Abstract
The planktonic diatom Fragilariopsis kerguelensis plays an important role in the biogeochemical cycles of the Southern Ocean, where remains of its frustules form the largest deposit of biogenic silica anywhere in the world. We assessed the genetic identity of 26 strains, from cells collected at various sites in the Southern Ocean, using three molecular markers, LSU and ITS rDNA and rbcL. The LSU sequences were identical among the tested strains, ITS sequences were highly similar, and only one base pair difference was detected among the rbcL sequences. These results, together with a large number of successful mating experiments demonstrated that the strains belong to a single biological species. We investigated the mating system and life cycle traits of F. kerguelensis. Cell size diminished gradually in clonal strains. Gamete formation only occurred when strains of opposite mating type – within a cell size range of 7–36 µm – were mixed together. Two binucleate gametes were formed in each gametangium and gamete conjugation produced a zygote that had four nuclei and was surrounded by thin siliceous scales. Two out of the four nuclei subsequently degenerated and the zygote expanded to form an auxospore surrounded by a transverse and a longitudinal perizonium. Staining with the fluorochrome PDMPO provided for the first time a clear demonstration that the longitudinal perizonium is formed after auxospore expansion is complete. Initial cells produced within the mature auxospores were 78–101 µm in length. Various authors have shown that the average valve size of F. kerguelensis varies in sediment samples collected in regions and seasons with different primary production regimes and this parameter has thus been proposed as a biological proxy for palaeo-productivity. A better understanding of the life cycle of F. kerguelensis should help the design of future investigations aimed at testing the link between cell size distribution in the natural environment and the role that environmental factors might have in the regulation of population cell size.
Introduction
The pennate diatom genus Fragilariopsis includes species that are important primary producers in the marine plankton and in the sea ice of polar seas (Hustedt, Citation1958; Lundholm & Hasle, Citation2010). In the Southern Ocean, the heavily silicified species Fragilariopsis kerguelensis has been reported as among the dominant diatoms in the water column of the ice-free, offshore Antarctic Circumpolar Current (ACC) (Hart, Citation1942; Smetacek et al., Citation2004).
Cells of Fragilariopsis kerguelensis can form long, ribbon-shaped chains, and they can be distinguished from other species in the genus by the strong silicification of their frustule and coarse pattern of the striae on the valve face (Hasle, Citation1965, Citation1968; Hasle & Syvertsen, Citation1997; Cefarelli et al., Citation2010). Mechanical stress-testing of frustules of live cells of F. kerguelensis have demonstrated that the frustule architecture provides high mechanical resistance to deformation and breakage, which is probably an adaptation against grazing (Hamm et al., Citation2003). The robust nature of the F. kerguelensis frustule is further reflected in the exceptionally high Si : N ratios of this species relative to most other diatoms (Brzezinski, Citation1985; Hoffmann et al., Citation2007). Together, these morphological and physiological characteristics demonstrate why this species has such a high demand for silica.
Fragilariopsis kerguelensis contributes up to 90% of the diatom frustules in the diatom ooze making up the Antarctic opal belt (Zielinski & Gersonde, Citation1997), which is the largest deposit of biogenic silica in the world ocean (Treguer et al., Citation1995). Therefore, F. kerguelensis is one of the most important diatoms in the global silicon cycle (Zielinski & Gersonde, Citation1997) and an indicator species of a silica-sinking regime in an otherwise iron-limited ecosystem (Smetacek et al., Citation2004). Valve size measurements of F. kerguelensis from sediment samples of different sites in the Atlantic and Pacific sector of the Southern Ocean and from sediment cores spanning the Holocene and the last glacial maximum (LGM) showed the presence of larger frustules close to the highly productive Antarctic Polar Front (APF) and in the LGM layers, while smaller frustules were recorded in the less productive zones north and south of the APF and in the interglacial layers (Cortese & Gersonde, Citation2007). Further size distribution measurements of F. kerguelensis in core samples of the Holocene showed the presence of longer and more abundant valves during the warmer Mid-Holocene period and of smaller and less abundant ones during the colder Late-Holocene period (Crosta, Citation2009). These results led the authors to propose F. kerguelensis as a biological proxy for palaeo-productivity and temperature conditions in the Southern Ocean. Productivity in this area is strongly linked to iron availability and an increased abundance of F. kerguelensis in response to artificial iron addition has been recorded in three Southern Ocean iron-fertilization experiments (Gall et al., Citation2001; Assmy et al., Citation2006, Citation2007).
