Abstract
Heat inactivation curves were constructed for Newcastle disease virus strain Herts 33/56 in artificially infected meat homogenate at 60°C, 65°C, 70°C, 74°C and 80°C. For the four higher temperatures the time taken to reduce the infectivity by 90% (1 log10) at the specified temperature (D t ) were estimated as: D 65=120 sec, D 70=82 sec, D 74=40 sec and D 80 29=sec.
Résumé
Inactivation par la chaleur du virus de la maladie de Newcastle (souche Herts 33/56) dans des broyats de viande de poulets infectés artificiellement
Les courbes d'inactivation par la chaleur à 60°C, 65°C, 70°C, 74°C et 80°C, de la souche Herts 33/56 du virus de la maladie de Newcastle, ont été construites à partir de broyats de viande infectés artificiellement. Pour les quatre températures les plus élevées, les temps nécessaires pour réduire l'infectiosité de 90% (1 log10), à la température spécifiée (D t ), ont été estimés comme ce qui suit: D 65 120 secs, D 70 82 secs, D 74 40 secs et D 80 29 secs.
Zusammenfassung
Hitzeinaktivierung des Virus der Newcastle-Krankheit (Stamm Herts 33/56) in künstlich infiziertem Hühnerfleischhomogenat
Für den Virusstamm Herts 33/56 der Newcastle-Krankheit wurden in künstlich infiziertem Hühnerfleischhomogenat bei 60°C, 65°C, 70°C, 74°C und 80°C Hitzeinaktivierungskurven erstellt. Für die vier höheren Temperaturen wurde die benötigte Zeit zur Reduzierung der Infektiösität um 90 % (1 log10) bei einer bestimmten Temperatur (D t ) geschätzt auf: D 65 120 Sekunden, D 70 82 Sekunden, D 74 40 Sekunden und D 80 29 Sekunden.
Resumen
Inactivación mediante calor de virus de la enfermedad de Newcastle (cepa Herts 33/56) en un homogenizado artificial de carne de pollo infectadaSe construyeron las curvas de inactivación por calor del virus de la enfermedad de Newcastle cepa Herts 33/56 en un homogenizado artificial de carne infectada a 60oC, 65oC, 70oC, 74oC y 80oC. Para las 4 temperaturas más altas, el tiempo necesario para reducir la infectividad en un 90% (1 log10) a la temperatura especificada (D t ) fue estimado como: D 65 120 secs, D 70 82 secs, D 74 40 secs and D 80 29 secs.
Introduction
In the current virus taxonomy Newcastle disease virus (NDV), or avian paramyxovirus type 1, is classified, with the other avian paramyxoviruses, in the genus Avulavirus, subfamily Paramyxovirinae, family Paramyxoviridae, order Mononegavirales (Lamb et al., Citation2000; Mayo, Citation2002); that is, it has a negative-sense, single-stranded, filamentous RNA genome and a glycoprotein/lipid membrane.
Newcastle disease and highly pathogenic avian influenza are the only two poultry diseases considered sufficiently serious to be placed in list A of the Office International des Epizooties (Citation2000) and, as such, justifiable trade restrictions may be put in place to prevent the introduction of NDV by poultry products to countries free of Newcastle disease. NDV, like most enveloped viruses, is considered heat labile, and trade in poultry products is considered to represent minimum risk, even from areas where Newcastle disease is endemic, if these products have been subjected to processing that involves heating or cooking. However, there is very little published information on the heat inactivation of NDV, especially in meat and other poultry products, and rarely have temperatures and times been specified. The OIE code for international trade (Office International des Epizooties, Citation2003) merely states “the meat products have been processed to ensure the destruction of the ND virus”, without specification. Where work has been published on heat inactivation of NDV this has usually involved batch processing (i.e. showing infectivity cannot be detected after a specified time at a specified temperature), infective amnio-allantoic (AAF) fluid and at temperatures not commonly used in food processing, and is thus unhelpful in assessing the inactivation of NDV in poultry products. The exception was a study of the inactivation of NDV in liquid whole egg by Gough (Citation1973), where an inactivation curve was constructed.
In the present study we have examined the inactivation of NDV in homogenized meat preparations at 60°C, 65°C, 70°C, 74°C and 80°C, have constructed heat inactivation curves and estimate the D t value for the four higher temperatures.
Materials and Methods
Virus
The virus strain used in this study was the virulent international challenge virus Herts 33/56 (Allan et al., Citation1978). The virus was grown and titrated in specific pathogen free (SPF) 9-day-old to 10-day-old embryonated fowls’ eggs.
