
Abstract
Bromalites are the group of ichnofossils that record the consumption, processing, and elimination of material through digestive systems. Each main stage of processing has been ascribed to distinct bromalite subgroups, such as cololites, coprolites, and regurgitalites, with little evidence for transitions between these stages. To expand this limited record, we consider a fossil horseshoe crab—Tachypleus syriacus—from the Cenomanian Hjoula Lagerstätte (Sannine Formation, Lebanon) that showcases one such transition. The specimen illustrates a bromalite both within the animal’s guts (cololite) and as expelled faecal matter (coprolite). This demonstrates a link between these two conditions for fossil chelicerates and highlights the level of exceptional preservation within the Hjoula Lagerstätte. Additionally, the specimen features prosomal nodules unique to limulids, suggesting new automorphic morphologies for T. syriacus.
Russell D. C. Bicknell [[email protected]; [email protected]], Division of Palaeontology (Invertebrates), American Museum of Natural History, New York City, NY, 10027, USA; Palaeoscience Research Centre, School of Environmental & Rural Science, University of New England, Armidale, NSW 2351, Australia; Mohamad Bazzi [[email protected]], Department of Earth and Planetary Sciences, Stanford University, Stanford, CA 94305, USA; Carl Mehling [[email protected]], Division of Palaeontology (Vertebrates), American Museum of Natural History, New York City, NY, 10027, USA; Anastasia Rashkova [[email protected]], Division of Palaeontology (Invertebrates), American Museum of Natural History, New York City, NY, 10027, USA; Madeline V. Pankowski [[email protected]], 16405 Fox Valley Terrace, Rockville, MD, 20853, USA; Mark Botton [[email protected]], Department of Natural Sciences, Fordham University, New York City, NY, 10023, USA; Carmela Cuomo [[email protected]], Invertebrate Palaeontology Division, Yale Peabody Museum, New Haven, CT, 06511, USA; Limulus Ranch LLC, Hamden, Connecticut, 06514, USA.
BROMALITES—the broad ichnofossil group that includes gut contents and prior gut contents—are useful fossils for understanding the diets and digestion processes of extinct animals (Knaust Citation2020). These traces have been categorized according to different stages of digestion (Hunt et al. Citation2012). Two groups that are especially useful within the arthropod fossil record are cololites and coprolites. Cololites (sensu Agassiz, Citation1833) are evidence of fossil digested food within the gastrointestinal tract posterior to the stomach (Kraft et al. Citation2023), while coprolites (sensu Buckland Citation1829) are fossil faecal material eliminated from the body (Häntzschel et al. Citation1968, Hunt et al. Citation2012). While coprolites can be linked to probable producers (Häntzschel et al. Citation1968, Kimmig & Strotz Citation2017, Kimmig & Pratt Citation2018), the majority of trace makers are unknown. Furthermore, the fossil record linking the two trace fossil groups together is very limited. To date, there are rare unpublished examples of insects in amber showing this association, and one published roach in Cretaceous amber showing a bromalite traversing the anus (Hinkelman & Vršanská Citation2020). Here, we expand the record of these transitional trace fossils by examining a fossil horseshoe crab (Xiphosurida)—Tachypleus syriacus (Woodward, Citation1879)—from the Hjoula Lagerstätte of Lebanon that preserves a cololite transitioning into a coprolite.
Geological setting
The late Cenomanian Lebanese marine fossil-fish locality at Hjoula (Arabic: حجولا) belongs to the Sannine Formation facies configuration (Nader et al. Citation2006) and is situated within the Byblos Governorate of Mount Lebanon in Central Lebanon. Precise palaeolatitude calculations (www.paleolatitude.org; van Hinsbergen et al. Citation2015) place the Hjoula limestone quarry at an average 14.36°N equatorial position within the 90-million-year age bracket. During the time of deposition, Lebanon formed part of the African platform at the northern frontiers of the Gondwana supercontinent (Philip et al. Citation1993). The fossiliferous rock units at Hjoula are recognized as a Konservat Lagerstätten producing rich assemblages of exceptionally well-preserved teleost fishes, cartilaginous fishes, cyclostomes, pterosaurs, and numerous invertebrate groups (Parry et al. Citation2015, Marramà et al. Citation2016, Bicknell et al. Citation2019a, Kellner et al. Citation2019, Miyashita et al. Citation2019, Klug et al. Citation2021). The laminated limestones of Hjoula were laid down under anoxic conditions in a confined, tectonically active carbonate basin (Hemleben Citation1977, Swinburne & Hemleben Citation1994, Wippich & Lehmann Citation2004).