Variations in valve size over geological time scales, as well as short-term changes resulting from iron addition experiments (Cortese et al., Citation2012), prompted us to investigate the life cycle of this important diatom. Changes in cell size frequency distribution (SFD) in many diatoms are linked to the functioning of the life cycle. Part of the timing mechanism is related to the morphology of the cell wall, which consists of two rigid siliceous thecae that are slightly different in size. This results in a progressive decrease of the average population cell size as vegetative division proceeds. This progressive reduction in cell size – mostly along the apical axis in pennate diatoms – can be circumvented by the onset of sexual reproduction and the formation of large-sized cells (Round et al., Citation1990; Chepurnov et al., Citation2004). In situ evidence for sexual reproduction comes either from direct observation of sexual stages (e.g. Crawford, Citation1995; Sarno et al., Citation2010; Holtermann et al., Citation2010) or from estimates of cell size spectra, where the detection of larger cell size classes is an indirect evidence for recent sexual events (e.g. Mann, Citation1988; Jewson, Citation1992; D’Alelio et al., Citation2010). Despite the ubiquity of F. kerguelensis in the Southern Ocean, evidence of sexual reproduction – e.g. the presence of gametangia, auxospores and initial cells – in field samples was reported for the first time only recently (Assmy et al., Citation2006).
The first aim of our study was to test if F. kerguelensis is a genetically homogeneous species or if it includes genetically different cryptic species. Cryptic diversity, i.e. morphologically similar or identical but genetically different species, have been detected within several diatoms (e.g. Amato et al., Citation2007; Sarno et al., Citation2007; Evans et al., Citation2008; Quijano-Scheggia et al., Citation2009). To this end, we obtained both nuclear ribosomal (LSU and ITS rDNA) and plastid (rbcL) sequences from strains isolated in different locations of the Atlantic sector of the Southern Ocean.
The second aim was to study the life cycle of F. kerguelensis in laboratory conditions, testing its mating system and the way in which gametes, auxospores and the large initial cells are produced. Insight into the mechanisms through which sexual reproduction occurs is a prerequisite for further investigations aimed at testing the link between cell size distribution in the natural environment and the role of environmental factors on the regulation of population cell size.
Materials and methods
Culture material
A total of 51 cultures of Fragilariopsis kerguelensis (Supplementary and Supplementary ) were established during cruises ANT-XXI/3 (EIFEX, European Iron Fertilization EXperiment: January–February 2004) and ANT-XXV/3 (LOHAFEX, ‘loha’ is the Hindi word for iron, Fertilization EXperiment: January–March 2009) carried out in the Atlantic sector of the Southern Ocean. Single cells or short chains were isolated with a micropipette from phytoplankton samples collected with a 20 µm-mesh-size phytoplankton hand net. At the Alfred Wegener Institute (AWI), strains were grown in 70-ml tissue culture bottles filled with 30 ml of f/2 medium (Guillard & Ryther, Citation1962) prepared with 0.2 µm-filtered and autoclaved Antarctic seawater (at a salinity of 34.6), modified to obtain a higher concentration of silicic acid (210 µmol l–1) and a lower concentration of nitrate (100 µmol l–1) and phosphate (6.25 µmol l–1). Strains were maintained at a temperature of 2°C, an irradiance of 50 µmol photons m–2 s–1 provided by Osram Biolux lamps (Osram L18W/965 Biolux: OSRAM, Munich, Germany), and a photocycle of 16 : 8 h light : dark. At the Stazione Zoologica Anton Dohrn (SZN), strains were grown in standard f/2 culture medium prepared with 0.2 µm-filtered and autoclaved oligotrophic Mediterranean seawater adjusted to a salinity of 35. Cultures were maintained at a temperature of 5°C, an irradiance of 50 µmol photons m–2 s–1, provided by Philips (Royal Philips, Amsterdam, the Netherlands) cool white lamps, and a photocycle of 12 : 12 h light : dark.
Table 1. The results of crossing experiments carried out with different strains of Fragilariopsis kerguelensis; in the first column and in the first row the strain code, in the second column the mating type (m.t., see text for further details), in the third column the average apical length of the strains (Size). Grey-filled squares indicate the absence of intraclonal sexual reproduction; + indicates the presence of sexual stages; 0 indicates the absence of sexual stages; n.a. = not available.
Figs 1–13. Light (Figs 1–4) and SEM (Figs 5–13) micrographs of Fragilariopsis kerguelensis vegetative cells. 1–3. Cells of different apical length arranged in chains; an arrowhead points to the nucleus (Fig. 1, strain PA_P6C3; Fig. 2, strain MM_P7A4; Fig. 3, strain MM_E8A6). 4. A single cell in valvar view (strain MM_E13C5). 5. A detail of a chain in which mucous material is visible between the valves of adjacent cells (large F1 generation cells of cross PA_P8B1 × MM_P13D2, experiment B). 6. An isopolar valve; the narrow raphe on the valve margin is arrowed (natural sample). 7. Epi- and hypotheca of a dividing cell showing the valvocopula and the two cingular bands; the diagonal suture in the valvocopula is marked with an arrow and the ligula on the second, thin cingular band is marked with arrowheads (cross PA_P8B1 × MM_P13D2, experiment B). 8. A detail of Fig. 7. 9. Epitheca showing valvocopula and two cingular bands; note the diagonal suture in the valvocopula (arrow) and the ligula on the narrow second cingular band (arrowhead) (cross PA_P8B1 × MM_P13D2, experiment B). 10. Detail of Fig. 9. 11. A heteropolar valve (strain Lynn 5). 12. Detail of the valve with the poroids (strain Lynn 5). 13. Detail of the same valve, showing a single poroid with minute perforations. Scale bars = 10 µm (Figs 1–5, 7, 11), 5 µm (Figs 6, 8, 9), 1 µm (Figs 10, 12) and 100 nm (Fig. 13).

