
ABSTRACT
Functional variants of immune-related genes may be implicated in the occurrence of colorectal cancer (CRC). In this study, Programmed cell death (PDCD)-1.6 (rs10204525 T/C), PDCD-1.7 (rs7421861 A/G), and PDCD-1.9 (rs2227982 A/G) loci were selected to explore gene expression and the potential susceptibility to the development of CRC. Here, 1,003 CRC patients and 1,303 controls were included and three PDCD-1 tagging loci were selected and analyzed by using SNPscan genotyping assays. SHESIS software was harnessed to obtain the haplotypes of the PDCD-1 gene. We found that the genotype and allele distribution of PDCD-1 tagging loci did not significantly affect the risk of CRC. Adjustment for body mass index, age, smoking, alcohol using and sex also found that PDCD-1 tagging loci did not influence the occurrence of CRC. In conclusion, this study suggests that the PDCD-1 tagging loci (rs10204525, rs7421861, and rs2227982) are not correlated with CRC susceptibility.
Introduction
Colorectal cancer (CRC) is a common malignancy worldwide. In 2018, CRC ranked fourth in term of incidence with a total of 1,096,601 cases diagnosed (Bray et al. Citation2018). A number of publications attribute CRC etiology to certain vital components of the environment (Campos et al. Citation2005; Jemal et al. Citation2011; Kune and Vitetta Citation1992; Tiemersma et al. Citation2002). Evaluating the trends of incidence and mortality in the most recent decade, Arnold et al. reported that both incidence and mortality increasing in China (Arnold et al. Citation2017). It is worth noting that the increasing trends of CRC incidence and mortality may be associated with some environment factors (e.g., physically inactive, alcohol consumption, tobacco using, low intake of vegetables and fruits, high intake of dietary fat, body fatness, and processed meat) (Al-Zalabani Citation2020; Cho et al. Citation2019; Dong et al. Citation2017; Gerber Citation2009; Lund et al. Citation2011; Nagle et al. Citation2015; Shirakami et al. Citation2017; Weitz et al. Citation2005; Xu et al. Citation2016). Besides these environmental factors mentioned above, evidence has also indicated that an individual’s genetic components are associated with CRC, and recent studies have identified immune related genes that may be implicated in the occurrence, progression, and prognosis of CRC (Chen and Chen Citation2014; Grasso et al. Citation2018; Zou et al. Citation2018).
The programmed cell death-1 gene (PDCD-1) is located on chromosome 2q37.3 and codes for a type I transmembrane glycoprotein with a 50–55 kD molecular weight. PDCD-1 is expressed on some activated immune cells (e.g., some dendritic cell, B cells, natural killer T, and CD8+ and CD4+ T cells) (Keir et al. Citation2008). PDCD-1 belongs to the immunoglobulin superfamily (IgSF), and is comprised of three domains: an extracellular and a cytoplastic domain and a transmembrane region. Immunoglobulin-like construction is located in extracellular domain, and the immunoreceptor tyrosine-based switch motif lies in transmembrane region. As well, the immunoreceptor tyrosine-based inhibitory motif is located in the cytoplasmic domain. When the interaction of PDCD-1 with its ligands PDCD-1 ligand 1 (PD-L1 and PD-L2) activates PDCD-1 that is a negative regulator for the effector function of T-lymphocytes, which may strongly block both cytokine production and proliferation of CD8+ and CD4+ T cells (Ai et al. Citation2020; Riley Citation2009). It has been reported that PD-L1 is expressed by certain CRC, which treated with aspirin, folinic acid, 5-FU, and oxaliplatin (Faruk et al. Citation2021; Peng et al. Citation2021; Siraj et al. Citation2021). Additionally, PDCD-1 was implicated in regulating the function of regulatory T-cells (Chen et al. Citation2020; Dong et al. Citation2020; Kumagai et al. Citation2020). In view of the negative regulation of PDCD-1 in cancer related immune response, we could hypothesize that PDCD-1 might be a useful candidate for individual’s hereditary susceptibility to CRC.
