
ABSTRACT
In this study, we describe the dental remains of a very minute beaver from the early Late Miocene locality Hammerschmiede (MN 7/8) located at the Northern Alpine Foreland Basin (Southern Germany, Bavaria). The finds represent the so far most comprehensive (>1.000 dental specimens) collection of the trogontheriine castorid Euroxenomys minutus. Metrically and morphologically, the large data set demonstrates an extensive intraspecific variability of this beaver from Hammerschmiede that covers most finds of other European localities and confirms previous species assignments. The subspecies Euroxenomys minutus rhenanus from Dorn-Dürkheim (Germany), clearly isolates from this range of variation and can thus be validated. The performed age-frequency distributions for Euroxenomys minutus from the local stratigraphic levels HAM 5 (rivulet) and HAM 4 (river) correspond roughly and also resemble Euroxenomys from the swamp deposit Rudabánya (Hungary). Since previous mortality studies on the also occurring larger beaver Steneofiber depereti from Hammerschmiede demonstrated dominance of behavioural or internal factors, this cross-ecosystem similarity in mortality for the small beaver species is likely explained by external factors, such as predation. Supported by a taphonomic bone analysis, a multitude of predators are finally considered to prey on Euroxenomys minutus and indicate a predatory bone assemblage.
Introduction
For almost half a century now, the Hammerschmiede fossil site has been providing new scientific discoveries (Fahlbusch and Mayr Citation1975; Mayr and Fahlbusch Citation1975). At least seven fossil-bearing strata are known to date, with most of the finds coming from the local stratigraphic levels HAM 5 and HAM 4, dated to 11.62 and 11.44 Ma, respectively, by Kirscher et al. (Citation2016). Since new excavation efforts by the University of Tübingen began in the early 2000s, the constantly growing amount of material available has increased to more than 30,000 specimens, which comprise at least 146 different vertebrate taxa. The still ongoing excavations continue to yield new discoveries year after year, which contribute to our knowledge on these unique early Late Miocene ecosystems in Central Europe. The most famous discovery of the Hammerschmiede is for sure the arboreal biped hominid Danuvius guggenmosi Böhme et al. (Citation2019), which attracted international interest (Böhme et al. Citation2019, Citation2020; Williams et al. Citation2020). Many different vertebrates from Hammerschmiede have already been subject of scientific research, including carnivores (Kargopoulos et al. Citation2021a, Citation2021b, Citation2021c, Citation2022), artiodactyles (Fuss et al. Citation2015; Hartung et al. Citation2020, Citation2022), small mammals (Mayr and Fahlbusch Citation1975; Prieto Citation2009, Citation2012; Prieto et al. Citation2011; Prieto and van Dam Citation2012) and birds (Mayr et al. Citation2020a, Citation2020b, Citation2022). A preliminary taxa list of the Hammerschmiede fauna was published by Kirscher et al. (Citation2016) and updated by Böhme et al. (Citation2019).
Beavers have also been included in scientific studies on the Hammerschmiede fauna. Mayr and Fahlbusch (Citation1975) have already mentioned the occurrence of two beaver species, a small form ‘Steneofiber minutus’ = Euroxenomys minutus (von Meyer Citation1838) and a large beaver species ‘Steneofiber jaegeri’ = Chalicomys jaegeri Kaup (Citation1832) were mentioned by a small number of teeth and fragments. The large form, referred to a single incisor fragment, was further listed by Hugueney (Citation1999), Kirscher et al. (Citation2016) and Böhme et al. (Citation2019) as Chalicomys jaegeri Kaup (Citation1832). Recently, Lechner and Böhme (Citation2022) assigned the now significantly more extensive material collection of this large beaver to the species of Steneofiber depereti Mayet (Citation1908). The small beaver form from Hammerschmiede was already correctly recognised by several authors as one of the various synonymous combinations like Trogontherium (Euroxenomys) minutum in Hugueney (Citation1999) and Euroxenomys minutus in Kirscher et al. (Citation2016), Böhme et al. (Citation2019) and Lechner and Böhme (Citation2022). Here, we follow the latter in their assignment to the combination Euroxenomys minutus (von Meyer Citation1838).
Euroxenomys represents with much over 1.000 dental and at least as many postcranial specimens a very abundant rodent in Hammerschmiede, being next to Anomalomys gaudryi (Anomalomyidae) the second most common small mammal species after Microcricetus molassicus (Cricetidae).
The current beaver biodiversity is limited to one genus including two species, whereas the family of Castoridae was much more diverse during the Miocene time period. The European Neogene beaver record is very rich and contains up to seven genera, Anchitheriomys Roger (Citation1898), Chalicomys Kaup (Citation1832), Dipoides Jäger (Citation1835), Eucastor? (Schreuderia) Aldana Carrasco (Citation1992), Euroxenomys Samson and Rădulesco (Citation1973), Steneofiber Geoffroy-Saint-Hilaire (Citation1833) and Trogontherium Fischer von Waldheim (Citation1809) (Hugueney Citation1999; Stefen Citation2009). There are localities with only one beaver species recorded and others, containing a higher beaver diversity (Hugueney Citation1999; Rekovets et al. Citation2020). It is quite rare that several large beavers occur simultaneously in one site, whereas the combination of one large beaver like Steneofiber or Chalicomys and the minute beaver Euroxenomys minutus is quite typical (Hugueney Citation1999).
The biology and ecology of fossil representatives of the Steneofiber-Chalicomys-Castor lineage can probably be considered quite similar to those of present-day castorids. Physiognomy with excellent swimming and burrowing ability and behavioural strategies or ecological parameters seem to differ only marginally from today’s beavers (Schreuder Citation1929; Hugueney and Escuillié Citation1995, Citation1996; Lechner and Böhme Citation2020, Citation2022). The knowledge about the ecology of the small beaver Euroxenomys minutus is much more limited and therefore it is not fully understood how coexistence with large beaver species is possible. It seems clear that the small beaver deviates from the typical beaver construction plan simply because of its small body size (size of a muskrat), but also other physiognomic differences (no wide and flattened tail; different long-bone proportions) and thus is likely to represent a different ecology, possibly comparable to the extant muskrat – Ondatra zibethicus (Stefen and Rummel Citation2003; Daxner-Höck Citation2004). We know from present-day beavers that they can easily exist sympatrically in the same habitats with the muskrat, which is also semiaquatic (Mott et al. Citation2013). For a deeper palaeoecological analysis, in addition to dental investigations, profound functional-morphological analyses of the entire skeleton must of course be carried out. Even though especially the long bones of the extremities are often fragmented and broken, more than 1000 pieces are available for such investigations. The postcranial fundus will be the subject of future investigations and is not part of the present study.
The aim of our study is to describe the large amount of dental material of the minute beaver Euroxenomys minutus from Hammerschmiede and compare with other finds of European localities. Since the Hammerschmiede material probably represents the most extensive collection of material of this fossil beaver species, a unique insight into its intraspecific variability can be gained here. Furthermore, the material is analysed taphonomically to find potential traces of predation. Finally, mortality analyses based on dental wear stages are created that can provide further evidence for the ecological classification of this small beaver and makes it possible to compare it with other beaver species from similar and different fossil habitats.
Geological setting
The Hammerschmiede fossil site is an actively operated clay mine near the small village of Pforzen (Bavaria, Southern Germany). The 26-metre-thick profile of the clay pit exhibits fluvio-alluvial flood plain deposits (clay, silt, fine-sand, lignite) of the Northern Alpine Foreland Basin. The two main fossiliferous levels HAM 5 and HAM 4 represent early Late Miocene (base of Tortonian, MN 7/8) channel fillings with very fossil enriched channel lag deposits and have been dated to 11.62 and 11.44 Myr, respectively (Kirscher et al. Citation2016). The older HAM 5 is assumed to represent a rivulet of approximately 5 m width, whereas HAM 4 corresponds to a larger river of about 50 m width and 4–5 m depth (Mayr et al. Citation2020a; Lechner and Böhme Citation2022).
Material and methods
The studied specimens were collected from the local fossil layers HAM 5 and HAM 4 of the Hammerschmiede site over the period from 2011 to 2021 (later collected specimens are not included). Since 2016, all excavated sediments of the find layers are screened for small finds (>1 mm) by means of screen washing and rotational sieving system. However, a large part of the sediments from the grain size range 1–5 mm has not yet been sorted out, so that a statistical distribution of tooth sizes of small teeth (last molars) to large teeth (premolars) is probably statistically biased. In total, 1115 teeth (981 specimens) were examined, of which 465 (452 specimens: 10 tooth row specimens, 442 isolated teeth) are from the HAM 5 layer and 650 (529 specimens: 75 tooth row specimens, 454 isolated teeth) from the HAM 4 layer.
The entire material is deposited in the palaeontological collection of the University of Tübingen, Germany (GPIT), and is labelled either with GPIT (for excavation years 2011 to 2019 inclusive) or SNSB-BSPG (Bavarian State Collection of Palaeontology and Geology in Munich, Germany; for excavation years 2020 to 2022). SNSB-BSPG 2020 XCIV encodes the inventory from the layer HAM 4 and SNSB-BSPG 2020 XCV from the layer HAM 5.
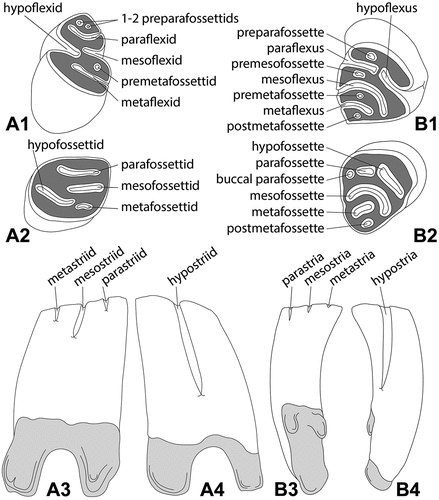
The morphological nomenclature of dental material follows Stirton (Citation1935) and Hugueney (Citation1999) (). The nomenclature of skull and mandibular features follows Freye (Citation1959) and Stefen and Rummel (Citation2003).
Figure 1. Morphological nomenclature and the general tooth scheme of left lower (A) and right upper (B) cheek teeth (premolars and molars) used for the descriptions and comparisons of the minute beaver Euroxenomys minutus (von Meyer Citation1838), from the early Late Miocene locality Hammerschmiede (Bavaria, Germany). (A1) and (B1), occlusal views of early wear stages; (A2) and (B2), occlusal views of later wear stages; (A3) and (B4), lingual views; (A4) and (B3), buccal views. Enamel in white, dentine in dark grey, roots in light grey. Line drawings are not based on actual specimens and are not to scale. Nomenclature follows Stirton (Citation1935) and Hugueney (Citation1999).
Measurements were taken with a digital calliper (rounded to the first decimal point). The maximum mesio-distal length (L) and bucco-lingual width (W) of cheek teeth were measured at the occlusal surface (occlusal measurements) and at the basal tooth crown position (basal measurements) where possible. In the upper and lower incisors, the length (L) corresponds to the antero-posterior and the width (W) to the mesio-distal extent.
Crown height measurements of lower premolars (p4) follow Kordos (Citation2020) and have been taken at the position of the buccal hypostriid: (1) from the occlusal surface of the tooth to the base of the hypoflexid and (2) from the base of the hypoflexid to the base of the tooth crown. Both measurements were added to determine a tooth crown height.