DNA extraction, amplification and sequence analysis
Cultures of F. kerguelensis were harvested during the exponential growth phase by filtration on 0.45 µm pore-size polycarbonate filters. DNA extraction and purification were performed as described by Kooistra et al. (Citation2003). PCR amplification of the hyper-variable D1–D3 domains in the nuclear-encoded LSU ribosomal RNA region, the nuclear-encoded internal transcribed spacer region (ITS1, 5.8S rDNA and ITS2), and the plastid-encoded rbcL region were carried out as described by Amato et al. (Citation2007). Sequence reactions were obtained with Big-Dye Terminator Cycle Sequencing technology (Applied Biosystems, Foster City, California, U.S.A.) and purified using a robotic Biomek FX station (Beckman Coulter, Fullerton, California, U.S.A.). Products were analysed on an Automated Capillary Electrophoresis Sequencer 3730 DNA Analyzer (Applied Biosystems). Sequences were aligned by eye with sequences downloaded from GenBank, using the sequence alignment editor Se-Al version 2.0a11 (Rambaut, Citation2002). The alignment of the ITS sequences is provided as Supplementary file 1. M-fold (http://mfold.rna.albany.edu) was used to explore the various possible secondary structure solutions for the ITS-2 sequences of the strains.
Life cycle
Cell size reduction
A first estimate of the monthly cell size reduction rate of F. kerguelensis was obtained using two time-point measurements for 47 strains with average apical length spanning 13–76 µm kept in the AWI standard growth conditions. The apical lengths of five cells for each strain were measured at time 0 and after 24 months.
Seven strains of F. kerguelensis (marked with § in Supplementary ), kept in the SZN growth conditions, were used to estimate cell size reduction rates. The apical length of 300 cells for each strain was measured at the beginning and at the end of the observation period (seven months); the monthly reduction rate of the apical length was estimated as the difference between the two average measures. The growth rate of the seven strains was measured as well to obtain an estimate of cell size reduction at each cell division. At the beginning of the observation period, an aliquot (between 1 and 3 ml, depending on cell concentration) of the exponentially growing strains was inoculated into glass tubes (three tubes for each strain) filled with 20 ml of culture medium. Growth was estimated by measuring fluorescence with a Turner Designs fluorometer model 10-005R (Turner Designs, Sunnyvale, California, U.S.A.) every two or three days. Cultures were monitored for 24 days, i.e. until the end of the exponential growth phase. The growth rate, as divisions·day–1, was estimated by calculating the linear regression over all data points between day 0 and 24. The growth curves of the seven strains were rather similar and the exponential growth phase was completed between day 18 and day 23. We have chosen to estimate growth rates over 24 days, because this was the average time interval (± 1 day) at which cultures were transferred during the experiment (lasting seven months).
Mating behaviour
Mating-type designation of F. kerguelensis was done on the basis of the results of two sets of crosses: in the first set, seven strains and in the second set 14 strains were mixed in all pairwise combinations (marked with * in Supplementary ). Crosses were carried out in 12-well tissue culture plates (Corning, Corning, New York, U.S.A) filled with 1 ml of modified f/2 medium. In each well, pairs of exponentially growing strains were added (c. 0.5 ml of each strain). Control wells were inoculated with double volume of a single strain. Culture plates were incubated in the AWI experimental conditions specified above and were checked every two days for the presence of sexual stages.
Sexual life cycle
Detailed observations on the life cycle of F. kerguelensis were carried out at different times and using different strains as follows: (1) experiment A was carried out in February 2009 with up to six strains mixed in the same culture plate (strains whose code starts with ‘L’ in Supplementary ; the average apical length of these strains was between 10 and 25 µm); (2) experiment B was carried out in August 2011 with strains PA_P8B1 (mean apical length: 16 µm) and MM_P13D2 (mean apical length: 15 µm); (3) experiment C was carried out in July 2010 with strains L2D6 (average apical length: 12.78 ± 1.09 µm, mean ± s.d.) and L9C3 (average apical length: 14.26 ± 1.60 µm).
In experiment A (carried out on board RV Polarstern), the wells of a 6-well culture plate were filled with exponentially growing cultures – 0.5 ml for each strain – of four or six different strains isolated during the ANT-XXV/3 cruise and 0.5 ml of growth medium. The culture plate was incubated at 5°C, 50 µmol photons m–2 s–1 and an 18 : 6 h L : D photocycle, and inspected daily for the presence of sexual stages. During the first days in which sexual stages were recorded, the material of one well was stained with the fluorochrome PDMPO [2-(4-pyridyl)-5-((4-(2-dimethylaminoethylaminocarbamoyl)methoxy)phenyl)oxazole: Molecular Probes, Eugene, Oregon, U.S.A.), following the protocol by Leblanc & Hutchins (Citation2005), to visualize newly formed silica frustule elements of the vegetative cells and auxospores.