Of late, some publications have studied the role of PDCD-1 single-nucleotide polymorphisms (SNPs) in a variety of tumors. Qiu reported that the PDCD-1.6 T/C polymorphism might decrease the risk of esophageal squamous cell carcinoma among males and younger patients (Qiu et al. Citation2014). A recent study found that PDCD-1.5 C/T could increase the susceptibility to gestational trophoblastic diseases (Dehaghani et al. Citation2009). In Chinese Han females, the frequencies of PDCD-1.5 CT and PDCD-1.1 GG were lower in breast cancer patients compared with controls (Hua et al. Citation2011). The combined genotypes of PDCD-1 +8669 AA/TIM3–1516 GT or TT was more frequent in hepatocellular carcinoma and cirrhosis patients (Li et al. Citation2013). Savabkar and colleagues demonstrated that PDCD-1.5 C/T was associated with the risk of gastric cancer (Savabkar et al. Citation2013). Our previous study also PDCD-1.7 (rs7421861 A/G) and PDCD-1.9 (rs2227982 A/G) could influence the risk of esophagogastric junction adenocarcinoma (Tang et al. Citation2017). Additionally, a case-control study found that PDCD-1.7 (rs7421861 A/G) could increase the risk of CRC (Ge et al. Citation2015). Additionally, in Chinese populations, the PDCD-1.1 (rs36084323) G allele promoted the advanced CRC TNM stage (Zhao et al. Citation2018). Mojtahedi et al. reported that PDCD-1.5 C/T could increase the susceptibility to colon cancer (Mojtahedi et al. Citation2012). However, due to the moderate sample size, the observations might be underpowered. The aim of this study was to clarify this issue more accurately. Here, we recruited 1,003 CRC patients and 1,303 controls to carried out a case-control study to evaluate the correlation between PDCD-1 tagging SNPs and the risk of CRC.
Materials and methods
Study population
The current case-control study was designed as a hospital-based investigation that consists of 1,303 cancer-free controls and 1,003 CRC patients. All the subjects were from Eastern China. They were unrelated Chinese populations. The participants were recruited at the No.1 people’s hospital of Zhenjiang City and Affiliated Union Hospital of Fujian Medical University. By using surgical or endoscopic biopsy specimens, CRC patients were diagnosed by two experienced pathologists. The included criterion were: (a) CRC patients were Chinese Han population, (b) CRC was confirmed by pathological diagnosis, and (c) cases were sporadic CRC patients. The excluded criterion of CRC patients were: (a) CRC patients having secondary or recurrent tumors, (b) patients who had received chemoradiotherapy and (c) patients who were diagnosed as hereditary CRC cases. A total of 1,303 cancer-free subjects matched to CRC patients by sex and age (within 5 years) were included as controls. When participants were interviewed, a structured questionnaire was harnessed to obtain the useful data on demographic information and risk factors. The weight/height2 (kg/m2) was used to calculate body mass index (BMI). The criterion of overweight and obesity for Chinese was BMI ≥ 24 kg/m2 (Zhai et al. Citation2010; Zhang et al. Citation2009). Those who currently or formerly smoked ≥1 cigarette per day over a year were considered as smokers and those who drank ≥3 times/week for more than 6 months were defined as drinkers (Tang et al. Citation2014). Informed consent was signed by each subject. The Ethics Committee of Fujian Medical University approved this investigation (No. KT 2018-003-01).
SNP selection and genotyping
The method of selecting PDCD-1 tagging SNPs has been summarized in our previous publications (Zhang et al. Citation2017; Zou et al. Citation2018). The PDCD-1.6 (rs10204525 T/C), PDCD-1.7 (rs7421861 A/G), and PDCD-1.9 (rs2227982 A/G) SNPs were selected for this study. The process for extraction of genomic DNA and quality control have been described in our previous report (Qiu et al. Citation2021). The SNPs of the PDCD-1 gene was genotyped by using a 48-Plex SNPscan assay (Ding et al. Citation2017). The quality control for genotyping was performed as follows: (1) the SNP scan assay was carried out without knowing the status of CRC cases or controls; and (2) two authors independently repeated assays and examined the data. Finally, we randomly selected and retest 4% of the samples. And those observations were 100% concordant.
Statistical analysis
The analysis was performed by using SAS 9.4 software for Windows (SAS Institute, Cary, USA). The Student’s t-test was used to assess the difference in continuous variables between CRC patients and cancer-free controls. We used the Pearson χ2 test to evaluate the difference of genotype frequencies and categorical variables between the groups. By using a goodness-of-fit χ2 test, the P values for Hardy-Weinberg equilibrium (HWE) for PDCD-1 tagging SNPs were calculated. PDCD-1 haplotypes have been obtained by using an online SHEsis software (http://analysis.bio-x.cn/myAnalysis.php) (Chen et al. Citation2018; Li et al. Citation2009; Liu et al. Citation2019; Qiu et al. Citation2017; Shi and He Citation2005; Wang et al. Citation2015). Odds ratios (ORs) and 95% confidence intervals (CIs) were used to express the potential relationship of PDCD-1 tagging SNPs with susceptibility of CRC. All P values of statistical analyses were two-tailed. And P < .05 was considered as the criterion of statistically significance.