The evaluation of dental wear stages (WS) used for tooth descriptions are modified according to Stefen (Citation1997, Citation2001, Citation2018), Stefen and Mörs (Citation2008), Heinrich and Maul (Citation2020) and Lechner and Böhme (Citation2022): (1) unworn: no wear can be observed, deciduous dentition in use; (2) slightly worn: first clear occlusal contact; (3) worn: para-/metaflexus/-id is closing or closed and mesoflexus/-id in most cases open; (4) medium worn: mesoflexus/-id is closing or just closed; (5) deeply worn: hypoflexus/-id is near to closing; (6) heavily worn: hypoflexus/-id is closed. Due to the very short lingual striids in the available material from Hammerschmiede, the wear stages from unworn to worn teeth represent approximately the first quarter of tooth crown height and therefore these wear stages defined on morphological criteria cannot be used for evenly graded age estimation and thus for a mortality analysis.
The dental wear stages for the preparation of mortality analyses were taken at the lower premolar tooth position and combine morphological and metric data. The first two wear stages are morphologically determined age stages defined by unused (WS 1, deciduous tooth in use) and first and slightly used lower p4 (WS 2, milk tooth replaced). During these two wear stages, the tooth crown of the lower p4 is not fully grown and the crown height cannot be measured. For the remaining p4 material (> WS 2), the wear stages are calculated based on the crown height. The difference between the largest and the smallest measured tooth crown height is divided into four evenly distributed size ranges and the teeth are assigned to these size categories as WS 3 to WS 6 according to their measured value. Due to the high metric variability within the existing tooth material, it did not seem reasonable to define more than 4 wear stages based on measurements.
Systematic palaeontology
Family Castoridae Hemprich (Citation1820)
Genus Euroxenomys Samson and Rădulesco (Citation1973)
Comment
The generic assignment of Euroxenomys is still under discussion. In some studies, Euroxenomys is classified at the subgenus level (e.g. Hugueney Citation1999; Stefen and Rummel Citation2003; Giersch et al. Citation2010; Kordos Citation2020) or as a valid genus (e.g. Samson and Rădulesco Citation1973; Korth Citation2001; Hugueney and Duranthon Citation2012; Prieto et al. Citation2014; Rekovets et al. Citation2020; Mörs et al. Citation2022).
Here, we follow Hugueney and Duranthon (Citation2012) and place Euroxenomys at generic level.
Type species
Chalicomys minutus von Meyer (Citation1838)
Type locality
Elgg (Switzerland), Middle Miocene.
Euroxenomys minutus (von Meyer Citation1838)
Subspecies of Euroxenomys: E. minutus minutus (von Meyer Citation1838), E. minutus rhenanus (Franzen and Storch Citation1975)
Euroxenomys minutus minutus (von Meyer Citation1838)
(; Measurements see , and Table 2 supplement)
Figure 2. Cranial fragments of the minute beaver Euroxenomys minutus (von Meyer Citation1838), from the early Late Miocene locality Hammerschmiede (Bavaria, Germany), local stratigraphic level HAM 4. (A) SNSB-BSPG 2020 XCIV-9124, maxillae and fragmentary palatine with left P4 and right P4-M1 in left buccal (A1), mesial (A2) and occlusal (A3) views. (B) SNSB-BSPG 2020 XCIV-9523, maxillae and fragmentary palatine without dentition but with preserved alveoli of left P4-M3 and right P4-M2 in dorsal (B1) and occlusal (B2) views. (C) SNSB-BSPG 2020 XCIV-9522, right maxilla and fragmentary palatine with right P4-M2 in occlusal (c1) and right buccal (C2) views. Scale bar equals 10 mm.
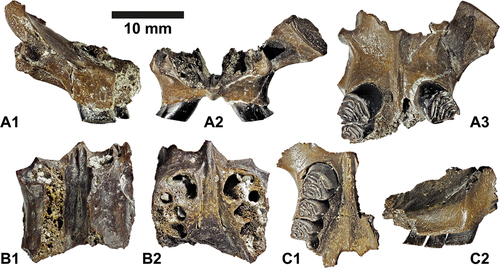
Figure 3. Upper cheek teeth of the minute beaver Euroxenomys minutus (von Meyer Citation1838), from the early Late Miocene locality Hammerschmiede (Bavaria, Germany), local stratigraphic levels HAM 5 and HAM 4. Deciduous premolars: (A-C); premolars (D-G); molars (H-P). Occlusal (A, B, C1, D1, E, F, G, H1, I1, K, L, M1, N1, O, P1), lingual (D3, H2, I3, M2, N2, P3), buccal (D2, I2, N3) and apical (P2) views. Scale bar equals 10 mm. Left DP4: (A) GPIT/MA/12001; (C) GPIT/MA/12058. Right DP4: (B) GPIT/MA/19158. Left P4: (G) GPIT/MA/18783. Right P4: (D) GPIT/MA/19060; (E) GPIT/MA/18780; (F) GPIT/MA/18835. Right M1/2: (H) GPIT/MA/18866; (I) GPIT/MA/18872; (K) SNSB-BSPG 2020 XCIV-6959; (L) GPIT/MA/19131. Left M3: (N) GPIT/MA/19088; (P) GPIT/MA/19127. Right M3: (M) SNSB-BSPG 2020 XCIV-6949; (O) SNSB-BSPG 2020 XCIV-2170.

Figure 4. Mandibles of the minute beaver Euroxenomys minutus (von Meyer Citation1838), from the early Late Miocene locality Hammerschmiede (Bavaria, Germany), local stratigraphic levels HAM 5 and HAM 4. Scale bar equals 10 mm. (A) GPIT/MA/12168, right mandible with angular process, part of the coronoid process, i2, p4-m2 in buccal (A1), lingual (A2) and occlusal (A3) views. (B) GPIT/MA/17127, right mandible with i2 in buccal view. (C) GPIT/MA/16523, left mandible with broken i2 and complete cheek toothrow p4-m3 in lingual (C1), buccal (C2) and occlusal (C3) views. (D) SNSB-BSPG 2020 XCIV-5287, left mandible fragment with complete cheek toothrow p4-m3 in occlusal (D1) and buccal (D2) views.

Figure 5. Lower cheek teeth of the minute beaver Euroxenomys minutus (von Meyer Citation1838), from the early Late Miocene locality Hammerschmiede (Bavaria, Germany), local stratigraphic levels HAM 5 and HAM 4. Deciduous premolars: (A-E); premolars (F-M); molars (N-X). Occlusal (A, B, C, D, E1, F G1, H, I1, K, L, M1, N1, O, P, Q1, R1, S1, T1, U1, V1, W, X1), lingual (E3, G3, I4, N2, Q2, R3, S2, T3, U2, V2, X4), buccal (E2, G2, I2, M2, N3, Q3, R2, S3, T2, U3, V3, X3), mesial (I3) and apical (X2) views. Scale bar equals 10 mm. Left dp4: (C) GPIT/MA/11999. Right dp4: (A) GPIT/MA/19166; (B) GPIT/MA/19095; (D) GPIT/MA/19093; (E) GPIT/MA/18988. Left p4: (H) GPIT/MA/18699; (L) SNSB-BSPG 2020 XCIV-6894. Right p4: (F) GPIT/MA/18678; (G) SNSB-BSPG 2020 XCIV-6889; (I) GPIT/MA/18732; (K) GPIT/MA/18676; (M) GPIT/MA/18725. Left m1/2: (N) GPIT/MA/19150; (Q) GPIT/MA/18964; (S) GPIT/MA/19021. Right m1/2: (O) GPIT/MA/19023; (P) GPIT/MA/19019; (R) SNSB-BSPG 2020 XCIV-7037. Left m3: (U) GPIT/MA/18982; (V) SNSB-BSPG 2020 XCIV-6983. Right m3: (T) SNSB-BSPG 2020 XCIV-6985; (W) GPIT/MA/19086; (X) GPIT/MA/19122.

Figure 6. Length/width dimensions of upper premolars of the minute beaver Euroxenomys minutus (von Meyer Citation1838), from the early Late Miocene locality Hammerschmiede (Bavaria, Germany), local stratigraphic levels HAM 5 and HAM 4, compared to Euroxenomys material from other Eurasian Miocene localities. Measurements for Hammerschmiede (HAM), Rudabánya and Dorn-Dürkheim additionally distinguish occlusal and basal tooth measurements while the other sites only include occlusal values. Data for Rudabánya from L. Kordos personal communication, for Dorn-Dürkheim own measurements, for Sansan from Hugueney and Duranthon (Citation2012), for Höwenegg from Giersch et al. (Citation2010), for Petersbuch 50 from Stefen and Rummel (Citation2003), for Anwil from Engesser (Citation1972), for Gratkorn from Prieto et al. (Citation2014), for Tagay from Mörs et al. (Citation2022), for Atzelsdorf from Daxner-Höck and Bernor (Citation2009) and for Felsötárkány from Hír (Citation2004)
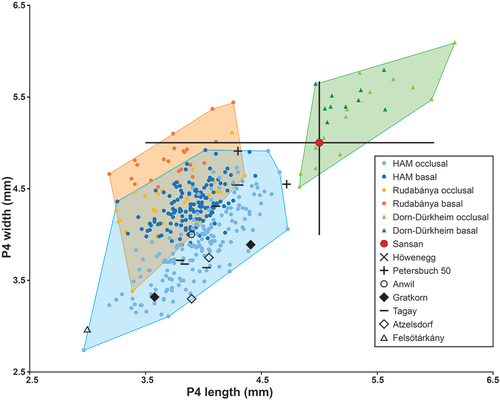
Figure 7. Length/width dimensions of lower premolars of the minute beaver Euroxenomys minutus (von Meyer Citation1838), from the early Late Miocene locality Hammerschmiede (Bavaria, Germany), local stratigraphic levels HAM 5 and HAM 4, compared to Euroxenomys material from other Eurasian Miocene localities. Measurements for Hammerschmiede (HAM), Rudabánya and Dorn-Dürkheim additionally distinguish occlusal and basal tooth measurements while the other sites only include occlusal values. Data for Rudabánya from L. Kordos personal communication, for Dorn-Dürkheim own measurements, for Sansan and Elgg from Hugueney and Duranthon (Citation2012), for Höwenegg from Giersch et al. (Citation2010), for Petersbuch 50 from Stefen and Rummel (Citation2003), for Anwil from Engesser (Citation1972), for Gratkorn from Prieto et al. (Citation2014), for Tagay from Mörs et al. (Citation2022), for Atzelsdorf from Daxner-Höck and Bernor (Citation2009) and for Felsötárkány from Hír (Citation2004)
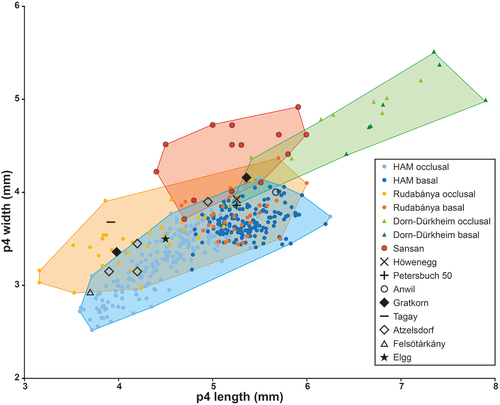
Table 1. Dimensions of upper and lower teeth of the minute beaver Euroxenomys minutus (von Meyer Citation1838), from the early Late Miocene locality Hammerschmiede (Bavaria, Germany), with combined treatment of the material from the local stratigraphic levels HAM 5 and HAM 4. L, mesio-distal length at occlusal surface and at basal crown position (where possible) for cheek teeth and length across anterior enamel band for incisors; W, bucco-lingual width at occlusal surface and at basal crown position (where possible) for cheek teeth; m, measurement; n, number of measurements (sum of occlusal and basal values); All measurements in mm.
