , D. L. 2008 . Morality: an evolutionary account . Perspectives on Psychological Science , 3 (3) : 149 – 172 .
- Lamb , S. and Zakhireh , B. 1997 . Toddlers’ attention to the distress of peers in a daycare setting . Early Education & Development , 8 (2) : 105 – 118 .
- Marshall , P. J. , Reeb , B. C. , Fox , N. A. , Nelson , C. A. and Zeanah , C. H. 2008 . Effects of early intervention on EEG power and coherence in previously institutionalized children in Romania . Development and Psychopathology , 20 (3) : 861 – 880 .
- McGowan , P.O. , Sasaki , A. , D’Alessio , A. , Dymov , S. , Labonté , B. and Szyf , M. 2009 . Epigenetic regulation of the glucocorticoid receptor in human brain associates with childhood abuse . Nature Neuroscience , 12 (3) : 342 – 348 .
- Meeus , W. H. J. and Raaijmakers , Q. A. W. 1995 . Obedience in modern society: the Utrecht studies . Journal of Social Issues , 51 (3) : 155 – 175 .
- Mikulincer , M. and Shaver , P. R. 2008 . Can’t buy me love: an attachment perspective on social support and money as psychological buffers . Psychological Inquiry , 19 (3–4) : 167 – 173 .
- Milgram , S. 1974 . Obedience to authority: an experimental view , London : Tavistock .
- Moll , J. , Krueger , F. , Zahn , R. , Pardini , M. , de Oliveira‐Souzat , R. and Grafman , J. 2006 . Human fronto‐mesolimbic networks guide decisions about charitable donation . Proceedings of the National Academy of Sciences of the United States of America , 103 (42) : 15623 – 15628 .
- Neale , M. C. , Boker , S. M. , Xie , G. and Maes , H. H. 1999 . Mx: statistical modeling , (5th edn) , Richmond : Medical College of Virginia, Department of Psychiatry .
- Nunnally , J. C. 1970 . Introduction to psychological measurement , New York : McGraw‐Hill .
- Oser , F. K. , Althof , W. and Higgins‐D’Allessandro , A. 2008 . The just community approach to moral education: system change or individual change? . Journal of Moral Education , 37 (3) : 395 – 415 .
- Pannebakker , F. D. 2007 . Morality from infancy to middle childhood , Leiden University . Unpublished doctoral thesis
- Piaget , J. 1932 . The moral judgment of the child , London : Kegan Paul .
- Power , F. C. 1979 . The moral atmosphere of a just community high school: a four‐year longitudinal study , Harvard Graduate School of Education . Unpublished doctoral dissertation
- Richters , J. E. and Waters , E. 1992 . “ Attachment and socialization: the positive side of social influence ” . In Social influences and behavior , Edited by: Lewis , M. and Feinman , S. 185 – 214 . New York : Plenum Press .
- Rushton , J. P. , Littlefield , C. H. and Lumsden , C. J. 1986 . Gene culture coevolution of complex social behavior: human altruism and mate choice . Proceedings of the National Academy of Sciences of the United States of America , 83 (19) : 7340 – 7343 .
- Sagi , A. and Hoffman , M. L. 1976 . Empathic distress in newborn . Developmental Psychology , 12 (2) : 175 – 176 .
- Sagi‐Schwartz , A. , Van IJzendoorn , M. H. , Grossmann , K. E. , Joels , T. , Grossmann , K. Scharf , M. 2003 . Attachment and traumatic stress in female holocaust child survivors and their daughters . American Journal of Psychiatry , 160 (6) : 1086 – 1092 .
- Schore , A. N. 2000 . Attachment and the regulation of the right brain . Attachment & Human Development , 2 (1) : 23 – 47 .
- Shanab , M. E. and Yahya , K. A. 1977 . Behavioral study of obedience in children . Journal of Personality and Social Psychology , 35 (7) : 530 – 536 .
- Singer , T. , Seymour , B. , O’Doherty , J. , Kaube , H. , Dolan , R. J. and Frith , C. D. 2004 . Empathy for pain involves the affective but not sensory components of pain . Science , 303 (5661) : 1157 – 1162 .
- Spiecker , B. 1991 . Emoties en morele opvoeding [Emotions and moral education] , Amsterdam : Boom .
- Trivers , R. L. 1974 . Parent‐offspring conflict . American Zoologist , 14 (1) : 249 – 264 .
- Van der Mark , I. L. , Van IJzendoorn , M. H. and Bakermans‐Kranenburg , M. J. 2002 . Development of empathy in girls during the second year of life: associations with parenting, attachment and temperament . Social Development , 11 (4) : 451 – 468 .
- Van IJzendoorn , M. H. 1984 . Answers without questions: a note on response style in questionnaires . Perceptual and Motor Skills , 59 (3) : 827 – 831 .
- Van IJzendoorn , M. H. 1997 . Attachment, emergent morality and aggression: toward a developmental socioemotional model of antisocial behaviour . International Journal of Behavioral Development , 21 (4) : 703 – 727 .
- Van IJzendoorn , M. H. , Bard , K. A. , Bakermans‐Kranenburg , M. J. and Ivan , K. 2009 . Enhancement of attachment and cognitive development of young nursery‐reared chimpanzees in responsive versus standard care . Developmental Psychobiology , 51 (2) : 173 – 185 .
- Van Rompay , T. J. L. , Vonk , D. J. and Fransen , M. L. 2009 . The eye of the camera: effects of security cameras on prosocial behavior . Environment and Behavior , 41 (1) : 60 – 74 .
- Volbrecht , M. M. , Lemery‐Chalfant , K. , Aksan , N. , Zahn‐Waxler , C. and Goldsmith , H. H. 2007 . Examining the familial link between positive affect and empathy development in the second year . Journal of Genetic Psychology , 168 (2) : 105 – 129 .
- Vul , E. , Harris , C. , Winkielman , P. and Pashler , H. 2009 . Puzzlingly high correlations in fMRI studies of emotion, personality and social cognition . Perspectives on Psychological Science , 4 (3) : 274 – 290 .
- Waddington , C. H. 1942 . Canalization of development and the inheritance of acquired characters . Nature , 150 : 563 – 565 .
- Zahn‐Waxler , C. , Radke‐Yarrow , M. , Wagner , E. and Chapman , M. 1992 . Development of concern for others . Developmental Psychology , 28 (1) : 126 – 136 .
- Zimbardo , P. 2007 . The Lucifer effect: how good people turn evil , New York : Random House .
- Zimbardo , P. , Maslach , C. and Haney , C. 2000 . “ Reflections on the Stanford prison experiment: genesis, transformations, consequences ” . In Obedience to authority: current perspectives on the Milgram paradigm , Edited by: Blass , T. 193 – 237 . Mahwah , NJ : Lawrence Erlbaum .