Materials and methods
The Tachypleus syriacus fossil, AMNH-IF 141422, was found in 2020 at a quarry in Hjoula at 34°07′59.8ʺ N 35°44′37.9ʺ E. AMNH-IF 141422 was prepared and partially exposed using a pneumatic tool. Due to breakage along a calcite vein, the specimen has been repaired with a Paraloid B-72 adhesive solution. The viscosity of the adhesive was modified by adjusting the polymer concentration to 50% and dissolving in acetone solution (Koob Citation1986). The adhesive was applied systematically in intervals to restructure the void which contained the calcite vein. Measurements of AMNH-IF 141422 were made using digital callipers. However, due to the partly exposed nature of the carapace, measurements represent minimal (incomplete) values. Images of AMNH-IF 141422 were made under normal LED light using an Olympus E-M1MarkIII. For comparison, a fresh horseshoe crab scat was collected from a living Limulus polyphemus specimen in captivity.
Institutional abbreviations
AMNH, American Museum of Natural History, New York, USA.
Results
AMNH-IF 141422 is a female Tachypleus syriacus individual (Lamsdell & McKenzie Citation2015) consisting of a partial prosoma that shows a doublure (92.9 mm long, 131.4 mm wide) and partial thoracetron (34.6 mm long, 81.2 mm wide) (). The gut is preserved as a mould and is a 18.22 mm long section (). The posterior-most section of the gut shows marked relief, and is 12.13 mm long, 5.66 mm wide anteriorly, tapering to 2.91 mm wide posteriorly. At the posterior-most point of the gut, an amorphous, sinuous structure is observed (). This sinuous structure is 18.6 mm long, showing marked disaggregation distally from the body fossil. Shelly material is identified within the sinuous structure, although the exact nature of these fragments is indeterminate.
Fig. 1. Examined Tachypleus syriacus with a cololite transitioning to a coprolite and modern xiphosurid scat. A, Complete specimen, AMNH-IF 141422. B, Close up of cololite transitioning to coprolite. C, Close up of prosomal border showing nodules (white arrows). D, Example of modern xiphosurid scat, morphologically comparable to observed coprolite.
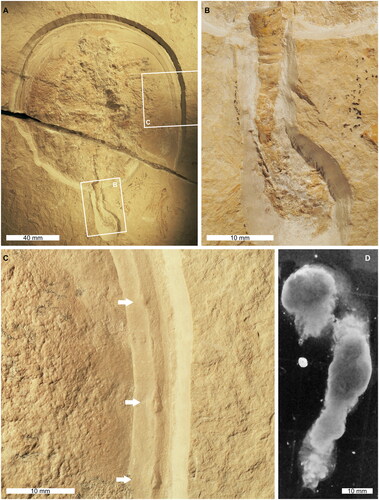
The prosomal rim is exceptionally preserved and shows unique taxonomic characteristics for T. syriacus. These are nodules along the rim proximal to the external margin (). The nodules are 10.78–27.29 mm apart and more prevalent on the right side.
Discussion
The gut section along the thoracetronic midline in AMNH-IF 141422 is comparable to previously documented horseshoe crab cololites (). As such, we confidently consider this structure a cololite. Furthermore, the sinuous structure at the terminus of the bromolite, where the xiphosurid anus is located, is comparable to expelled modern horseshoe crab faeces (Bicknell et al. Citation2022a; compare ). We therefore conclude that this sinuous structure is a coprolite. Taken together, we propose that AMNH-IF 141422 represents a transitional horseshoe crab bromalite. Moreover, the coprolite shows evidence of shelly material (), confirming the hypotheses that ancient xiphosurids had diets comparable to modern forms (Bicknell et al. Citation2019a), and processed shelly material using walking and pushing legs (Bicknell et al. Citation2018a, Citation2018b, Citation2021b).