Material
Hammerschmiede locality, Germany, early Late Miocene, base of Tortonian, MN 7/8. For measurements see and Table 2 supplement. The material comprises a total of 1115 teeth (981 specimens: 896 isolated teeth; 32 jaws with in situ dentition) from layer HAM 5 and HAM 4 together. The HAM 5 material includes 465 teeth (452 specimens: 442 isolated teeth; 4 jaws with in situ dentition) and the HAM 4 material includes 650 teeth (529 specimens: 454 isolated teeth; 28 jaws with in situ dentition). HAM 5: 24 i2, 34 dp4, 68 p4, 84 m1/2, 21 m3, 18 I2, 17 DP4, 67 P4, 91 M1/2, 37 M3; HAM 4: 89 i2, 9 dp4, 143 p4, 130 m1/2, 19 m3, 45 I2, 6 DP4, 115 P4, 72 M1/2, 22 M3. The MNI (minimum number of individuals) based on the maximum number of a tooth position on one side is 35 for HAM 5 (based on right lower p4 and left upper P4) and 82 for Ham 4 (right lower p4).
Diagnosis
See Prieto et al. (Citation2014) and emended diagnosis therein. Furthermore, see diagnosis in Samson and Rădulesco (Citation1973) for the then assumed monospecific genus Euroxenomys and see Hugueney (Citation1999), Daxner-Höck and Bernor (Citation2009) and Kordos (Citation2020) for detailed dental descriptions of Euroxenomys minutus.
Description
()
Maxilla and palatine ()
There are several cranial specimens of Euroxenomys minutus available from Hammerschmiede, mostly comprising fragments of the toothrow or maxilla and palatine. The description of maxilla and palatine follows the rather complete specimens (SNSB-BSPG 2020 XCIV-9124, 9522, 9523). In ventral view, the tooth rows diverge from anterior to distal (Fig. 2A3, B2, C1). The palatine shows a small anterior palatine foramen and a slightly larger lateral distal palatine foramen is fragmentary preserved in SNSB-BSPG 2020 XCIV-9522 (Fig. 2C1). The distal end of the palatine is medially trapezoidal carved without an aboral nasal spine as it typically occurs with Castor. The anterior palatine suture separates the maxillaries almost in a straight line on between M1 and M2. Maxillar grooves start at the anterior palatal foramen and extend slightly anterior to P4 enclosing a medially aligned palatine sulcus. A laterally very widely projecting zygomatic process is fragmentarily indicated (Fig. 2A2).
I2
Upper incisors are robust, rounded triangular and show a smooth and convex anterior enamel surface, a convex to nearly straight mesial and a convex rounded distal tooth surface. The mesial and distal tooth surfaces may show slight longitudinal striations of irregular arrangement (GPIT/MA/19176, 19177). In contrast to the lower incisors, the wear area in upper I2 is short and the anterior enamel edge is straight. There are two wear facets, one runs evenly diagonally over the distal wear area, whereas the other dips steeply into the dentin along the anterior enamel band and results in a mesial depression in which the tip of the lower i2 can insert (GPIT/MA/19177), as in the modern muskrat Ondatra zibethicus.
DP4 ()
The upper deciduous premolar is very short crowned (brachydont) and exhibits in unused specimens a rather rectangular, with increasing wear rather triangular occlusal outline. The buccal and posterior tooth margins are straight and the anterolingual tooth wall is convex rounded. The buccal side of the DP4 is clearly lower crowned than the lingual one. The hypoflexus is open in all available specimens and the hypostria almost reaches the tooth crown base. Only in unworn or slightly worn DP4, the buccal para-, meso- and metaflexus/-stria is open. Most specimens show moderate wear stages with all of the buccal flexus/striae close to the fossettes. The long and straight paraflexus (GPIT/MA/12002, unworn) is oriented posterolingually and meets straight in line with the obliquely oriented hypoflexus. The mesoflexus is long and bent, runs across the entire tooth width and ends at the posterolingual corner. In barely worn specimens, the mesoflexus can be connected to the metaflexus/-fossette. In more worn DP4, the long mesofossette spans the small round metafossette, that is placed at the posterobuccal edge (GPIT/MA/19158). The upper deciduous premolar is three-rooted with one large transverse lingual and two equally small and circular buccal roots at the anterolingual and posterolingual corners (GPIT/MA/12001, GPIT/MA/12058).
P4 ()
The upper P4 is very high-crowned and represents the largest tooth of the upper tooth row. In most of the specimens (unworn to higher worn P4) the occlusal outline is triangular with convex anterolingual and buccal sides. In very heavily worn specimens, also the buccal side can be straight or slightly concave (GPIT/MA/18783). The straight posterior tooth wall together with the buccal side forms a posterobuccal apex. The entire tooth crown is bent diagonally towards posterobuccal resulting in a very high anterolingual and a shorter posterobuccal crown. The lingual hypoflexus/-fossette, and the buccal para-, meso- and metaflexus/-fossette are general occlusal elements of the P4. The paraflexus/-fossette is straight, oriented posterolingually and meets in straight line the equally sized hypoflexus/-fossette, which is directed oblique anterobuccally. In heavily worn upper premolars the hypofossette is short, protrudes slightly between para- and mesofossette (GPIT/MA/18783, 18835) and can be angled posteriorly (GPIT/MA/18783).
The mesoflexus/-fossette is the longest occlusal element, can be less, more or heavily convex and ends at the posterior margin of the P4. Similar to the DP4 the short metaflexus/-fossette is straight or very slightly curved and enclosed by the mesoflexus/-fossette at the posterobuccal corner. A wide range of variations within the occlusal elements of the extensive material can be found: a divided parafossette (GPIT/MA/19066), an additional preparafossette (GPIT/MA/19059, SNSB-BSPG 2020 XCIV-7000) or a small premetafossette at the posterior margin between the meso- and the metafossette (GPIT/MA/16866). In SNSB-BSPG 2020 XCIV-7001, the meta- and mesofossette are fused resulting in a U-shape and in the special case of SNSB-BSPG 2020 XCIV-7025 an additional elongated fossette is placed and oriented perpendicular between the ends of the hypo- and parafossette, in anterodistal orientation.
The hypostria is the longest of all striae and in unworn or slightly worn P4 (GPIT/MA/19060), it runs down half of the tooth crown, closing at high wear stages. The parastria is the longest buccal stria closing within the first quarter of the tooth-crown at moderate dental wear stages. The mesostria, in general the second shortest stria, closes within the first millimetre of tooth wear (assessed on unworn teeth). A very short metastria/-flexus is only present in unworn or very slightly worn P4 (GPIT/MA/12436, 16191, 17302, 19055, 19056, SNSB-BSPG 2020 XCIV-3449, 7004, 7017, 7023). In most of the material the metastria is the shortest stria but in two worn P4, the mesostria is already closed while para- and metastria are still open (GPIT/MA/19052, SNSB-BSPG 2020 XCIV-7008). In a single case is the mesostria the longest and only buccal stria, while para- and metastria are already closed (GPIT/MA/19066). Upper premolars show two heteromorphic roots, one massive that follows the anterolingual outline of the tooth in a crescent shape and one short, small circular and columnar shaped posterobuccal root.
M1/2 ()
In general, the upper first and second molars occlusally show high morphological similarities with upper premolars. Deviating from this, upper M1/2 are smaller and more square in occlusal outline. With wear, the mesio-distal length gets shorter, while the width remains approximately the same (rectangular outline). The buccal and posterior tooth walls are straight and oriented perpendicular to each other, while the anterior and lingual ones are more rounded. The tooth crowns of the M1/2 are bent diagonally so that the posterobuccal corner slightly overhangs. Very young dental ages (unworn or slightly worn) of M1/2 show all typical occlusal elements of the P4. The lingual hypoflexus/-fossette is long, straight and oriented oblique in anterobuccal direction, crossing half of the tooth width. The paraflexus/-fossette is short, straight or slightly bent posteriorly and ends slightly directing posteriorly to the end of the hypoflexus/-fossette. Like with the P4, the mesoflexus/-fossette is the longest occlusal element. It is slightly, more or heavily curved posteriorly and ends at the posterior margin of the M1/2. Similar to DP4 and P4 the short metaflexus/-fossette is straight or very slightly curved and enclosed by the mesoflexus/-fossette at the posterobuccal corner. The lingual hypostria runs down half of the tooth crown, being the longest stria. Meso- and parastria correspond to each other in length but are very short. There are slightly worn M1/2 that show a buccally open para- and mesoflexus (GPIT/MA/18866, 18872, 19121) and in two worn specimens only the mesoflexus/-stria is open at the buccal tooth wall (GPIT/MA/12057, 19136). The metastria is the shortest of all striae – if existing at all. There are two specimens representing slightly worn M1/2, that show all four flexus/striae in an open phase (hypo-, para-, meso- and metaflexus/-stria; GPIT/MA/19009, 19010). In highly worn upper first and second molars, the hypoflexus/-stria is closed (GPIT/MA/12028, 12030, 18863, 18896, 18909, 19007, 19120, SNSB-BSPG 2020 XCIV-6962). M1/2 of very high wear stages show only occlusal fossettes and in two cases the metafossette is worn down and missing (GPIT/MA/18863, 19120). Upper M1/2 are three-rooted with one large and massive root that follows in a crescent shape the anterolingual margin of the tooth wall and two short, small and round roots located at the anterobuccal and posterobuccal corners.
M3 ()
Upper third molars are triangular in occlusal outline and highly elongated in anterodistal direction. The convex anterior face marks the greatest tooth width and the straight buccal and posterolingual faces merge in a distally pointed and round corner. The tooth crown of the M3 is slightly bent and overhanging at the posterobuccal tip. Furthermore, the anterolingual margin is higher crowned than the posterobuccal one. In early dental wear stages, the M3 is rather rectangular and shows a weak distal elongation, which strengthens and lengthens with higher wear stages. With wear, mesio-distal length gets longer, while the width remains approximately the same. As a basic pattern, a lingual hypoflexus/-fossette and the buccal para-, meso- and metaflexus/-fossette can be observed. The straight anterobuccally oriented hypoflexus/-fossette terminates at median tooth width anteriorly to the paraflexus/-fossette. Mesoflexus/-fossette in the upper third molars, as in all other upper cheek teeth, is more or less curved or angled posteriorly and encloses the metaflexus/-fossette anteriorly. Para- and metaflexus/-fossette are equally small, shorter than the mesoflexus/-fossette and display marked variability in size, orientation and shape. Occurring occlusal variations are: a split in two parafossettes (or additional, a small and circular buccal parafossette; GPIT/MA/18752); an additional premesofossette (GPIT/MA/19124); an additional small and circular fossette lingually (GPIT/MA/18753, 19124) or posteriorly to the metafossette (postmetafossette: GPIT/MA/19125, 19126); a straight buccolingually oriented (SNSB-BSPG 2020 XCIV-6949), an anteroposteriorly oriented (GPIT/MA/19127), a posteriorly inverted U-shaped (GPIT/MA/19124) or a Y-shaped metafossete (GPIT/MA/19125). The hypostriid is the longest striid closing at very high wear stages (GPIT/MA/18760, 19127, SNSB-BSPG 2020 XCIV-4606, 6956). GPIT/MA/18997 is the only M3 showing three open buccal striae (slightly worn) but all very short. In other cases, the parastria is closed while meso- and metastria are open (worn; GPIT/MA/18995, SNSB-BSPG 2020 XCIV-6954); the para- and mesostria are open, while the metastria is closed (slightly worn or worn; GPIT/MA/18999, 19,124, SNSB-BSPG 2020 XCIV-6953), or only the mesostria is open buccally (slightly worn or worn; GPIT/MA/18752, 18753, SNSB-BSPG 2020 XCIV-6955). The third upper molar shows two or three roots. A marked crescent-shaped root runs along the anterior tooth margin and a slightly smaller and round root is placed under the posterior tooth tip. A third, very small root may occur adjacent to the anterolingual margin of the former main root (GPIT/MA/19088) or is somewhat more pronounced and shifted further into the lingual centre of the tooth (GPIT/MA/19127).