In experiment B (carried out at AWI), strains PA_P8B1 and MM_P13D2 were mixed and dispensed into a 12-well tissue culture plate (Corning, Corning, New York, U.S.A.) and 12 Utermöhl chambers. All vessels were incubated in the standard AWI conditions and inspected twice per day for 10 days. At every inspection, the material from one Utermöhl chamber was fixed with hexamethylenetetramine-buffered formaldehyde at a final concentration of 2% and stained with DAPI (4′,6-diamidino-2-phenylindole; Sigma–Aldrich, St. Louis, Missouri, U.S.A.) at a final concentration of 0.5 µg ml–1 to visualize the nuclei. On day 6, when sexual stages were detected in the culture, the material from three culture wells was incubated with PDMPO for 12 h.
In experiment C (carried out at SZN), a stock culture was prepared with 150 ml of f/2 culture medium in which exponentially growing strains L2D6 and L9C3 were inoculated together to reach a final concentration of about 4000 cells ml–1 (i.e. 2000 cells ml–1 for each mating type); aliquots of 4 ml were dispensed in two 6-well tissue culture plates and incubated at the standard SZN culture conditions. Four ml of each parental strain were placed in a plastic vial and fixed with formaldehyde solution at a final concentration of 1.6%; these two samples were used to estimate the percentage composition of single cells and cells arranged in chains in the parental strains. On day 7, when cells were observed detaching in the dual cultures approaching the sexual phase, the material from three wells was fixed with formaldehyde; this sample was used to estimate the percentage composition of single cells and cells arranged in chains. On each of the subsequent three days, the material of two wells was pooled in vials, fixed, and used for the observation of sexual stages. The fixed samples were stained with DAPI as described above.
The culture material from all three experiments was examined and photographed with a Zeiss Axiovert 200 epifluorescence microscope (Zeiss, Oberkochen, Germany) equipped with a Zeiss AxioCam, a long pass DAPI filter set (EX G 365, BS FT 395, EM LP 420) and a band pass DAPI filter set (EX G365, BS FT 395, EM BP 445/50).
Scanning electron microscopy
For observation of different life stages by scanning electron microscopy, the material from experiments B and C was fixed with glutaraldehyde at a final concentration of 2% at 4°C for 1 h. The fixed samples were gently filtered over a membrane filter placed in a filter holder connected to a syringe and subsequently dehydrated with ethanol at increasing concentrations (10%, 25%, 50%, 75%, 95%, 2 × 100%) for at least 10 min for each step. After using 100% ethanol, the filter was placed on an aluminium stub, critical point dried, and sputter coated with gold-palladium. Samples were observed with a FEI Quanta 200F (FEI, Hillsboro, OR, USA) SEM at AWI or a JEOL 6700-F (JEOL, Tokyo, Japan) at SZN. Two natural samples collected during the Lohafex cruise were also prepared for SEM, using the protocol described above.
Results
Genetic and morphological characterization of Fragilariopsis kerguelensis
The LSU sequences of the 26 analysed strains were identical. The rbcL region was sequenced for 11 strains and sequences were identical with one exception: two strains (MM_E13C5 and MM_E13B2) showed a C at position 1074 in the gene region (1055 in our alignment) whereas the remainder showed an A at that position. The ITS sequences of the 11 tested strains were also highly similar. Differences were restricted to 13 positions in the alignment, exhibiting base changes or ambiguities, but without any phylogenetic pattern (Supplementary ). One of these positions, position 115 from the 5′-end of ITS-2, exhibited either an A, a T or a W (= ambiguity A and T). The various alternative RNA folding patterns provided by M-fold generally located this position against a U at position 249 from the 5′-end of ITS-2 in a stem region, flanking a small internal loop. Therefore, an A at position 115 in ITS-2 resulted in an A–U bond whereas a U at that position increased the size of the internal loop.
Table 2. Rate of reduction in cell size (apical length) and growth rate for seven Fragilariopsis kerguelensis strains. The cell size is the average cell size on day 0 and the growth rate is the average of three measurements.
The apical length of the strains used in this study ranged from 70 µm (strain PA_P10C3) to less than 10 µm, the average cell length below which strains died. Cells were single () or arranged in chains of different length (–3), where cells were joined together valve to valve by mucilaginous material (). In smaller cells (up to 15–20 µm long), the contact between neighbouring cells was along the whole surface of the two adjacent valves, while longer cells were in contact along the central portion of the cells; this suggests that larger cells are slightly inflated in their central portion in girdle view (). The nucleus was situated in the central part of the cell, with two large chloroplasts located on either side ().