Table 1. Summary of previously documented xiphosuran cololites. Ordered by time period and higher taxonomic assignment following Bicknell & Pates (Citation2020) and Lamsdell (Citation2020).
The digestive tract of a horseshoe crab is a simple chitin-lined tube, subdivided into several regions (Botton et al. Citation2003). The oesophagus connects the mouth to the muscular proventriculus (gizzard), which is lined with thick ridges of chitin that macerate material. Distally, the food, often mixed with residual sediment and shell matter, is passed through the pyloric valve into the linear, non-convoluted intestine for enzymatic digestion and absorption. The faecal material is encased in mucus, and eliminated at the anus, located ventrally to the telson.
Horseshoe crabs feed on a variety of benthic organisms, such as molluscs, crustaceans, echinoderms and polychaetes (Botton Citation1984, Botton & Haskin Citation1984, Botton & Ropes Citation1989, Debnath et al. Citation1989, Chatterji et al. Citation1992, Razak et al. Citation2017). The consumption of ambient sediment particles is customary when the animals ingest their prey, and crushed shell debris and sediment are commonly found in the faecal material of extant limulids (Botton Citation1984). Thus, we regard the shelly material and sediment in the observed bromalite as typical horseshoe crab feeding behaviour.
AMNH-IF 141422 expands the limited evidence of horseshoe crab cololites that range from the Late Devonian through to the Cenomanian (). At least four of these examples demonstrate cololites with relief compared to the fossil (Bicknell et al. Citation2019b, figs 1A, E; Bicknell et al. Citation2019a, figs 2, 3d; ). This relief suggests that the gut was likely infilled with sediment, reflecting the condition where modern horseshoe crabs consume and pass sediment through their gut while foraging (Botton & Haskin Citation1984, Carmichael et al. Citation2004). We can therefore infer that extinct horseshoe crabs may have consumed sediment as a byproduct of scavenging or predation. However, as we cannot confidently rule out post-mortem sediment infilling, we are tentative in this proposal.
Modern horseshoe crab faecal material decomposes quickly in the marine environment (Cuomo, pers. obs.). The preservation of horseshoe crab coprolites is therefore expected to be rare. Alternatively, these fossils have been documented in exceptional preservation conditions, but are not associated with xiphosurid body fossils. In either case, this area of research should be expanded to further understand these important ichnofossils.
Useful frameworks for documenting and interpreting vertebrate bromalites have been presented (Hunt et al. Citation2012, fig. 1; Přikryl et al. 2012; Hunt & Lucas Citation2021). These can also be applied to arthropod bromalites (Knaust, Citation2020; Bicknell et al. Citation2023, Bicknell et al. Citation2024). Despite this research, no term has been presented to describe transitional bromalites. Therefore, the trace fossil presented here is simultaneously a cololite proximally and a coprolite distally.
Prosomal nodules observed on AMNH-IF 141422 are unique within Limulidae (Bicknell et al. Citation2021a). Furthermore, they have not previously been noted on Tachypleus syriacus prosomae (Woodward Citation1879, Lamsdell & McKenzie Citation2015, Bicknell et al. Citation2019a). As such, these structures may represent a novel autapomorphy for the species. The palaeobiological implications for these nodules are also interesting to consider. Nodules in Limulidae are commonly associated with dorsal spines, including in T. syriacus (see Lamsdell & McKenzie Citation2015). However, adult xiphosurids lack spines along the anterior and lateral prosomal margin (see Bicknell & Pates Citation2020, Lamsdell Citation2020). These nodules must therefore have had another use. One possibility is that they represent a sensory structure. Juvenile horseshoe crabs have sensory mechanoreceptors along the prosomal margin (Fornshell Citation2022). However, these structures are lost through ontogeny (Fornshell Citation2022). In T. syriacus, modified sensory mechanoreceptors may have been retained, producing these nodules, although it is impossible to state if these were functional or vestigial. Alternatively, these morphologies may be associated with the sexually dimorphic nature of the species (Lamsdell & McKenzie Citation2015). However, the exact use remains unknown.