Mandible ()
There are several mandibular specimens of Euroxenomys minutus available from Hammerschmiede mostly comprising fragments of the toothrow or anterior portion. GPIT/MA/12168 represents the most complete specimen with the ramus and angular process preserved but crushed and slightly displaced. Furthermore, the description of mandibles follows the rather complete specimens (GPIT/MA/16523, 17127, SNSB-BSPG 2020 XCIV-0310, 4551, 5287, 5289). The tooth row lengths show values between 13.38 and 14.59 mm (SNSB-BSPG 2020 XCIV-5287, 13.38 mm; GPIT/MA/17444, 13.4 mm; GPIT/MA/17700, 13.61 mm; GPIT/MA/16523, 14.1 mm; SNSB-BSPG 2020 XCIV-4551, 14.58 mm; SNSB-BSPG 2020 XCIV-0310, 14.59 mm). The alveolar length of the mandibular toothrow is between 14.36 and 15.62 mm (SNSB-BSPG 2020 XCIV-5287, 14.36; SNSB-BSPG 2020 XCIV-4551, 14.42 mm; GPIT/MA/16523, 14.77 mm; SNSB-BSPG 2020 XCIV-0310, 14.99 mm; GPIT/MA/17444, 15.09 mm; GPIT/MA/17127, 15.12; GPIT/MA/16521, 15.22 mm; GPIT/MA/17419, 15.25 mm; GPIT/MA/17700, 15.62 mm).
Broken margins of the ventral side of the posterior portion of the mandibles indicate a large pterygoid fossa with a wide shelf. On the buccal side, the masseteric fossa is marked and placed at the level of the occlusal surface of the toothrow in superposition to the posterior end of the incisor, between the coronoid process and the angular process. There is a distinct groove at the anterior ascending ramus that runs anteroventrally until it terminates at a shallow and tiny punctual depression, arranged in line with the buccal hypostriid of the lower premolar and placed slightly occlusal to the foramen mentale. This depression is the origin of another groove that is running posteriorly at a more ventral position and terminates at about the level of the posterior m3.
In buccal view, the small chin process is in line with the foramen mentale, shortly anterior to the premolar. The diastema is posteriorly asymmetrical and reaches approximately the length of the occlusal p4-m2 (10 mm). The posterior end of the lingual symphysis is placed below the anterior margin of the m1.
i2
Lower incisors are robust, show a smooth and convex anterior enamel surface, a convex distal and a straight mesial tooth surface that roughly describe a triangular cross-section. The wear area is elongated (longer than in the upper I2) and separated into a slightly mesially and a slightly distally angled wear facet, terminating in a central tooth tip, as in the modern muskrat Ondatra zibethicus.
dp4 ()
The lower deciduous premolar corresponds morphologically to a brachydont miniature of the permanent lower premolar. Basic elements include a buccal hypoflexid with a hypostriid that does not reach the crown base, a parafossettid (closed in all specimens), a metaflexid/fossettid with a metastriid as the second longest lingual striid and a mesoflexid/fossettid with the longest of the lingual striids, the mesostriid. Two of the hardly worn specimens show an open metaflexid that is closed in all other dp4 to a metafossettid. Mesoflexid is only closed in some dp4 of higher wear stages forming a mesofossettid (GPIT/MA/18993, 19096, 19097, 19098). In exceptional cases, the following variations may occur in deviation from the basic construction plan: presence of one or two small and circular preparafossettids (one preparafossettid: GPIT/MA/19093, 19169, 19172, 19173; two preparafossettids: GPIT/MA/18988, 19092, 19166); parafossettid is U-shaped (GPIT/MA/19095); In a single case, the paraflexid is connected to the mesoflexid, elongating the mesoflexid in anterio-buccal direction (GPIT/MA/11999). In the HAM 5 material 2 out of 34 dp4 have preserved roots, while in the HAM 4 material all 9 available lower deciduous premolars are only preserved as rootless tooth crowns. The dp4 is two-rooted with one anteriorly and one posteriorly diverging root (GPIT/MA/18988, 18993).
p4 ()
The lower premolar clearly represents the largest tooth of the lower tooth row and is slightly inclined anteriorly. The occlusal outline is elongated and figure eight shaped and generally four flexids/fossettids are exposed, one buccal hypoflexid and three lingual flexids (para-, meso- and metaflexid). With progressive wear, the occlusal tooth dimensions increase in longitudinal and transverse direction with a stronger increase of the mesio-distal length. Lingual flexids are straight or slightly curved and oriented mesiobuccally, opposing the hypoflexid. All striids end before reaching the crown base. The hypoflexid presents the longest striid and is open in all available p4 and age categories, except for a single, very heavily worn p4 in which a hypofossettid is formed (GPIT/MA/18725) and one tooth exposes a closing hypoflexid (GPIT/MA/18711). In a few cases of very young age classes (unworn or slightly worn teeth), a very short and mesially open paraflexid can be present (GPIT/MA/12480, 17240, 17307, 18671, 18682, 18694, 18695, 18700, 18702, 18704, 18707, 18723, 18727, 18729, 18732, SNSB-BSPG 2020 XCIV-6870, 6876, 6899), but in most cases, it is close to a parafossettid. A similar observation can be made for the metaflexid, that can be lingually open (and slightly longer than the parastriid), in unworn and slightly worn p4. The mesoflexid exhibits the longest lingual striid and remains open until higher age classes (only medium, deeply and highly worn p4 show a mesofossettid). In unworn and slightly worn p4, it can be connected to the hypoflexid. Further features can be one to two preparafossettids (one: GPIT/MA/12480, 17240, 17419, 17444, 18664, 18670, 18674, 18685, 18693, 18699, 18716, SNSB-BSPG 2020 XCIV-6889; two: GPIT/MA/10756, 17700, 18668, 18678, 18730, SNSB-BSPG 2020 XCIV-6874, 6893) and/or premetafossettids (one: GPIT/MA/18664, 18668, 18683, 18722, SNSB-BSPG 2020 XCIV-6889; two: SNSB-BSPG 2020 XCIV-6887, divided parafossettids (SNSB-BSPG 2020 XCIV-6875) and/or metafossettids (GPIT/MA/18693) or bifurcated parafossettids (GPIT/MA/17444, SNSB-BSPG 2020 XCIV-6894, 6903). The lower premolar has two equally sized roots with a crescent shaped cross-section.
m1-2 ()
The m1/2 is shorter crowned than the p4 and inclined anteriorly within the lower tooth row. The occlusal outline of lower m1/2 is nearly square with a straight and transverse anterior and a slightly convex posterior tooth wall. With wear, mesio-distal length becomes shorter, while the width remains approximately the same (rectangular outline). The principal morphology is similar to that of the p4 but teeth are shorter. In general, an oblique posteriorly oriented hypoflexid and a slightly anteriorly oriented mesoflexid are present. Both flexids end at the median line of the tooth with the hypoflexid located between the mesoflexid and metaflexid/-fossettid. Only in a few unworn and slightly worn m1/2 an open paraflexid is present (GPIT/MA/19035, 19150, SNSB-BSPG 2020 XCIV-0725-2), while an open metaflexid is more frequently observed (GPIT/MA/10995, 17710–2, 17949–1, 17949–2, 18964, 19030, 19032, 19035, 19037, 19104, 19140, 19150, SNSB-BSPG 2020 XCIV-0725–2, 6915–2, 6978, 6981, 7035, 7041, 7044–2). The mesoflexid closes at medium wear stages and in a few cases of very heavily and deeply worn m1/2 the hypoflexid is close to a hypofossettid (GPIT/MA/10757, 12479–1, 18934, 18949, 18968, 19021, 19149, SNSB-BSPG 2020 XCIV-4235). The paraflexid/-fossettid is curved and oriented anteriorly and is placed in the centre of the anterior m1/2. In many cases of up to moderately worn m1/2, a tiny preparafossettid exists (GPIT/MA/10995, 12432, 12480–1, 12480–2, 16433–1, 17240–2, 17444–1, 17444–2, 17710–1, 17710–2, 17766–2, 17949–1, 17949–2, 17952, 19019, 19029, 19030, 19032, 19037, 19039, 19040, 19041, 19044, 19102, 19103, 19104, 19141, 19143, 19144, 19145, SNSB-BSPG 2020 XCIV-0725-1, 0725–2, 6915–1, 6915–2, 6975, 6977, 6978, 6980, 6981, 7035, 7038, 7044–1, 7044–2), that sometimes can be connected with the paraflexid/-fossettid to form a lingually open U-bend (GPIT/MA/19035, 19146, 19148, 19150). In a single case, two preparafossettids exist and are connected to form a horizontal figure eight (GPIT/MA/19023). The metaflexid/-fossettid is rather straight and aligned in parallel to the posterior tooth margin. The hypostriid is the longest striid that ends short above the base of the tooth crown. The mesostriid is the second longest striid reaching approximately halve of the tooth crown, while para- and metastriid are only present in some cases and often only observed in early wear stages. The tree-rooted m1/2 show two equally short and small anterior roots and one large transverse posterior one.
m3 ()
The m3 is the shortest-crowned tooth of the lower tooth row, strongly inclined anteriorly, comprising a similar principal morphology as the m1/2 but instead of a square occlusal outline, the molar is slightly elongated and tapers buccally from the posterior half of the tooth. No significant dimensional changes can be observed with progressive wear. Lower m3 comprises a straight obliquely oriented buccal hypoflexid, and the tree lingual para-, meso- and metaflexids/-fossettids. A paraflexid can only be observed in few slightly worn specimens (GPIT/MA/12269, 18982, 18983, 19110), whereas in the other teeth a parafossettid is expressed that is slightly concave and spans transversely the entire anterior portion of the tooth. One to two small preparafossettids can be placed within this anterior-opened arch (one: GPIT/MA/17444-3, 17700–3, 19086, 19109, 19110, SNSB-BSPG 2020 XCIV-6983, 6984; two: SNSB-BSPG 2020 XCIV-6985). In less worn teeth, this preparafossettids can be connected to the anterior tooth wall and the parafossettid can be open anteriorly (GPIT/MA/18974, SNSB-BSPG 2020 XCIV-6985). The mesoflexid/-fossettid is slightly shorter than the metaflexid/-fossettid, which both are straight and reach near to the half of the tooth width and terminate near to the anterior and posterior end of the hypoflexid. The hypostriid is the longest striid reaching nearly the end of the tooth crown but still closes in a heavily worn specimen (GPIT/MA/19122). The mesostriid is the second longest striid and reaches the first third of the tooth crown, closely followed by the shorter metastriid. A parastriid is rarely formed and when present it is extremely short and closes during the very first wear. The m3 is three rooted, with two very tiny anterior roots and one large one, that runs transverse along the convex posterior tooth wall.
Results and discussion
Morphologic comparison
The dental material from the HAM 5 and HAM 4 levels of the Hammerschmiede fossil site show the typical characters for Euroxenomys minutus following Hugueney (Citation1999) and Daxner-Höck and Bernor (Citation2009) with additions after Prieto et al. (Citation2014), Kordos (Citation2020) and Mörs et al. (Citation2022).