The frustule included two thecae, each with a valve, a valvocopula and two open cingular (girdle) bands (–). The opposite ends of the valvocopula were pointed and joined forming a diagonal suture on the portion of the cingulum close to the cell apex (). The first cingular band was hairpin-shaped and its ends were not in direct contact. The second band was very thin and its ends were almost in contact on the narrow side of the cingulum; a ligula was present in its central portion, on the other narrow side of the cingulum (, ). This ligula was inserted between the ends of the first cingular band. A thin raphe ran along the margin of each valve and the raphes of the two valves of each frustule were in the trans positions (), i.e. the raphes lay diagonally opposite each other in intact frustules (nitzschioid symmetry). Large cells were lanceolate in valvar view, with a heteropolar outline (), while smaller cells were isopolar (). The number of striae in 10 µm ranged between 6 and 8 and the number of poroids in 10 µm ranged between 8 and 10. The poroids were 200–300 nm in diameter and were closed by a hymenate thin layer with minute perforations (, ).
Mating behaviour
Gametangia, auxospores and initial cells were never observed in clonal strains, but were only observed when mixing strains of compatible mating type. The average cell lengths of parental strains that were involved in successful crosses ranged between 12 and 36 µm (). The results of the first set of crosses (upper matrix in ) involving seven strains revealed that three of them belonged to one mating type [defined as Fk1, following the recommendation of Chepurnov et al. (Citation2005)] and the other four to the opposite mating type Fk2. All combinations between crosses of the opposite mating type produced auxospores and viable initial cells. In the second set of crosses carried out with 14 strains (lower matrix in ), nine of them belonged to one mating type and five to the opposite one. In this case, sexual stages were recorded in all combinations of opposite mating types except one (L8B1 × L8B6). Assignment of the mating type in F. kerguelensis is only based on the results provided by the matrix of crosses because no morphological or behavioural differences could be detected between strains of different mating type (see the following section). The designations Fk1 and Fk2 thus denote only that strains belong to opposite mating types, without assigning any biological meaning, i.e. ‘male’ vs ‘female’ or ‘active’ vs ‘passive’.
Life cycle
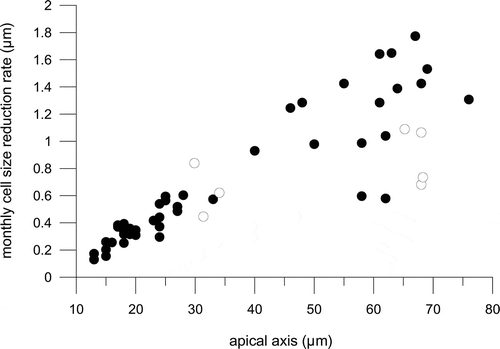
All clonal strains showed a progressive reduction of the apical axis of the cells over time. Cell size reduction estimates based on measurements of 47 cultivated strains over a duration of 24 months showed that the average reduction in apical length was 0.13–1.77 µm per month and it was larger for longer cells (≥ 50 µm long). However, considerable variability in cell size reduction rates was detected in these longer cells (regression coefficient for size reduction vs length = 0.11; ), whereas for cells ≤ 50 µm a tighter linear relationship was detected between average cell size and the monthly reduction rate (regression coefficient: 0.9; ). Seven strains with an average apical length spanning 29 and 68 µm were measured after a time interval of seven months and showed average cell size reduction rates between 0.45 and 1.06 µm∙month–1 (, , Supplementary ). The average growth rates estimated over 24 days ranged from 0.076 to 0.121 divisions day–1 and the rate of length reduction per division was 0.18–0.47 µm ().
Fig. 14. Average monthly reduction rate of the apical length of Fragilariopsis kerguelensis clonal strains of different cell size; black circles represent the 47 strains measured after a time interval of 24 months, white circles represent the seven strains measured after a time interval of 7 months.
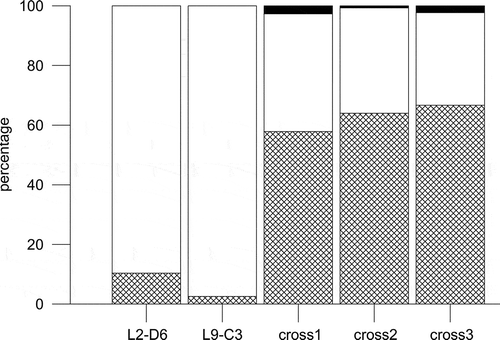
The first sign of interaction between strains of opposite mating type (within a few days of inoculation in the same culture vessel) was the detachment of the cells from the chain (). A quantitative estimate was obtained for the cross L2-D6 × L9-C3. About 90% of the cells were arranged in chains in the individual parental strains, but on day 7 after the mixing of the two parental strains, when the first sexual stages were detected, only 30% of the cells were arranged in chains (). A similar timing and behaviour was observed also in the cross PA_P8B1 × MM_P13D2, where most of the cells were detached within three to four days after inoculation. After detachment from the chains, single cells increased mobility and several formed pairs (). The contact point between paired cells was variable. Most of the time cells were in contact at the level of the cingular bands, but their orientations varied; in some cases the valves were parallel to each other, in other cases perpendicular ( and ). The observation in SEM of a sample from cross PA_P8B1 × MM_P13D2 containing numerous paired cells showed the presence of mucous threads joining cells along the cingular bands (, ). At the stage in which chains were disassembled, many cells – either paired or single – showed a considerably enlarged nucleus (). In these cells, the two chloroplasts were appressed to the cell wall. These cells expanded along their pervalvar axis by the deposition of additional cingular bands in the hypotheca (, , ) and were interpreted as gametangia. Figure 22 shows the hypovalve of the parental gametangium still connected to the auxospore.