Acknowledgements
This research was funded by a MAT Program Postdoctoral Fellowship (to RDCB), a Knut and Alice Wallenberg Foundation grant (KAW 2022.0330 to MB). We thank Bushra Hussaini for aid with collections. David Rudkin and the Editorial Board of Alcheringa suggested changes that improved the manuscript.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Correction Statement
This article has been corrected with minor changes. These changes do not impact the academic content of the article.
References
- Agassiz, L., 1833. Neue Entdeckungen über fossile Fische. Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefaktenkunde 1833, 675–677.
- Bicknell, R.D.C., Błażejowski, B., Wings, O., Hitij, T. & Botton, M.L., 2021a. Critical re-evaluation of Limulidae reveals limited Limulus diversity. Papers in Palaeontology 7, 1525–1556.
- Bicknell, R.D.C., Birch, S.A., Charbonnier, S., Sautereau, F., Hitij, T. & Campione, N.E., 2019a. On the appendicular anatomy of the xiphosurid Tachypleus syriacus and the evolution of fossil horseshoe crab appendages. Die Naturwissenschaften 106, 38.
- Bicknell, R.D.C., Bruthansová, J. & Kimmig, J., 2024. Shelly coprolites record durophagous predation in the Late Ordovician Bohdalec Formation (Katian; Prague Basin, Czech Republic). Geobios 82, 1–11.
- Bicknell, R.D.C., Holmes, J.D., Edgecombe, G.D., Losso, S.R., Ortega-Hernández, J., Wroe, S. & Paterson, J.R., 2021b. Biomechanical analyses of Cambrian euarthropod limbs reveal their effectiveness in mastication and durophagy. Proceedings. Biological Sciences 288, 20202075.
- Bicknell, R.D.C., Holmes, J.D., Pates, S., García-Bellido, D.C. & Paterson, J.R., 2022a. Cambrian carnage: trilobite predator-prey interactions in the Emu Bay Shale of South Australia. Palaeogeography, Palaeoclimatology, Palaeoecology 591, 110877.
- Bicknell, R.D.C., Ledogar, J.A., Wroe, S., Gutzler, B.C., Watson, W.H., III,. & Paterson, J.R., 2018a. Computational biomechanical analyses demonstrate similar shell-crushing abilities in modern and ancient arthropods. Proceedings of the Royal Society B 285, 20181935.
- Bicknell, R.D.C., Paterson, J.R., Caron, J.-B. & Skovsted, C.B., 2018b. The gnathobasic spine microstructure of recent and Silurian chelicerates and the Cambrian artiopodan Sidneyia: functional and evolutionary implications. Arthropod Structure & Development 47, 12–24.
- Bicknell, R.D.C., Lustri, L. & Brougham, T., 2019b. Revision of ‘Bellinurus’ carteri (Chelicerata: Xiphosura) from the Late Devonian of Pennsylvania, USA. Comptes Rendus Palevol 18, 967–976.
- Bicknell, R.D.C., Naugolnykh, S.V. & McKenzie, S.C., 2022b. On Paleolimulus from the Mazon Creek Konservat-Lagerstätte. Comptes Rendus Palevol 21, 303–322.
- Bicknell, R.D.C. & Pates, S., 2020. Pictorial atlas of fossil and extant horseshoe crabs, with focus on Xiphosurida. Frontiers in Earth Science 8, 98.
- Bicknell, R.D.C., Smith, P.M. & Kimmig, J., 2023. Novel coprolitic records from the Silurian (Přídolí) Wallace Shale of New South Wales. Alcheringa 47, 24–30.
- Botton, M.L., 1984. Diet and food preferences of the adult horseshoe crab Limulus polyphemus in Delaware Bay, New Jersey, USA. Marine Biology 81, 199–207.