(1) Small size; (2) upper premolar (P4) subtriangular in occlusal outline and clearly larger than upper molars; (3) lower premolar (p4) that is elongated, significantly larger than lower molars and forming an anteriormost edge; (4) rooted and high crowned cheek teeth with mostly parallel-oriented flexus/fossettes and flexids/fossettids without cement in synclines and hypostriae/-ids that do not reach the crown base; (5) characteristic enlarged upper third molar (M3)
The extensive dental material collection of Euroxenomys minutus from the Hammerschmiede fossil site is morphologically highly variable but fits well with other Eurasian Miocene localities, such as Tagay (MN 5) in Siberia (Mörs et al. Citation2022), Sansan (MN 6) in France (Hugueney and Duranthon Citation2012), Gratkorn (MN 7/8) in Germany (Prieto et al. Citation2014), Mataschen (MN 7/8) in Austria (Daxner-Höck Citation2004), Anwil (MN 7/8) in Switzerland (Engesser Citation1972), Felsötárkány (MN 7/8) in Hungary (Hír Citation2004), Atzelsdorf (MN 9) in Austria (Daxner-Höck and Bernor Citation2009), Höwenegg (MN 9) in Germany (Giersch et al. Citation2010) and Rudabánya (MN 9) in Hungary (Kordos Citation2020). An additional mesial preparafossettid/fossette (occurring in Hammerschmiede material at all lower cheek tooth positions and the first time documented for the upper P4) usually visible at earlier wear stages is already described in material from Tagay (p4 and m3), Rudabanya (m2 and m3), Anwil (dp4, m3), Sansan and Atzelsdorf (p4 and m1/2) (Engesser Citation1972; Daxner-Höck and Bernor Citation2009; Hugueney and Duranthon Citation2012; Kordos Citation2020; Mörs et al. Citation2022). Moreover, duplicate preparafossettids, or additional premetafossettid/fossette, premesofossette or postmetafossette can be varying supplementary characters (). Further possible variations within the enormous cheek teeth collection are differences in length, course and orientation as well as possible bipartitions, bifurcations or connections of occlusal flexids/flexus or fossettids/fossettes.
Metric comparison ()
In order to investigate the subjective influence of the measurer on values obtained, additionally to the Hammerschmiede material, own measurements were taken for Euroxenomys minutus rhenanum from the Dorn-Dürkheim site, where values deviate slightly from those from the literature and differences mainly concern the width values. This methodological uncertainty may explain to some extent differences in especially the width values from teeth of the compared fossil sites taken from literature, which means that slight ranges of metric variation should not be weighted too much and could be based on methodological differences in measurement techniques.
Measurements for lower and upper teeth from the local stratigraphic levels HAM 5 and HAM 4 from the Hammerschmiede deposits exactly overlap and can unambiguously be used as one dataset (HAM). The smallest values of upper premolars correspond to occlusal measurements of less worn teeth. In contrast to lower premolars, basal measurements of upper premolars and thus also teeth of higher wear stage and age show similar length values, but a slightly higher width and occlusal measurements show a higher scatter than basal ones (). A similar pattern can be observed in the collection of Euroxenomys minutus teeth from Rudabánya in Hungary (). The smallest values of lower premolars correspond to occlusal measurements of less worn teeth, the larger values enclose measurements taken at the base of the crown, which in turn match occlusal values of some very worn teeth. With progressive tooth wear (age), lower premolars become slightly wider but significantly longer ().
Metrically, the upper premolars from Hammerschmiede fit well with Euroxenomys minutus from Tagay (MN 5), Gratkorn (MN 7/8), Anwil (MN 7/8) and Atzelsdorf (MN 9) (). The single P4 from Petersbuch 50 (MN 8) corresponds with the highest, and from Felsötárkány (MN 7/8) with the lowest values of Hammerschmiede specimens. The large collection from Rudabánya (MN 9) fits well with the highest width values and overlaps best the basal measurements of upper Hammerschmiede premolars. The Rudabánya material also shows even greater width values at the basal P4 (). Upper premolars from Sansan (MN 6) partly match the highest values and Dorn-Dürkheim specimens (Euroxenomys minutus rhenanus) are substantially larger than those from Hammerschmiede ().
The lower p4 from Hammerschmiede match with material from Elgg (MN 5), Anwil (MN 7/8), Felsötárkány (MN 7/8), Petersbuch 50 (MN 8), Atzelsdorf (MN 9) and Höwenegg (MN 9) (). Lower p4 from Gratkorn (MN 7/8) are within the maximum width limit of the Hammerschmiede material and the Tagay (MN 5) specimen is slightly wider, but within the length range of Hammerschmiede p4 (). Rudabánya (MN 9) p4 fit very well, but the shortest length values show a higher width maximum at the occlusal tooth portion, while basal tooth measurements agree with the values for lower premolars from Hammerschmiede (). Rudabánya and Hammerschmiede both show a very high variability in p4 length with a close match in the range of values (). Lower p4 from Sansan (MN 6) show consistency in their lower range of values, however most measurements are wider than the maximum width of Hammerschmiede specimens (possibly different measuring sections are used) (). Dorn-Dürkheim p4 (own measurements) are larger and wider and follow a different regression line than the Hammerschmiede measurements ().
To distinguish finally between the two subspecies (or possibly species) Euroxenomys minutum minutum and E. minutum rhenanum, the differences already mentioned by Franzen and Storch (Citation1975) can be maintained and confirmed. Even the extremely large data set from Hammerschmiede does not manage to match the range of large values for upper and lower premolars from Dorn-Dürkheim. Finally, the diagnosis for E. minutum rhenanum made by Franzen and Storch (Citation1975) remains valid and the best criteria to distinguish from E. minutum minutum are (1) the larger premolars in absolute terms and in comparison to M1/2 and m1/2, (2) the M3, which are significantly longer in absolute terms and compared to M1/2 and (3) M1 and m1, which are wider compared to M2 and m2 in E. minutum rhenanum. This also results in a longer jaw tooth row length in E. minutum rhenanum.
Taphonomy and signs of predation ()
The presented tooth and jaw material of Euroxenomys minutus from Hammerschmiede shows a significantly high quantity of isolated teeth compared to in situ tooth rows and jaw fragments (HAM 5: 442/10; HAM 4: 454/75). In both studied strata, this ratio of disarticulation is high with 83% isolated dental specimens at HAM 4 and very high with 98% at HAM 5. A considerable proportion of dental specimens shows moderate and heavy corrosive lesions (). In most cases, it is mainly the enamel that is affected, and in jaw specimens with in situ dentition, the bone itself is hardly or not at all dissolved, while the crowns of the teeth have been considerably corroded. Corrosive lesions mainly affect the visible part of the enamel. Areas of enamel that are still hidden in the jawbone remain largely undamaged as it is best visible on incisors (Figs. 8B1, C2). It is obvious that corrosive effects on molars are greatest at the more basal enamel areas, just above the alveolar rim of the jawbone, while enamel close to the occlusal surface exhibits only weak corrosive lesions (Figs. 8A3, A4, C3).
Figure 8. Signs of digestive corrosion on dental material of the minute beaver Euroxenomys minutus (von Meyer Citation1838), from the early Late Miocene locality Hammerschmiede (Bavaria, Germany), local stratigraphic levels HAM 5 and HAM 4. (a) SNSB-BSPG 2020 XCIV-6917, left mandible with corrosion on p4 and m1 enamel in lingual (A1, A4), buccal (A2-A3) views and double magnifications of the corroded enamel regions (A3, A4). (B) GPIT/MA/19190, left lower incisor i2 with corrosion on enamel regions in labial/anterior (B1) and mesial (B2) views. Enamel of the tooth tip region is already completely missing while the proximal half preserves some portions of enamel. (C) GPIT/MA/17703, left mandible with broken i2 and cheek teeth p4-m2 with corrosion of the tooth enamel at the i2 and at the base of the molars in buccal (C1-C3) and double magnified (C2-C3) views. (D) GPIT/MA/19188, left lower incisor i2 without corrosion effects for comparison purposes with the corroded incisors in buccal (D1) and labial/anterior (D2) views. (E) SNSB-BSPG 2020 XCIV-0607, upper right incisor I2 in buccal (E1) and labial/anterior (E2) views, with signs of dental enamel corrosion that possibly amplify the incremental rhythm of growth lines. Scale bar equals 10 mm except for magnifications A3, A4, C2 and C3 where it corresponds to 5 mm.

Some incisors show strongly corroded, striated enamel patterns, which were carved out and highlighted by the corrosion and presumably represent the rhythm of diurnal incremental tooth growth lines (Figs. 8C2, E1, E2). Possibly cyclic changes in tooth properties provide weak points or inhibitions for acids to work.
In addition to dental corrosions, it is striking that long bones in particular are in many cases only preserved in broken form. According to several studies, the degree and position of the corrosive lesions could correspond to digestive corrosions observed on prey bones in recent predators that have an acidic digestive system, such as mammalian carnivores (Andrews and Evans Citation1983; Lopez et al. Citation2017, Figure 3; Marin-Monfort et al. Citation2019) or predatory birds, e.g., owls (Andrews Citation1990; Fernández-Jalvo and Andrews Citation2016, Fig. A767; Fernández et al. Citation2019). According to Andrews (Citation1990), there is no other alteration process that can replicate the corrosive effects of digestion, which makes such traces, especially on teeth, a clear indicator of predatory bone assemblages. In contrast to digestive corrosion, bone fractures alone are not good indicators of a predator assemblage, as it cannot be ruled out that further modifications may have been caused by processes like weathering, trampling or sediment compression (Andrews Citation1990), including excavation artefacts. Generally, each predator species produces a different type of alteration on the tooth and bone material, which means that some predators can be assigned to their traces (Andrews Citation1990). With approximately the body size of a muskrat, large birds of prey as well as medium- and large-sized mammalian carnivores are certainly possible predators of Euroxenomys minutus at the Hammerschmiede fossil site. However, we lack corresponding studies on the respective individual corrosive effects of digestion of the numerous possible fossil predator species.
Mortality analysis ()
To map the entire lifespan of beaver individuals in a mortality analysis, the tooth position of the lower dp4 and p4 is used (Heinrich and Maul Citation2020; Kordos Citation2020; Lechner and Böhme Citation2022). The general problem with using deciduous teeth (here dp4) in the mortality analysis is the possible duplication of individuals, by interpreting teeth as deceased juvenile individuals and later duplicating the same individual by an adult p4 with the actual wear stage it died. For this reason, when using deciduous teeth, meticulous care must be taken to ensure that only teeth that show no signs of resorbed roots are used. In addition, when using deciduous teeth, the permanent p4 must not be supplemented at the same time if it shows no signs of wear, as both are in the juvenile jaw at the same time.
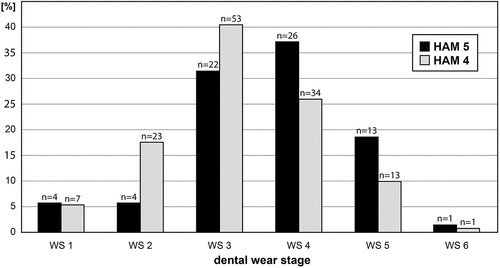
Figure 9. Mortality profiles (age-frequency distribution) of the minute beaver Euroxenomys minutus (von Meyer Citation1838), from the early Late Miocene locality Hammerschmiede (Bavaria, Germany), based on lower premolars from the local stratigraphic levels HAM 5 and HAM 4. Each bar corresponds to an age class, defined by either morphologic criteria (WS 1-WS 2) or mathemathical spread of the tooth crown height (WS 3-WS 6) as defined in the materials and methods section. Vertical axis represents the percentage of individuals within the corresponding local stratigraphic level.