Fig. 15. Percentage distribution of single cells (grids), cells in chains (white) and sexual stages (black) in monoclonal parental strains (L2-D6 and L9-C3) of Fragilariopsis kerguelensis and in three replicate crosses on day 7 after inoculation.
Figs 16–22. Light (Figs 16, 17) and SEM (Figs 18–22) micrographs of gametangia of Fragilariopsis kerguelensis. 16. A few days after the inoculation together of strains of opposite mating type: cells have detached from chains and several cells are in contact (arrows) (mixture of different strains in experiment A). 17. Picture taken on the same day as , where single cells in girdle view have an enlarged nucleus (arrowhead) and chloroplasts appressed to the valves (experiment A). 18. Two cells (gametangia) close to and perpendicular to each other, in contact via the cingulum (cross PA_P8B1 × MM_P13D2, experiment B). 19. Detail of the same pair of cells showing the mucous material extruded from the girdle region that keeps the two adjacent cells together. 20. A gametangium in girdle view with extra cingular bands in the hypotheca (arrows) (cross L2D6 × L9C3, experiment C). 21. Gametangium with extra cingular bands in the hypotheca (cross PA_P8B1 × MM_P13D2, experiment B). 22. An auxospore and three gametangial hypothecae with extra cingular bands (cross L2D6 × L9C3, experiment C). Scale bars = 20 µm (, ), 5 µm (, –) and 1 µm ().

In the gametangia, the two-step meiotic division took place. Meiosis I was followed by cytokinesis (, ), but the two took place without an accompanying cell division (acytokinetic division) and, in several cases, it was not synchronous in the two protoplasts within one gametangium. After completion of meiosis II, two binucleate gametes were formed (–). In most cases, the two paired gametangia remained connected during meiotic division. The gametangium opened along the cingular bands and the two gametes became exposed, but each still lay within one gametangial theca (, ). We could not follow gamete conjugation in real time and we cannot state if conjugation occurred between gametes while they were still situated within the appressed gametangia, or if the gametes escaped from the gametangial thecae and moved towards other gametes. We observed only a few naked binucleate stages (gametes) in the samples and we thus hypothesize that conjugation occurred between gametes still associated closely with the appressed gametangia. There appeared to be no mucous envelopes surrounding the paired gametangia.
Figs 23–41. Light and epifluorescence micrographs of auxosporulation in Fragilariopsis kerguelensis. 23. Two appressed gametangia in girdle view; the cell on the right shows the cleavage of cytoplasm (cytokinesis) (cross L2D6 × L9C3, experiment C). 24. The same gametangia with DAPI-stained nuclei; the cell on the left has an expanded nucleus in meiotic prophase, the cell on the right has undergone the second meiotic division and each protoplast has two nuclei. 25. A gametangium after completion of cytokinesis and the formation of two gametes (cross L2D6 × L9C3, experiment C). 26. The same gametangium with DAPI-stained nuclei; each gamete has two nuclei. 27. A zygote still attached to a theca of the gametangium (cross L2D6 × L9C3, experiment C). 28. The same zygote with four DAPI-stained nuclei (arrowheads). 29. A zygote still attached to a theca of the gametangium (cross PA_P8B1 × MM_P13D2, experiment B). 30. The same zygote stained with PDMPO, showing thin siliceous platelets in its wall (arrowheads). 31. A young auxospore still attached to one theca of a gametangium (cross PA_P8B1 × MM_P13D2, experiment B). 32. The same auxospore stained with PDMPO, showing thin siliceous platelets at both ends of the auxospore (arrowheads). 33. An elongating auxospore still attached to one theca of a gametangium (mix of different strains in experiment A). 34. The same auxospore with two DAPI-stained nuclei. 35. An elongated auxospore still attached to the theca of a gametangium; mix of different strains in experiment A). 36. The same auxospore stained with PDMPO, showing the bands of the transverse perizonium. 37. An elongated auxospore still attached to the theca of a gametangium (mix of different strains in experiment A). 38. The same auxospore stained with PDMPO, showing the terminal bands of the transverse perizonium (arrowheads) and the longitudinal perizonium. 39. An elongated auxospore; the sample was incubated with PDMPO and the internal longitudinal perizonium, the thin bands of the transversal perizonium and the terminal caps (arrowheads) are visible (cross PA_P8B1 × MM_P13D2, experiment B). 40. A mature auxospore within which one valve of the initial cell has been deposited (mix of different strains in experiment A). 41. The same auxospore with two DAPI-stained nuclei and PDMPO-stained valve (on the top side). Scale bars = 10 µm.