Results
summarizes clinical features, demographic information, and risk factors of cases and controls. Two groups were well matched with gender and age in Pearson χ2 test (P = .867and .605). In the case group, the mean age was 61.10 ± 12.17 years and the mean age was 61.40 ± 9.61 years in control group. When the mean age was compared in two groups, the P value was .496. In total, 620 males and 383 females were recruited in the case group. In addition, the controls contained 1,303 cancer-free subjects (801 males and 502 females). The distribution of smoking status, drinking, and BMI were different in the two groups [smoking status of CRC patients vs. controls (ever/never): 259/744 vs. 265/1,038, P = .002; drinking of CRC patients vs. controls: (ever/never): 174/829 vs. 136/1,167, P < .001 and BMI of CRC patients vs. controls: (<24 kg/m2/≥24 kg/m2): 670/333 vs. 688/615, P < .001]. The genotype of the PDCD-1.6, PDCD-1.7, and PDCD-1.9 SNPs in controls were in accordance with HWE. The minor allele frequency (MAF) of PDCD-1.6, PDCD-1.7, and PDCD-1.9 SNPs was 0.277, 00.164,and 0.495, respectively (). The detailed genotypes of PDCD1 are summarized in Table S1.
Table 1. Distribution of selected risk factors and demographic variables in CRC cases and controls
Table 2. Data for PDCD-1 tagging SNPs
Association of PDCD-1 tagging SNPs with CRC
The distribution of PDCD-1.6, PDCD-1.7, and PDCD-1.9 genotypes in CRC patients and controls are summarized in .
Table 3. Logistic regression analyses of associations between PDCD-1 polymorphisms and the risk of CRC
The variant frequency of PDCD-1.6 was 51.98% (TT), 39.45% (CT), and 8.66% (CC) in CRC ccases,and 52.92% (TT), 38.85% (CT), and 8.23% (CC) in controls. When PDCD-1.6 TT frequency was used as a reference, the PDCD-1.6 TC genotype could not change a risk of CRC (P = .694). When compared to the PDCD-1.6 TT frequency, the PDCD-1.6 CC genotype did not alter the susceptibility of CRC (P = .649). When compared to the PDCD-1.6 TT/TC frequency, the PDCD-1.6 CC genotype did not change the susceptibility of CRC (P = .711). When the PDCD-1.6 TT genotype frequency was used as a reference, the PDCD-1.6 TT/CT genotype also did not affect CRC susceptibility (P = .623). Adjustments for included risk factors and demographic information, we concluded that PDCD-1.6 did not change CRC risk (all adjusted P ≥ .05; ).
The variant frequencies of PDCD-1.7 were 70.51% (AA), 26.53% (AG), and 2.96% (GG) in CRC cases and 70.38% (AA), 26.38% (AG), and 3.23% (GG) in controls. When the PDCD-1.7 AA frequency was used as a reference, the PDCD-1.7 AG genotype did not change the risk of CRC (P = .969). When compared to the PDCD-1.7 AA frequency, the PDCD-1.7 GG genotype did not influence the susceptibility to CRC (P = .718). When compared to the PDCD-1.7 AA frequency, the PDCD-1.7 AG/GG genotype did not alter the susceptibility of CRC (P = .948). When the PDCD-1.7 AA/AG genotype frequency was used as reference, the PDCD-1.7 GG genotype also did not modulate the risk of CRC (P = .714). We identified that PDCD-1.7 did not change CRC risk after the multiple regression (all adjusted P ≥ .05; ).
The variant frequencies of PDCD-1.9 were 25.92% (AA), 50.51% (AG), and 23.57% (GG) in the CRC cases, and 25.62% (AA), 49.69% (AG), and 24.69% (GG) in controls. When the PDCD-1.9 AA frequency was used as a reference, the PDCD-1.9 AG genotype did not modify the risk of CRC (P = .964). When compared to the PDCD-1.9 AA frequency, the PDCD-1.9 GG genotype did not influence the susceptibility to CRC (P = .628). When compared to the PDCD-1.9 AA frequency, the PDCD-1.7 AG/GG genotype did not alter the susceptibility to CRC (P = .870). When the PDCD-1.9 AA/AG genotype frequency was used as a reference, the PDCD-1.9 GG genotype also did not affect the risk of CRC (P = .537). With adjustments for included factors, we identified that PDCD-1.9 did not change CRC risk (all adjusted P ≥ .05; ).
Association of PDCD-1 tagging SNPs with CRC in subgroup analysis
The associations of PDCD-1.6, PDCD-1.7, and PDCD-1.9 genotypes with the risk of CRC in subgroup analyses were summarized in , respectively. We found that there was null association in any comparison.