Medium
The medium used to assess the heat inactivation of the virus was homogenized chicken muscle/skin to which infective allantoic fluid was added. To achieve this, muscle from SPF chickens with 15% w/w added skin and fat, which was considered to more closely reflect commercial meat products than muscle alone, was homogenized as a 50% w/v suspension in normal saline at pH 7.2 (containing antibiotics) with sufficient infective allantoic fluid added to give a final titre of about 107 mean egg infectious doses (EID50)/0.1 ml supernatant after clarification by centrifugation at 3000×g for 10 min. Sufficient infective homogenate was prepared for all tests and stored in aliquots of 10 ml at −70°C until the heat inactivation experiments were carried out.
Experimental procedure
Sufficient aliquots for each test were removed from refrigeration and allowed to thaw to room temperature (22°C). Glass ‘bijou’ bottles (7 ml capacity, 5 cm high, with external diameter 2 cm internal diameter 1.7 cm at the bottom tapering after 2 cm to 1.5 cm, below the aluminium screw cap with rubber insert, at 3.5 cm) sufficient for the number of samples to be taken were placed in a laminated wire ‘bijou’ rack in a water bath at the specified test temperature, so that each bottle was immersed in the water to a minimum depth of 2 cm (i.e. well above the level of the added suspension), one bottle was placed directly into an ice bath. After 10 min, 1 ml thawed homogenate was placed in each of the bottles. The rack containing the bottles was agitated continuously by hand for the shorter times and for 10 sec in every 20 sec for the longer periods. At the times specified in one bottle was removed and placed directly into the ice bath and then held at 4°C until the titrations in SPF eggs were performed, which was immediately after the longest treatment.
Table 1. Results of heat inactivation of Newcastle disease virus in meat homogenate
The heat-treated samples were titrated by preparing 10-fold dilutions over the range 100 to 10−6 in phosphate-buffered saline and inoculating each of five 9-day-old to 10-day-old embryonated SPF fowls eggs with 0.1 ml each dilution by the allantoic route. All eggs as they died and all survivors after 5 days were tested for the presence of haemagglutinin activity. Titres of virus in 0.1 ml inoculated material, expressed as EID50, were calculated by the Spearman–Karber method.
For comparative purposes heat inactivation curves for infective AAF were constructed at 65°C and 80°C following the same experimental procedure.
Interpretation of results
Heat inactivation curves at each of the five temperatures were prepared by plotting log10 EID50 of virus against treatment time. Where no virus was detected points were given the value −0.5 log10 EID50; that is, assuming all eggs would have been positive at a 10-fold concentration of the sample. D t values (the time taken to reduce the infectivity by 90% [1 log10] at the specified temperature) were deduced from the slope of straight line curves of ‘best fit’ plotted by eye; where curves were bi-phasic, the D t value was calculated from the gradient of the second (shallowest) part.
Results
The results obtained are presented in and graphically in –.
Figure 1. Heat inactivation of Newcastle disease virus in meat homogenate at 60°C.
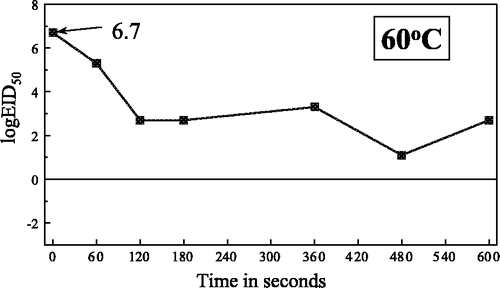
Figure 2. Heat inactivation of Newcastle disease virus in meat homogenate at 65°C.
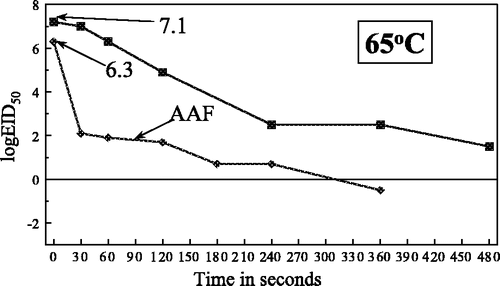
Figure 3. Heat inactivation of Newcastle disease virus in meat homogenate at 70°C.
Figure 4. Heat inactivation of Newcastle disease virus in meat homogenate at 74°C.
Figure 5. Heat inactivation of Newcastle disease virus in meat homogenate at 80°C.
Considerable problems were encountered during the heat inactivation experiments due to a tendency for the homogenized meat material to clump when heated and even solidify at higher temperatures. To some extent this could be overcome by manual agitation during heat inactivation, but it may account for some of the unexpected, out of line, results obtained.
The estimated D t values for meat homogenate at the specified temperatures were: at 65°C, 120 sec; at 70°C, 82 sec; at 74°C, 40 sec and at 80°C, 29 sec.
Discussion
Estimation of the heat inactivation of viruses in a complex medium such as meat poses considerable problems, both in terms of the practicable experimentation and in the interpretation of the data obtained. The ‘cooking’ effect on the consistency of the meat homogenate caused some technical problems in this study, but the most erratic results were obtained at 60°C, the lowest temperature used (). Whether or not the ‘cooking’ effect has any adverse effect on the overall validity of the results obtained, or their extrapolation to commercial processing of poultry products, is unclear.