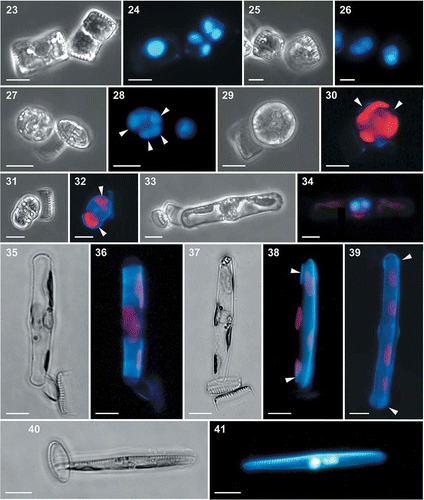
Rounded stages with four nuclei were observed, almost always attached to the empty valve of one gametangium (–). These stages were interpreted as zygotes. Support for this conclusion also derived from the observation of samples incubated with PDMPO, which showed the presence of thin, slightly silicified scales on the outer membrane of the rounded zygote () and on both ends of the auxospore (, ). SEM preparations showed that the caps at both ends of the auxospore consisted of thin, tightly appressed and partially overlapping round scales with diameters of 2.3–3.7 µm (). The zygote had four nuclei () and the elongating auxospore had two (, ); we thus assume that the degeneration of the two supernumerary nuclei occurred in the young auxospore. The vast majority of auxospores remained attached by a mucous plug to a valve of one gametangium until they had completed development. The incubation of samples with PDMPO allowed visualization of the formation of the transverse (, ) and longitudinal perizonium (, ). The transverse perizonium, which allows the bipolar expansion of the auxospore, was composed of a series of slightly silicified perizonial bands (, , , ). The longitudinal perizonium was synthesized when the auxospore had completed elongation and when the cytoplasm started detaching from the transverse perizonium (, ). Figures 37 and 38 show that, within the period of incubation with PDMPO (12 h), the auxospore synthesized the distal bands of the transverse perizonium and part of the longitudinal perizonium, which is presumably composed by thin longitudinal bands, though the exact number could not be determined. Figures 40 and 41 represent an auxospore in which one valve has been formed during incubation with PDMPO. This cell has two nuclei and one of them seems to be degenerating: we interpret this as the acytokinetic mitotic division that accompanies the deposition of one valve of the initial cell. While still wrapped by the perizonium, the initial cell contained a single nucleus (, ).
Figs 42–45. Auxospores of Fragilariopsis kerguelensis. 42. Light micrograph of a mature auxospore containing the initial cell (mix of different strains in experiment A). 43. Epifluorescence micrograph of the same auxospore showing the fusion of the two DAPI-stained nuclei. 44. SEM micrograph of an auxospore (cross L2D6 × L9C3, experiment C), showing caps at the ends and the transverse perizonium (the faint rings around the auxospore: cross L2D6 × L9C3, experiment C). 45. A detail of showing round scales covering the terminal portion of the auxospore. Scale bars = 10 µm (Figs 42–44) and 1 µm ().

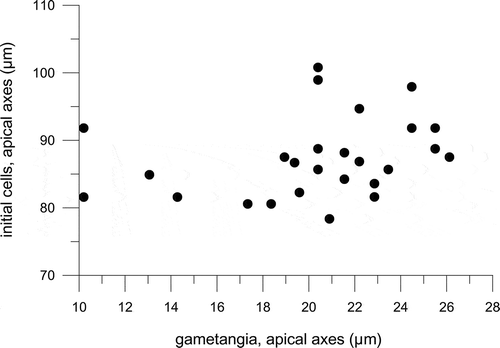
The average cell length of parental strains that produced sexual stages, both in the crosses for testing mating behaviour () and in those carried out to study the life cycle, ranged between 7 and 36 µm. We also measured the sizes of 57 empty gametangial thecae still connected to auxospores or initial cells in crosses carried out within experiment A: their lengths were 6.9–26.1 µm. Within the same experiment, we also measured the length of 27 initial cells, i.e. cells still wrapped in the perizonium but in which the siliceous valves were already visible, and of the nearby gametangial theca: initial cells measured 78.4–100.8 µm and gametangia 10.2–26.1 µm ().
Fig. 46. The lengths of 27 initial cells, still wrapped in their perizonium, and the corresponding gametangial thecae attached to them, measured in crosses carried out in experiment A.
Discussion
The results obtained with the different genetic markers showed no evidence for cryptic diversity in Fragilariopsis kerguelensis. All tested strains shared the same LSU and their highly similar ITS sequences differed only in positions showing variation between a particular base and an ambiguity consisting of that same base and another base, and in one case, variation between A, T and ambiguity W. These few variable positions did not reveal any phylogenetic pattern among the sequences of F. kerguelensis presented. Both markers have been used extensively in assessing phylogenetic relationships amongst closely related diatom species and for barcoding purposes, and they have proved to represent reliable markers for species identification, although with slightly different results amongst the taxa tested (e.g. Moniz & Kaczmarska, Citation2010; Hamsher et al., Citation2011). Amato et al. (Citation2007) demonstrated that strains of Pseudo-nitzschia with identical ITS sequences were reproductively compatible if crossed with strains of the opposite mating type. Far more pronounced sequence differences than the ambiguities observed among the ITS-sequences from strains of F. kerguelensis were observed among strains within Pseudo-nitzschia multistriata, but even these strains were sexually perfectly compatible as long as they were of the opposite mating type (D’Alelio et al., Citation2009b). The single base-pair difference in the rbcL sequences among the tested strains cannot be considered as a proof of species difference either, because Levialdi Ghiron et al. (Citation2008) have shown that strains of Pseudo-nitzschia exhibiting a total of four base pair differences among their rbcL sequences mated successfully. Comparison of Fragilariopsis and Pseudo-nitzschia is sound because these two planktonic genera are close relatives (Lundholm et al. Citation2002). The results of the molecular analyses, together with the large number of successful mating experiments carried out in this study, demonstrate that all the tested F. kerguelensis strains belong to a single biological species.