Table 4. Stratified analyses between PDCD-1.6 polymorphism and CRC risk by sex, age, BMI, smoking status, and alcohol consumption
Table 5. Stratified analyses between PDCD-1.7 polymorphism and CRC risk by sex, age, BMI, smoking status, and alcohol consumption
Table 6. Stratified analyses between PDCD-1.9 polymorphism and CRC risk by sex, age, BMI, smoking status, and alcohol consumption
Association of PDCD-1 haplotypes with CRC
TAA, TGA, CGG, CGA, TGG, and others (PDCD-1.6, PDCD-1.7, and PDCD-1.9 order) PDCD-1 haplotypes were, obtained by using an online SHEsis software (Tang et al. Citation2019; Yang et al. Citation2019; Zhang et al. Citation2019) (). When the TAA haplotype was used as a reference, those PDCD-1 haplotypes indicated that they did not alter the susceptibility of CRC (P = .181, .654, .380, .072, and .294, respectively) ()
Figure 1. Linkage disequilibrium correlations among different loci.
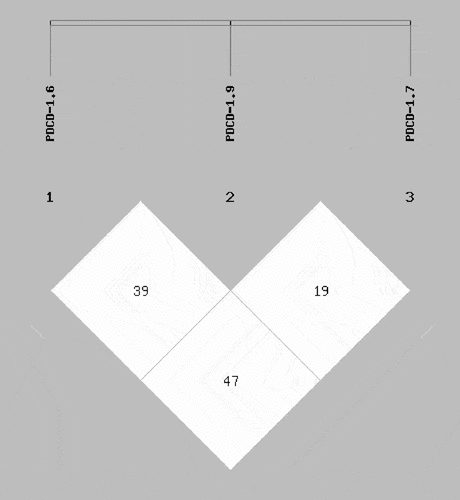
Table 7. Combination analysis of PDCD-1 polymorphisms in CRC patients and controls
Discussion
In the present investigation, three tagging SNPs of PDCD-1 were explored in 1,003 CRC cases and 1,303 cancer-free controls; however, no difference between tagging SNPs of PDCD-1 and the risk of CRC was found.
The present investigation has been launched to investigate the hypothesis that CRC occurrence may be related to an immune disorder. A PDCD-1 SNP may alter PDCD-1 expression on the surface of activated T-lymphocytes and result in an imbalance that influences the immune activity of T cells.
Due to the vital role of PDCD-1 tagging SNPs (rs10204525 on the 3’-UTR, rs7421861 on intron 1, and rs2227982 on the exon 5 region) (Qiu et al. Citation2014; Tang et al. Citation2017), these SNPs have been widely explored for their relationship with multiple cancers (Ge et al. Citation2015; Qiu et al. Citation2014; Tang et al. Citation2015; Zang et al. Citation2020; Zhou et al. Citation2016). Previous studies obtained inconsistent results. For instance, Tang et al. reported that the PDCD-1.7 A/G significantly increased the risk of esophagogastric junction adenocarcinoma in Chinese populations (Tang et al. Citation2017), whereas a previous study had found that the PDCD-1.7 G allele decreased the risk of esophageal cancer (Zang et al. Citation2020). Two recent meta-analyses showed that PDCD-1.6, PDCD-1.7, and PDCD-1.9 SNPs were not associated with cancer development (Dong et al. Citation2016; Zhang et al. Citation2016). In the future, more studies should be performed to detect the relationship of PDCD-1.6, PDCD-1.7, and PDCD-1.9 SNPs with the development of cancer. Our data also should be used to help direct more detailed analysis.
Although these findings of the current investigation could not confirm a relationship of the PDCD-1.6, PDCD-1.7, and PDCD-1.9 haplotypes and variants with a risk of CRC, a previous publication of results from a Chinese Han population suggested that PDCD-1.7, and PDCD-1.9 could be implicated in the occurrence of CRC (Ge et al. Citation2015). In this report, we investigated the conflicting findings between these recent investigations and the found that inconsistencies could be due to various causes.
Firstly, CRCs located in different regions may have diverse pathological characteristics. A recent study has suggested that patients with CRC who have a habit of alcohol consumption and smoking are more likely to develop proximal colon cancers (Wang et al. Citation2020). For family history of CRC and age, Wang et al. reported a positive correlation with a reduced risk of cancer, from cecum to rectum (Nagai et al. Citation2016; Wang et al. Citation2020). Additionally, CRCs in the distal or proximal site were related to inconsistent clinical-pathological characteristics (Oh et al. Citation2008).