In the present case of the Hammerschmiede, the data from dp4 and unused p4 were compared with each other, and it turns out that only two out of 34 dp4 at the HAM 5 and none of the nine from HAM 4 show preserved roots. All ‘unrooted’ dp4 have lost their roots, possibly through root resorption (shedding) or have been mechanically eroded during transportation processes in the alluvial system, what cannot be definitively determined. In contrast, there are four unused p4 from HAM 5 and seven from HAM 4, which should finally generate a larger and thus better data security for this age group, also since the larger lower premolars have a better chance of being found in the field and during wet sieving than the very small dp4. Although the shed deciduous premolars cannot be used for the mortality study, as they represent a ‘life process’, they indicate autochthony of Euroxenomys minutus at both HAM 5 and HAM 4 habitats.
All available lower premolars of appropriate preservation from HAM 5 and HAM 4 strata representing the small beavers Euroxenomys minutus were counted and categorised by wear-stage definitions provided in the material and methods paragraph. With the final counts on the six wear stages, an age-frequency distribution – Mortality profile – following Lyman (Citation1994) was performed ().
The age-frequency distributions for Euroxenomys minutus are based on 70 lower premolars from the local stratigraphic level HAM 5 and 131 p4 from the HAM 4 layer. In general, the shape of the two resulting mortality profiles is quite similar but a slight shift by one wear stage, to an older age at death in the HAM 5 material can be observed (). In the HAM 5 profile, WS 1 (5.7%, n = 4) and WS 2 (5.7%, n = 4) show very low values, followed by the most dominant WS 3 (31.4%, n = 22) and WS 4 (37.1%, n = 26) counts. WS 5 (18.6%, n = 13) still exposes a quite high value, while WS 6 (1.4%, n = 1) is the most underrepresented age class. The age-frequency distribution of lower premolars from HAM 4 reveals a similarly low WS 1 (5.3%, n = 7) as in HAM 5. In contrast to the latter, WS 2 already exhibits a considerably higher score (17.6%, n = 23) in the HAM 4 material, followed by the highest mortality proportion during WS 3 (40.1%, n = 53) and a slightly lower value in WS 4 (26%, n = 34). In contrast to HAM 5, WS 5 (9.9%, n = 13) from HAM 4 is already low, while both share a very low value for WS 6 (0.8%, n = 1).
The mortality profiles of both HAM 5 and HAM 4 layers neither correspond with the typical U-shaped (attritional death model) age-frequency distribution nor with the characteristic L-shaped (catastrophic death model) mortality pattern after Voorhies (Citation1969) and Lyman (Citation1994). In the case of both Hammerschmiede mortality profiles, the shape is more likely to be described as a ‘pyramid’-shaped pattern. A typical peak in the form of a high mortality in juveniles is not observed in both HAM 5 and HAM 4 age-frequency distributions. Instead, the highest mortality is among individuals in the ‘prime’ age group (WS 3 and WS 4). The peak of the mortality ‘pyramid’ of HAM 5 is located at WS 4 and in HAM 4 at WS 3, and thus shifted by one wear stage. Apart from that, the two profiles are extremely similar.
In a second mortality analysis, a ternary diagram following the basic methods introduced by Stiner (Citation1990) and modified after Discamps and Costamagno (Citation2015) was created (). Here only three age groups are used (juvenile, prime adult and old adult). These three groups are obtained by always combining the counts of two wear stages (juvenile = WS 1+ WS 2, prime adult = WS 3+ WS 4 and old adult = WS 5+ WS 6). In addition to the classic L-shaped (JPO dominated zone) and U-shaped (JOP dominated zone) mortality profiles, the ternary diagram can be used to compare more options. To display the four dominance zones (Juveniles-Old-Prime = JOP; Juveniles-Prime-Old = JPO; Old = O; and Prime = P) introduced by Discamps and Costamagno (Citation2015) a reference point must be defined, which is maintained due to the species-specific lifetime distribution. Since we do not know the relative distribution of the three age groups compared to the average lifetime expectancy in Euroxenomys minutus, this point was derived as follows. While for the large beaver species from Hammerschmiede Steneofiber depereti, the lifetime distribution is assumed to be similar to today’s beavers, this is not necessarily applicable to Euroxenomys minutus, especially due to the tiny body size. Given the extremely high functional morphological and presumably ecological similarity of Euroxenomys minutus to today’s muskrat (Ondatra zibethicus), its life cycle could be a possible assumption for comparison. Muskrats have a significantly shorter life span of about 3 years (Godin Citation1977; Willner et al. Citation1980; Nowak Citation1991). The muskrat reaches sexual maturity and thus the end of the juvenile period with about 7 months (Heidecke and Seide Citation1989; Birnbaum Citation2013). Until the end of the second year of life, the muskrat is considered a prime adult and from the second year of life onwards an old adult. This roughly corresponds to a distribution of 20% juvenile (7 months), 47% prime adult (17 months) and 33% old adult (12 months). Since the reference point used for Castor fibre from extant studies and Pleistocene sites of a corresponding analysis from Campbell (Citation2009) and Heinrich and Maul (Citation2020) marks a similar point (24% joung, 43% prime, 33% old) in the ternary diagram as the calibration just derived for the muskrat, the decision between both calibration points is not necessary and we continue to work with the muskrat data.
Figure 10. Ternary diagram of the relative age-frequency distributions (mortality) differentiated into the three age groups juvenile, prime adult and old adult individuals (in %) of the small beaver species Euroxenomys minutus (von Meyer Citation1838) from the early Late Miocene localities Hammerschmiede (Bavaria, Germany), including the local stratigraphic levels HAM 5 and HAM 4 and from Rudabánya (Hungary; Data from Kordos Citation2020) based on lower premolars. Calibration of the zoning point corresponds to the relative lifetime distribution of the extant muskrat, Ondatra zibethicus (see text). Abbreviations of zones follow Discamps and Costamagno (Citation2015): JOP, juveniles-old adults-prime adults dominated zone; JPO, juveniles-prime adults-old adults dominated zone; O, old adults dominated zone; P, prime adults dominated zone. All three fossil samples are located in the prime dominated zone.
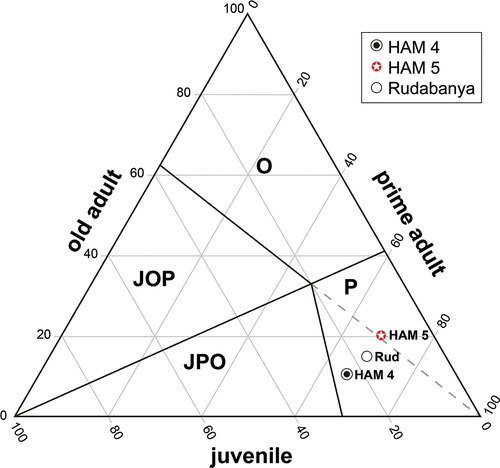
As already indicated in the mortality profile, the mortalities of the compared Euroxenomys sites do not correspond to any of those classically differentiated zones, but in a separate area called ‘P’ (prime dominated zone). Hereby, it can be further specified that HAM 5 is located in a subzone P > O > J and HAM 4 and Rudabanya are located in the subzone P > J > O. However, these subtle differences are probably of marginal importance and are not the focus of our further discussion. It must be emphasised that the points are not located in the standard fields, but in the prime dominated zone, which should not be the case in a classical normal mortality (Discamps and Costamagno Citation2015). Thus, a special influence is required for a mortality to shift from juvenile dominance, which would be expected due to typically high infant mortality, to prime adult dominance. Such prime adult dominance has hardly been described in beavers so far, but is rather found in archaeological contexts. Heinrich and Maul (Citation2020) describe an almost identical mortality in Castor fibre from the Pleistocene sites Bilzingsleben II, Weimar-Ehringsdorf and Weimar-Taubach. The authors explain this taphonomic filter as an accumulation of similarly aged beavers that were selectively hunted by hominins. Since we exclude hominin/hominid hunting on beavers for Hammerschmiede, a dominating influence by other predators is the most likely explanation.
We already published the age-frequency distribution of another, much larger beaver species from Hammerschmiede locality – Steneofiber depereti (Lechner and Böhme Citation2022). For this large beaver, a massive difference in the mortality distribution between the two layers (HAM 5 and HAM 4) was found, which we attribute mainly to ecological reasons of Steneofiber depereti and the observation that HAM 5 represents a rivulet and HAM 4 a larger river with different environmental parameters. S. depereti, a beaver slightly smaller in size than today’s beavers, most probably preferred the larger river (represented by HAM 4 sediments) as the core habitat, while the HAM 5 rivulet was a temporary habitat for young adult beavers in search of their own territory (Lechner and Böhme Citation2022). Although such observations and differences in mortality can be made for the large beaver species from Hammerschmiede, the age-frequency distribution of the tiny Euroxenomys minutus shows a totally different pattern. A possible explanation for these major differences could be a different ecologic lifestyle of these beaver species and also the huge difference in size. Euroxenomys minutus possibly is subject to strong predation influence due to its small size, in contrast to the much larger and therefore more defensible Steneofiber depereti. The body size of Euroxenomys minutus should be about the same as the extant muskrat, with an average weight of approximately 1100 g (750–1575 g) provided for the muskrat by Fuller (Citation1951). For rodents of this weight class, a very large number of avian and mammalian predators are available in the Hammerschmiede ecosystems, e.g., several aquatic/semiaquatic as well as terrestrial carnivores and birds of prey. A predatory selection in Euroxenomys minutus according to life age seems possible if especially the young animals are not accessible for the predators for certain reasons. However, the low number of very young Euroxenomys beavers in the mortality is possibly based on habitat or behavioural reasons. Perhaps young Euroxenomys stayed hidden in burrows inaccessible to predators or they moved mainly in more by vegetation protected areas near the dwelling, while adults that cover much greater distances in search of food and resources, were easy targets of predators in the water and on land.
As already described before, there is no data on the lifetime of the fossil Euroxenomys minutus and consequently we can only assume which time-steps the age-frequency distribution covers. While in the large Steneofiber depereti, the lifetime probably corresponds to that of today’s large beaver species and other large rodents (up to 15–24 years living in the wild; Brown Citation1979; Grzimek Citation1990; Bobick and Peffer Citation1993), the lifetime expectancy of Euroxenomys minutus is assumed to correspond with that of smaller rodents like the muskrat (Ondatra zibethicus) with an estimated 3 years of life (Godin Citation1977; Willner et al. Citation1980; Nowak Citation1991). With such a short-life expectancy, mortality analyses would react much more sensitive and could depict processes that take place over a period of a few months only. The probability of dying during a short juvenile phase (Ondatra) is of course much lower than if this phase lasts longer (Castor). This certainly has an influence on the relative frequency of juvenile teeth to adult teeth.
For an intraspecific comparison of Euroxenomys minutus, the age-frequency distribution from the slightly younger fossil site of Rudabánya in Hungary is compared (Kordos Citation2020). Here, very young and very old beavers are also quite rare in the mortality analysis, while the main count is represented by medium-aged beavers (Kordos Citation2020). The age frequency distribution in the mortality analyses for Euroxenomys minutus from Rudabánya is therefore very similar to that observed at HAM 5 and HAM 4 (). This finding is particularly interesting also by the fact that Rudabánya represents a fossil swamp environment (Kordos Citation2020). Thus, in addition to the channel of HAM 5 interpreted as a small rivulet and the HAM 4 channel representing larger river deposits, a third environment interpreted as a swamp (Rudabánya) now also shows a very comparable mortality distribution (Böhme et al. Citation2019; Kordos Citation2020). In contrast to the larger Miocene beaver species Steneofiber depereti (which is not found at Rudabánya site) our results show that on one hand Euroxenomys minutus is an ubiquitary dweller of various aquatic habitats (swamp, rivulet and river), much as the muskrats today, on the other hand, the small beaver in all of these habitats seems to be under an influence that produces a very similar age distribution and mortality. As a possibly very dominant external factor, especially because of the small body size of Euroxenomys minutus as a prey, a selective predation on prime age Euroxenomys by the afore mentioned large amount of different available predators (diurnal and nocturnal avian and terrestrial and aquatic/semiaquatic mammalian carnivores) is probably one of the most causal reasons for these similarities in the age-frequency distributions of those habitats. Another explanation could be a special behaviour of the young beavers, which makes them more difficult to reach for predators.