- Botton, M.L. & Haskin, H.H., 1984. Distribution and feeding of the horseshoe-crab, Limulus polyphemus, on the continental-shelf off New-Jersey. Fishery Bulletin 82, 383–389.
- Botton, M.L. & Ropes, J.W., 1989. Feeding ecology of horseshoe crabs on the continental shelf, New Jersey to North Carolina. Bulletin of Marine Science 45, 637–647.
- Botton, M.L., Shuster, C.N., Jr.,. & Keinath, J.A., 2003. Horseshoe crabs in a food web: Who eats whom. In The American Horseshoe Crab. Shuster Jr., C.N., Barlow, R.B. & Brockmann, H.J., eds. Cambridge: Harvard University Press, pp. 133–153.
- Buckland, W., 1829. XII. – On the Discovery of Coprolites, or Fossil Fæces, in the Lias at Lyme Regis, and in other Formations. Transactions of the Geological Society of London 3, 223–236.
- Carmichael, R.H., Rutecki, D., Annett, B., Gaines, E. & Valiela, I., 2004. Position of horseshoe crabs in estuarine food webs: N and C stable isotopic study of foraging ranges and diet composition. Journal of Experimental Marine Biology and Ecology 299, 231–253.
- Chatterji, A., Mishra, J.K. & Parulekar, A.H., 1992. Feeding behaviour and food selection in the horseshoe crab, Tachypleus gigas (Müller). Hydrobiologia 246, 41–48.
- Debnath, R., Nag, S.K., Choudhury, A., Dasgupta, R. & Sur, R., 1989. Feeding habit and digestive physiology of the Indian horseshoe crab, Tachypleus gigas (Müller). Indian Journal of Physiology and Allied Sciences 43, 44–49.
- Eller, E.R., 1940. Belinurus carteri: a new xiphosuran from the Upper Devonian of Pennsylvania. Annals of the Carnegie Museum 28, 133–136.
- Fornshell, J.A., 2022. The potential functions of mechanoreceptors found on trilobite larva of Limulus polyphemus (Linnaeus, 1758). Arthropods 11, 127–134.
- Häntzschel, W., El-Baz, F. & Amstutz, G.C., 1968. Coprolites an Annotated Bibliography, Geological Society of America, Boulder, CO.
- Hemleben, Cv., 1977. Rote Tiden und die oberkretazischen Plattenkalke im Libanon. Neues Jahrbuch für Geologie und Paläontologie. Monatshefte 1, 239–255.
- Hinkelman, J. & Vršanská, L., 2020. A Myanmar amber cockroach with protruding feces contains pollen and a rich microcenosis. The Science of Nature107, 13.
- Hunt, A.P. & Lucas, S.G., 2021. The Ichnology of Vertebrate Consumption: Dentalites, Gastroliths and Bromalites. Bulletin of the New Mexico Museum of Natural History and Science 87, 1–216.
- Hunt, A.P., Milàn, J., Lucas, S.G. & Spielmann, J.A., 2012. Vertebrate coprolite studies: status and prospectus. New Mexico Museum of Natural History and Science Bulletin 57, 5–24.
- Kellner, A.W.A., Caldwell, M.W., Holgado, B., Vecchia, F.M.D., Nohra, R., Sayão, J.M. & Currie, P.J., 2019. First complete pterosaur from the Afro-Arabian continent: insight into pterodactyloid diversity. Scientific Reports 9, 17875.
- Kimmig, J. & Pratt, B.R., 2018. Coprolites in the Ravens Throat River Lagerstätte of Northwestern Canada: implications for the middle Cambrian food web. PALAIOS 33, 125–140.
- Kimmig, J. & Strotz, L.C., 2017. Coprolites in mid-Cambrian (Series 2-3) Burgess Shale-type deposits of Nevada and Utah and their ecological implications. Bulletin of Geosciences 92, 297–309.
- Klug, C., Pohle, A., Roth, R., Hoffmann, R., Wani, R. & Tajika, A., 2021. Preservation of nautilid soft parts inside and outside the conch interpreted as central nervous system, eyes, and renal concrements from the Lebanese Cenomanian. Swiss Journal of Palaeontology 140, 15.