Secondly, on the basis of the consensus molecular subtype (CMS) by The CRC Subtyping Consortium (CRCSC), the vital CMSs of CRCs are CMS1, CMS2, CMS3, and CMS4. CMS1 is an immune activated type of microsatellite instability, which shows mismatch repair gene mutation and microsatellite instability. CMS2 is a typical subtype with abnormal activation in the Wnt and Myc signaling pathways and significant copy number variation. CMS3 is a metabolic subtype with a higher ratio of KRAS mutations. CMS4 is a subtype of mesenchyme, in which the transforming growth factor-β signaling pathway is abnormally activated. And then, 13% of CRC patients are mixed. Among them, CMS1 cases are more likely to originate in right-sided colon and CMS2 and CMS3 cases are more likely to originate in the left-sided colon (Lee et al. Citation2017; Wielandt et al. Citation2017). CMS4 cases show in advanced stages and has a presence during metastasis (Wielandt et al. Citation2017).
Thirdly, the individual’s inherited background, different exposure level to risk factors, intervention of lifestyles, and diabetes history might modify the findings.
In this study, several advantages should be addressed: (a) we have recruited more participants than previous studies. The current study has included 1,003 CRC cases and 1,303 controls, which decreases selection bias and some uncharted confounders; (b) PDCD-1 tagging SNPs were selected and studied, which might represent wider and dependable findings for a relationship of PDCD-1 tagging SNPs with CRC risk.
Although there were several advantages mentioned above, some insufficiencies of design should be acknowledged. For example, in some participants, the data of family history and CMS were not collected. As we know, family history can influence the occurrence and development for CRC. Besides, in non-MSI CRCs, the expression of immunoscore-like metagenes led to a better survive, and displays a lower level of both immune checkpoints and tumor-infiltrating T cells (Marisa et al. Citation2018). Additionally, although this study could not attest to the relationship between PDCD-1 SNPs and risk of CRC, some epigenetic biomarkers might have influenced CRC susceptibility, which should be investigated thoroughly in another study. Thirdly, due to lack of some clinical information, we did not carry out subgroup analysis for them. Finally, although some confounding factors (alcohol use, age, sex, smoking status, and BMI status) have been adjusted to treat the difference between cases and controls, this study was designed as a hospital-based investigation and the bias of selection cannot be ignored.
Conclusion
PDCD-1 SNPs (PDCD-1.6, PDCD-1.7, and PDCD-1.9) may not influence the development of CRC in Chinese populations.
Acknowledgements
We appreciate all subjects who participated in this study. We wish to thank Dr. Yan Liu (Genesky Biotechnologies Inc., Shanghai, China) for technical support.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
References
- Ai L, Xu A, Xu J. 2020. Roles of PD-1/PD-L1 pathway: signaling, cancer, and beyond. Adv Exp Med Biol. 1248:33–59.
- Al-Zalabani A. 2020. Preventability of colorectal cancer in Saudi Arabia: fraction of cases attributable to modifiable risk factors in 2015–2040. Int J Environ Res Public Health. 17:320.
- Arnold M, Sierra MS, Laversanne M, Soerjomataram I, Jemal A, Bray F. 2017. Global patterns and trends in colorectal cancer incidence and mortality. Gut. 66:683–91.
- Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. 2018. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 68:394–424.
- Campos FG, Logullo Waitzberg AG, Kiss DR, Waitzberg DL, Habr-Gama A, Gama-Rodrigues J. 2005. Diet and colorectal cancer: current evidence for etiology and prevention. Nutr Hosp. 20:18–25.
- Chen J, Chen Z. 2014. The effect of immune microenvironment on the progression and prognosis of colorectal cancer. Med Oncol. 31:82.
- Chen S, Qiu H, Liu C, Wang Y, Tang W, Kang M. 2018. Relationship between IGF2BP2 and IGFBP3 polymorphisms and susceptibility to non-small-cell lung cancer: a case-control study in Eastern Chinese Han population. Cancer Manag Res. 10:2965–75.
- Chen JX, Yi XJ, Gao SX, Sun JX. 2020. The possible regulatory effect of the PD-1/PD-L1 signaling pathway on Tregs in ovarian cancer. Gen Physiol Biophys. 39:319–30.
- Cho YA, Lee J, Oh JH, Chang HJ, Sohn DK, Shin A, Kim J. 2019. Genetic risk score, combined lifestyle factors and risk of colorectal cancer. Cancer Res Treat. 51:1033–40.
- Dehaghani AS, Kashef MA, Ghaemenia M, Sarraf Z, Khaghanzadeh N, Fattahi MJ, Ghaderi A. 2009. PDCD1, CTLA-4 and p53 gene polymorphism and susceptibility to gestational trophoblastic diseases. J Reprod Med. 54:25–31.
- Ding G, Wang Y, Chen Y, Yin J, Liu C, Fan Y, Qiu H, Tang W, Chen S. 2017. Methylenetetrahydrofolate reductase tagging polymorphisms are associated with risk of esophagogastric junction adenocarcinoma: a case-control study involving 2,740 Chinese Han subjects. Oncotarget. 8:111482–94.