Conclusion
Beavers are extremely abundant finds at the early Late Miocene Hammerschmiede locality. The two local stratigraphic levels worked on here represent a small rivulet (HAM 5) and a river (HAM 4) and in both there occurs a common large beaver (Steneofiber depereti) as well as an even more frequent small beaver form. We assign the small form to the trogontheriine beaver Euroxenomys minutus (von Meyer Citation1838). The extensive set of dental remains from Hammerschmiede represent the largest Miocene record for this beaver species at all. This facilitates a very realistic insight into the metric and morphological intraspecific variability of this small beaver, which is marked by an enormous range of variation in these points. Euroxenomys minutus from Hammerschmiede includes the hitherto rather heterogeneous individual data from the many other Eurasian discovery sites and proves their affiliation to one species. Furthermore, a clear delimitation to teeth from the locality Dorn-Dürkheim (Germany) can be confirmed and the status of this morphotype at least at subspecies level (Euroxenomys minutus rhenanus) is approved as correct.
In contrast to the large beaver species Steneofiber depereti from Hammerschmiede, where the local stratigraphic levels HAM 5 (rivulet) and HAM 4 (river) demonstrate significantly different mortality patterns, the age-frequency distributions of Euroxenomys minutus reveal a higher degree of similarity for both of these fossil habitats. In addition to that, the age-frequency distribution of Euroxenomys minutus from Rudabánya in Hungary, that is assumed to be a fossil swamp, is consistent with this image, despite the fact that no large beaver species is found here. In all three habitats a very low number of individuals is represented by very young beavers, a high mortality indicates the midfield and only few very old animals were found. This high similarity in the mortality profiles of the river, rivulet and swamp habitats indicates, that on one hand, the ecology of the small Euroxenomys minutus could be different from that of the large beaver, and on the other hand, that the small beaver is probably more vulnerable to external factors like predators, due to its size. The latter hypothesis is supported by traces of corrosive lesions on several dental specimens, which could be attributed to corrosive effects of digestion by mammalian carnivores or predatory birds. Furthermore, the extremely high proportion of fragmented cranial (more than 90% of dental material are single teeth) and postcranial material and especially the long bones could indicate that finds of Euroxenomys minutus at the Hammerschmiede represent a predatory bone assemblage.
Supplemental Material
Download MS Excel (82.4 KB)Acknowledgments
The authors would like to thank I. Werneburg (GPIT) for providing access to the objects stored in the Tübingen collection (GPIT and SNSB-BSPG stored at GPIT) and T. Lehmann for access to the specimens from Dorn-Dürkheim under his care (Senckenberg, Frankfurt, Germany). Moreover, we are very grateful to L. Kordos (Eötvös Loránd University, Budapest) for providing his records and measurements on the beaver material from Rudabánya. Further, we thank W. Lechner (Nawilab, Trostberg, Germany) for providing the most recent reference material. We thank A.-C. Grupp (GPIT and SNSB-BSPG), D. Wedekind (Senckenberg Centre HEP, Tübingen, Germany) and J. Franke (SNSB-BSPG) and H. Stöhr (GPIT) for the preparation of the specimens. We further acknowledge all participants of the numerous excavations in the Hammerschmiede fossil site, who helped to detect and collect the studied material, with special thanks to the members of the organisation team of the excavation years 2020–2022: F. Augustin, A. Ayvazyan, C. Dietzel, H. El Atfy, J. Hartung, P. Kampouridis, N. Kargopoulos, U. Kirscher, C. Kyriakouli, M. Lex, T. Massonne, A. Matzke and M. Neuhof. The excavations and associated research were supported by the Bavarian State Ministry of Research and the Arts and by the Bavarian Natural History Collections (SNSB). We would also like to thank I. Gold, C. Dietzel, P. Kampouridis, F. Augustin, and A. Matzke (all GPIT) for fruitful discussions and improvement of the manuscript. Finally, we thank the editor G. Dyke, and the reviewers T. Mörs and an anonymous reviewer for helpful comments and suggestions on the manuscript.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Supplemental data
Supplemental data for this article can be accessed online at https://doi.org/10.1080/08912963.2023.2215236.
Additional information
Funding
References
- Aldana Carrasco E. 1992. Los Castoridae (Rodentia, Mammalia) del Neógeno de Cataluña (España). Treballs del Museu de Geologia de Barcelona. 2:99–141. Spanish
- Andrews P. 1990. Owls, Caves and Fossils. Chicago: University of Chicago Press; pp. 231.
- Andrews P, Evans EM. 1983. Small mammal bone accumulations produced by mammalian carnivores. Paleobiology. 9(3):289–307. doi:10.1017/S0094837300007703.
- Birnbaum C 2013. NOBANIS – invasive alien species fact sheet – Ondatra zibethicus. – from: online database of the European Network on invasive alien species – NOBANIS www.nobanis.org, Date of access 2023 March 30.
- Bobick JE, Peffer M 1993. Science and technology desk reference. Gale research Inc. Washington (D.C).
- Böhme M, Spassov N, DeSilva JM, Begun DR. 2020. Reply to: Reevaluating bipedalism in Danuvius. Nature. 586(7827):E4–E5. doi:10.1038/s41586-020-2737-3.
- Böhme M, Spassov N, Fuss J, Tröscher A, Deane AS, Prieto J, Kirscher U, Lechner T, Begun DR. 2019. A new Miocene ape and locomotion in the ancestor of great apes and humans. Nature. 575(7783):489–493. doi:10.1038/s41586-019-1731-0.
- Brown MK. 1979. Two old beavers from the Adirondacks. New York Fish and Game J. 26:92.
- Campbell RD 2009. Demography and life history of the Eurasian beaver Castor fiber. Ph.D. thesis. – MS, Department of Zoology, University of Oxford, Oxford, 282 pp. (thesis available online)
- Daxner-Höck G. 2004. Biber und ein Zwerghamster aus Mataschen (Unter-Pannonium, Steirisches Becken). Joannea, Geologie und Paläontologie. 5:19–33. German
- Daxner-Höck G, Bernor RL. 2009. The early Vallesian vertebrates of Atzelsdorf (Late Miocene, Austria) 8. Anchitherium, Suidae and Castoridae (Mammalia). Annalen des Naturhistorischen Museums in Wien. 111(A):557–584.
- Discamps E, Costamagno S. 2015. Improving mortality profile analysis in zooarchaeology: a revised zoning for ternary diagrams. Journal of Archaeological Science. 58:62–76. doi:10.1016/j.jas.2015.03.021.
- Engesser B. 1972. Die obermiozäne Säugetierfauna von Anwil (Baselland). Tätigkeitsbericht Naturforschende Gesellschaft Baselland. 28:1–363. German
- Fahlbusch V, Mayr H. 1975. Microtoide Cricetiden (Mammalia, Rodentia) aus der Oberen Süßwasser-Molasse Bayerns. Paläontologische Zeitschrift. 49(1/2):78–93. doi:10.1007/BF02988068. German
- Fernández FJ, Hadler P, Cherem JJ, Saldanha JP, Stutz NS, Dias AS, Pardiñas UFJ. 2019. Holocene small mammals hunted by owls and humans in southern Brazil: taphonomic evidence and biological significance. Boreas. 48(4):953–965. doi:10.1111/bor.12399.
- Fernández-Jalvo Y, Andrews P. 2016. Atlas of taphonomic identifications. Dordrecht: Springer; pp. 359.
- Fischer von Waldheim G. 1809. Sur l’Elasmotherium et le Trogontherium, deux animaux fossiles et inconnus de la Russie. Études palaeontologiques sur les environs de Moscou 2: 250–268. French
- Franzen JL, Storch G. 1975. Die unterpliozäne (turolische) Wirbeltierfauna von Dorn-Dürkheim, Rheinhessen (SW-Deutschland); 1. Entdeckung, Geologie, Mammalia: Carnivora, Proboscidea, Rodentia. Grabungsergebnisse 1972–1973. Senckenbergiana lethaea. 56. 233–303. German
- Freye H-A. 1959. Deskriptive Anatomie des Craniums vom Elbe-Biber (Castor fiber albicus MATSCHIE 1907). Wissenschaftliche Zeitschrift der Martin-Luther-Universität Halle-Wittenberg. Math-naturwissenschaftliche Reihe. VIII. 913–962. German
- Fuller WA. 1951. Measurements and weights of Northern Muskrats. Journal of Mammalogy. 32(3):360–362. doi:10.1093/jmammal/32.3.360.
- Fuss J, Prieto J, Böhme M. 2015. Revision of the boselaphin bovid Miotragocerus monacensis Stromer, 1928 (Mammalia, Bovidae) at the Middle to Late Miocene transition in Central Europe. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen. 276(3):229–265. doi:10.1127/njgpa/2015/0481.
- Geoffroy-Saint-Hilaire É. 1833. Palaeontographie. Considérations sur des ossemens fossiles la plupart inconnus trouvés et observés dans les bassins de l’Auvergne. Revue Encyclopédique. 59(LIX):76–95. French
- Giersch S, Munk W, Ziegler R. 2010. The first record of a beaver – Trogontherium (Euroxenomys) minutum – in the Höwenegg fauna (Miocene, southern Germany). Palaeodiversity. 3:235–239.
- Godin AJ. 1977. Wild mammals of New England. Baltimore: Johns Hopkins University Press; p. 304.
- Grzimek B. ed. 1990. Grzimek’s animal life encyclopedia. Mammals I - IV. ed. Series. Grzimek, B. Vol. I-IV. New York: McGraw-Hill Publishing Company.
- Hartung J, Böhme M. 2022. Unexpected cranial sexual dimorphism in the tragulid Dorcatherium naui based on material from the middle to late Miocene localities of Eppelsheim and Hammerschmiede (Germany). PLOS ONE. 17(5):e0267951. [published online] 10.1371/journal.pone.0267951
- Hartung J, Lechner T, Böhme M. 2020. New cranial material of Miotragocerus monacensis (Mammalia: Bovidae) from the late Miocene hominid locality Hammerschmiede (Germany). Neues Jahrbuch für Geologie und Paläontologie Abhandlungen. 298(3):269–284. doi:10.1127/njgpa/2020/0948.
- Heidecke D, Seide P. 1989. Bisamratte Ondatra zibethicus (L.). Buch der Hege Band 1 Haarwild. Stubbe H ed., Berlin: VEB Deutscher Landwirtschaftsverlag Berlin. German. p. 640–666.
- Heinrich W-D, Maul LC. 2020. Mortality profiles of Castor and Trogontherium (Mammalia: Rodentia, Castoridae), with notes on the site formation of the Mid-Pleistocene hominin locality Bilzingsleben II (Thuringia, Central Germany). Fossil Imprint. 76(1):40–58. doi:10.37520/fi.2020.004.
- Hemprich W 1820. Grundriss der Naturgeschichte für höhere Lehranstalten. 29 pp. August Rücker, Berlin. German.
- Hír J. 2004. The Middle Miocene (late Astaracian, MN7-8) rodent Fauna of Felsötárkány 3/2 (Hungary). Acta Palaeontol Rom. 4:125–136.
- Hugueney M. 1999. Family Castoridae. In: Rössner GE, Heissig K, editors. The Miocene Land Mammals of Europe. Munich: Verlag Friedrich Pfeil; p. 281–300.