The heat inactivation curves were biphasic as expected ( –) although at the lower temperatures this was less marked, which is in keeping with the observation by Picken (1964) for NDV that the heat inactivation curves tend to become single phase as the heat stress declines. Comparisons of the curves obtained for meat homogenate and infective AAF at 65°C and 80°C indicated that in AAF the slope of the initial phase was considerably steeper and more precipitous than in meat, to below the detection level at 80°C. At 65°C the slopes of the second phase were remarkably similar for both AAF and meat homogenate. It was assumed that the different results obtained with this virus in the two media was a result of the greater concentration of protein and other organic matter in the meat homogenate giving a protective effect, especially from the initial heat stress.
Estimations of D t values also pose something of a problem, especially for bi-phasic or multi-phasic curves. Square root transformation and linear regression techniques can be applied to the data (MAF NZ, Citation1999), but ultimately the ‘best fit by eye’ method we have used is likely to be a reasonable estimate, especially in view of the probable experimental and biological variability. The D t values derived in this report are based on the second phase of the curve and are therefore likely to represent an extremely demanding test of heat resistance. However, pragmatically, these are the values most likely to be considered desirable by agencies involved in the importation of processed chicken meat. There should be a linear relationship between log D t values and temperature (Russell et al., Citation1992) and this allows extrapolation and estimation of the probable D t values at temperatures other than those examined in the present study ().
Figure 6. Logarithmic plot of time taken to reduce the infectivity of NDV in meat homogenate by 90% (1 log10) at the specified temperatures (Dt values).
There have been remarkably few comparable studies on NDV reported in the literature. Gough (Citation1973) studied the inactivation of NDV in liquid whole egg, and from the data published in that study Alexander & Chettle (Citation1998) deduced that the mean time taken to bring about reduction of 1 log10 EID50 NDV strain Beaudette C in liquid whole eggs was 38 sec at a temperature of 64.4°C (D 64.4=38 sec). This is considerably less than the D 65 of 120 sec for Herts 33/56 in meat homogenate determined in the present study.
We conclude that while NDV may be regarded as a heat labile virus, showing considerably less heat resistance than infectious bursal disease virus, for example (Alexander & Chettle Citation1998), its survival during heat stress will be dependent on the medium involved and this will be an important factor in assessing the risk of introduction of NDV in processed meat products. In the absence of other studies, regulatory authorities setting levels for temperature and time for imported processed poultry products may find the data presented here useful, but should interpret the results obtained in a laboratory situation with care when applying to commercial processing.
Acknowledgments
This work was funded by the Commonwealth of Australia under contract No. FT0513 with the Veterinary Laboratories Agency, and the authors are indebted to the Australian authorities for their permission to publish the work.
Related Research Data
References
- Alexander , DJ and Chettle , NJ . 1998 . Heat inactivation of serotype 1 infectious bursal disease virus . Avian Pathology , 27 : 97 – 99 .
- Allan WH Lancaster JE Toth B 1978 Newcastle disease vaccines—their production and use FAO Animal Production Series No. 10 Rome FAO
- Gough , RE . 1973 . Thermostability of Newcastle disease virus in liquid whole egg . Veterinary Record , 93 : 632 – 633 .
- Lamb RA Collins PL Kolakofsky D Melero Y Nagai Y Oldstore MBA Pringle CR Rima BK 2000 Family Paramyxoviridae In van Regenmortel M.V.H. (Ed.) Virus Taxonomy, Seventh report of the International Committee on Taxonomy of Viruses pp. 549–561 New York Academic Press
- MAF NZ 1999 Import Risk Analysis: Chicken Meat 228 pp Wellington New Zealand Ministry of Agriculture and Forestry
- Mayo , MA . 2002 . A summary of the changes recently approved by ICTV . Archives of Virology , 147 : 1655 – 1656 .
- Office International des Epizooties 2000 Manual of Standards for Diagnostic Tests and Vaccines, 2000 957 pp Paris Office International des Epizooties
- Office International des Epizooties 2003 Chapter 2.1.15, Newcastle disease Terrestrial Animal Health Code 2003 Paris Office International des Epizooties http://www.oie.int/eng/normes/mcode/A_00044.htm
- Picken JC 1968 Thermostability of Newcastle disease virus In Hanson R.P. (Ed.) Newcastle disease virus an evolving pathogen pp. 167–188 The University of Wisconsin Press Wisconsin USA
- Russell AD Hugo WB Ayliffe GAJ 1992 Principles and Practices of Disinfection, Preservation and Sterilisation 2nd edn Oxford Blackwell Scientific Publications