- Dong W, Gong M, Shi Z, Xiao J, Zhang J, Peng J. 2016. Programmed cell death-1 polymorphisms decrease the cancer risk: a meta-analysis involving twelve case-control studies. PloS One. 11:e0152448.
- Dong Y, Han Y, Huang Y, Jiang S, Huang Z, Chen R, Yu Z, Yu K, Zhang S. 2020. PD-L1 is expressed and promotes the expansion of regulatory T cells in acute myeloid leukemia. Front Immunol. 11:1710.
- Dong Y, Zhou J, Zhu Y, Luo L, He T, Hu H, Liu H, Zhang Y, Luo D, Xu S, et, and Teng Z. 2017. Abdominal obesity and colorectal cancer risk: systematic review and meta-analysis of prospective studies. Biosci Rep. 37(6):BSR20170945.
- Faruk M, Ibrahim S, Aminu SM, Adamu A, Abdullahi A, Suleiman AM, Rafindadi AH, Mohammed A, Iliyasu Y, Idoko J, et al. 2021. Prognostic significance of BIRC7/Livin, Bcl-2, p53, Annexin V, PD-L1, DARC, MSH2 and PMS2 in colorectal cancer treated with FOLFOX chemotherapy with or without aspirin. PloS One. 16:e0245581.
- Ge J, Zhu L, Zhou J, Li G, Li Y, Li S, Wu Z, Rong J, Yuan H, Liu Y, et al. 2015. Association between co-inhibitory molecule gene tagging single nucleotide polymorphisms and the risk of colorectal cancer in Chinese. J Cancer Res Clin Oncol. 141:1533–44.
- Gerber M. 2009. Background review paper on total fat, fatty acid intake and cancers. Ann Nutr Metab. 55:140–61.
- Grasso CS, Giannakis M, Wells DK, Hamada T, Mu XJ, Quist M, Nowak JA, Nishihara R, Qian ZR, Inamura K, et al. 2018. Genetic mechanisms of immune evasion in colorectal cancer. Cancer Discov. 8:730–49.
- Hua Z, Li D, Xiang G, Xu F, Jie G, Fu Z, Jie Z, Da P, Li D. 2011. PD-1 polymorphisms are associated with sporadic breast cancer in Chinese Han population of Northeast China. Breast Cancer Res Treat. 129:195–201.
- Jemal A, Bray F, Center MM, Ferlay J, Ward E, Forman D. 2011. Global cancer statistics. CA Cancer J Clin. 61:69–90.
- Keir ME, Butte MJ, Freeman GJ, Sharpe AH. 2008. PD-1 and its ligands in tolerance and immunity. Annu Rev Immunol. 26:677–704.
- Kumagai S, Togashi Y, Kamada T, Sugiyama E, Nishinakamura H, Takeuchi Y, Vitaly K, Itahashi K, Maeda Y, Matsui S, et al. 2020. The PD-1 expression balance between effector and regulatory T cells predicts the clinical efficacy of PD-1 blockade therapies. Nat Immunol. 21:1346–58.
- Kune GA, Vitetta L. 1992. Alcohol consumption and the etiology of colorectal cancer: a review of the scientific evidence from 1957 to 1991. Nutr Cancer. 18:97–111.
- Lee MS, Menter DG, Kopetz S. 2017. RIght versus left colon cancer biology: integrating the consensus molecular subtypes. J Natl Compr Canc Netw. 15:411–19.
- Li Z, Li N, Zhu Q, Zhang G, Han Q, Zhang P, Xun M, Wang Y, Zeng X, Yang C, et al. 2013. Genetic variations of PD1 and TIM3 are differentially and interactively associated with the development of cirrhosis and HCC in patients with chronic HBV infection. Infect Genet Evol. 14:240–46.
- Li Z, Zhang Z, He Z, Tang W, Li T, Zeng Z, He L, Shi Y. 2009. A partition-ligation-combination-subdivision EM algorithm for haplotype inference with multiallelic markers: update of the SHEsis (http://analysis.Bio-x.Cn). Cell Res. 19:519–23.
- Liu J, Tang W, Lin W, Wang Y, Chen Y, Wang J, Liu M, Xu W, Lin J, Zhang L. 2019. Lack of association between CTLA-4 genetic polymorphisms and noncardiac gastric cancer in a Chinese population. DNA Cell Biol. 38:443–48.
- Lund EK, Belshaw NJ, Elliott GO, Johnson IT. 2011. Recent advances in understanding the role of diet and obesity in the development of colorectal cancer. Proc Nutr Soc. 70:194–204.