- Hugueney M, Duranthon F. 2012. Les Castoridae (Rodentia) de Sansan. In: Peigné S, editor. Mammifères de Sansan, Mémoires du Muséum National d’Histoire Naturelle. French: Publications Scientifiques du Muséum, Paris; p. 95–118.
- Hugueney M, Escuillié F. 1995. K-strategy and adaptative specialization in Steneofiber from Montaigu-le-Blin (dept. Allier, France; Lower Miocene, MN 2a, ± 23 Ma): first evidence of fossil life-history strategies in castorid rodents. Palaeogeography Palaeoclimatol Palaeoecol. 113(2–4):217–225. doi:10.1016/0031-0182(95)00050-V.
- Hugueney M, Escuillié F. 1996. Fossil evidence for the origin of behavioral strategies in early Miocene Castoridae, and their role in the evolution of the family. Paleobiology. 22(4):507–513. doi:10.1017/S0094837300016493.
- Jäger GF 1835. Über die fossilen Säugethiere, welche in Württemberg aufgefunden worden sind. 70 pp. C. Erhard, Stuttgart. German.
- Kargopoulos N, Kampouridis P, Lechner T, Böhme M. 2021a. A review of Semigenetta (Viverridae, Carnivora) from the Miocene of Eurasia based on material from the hominid locality of Hammerschmiede (Germany). Geobios. 69:25–36. doi:10.1016/j.geobios.2021.07.001.
- Kargopoulos N, Kampouridis P, Lechner T, Böhme M. 2021b. Hyaenidae (Carnivora) from the Late Miocene hominid locality of Hammerschmiede (Bavaria, Germany). Historical Biology. doi:10.1080/08912963.2021.2010193.
- Kargopoulos N, Valenciano A, Abella J, Kampouridis P, Lechner T, Böhme M. 2022. The exceptionally high diversity of small carnivorans from the Late Miocene hominid locality of Hammerschmiede (Bavaria. Germany). PLOS ONE. 17(7):e0268968. doi:10.1371/journal.pone.0268968.
- Kargopoulos N, Valenciano A, Kampouridis P, Lechner T, Böhme M. 2021c. New early late Miocene species of Vishnuonyx (Carnivora, Lutrinae) from the hominid locality of Hammerschmiede, Bavaria, Germany. Journal of Vertebrate Paleontology. 41(3): [published online] 10.1080/02724634.2021.1948858
- Kaup JJ. 1832. Beschreibung Dreyer Gattungen urweltlicher Nager des zoologischen Museums zu Darmstadt, welche von den jetzt lebenden Genera verschieden sind. Isis. IX. 992–996. German
- Kirscher U, Prieto J, Bachtadse V, Abdul Aziz H, Doppler G, Hagmaier M, Böhme M. 2016. A biochronologic tie-point for the base of the Tortonian stage in Europe terrestrial settings: magnetostratigraphy of the topmost upper freshwater molasse sediments of the North Alpine Foreland Basin in Bavaria (Germany). Newsl on Stratigr. 49(3):445–467. doi:10.1127/nos/2016/0288.
- Kordos L. 2020. Trogontherium minutum (Rodentia, Castoridae) from the late Miocene of Rudabánya, Hungary. Tertiary & Quaternary Strata. 2020(1):1–7.
- Korth WW. 2001. Occurrence of the European genus of beaver Euroxenomys (Rodentia: Castoridae) in North America. Paludicola. 3:73–79.
- Lechner T, Böhme M. 2020. Castor-like postcranial adaptation in an uppermost Miocene beaver from the Staniantsi Basin (NW Bulgaria). Fossil Imprint. 76(1):128–164. doi:10.37520/fi.2020.009.
- Lechner T, Böhme M. 2022. The beaver Steneofiber depereti from the lower Upper Miocene hominid locality Hammerschmiede and remarks on its ecology. Acta Palaeontologica Polonica. 67(4):807–826. doi:10.4202/app.00997.2022.
- López JM, Rosi MI, Tabeni S, Bender B, Chiavazza H. 2017. Taphonomic analysis of small mammal bone remains preyed upon by wildcats (Carnivora: Felidae) from the central Monte Desert (Mendoza. Argentina). Boreas. 46(2):282–293. doi:10.1111/bor.12211.
- Lyman RL 1994. Vertebrate taphonomy. Cambridge manuals in archaeology. Cambridge University Press, Cambridge, New York, Melbourne. 524 pp.
- Marin-Monfort MD, García-Morato S, Olucha R, Yravedra J, Piñeiro A, Barja I, Andrews P, Fernández-Jalvo Y. 2019. Wildcat scats: taphonomy of the predator and its micromammal prey. Quaternary Science Reviews. 225:1–16. doi:10.1016/j.quascirev.2019.106024.
- Mayet L. 1908. Étude des Mammifères Miocènes des Sables de l’Orléanais et des Faluns de la Touraine. Annales de L’Universitè de Lyon - Nouvelle Série, I. Sciences, Médecine. 24. 1–336. French
- Mayr H, Fahlbusch V. 1975. Eine unterpliozäne Kleinsäugerfauna aus der Oberen Süßwasser-Molasse Bayerns. Mitt der Bayerischen Staatssammlung für Paläontol und Historische Geol. 15. 91–111. German
- Mayr G, Lechner T, Böhme M. 2020a. A skull of a very large crane from the late Miocene of Southern Germany, with notes on the phylogenetic interrelationships of extant Gruinae. J Ornith. 161(4):923–933. doi:10.1007/s10336-020-01799-0.
- Mayr G, Lechner T, Böhme M. 2020b. The large-sized darter Anhinga pannonica (Aves, Anhingidae) from the late Miocene hominid Hammerschmiede locality in Southern Germany. PLOS ONE. 15(5):0232179. doi:10.1371/journal.pone.0232179.
- Mayr G, Lechner T, Böhme M. 2022. Nearly complete leg of an unusual, shelduck-sized anseriform bird from the earliest late Miocene hominid locality Hammerschmiede (Germany). Historical Biology. doi:10.1080/08912963.2022.2045285.
- Meyer H von. 1838. Mittheilungen, an Professor Bronn gerichtet. In Leonhard KC, Bronn HG, editors. Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefaktenkunde.(1838):413–418. German
- Mörs T, Hägglund S, Erbajeva MA, Alexeeva N, Shchetnikov AA, Daxner-Höck G. 2022. The northernmost Eurasian Miocene beavers: Euroxenomys (Castoridae, Mammalia) from Olkhon Island, Lake Baikal (Eastern Siberia). Palaeobiodivers and Palaeoenviron. 102(4):873–883. doi:10.1007/s12549-022-00555-x.
- Mott CL, Bloomquist CK, Nielsen CK. 2013. Within-lodge interactions between two ecosystem engineers, beavers (Castor canadensis) and muskrats (Ondatra zibethicus). Behav. 150(11):1325–1344. doi:10.1163/1568539X-00003097.
- Nowak RM. 1991. Walker’s Mammals of the World. Vol. I, 5th ed. Baltimore: Johns Hopkins University Press.
- Prieto J. 2009. Evolution of the genus Collimys Daxner-Hock, 1972 (Rodentia, Cricetidae) a key to Middle to Late Miocene biostratigraphy in Central Europe. Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 252(2):237–247. doi:10.1127/0077-7749/2009/0252-0237.
- Prieto J. 2012. The genus Eomyops Engesser, 1979 (Rodentia, Eomyidae) from the youngest deposits of the German part of the North Alpine Foreland Basin. Swiss Journal of Palaeontology. 131(1):95–106. doi:10.1007/s13358-011-0033-4.
- Prieto J, Casanovas-Vilar I, Gross M. 2014. Euroxenomys minutus minutus (Rodentia, Castoridae) from Gratkorn (Austria, Styria). In: Böhme M, Gross M, Prieto J, editors, The Sarmatian vertebrate locality Gratkorn, Styrian Basin. Palaeobiodiversity and Palaeoenvironments. Vol. 94(1). 163–170
- Prieto J, van Dam JA. 2012. Primitive Anourosoricini and Allosoricinae from the Miocene of Germany. Geobios. 45(6):581–589. doi:10.1016/j.geobios.2012.03.001.
- Prieto J, van den Hoek Ostende LW, Böhme M. 2011. Reappearance of Galerix (Erinaceomorpha, Mammalia) at the Middle to Late Miocene transition in South Germany: biostratigraphic and palaeoecologic implications. Contributions to Zoology. 80(3):179–189. doi:10.1163/18759866-08003002.
- Rekovets L, Stefen C, Demeshkant V. 2020. Beavers (Castoridae, Rodentia) from the late Miocene (MN 9) locality Grytsiv in Ukraine. Fossil Imprint. 76(1):165–173. doi:10.37520/fi.2020.010.
- Roger O. 1898. Wirbeltierreste aus dem Dinotheriensande der bayerisch-schwäbischen Hochebene. Bericht des Naturwissenschaftlichen Vereins für Schwaben und Neuburg. 33: 1–46. German
- Samson P, Rădulesco C. 1973. Rémarques sur l’évolution des Castoridés (Rodentia, Mammalia). In: Orghidan T, editor. Livre du cinquantenaire de L’Institut de Spéléologie “Emile Racovitza”. French: Academiae Republicii Socialiste România, Bucarest. Romanian; p. 437–449.
- Schreuder A. 1929. Conodontes (Trogontherium) and Castor from the Teglian clay compared with the Castoridae from other localities. Archives du Musee’ Teyler. 3:99–320.
- Stefen C 1997. Steneofiber eseri (Castoridae, Mammalia) von der Westtangente bei Ulm im Vergleich zu anderen Biberpopulationen. Stuttgarter Beiträge zur Naturkunde - Serie B (Geologie und Paläontologie) 255:1–73. German.
- Stefen C. 2001. Barstovian (Miocene) beavers from Stewart Valley, Nevada, and a discussion of the genus Monosaulax based on tooth morphology. PaleoBios. 21(1):1–14.
- Stefen C. 2009. The European Tertiary beaver Chalicomys jaegeri (Rodentia: Castoridae) revisited. Kaupia - Darmstädter Beiträge zur Naturgeschichte. 16:161–174.
- Stefen C. 2018. The castorids (Mammalia, Castoridae) from the (early) middle Miocene of Gračanica (Bosnia-Herzegovina). Palaeobiodivers and Palaeoenviron. 100(2):301–305. doi:10.1007/s12549-018-0365-9.
- Stefen C, Mörs T. 2008. The beaver Anchitheriomys from the Miocene of Central Europe. J Paleontol. 82(5):1009–1020. doi:10.1666/06-049.1.
- Stefen C, Rummel M. 2003. Trogontherium (Euroxenomys) minutum (Castoridae: Mammalia) from Petersbuch 50, Miocene, South Germany. Neues Jahrb für Geol und Paläontol, Monatsh. 2003(1):11–34. doi:10.1127/njgpm/2003/2003/11.
- Stiner MC. 1990. The use of mortality patterns in archaeological studies of hominid predatory adaptations. Journal of Anthropological Archaeology. 9(4):305–351. doi:10.1016/0278-4165(90)90010-B.
- Stirton RA. 1935. A review of the tertiary beavers. Univ Calif Publ Geol Sci. 23:391–458.
- Voorhies MR. 1969. Taphonomy and population dynamics of an early Pliocene vertebrate fauna, Knox county, Nebraska. Rocky Mountain Geol. 8:1–69.
- Williams SA, Prang TC, Meyer MR, Russo GA, Shapiro LJ. 2020. Reevaluating bipedalism in Danuvius. Nature. 586(7827):E1–E3. doi:10.1038/s41586-020-2736-4.
- Willner GR, Feldhamer GA, Zucker EE, Chapman JA. 1980. Ondatra zibethicus. Mamm Species. 141(141):1–8. doi:10.2307/3504016.