- Knaust, D., 2020. Invertebrate coprolites and cololites revised. Papers in Palaeontology 6, 385–423.
- Koob, S.P., 1986. The use of Paraloid B-72 as an adhesive: its application for archaeological ceramics and other materials. Studies in Conservation 31, 7–14.
- Kraft, P., Vaškaninová, V., Mergl, M., Budil, P., Fatka, O. & Ahlberg, P.E., 2023. Uniquely preserved gut contents illuminate trilobite palaeophysiology. Nature 622, 545–551.
- Lamsdell, J.C., 2020. The phylogeny and systematics of Xiphosura. Peerj. 8, e10431.
- Lamsdell, J.C. & McKenzie, S.C., 2015. Tachypleus syriacus (Woodward) – a sexually dimorphic Cretaceous crown limulid reveals underestimated horseshoe crab divergence times. Organisms Diversity & Evolution 15, 681–693.
- Marramà, G., Villier, B., Dalla Vecchia, F.M. & Carnevale, G., 2016. A new species of Gladiopycnodus (Coccodontoidea, Pycnodontomorpha) from the Cretaceous of Lebanon provides new insights about the morphological diversification of pycnodont fishes through time. Cretaceous Research 61, 34–43.
- Miyashita, T., Coates, M.I., Farrar, R., Larson, P., Manning, P.L., Wogelius, R.A., Edwards, N.P., Anné, J., Bergmann, U., Palmer, A.R. & Currie, P.J., 2019. Hagfish from the Cretaceous Tethys Sea and a reconciliation of the morphological–molecular conflict in early vertebrate phylogeny. Proceedings of the National Academy of Sciences of the United States of America 116, 2146–2151.
- Nader, F.H., Abdel-Rahman, A.-F.M. & Haidar, A.T., 2006. Petrographic and chemical traits of Cenomanian platform carbonates (central Lebanon): implications for depositional environments. Cretaceous Research 27, 689–706.
- Parry, L.A., Wilson, P., Sykes, D., Edgecombe, G.D. & Vinther, J., 2015. A new fireworm (Amphinomidae) from the Cretaceous of Lebanon identified from three-dimensionally preserved myoanatomy. BMC Evolutionary Biology 15, 256.
- Philip, J., Babinot, J.F., Tronchetti, G., Fourcade, E., Ricou, L.E., Guiaud, R., Bellion, Y., Herbin, J.P., Combes, P.E., Conee, J.J. & Dercourt, J., 1993. Late Cenomanian palaeoenvironments (94 to 92 Ma). In Atlas Tethys Palaeoenvironmental Maps. Dercourt, J., Ricou, L.E. & Vrielynck, B., eds. Paris: Gauthier-Villars, p. 153–178.
- Přikryl, T., Košták, M., Mazuch, M. & Mikuláš, R., 2012. Evidence for fish predation on a coleoid cephalopod from the Lower Jurassic Posidonia Shale of Germany. Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen 263, 25–33.
- Razak, M.R.M., Kassim, Z., Sabuti, A.A. & Ismail, A., 2017. Feeding ecology and food preferences of Cherok Paloh, Pahang horseshoe crab, Tachypleus gigas. Malaysian Journal of Fundamental and Applied Sciences 13, 198–202.
- Swinburne, N.H.M. & Hemleben, C., 1994. The plattenkalk facies: a deposit of several environments. Geobios 27, 313–320.
- van Hinsbergen, D.J.J., De Groot, L.V., van Schaik, S.J., Spakman, W., Bijl, P.K., Sluijs, A., Langereis, C.G. & Brinkhuis, H., 2015. A paleolatitude calculator for paleoclimate studies. PLoS ONE 10, e0126946.
- Wippich, M.G.E. & Lehmann, J., 2004. Allocrioceras from the Cenomanian (mid‐Cretaceous) of the Lebanon and its bearing on the palaeobiological interpretation of heteromorphic ammonites. Palaeontology 47, 1093–1107.
- Woodward, H., 1879. Contributions to the knowledge of fossil Crustacea. Quarterly Journal of the Geological Society 35, 549–556.