- Marisa L, Svrcek M, Collura A, Becht E, Cervera P, Wanherdrick K, Buhard O, Goloudina A, Jonchere V, Selves J, et al. 2018. The balance between cytotoxic T-cell lymphocytes and immune checkpoint expression in the prognosis of colon tumors. J Natl Cancer Inst. 110:68–77.
- Mojtahedi Z, Mohmedi M, Rahimifar S, Erfani N, Hosseini SV, Ghaderi A. 2012. Programmed death-1 gene polymorphism (PD-1.5 C/T) is associated with colon cancer. Gene. 508:229–32.
- Nagai Y, Hata K, Kawai K, Murono K, Yasuda K, Otani K, Nishikawa T, Tanaka T, Kiyomatsu T, Nozawa H, et al. 2016. Clinicopathological features of colorectal cancer patients under the age of 50: recent experience and case-control study of prognosis in a Japanese cohort. Digestion. 93:272–79.
- Nagle CM, Wilson LF, Hughes MC, Ibiebele TI, Miura K, Bain CJ, Whiteman DC, Webb PM. 2015. Cancers in Australia in 2010 attributable to inadequate consumption of fruit, non-starchy vegetables and dietary fibre. Aust N Z J Public Health. 39:422–28.
- Oh SW, Kim YH, Choi YS, Chang DK, Son HJ, Rhee PL, Kim JJ, Rhee JC, Yun SH, Lee WY, et al. 2008. The comparison of the risk factors and clinical manifestations of proximal and distal colorectal cancer. Dis Colon Rectum. 51:56–61.
- Peng QH, Wang CH, Chen HM, Zhang RX, Pan ZZ, Lu ZH, Wang GY, Yue X, Huang W, Liu RY. 2021. CMTM6 and PD-L1 coexpression is associated with an active immune microenvironment and a favorable prognosis in colorectal cancer. J Immunother Cancer. 9:e001638.
- Qiu H, Wang Y, Kang M, Ding H, Liu C, Tang W, Xiao Z, Chen Y. 2017. The relationship between IGF2BP2 and PPARG polymorphisms and susceptibility to esophageal squamous-cell carcinomas in the eastern Chinese Han population. Onco Targets Ther. 10:5525–32.
- Qiu H, Xie Z, Tang W, Liu C, Wang Y, Gu H, Zheng Q. 2021. Association between microRNA-146a, -499a and -196a-2 SNPs and non-small cell lung cancer: a case-control study involving 2249 subjects. Biosci Rep. 41(2):BSR20201158.
- Qiu H, Zheng L, Tang W, Yin P, Cheng F, Wang L. 2014. Programmed death-1 (PD-1) polymorphisms in Chinese patients with esophageal cancer. Clin Biochem. 47:612–17.
- Riley JL. 2009. PD-1 signaling in primary T cells. Immunol Rev. 229:114–25.
- Savabkar S, Azimzadeh P, Chaleshi V, Nazemalhosseini Mojarad E, Aghdaei HA. 2013. Programmed death-1 gene polymorphism (PD-1.5 C/T) is associated with gastric cancer. Gastroenterol Hepatol Bed Bench. 6:178–82.
- Shi YY, He L. 2005. Shesis, a powerful software platform for analyses of linkage disequilibrium, haplotype construction, and genetic association at polymorphism loci. Cell Res. 15:97–98.
- Shirakami Y, Ohnishi M, Sakai H, Tanaka T, Shimizu M. 2017. Prevention of colorectal cancer by targeting obesity-related disorders and inflammation. Int J Mol Sci. 18:908.
- Siraj AK, Parvathareddy SK, Annaiyappanaidu P, Haqawi W, Al-Rasheed M, AlManea HM, AlHussaini HF, Al-Dayel F, Al-Kuraya KS. 2021. PD-L1 expression is associated with deficient mismatch repair and poor prognosis in middle eastern colorectal cancers. J Pers Med. 11:73.
- Tang W, Chen S, Chen Y, Lin J, Lin J, Wang Y, Liu C, Kang M. 2017. Programmed death-1 polymorphisms is associated with risk of esophagogastric junction adenocarcinoma in the Chinese Han population: a case-control study involving 2,740 subjects. Oncotarget. 8:39198–208.
- Tang W, Chen Y, Chen S, Sun B, Gu H, Kang M. 2015. Programmed death-1 (PD-1) polymorphism is associated with gastric cardia adenocarcinoma. Int J Clin Exp Med. 8:8086–93.
- Tang W, Chen S, Kang M, Liu J, Liu C. 2019. Investigation of BTLA tagging variants with risk of esophagogastric junction adenocarcinoma. Biosci Rep. 39(12):BSR20191770.
- Tang W, Zhang S, Qiu H, Wang L, Sun B, Yin J, Gu H. 2014. Genetic variations in MTHFR and esophageal squamous cell carcinoma susceptibility in Chinese Han population. Med Oncol. 31:915.
- Tiemersma EW, Kampman E, Bueno de Mesquita HB, Bunschoten A, van Schothorst EM, Kok FJ, Kromhout D. 2002. Meat consumption, cigarette smoking, and genetic susceptibility in the etiology of colorectal cancer: results from a Dutch prospective study. Cancer Causes Control: CCC. 13:383–93.
- Wang Y, Chen S, Kang M, Tang W, Gu H, Yin J, Huang Z. 2015. Genetic variations in MTHFR and gastric cardia adenocarcinoma susceptibility in the Chinese Han population. Int J Clin Exp Med. 8:18936–44.
- Wang L, Lo CH, He X, Hang D, Wang M, Wu K, Chan AT, Ogino S, Giovannucci EL, Song M. 2020. Risk factor profiles differ for cancers of different regions of the colorectum. Gastroenterology. 159:241–56 e213.
- Weitz J, Koch M, Debus J, Hohler T, Galle PR, Buchler MW. 2005. Colorectal cancer. Lancet. 365:153–65.
- Wielandt AM, Villarroel C, Hurtado C, Simian D, Zamorano D, Martinez M, Castro M, Vial MT, Kronberg U, Lopez-Kostner F. 2017. Characterization of patients with sporadic colorectal cancer following the new Consensus Molecular Subtypes (CMS). Rev Med Chil. 145:419–30.
- Xu M, Chen YM, Huang J, Fang YJ, Huang WQ, Yan B, Lu MS, Pan ZZ, Zhang CX. 2016. Flavonoid intake from vegetables and fruits is inversely associated with colorectal cancer risk: a case-control study in China. Br J Nutr. 116:1275–87.
- Yang J, Liu J, Chen Y, Tang W, Liu C, Sun Y, Chen J. 2019. Association of CTLA-4 tagging polymorphisms and haplotypes with hepatocellular carcinoma risk: a case-control study. Medicine. 98:e16266.
- Zang B, Chen C, Zhao JQ. 2020. PD-1 gene rs10204525 and rs7421861 polymorphisms are associated with increased risk and clinical features of esophageal cancer in a Chinese Han population. Aging. 12:3771–90.
- Zhai Y, Zhao WH, Chen CM. 2010. Verification on the cut-offs of waist circumference for defining central obesity in Chinese elderly and tall adults. Zhonghua Liu Xing Bing Xue Za Zhi = Zhonghua Liuxingbingxue Zazhi. 31:621–25.
- Zhang S, Chen S, Chen Y, Kang M, Liu C, Qiu H, Wang Y, Tang W. 2017. Investigation of methylenetetrahydrofolate reductase tagging polymorphisms with colorectal cancer in Chinese Han population. Oncotarget. 8:63518–27.
- Zhang S, Lin J, Jiang J, Chen Y, Tang W, Liu L. 2019. Association between methylenetetrahydrofolate reductase tagging polymorphisms and susceptibility of hepatocellular carcinoma: a case-control study. Biosci Rep. 39(11):BSR20192517.
- Zhang X, Zhang S, Li Y, Detrano RC, Chen K, Li X, Zhao L, Benjamin EJ, Wu Y. 2009. Association of obesity and atrial fibrillation among middle-aged and elderly Chinese. Int J Obes. 33:1318–25.
- Zhang J, Zhao T, Xu C, Huang J, Yu H. 2016. The association between polymorphisms in the PDCD1 gene and the risk of cancer: a PRISMA-compliant meta-analysis. Medicine. 95:e4423.
- Zhao Y, Mao Z, Pang H, Zhao X, Zhang S, Gao Z, Yang Y, Fang T, Ma Q, Ma X, et al. 2018. Association of programmed cell death 1 (PDCD1) gene polymorphisms with colorectal cancer among Han Chinese population. Zhonghua Yi Xue Yi Chuan Xue Za Zhi = Zhonghua Yixue Yichuanxue Zazhi = Chinese Journal of Medical Genetics. 35:219–23.
- Zhou RM, Li Y, Wang N, Huang X, Cao SR, Shan BE. 2016. Association of programmed death-1 polymorphisms with the risk and prognosis of esophageal squamous cell carcinoma. Cancer Genet. 209:365–75.
- Zou C, Qiu H, Tang W, Wang Y, Lan B, Chen Y. 2018. CTLA4 tagging polymorphisms and risk of colorectal cancer: a case-control study involving 2,306 subjects. Onco Targets Ther. 11:4609–19.