The gross morphology of the cell and the valve ultrastructure of the strains examined in light and electron microscopy fit the descriptions provided for F. kerguelensis by different authors (Hasle, Citation1965, Citation1968; Hasle & Syvertsen, Citation1997; Cefarelli et al., Citation2010). The ultrastructure of the poroid hymen of our specimens corresponds perfectly with the TEM images provided by Hasle (Citation1965), thus further confirming the uniformity of the species. The pioneering work carried out by Hasle (Citation1965, Citation1968) did not include information on the number and ultrastructure of the girdle bands. Recently, Cefarelli et al. (Citation2010) reported the presence of one valvocopula and one girdle band in F. kerguelensis. We here provide evidence for the presence of an additional thin band at the abvalvar end of the cingulum; this band bears a ligula that inserts between the open ends of the first cingular band. Similar narrow bands have been detected in other Bacillariaceae, e.g. some Nitzschia species (Trobajo et al., Citation2013).
Gradual cell size reduction was detected in all clonal strains of F. kerguelensis and the results of mating experiments showed that the sexual phase was induced only when compatible strains within the gametangial size window were mixed together. These facts indicate that F. kerguelensis is heterothallic. Furthermore, we never observed the formation of auxospores nor detected a sudden increase in cell size in any of the clonal cultures (which were monitored for a long time), indicating the absence of homothallic or uniparental (automictic or apomictic) auxosporulation (Kaczmarska et al., Citation2013). Fragilariopsis kerguelensis is the only species in the genus Fragilariopsis for which information on the life cycle is available (Assmy et al., Citation2006; this study) and its general pattern conforms to what has been described for other raphid pennate diatoms (Chepurnov et al., Citation2004). Nevertheless, differences occur between genera and in the following we discuss the main features of the life cycle in the frame of the available literature information.
Transition from the vegetative to the sexual phase and formation of gametes
The first evidence of interactions between strains of opposite mating type placed in the same culture vessel was the detachment of chains into single cells. When joined in chains, cells of F. kerguelensis in the cell size range for sexualization are in close contact along the whole surface of their valves and it is reasonable to assume that this arrangement would impede the contact between individual cells that precedes the differentiation of gametangia and their subsequent conjugation. The detachment of cells arranged in chains has been reported for other chain-forming planktonic diatoms, such as Pseudo-nitzschia pungens (Chepurnov et al., Citation2005) and P. multistriata (E. Scalco, unpublished data). However, in Pseudo-nitzschia cells are joined together by their tips and the cell surface available for contacts with other cells is considerably larger. In fact, the formation of auxospores on gametangia still joined in a chain has been reported for natural populations of P. cf. delicatissima and P. cf. calliantha (Sarno et al., Citation2010) and for P. pungens in culture (Chepurnov et al., Citation2005).
In F. kerguelensis, the detachment of chains was followed by cell pairing, which involved girdle-to girdle contact in a variable fashion and was mediated by thin mucilage threads. Pairing modality differs amongst Bacillariaceae, where pairing of gametangial cells generally occurs along the adjacent valves in species of the genus Pseudo-nitzschia (e.g. Davidovich & Bates, Citation1998; Chepurnov et al., Citation2005) and along the cingular bands in Nitzschia species (e.g. Mann, Citation1986; Kaczmarska et al., Citation2007; Trobajo et al., Citation2009). In F. kerguelensis, the paired gametangia showed the presence of a higher number of cingular bands in the hypovalve, which allowed their enlargement in the transapical direction. The synthesis of additional cingular bands was detected only in gametangia. Paired cells of F. kerguelensis showed an enlarged nucleus, which we interpret as being in early meiotic I prophase, by analogy with what has been observed in other pennate diatoms (e.g. Seminavis robusta, Chepurnov et al., Citation2002; Navicula oblonga, Mann & Stickle, Citation1989). In the gametangia of F. kerguelensis, the first meiotic division is followed by protoplast division, while the second meiotic division is acytokinetic, i.e. the nuclear division is not followed by cytokinesis. In this way, two binucleate gametes are formed by each of the paired gametangia. This pattern corresponds to ‘type I auxosporulation’, following the classification scheme provided by Geitler (Citation1973). This sequence of events is similar to what has been reported for some other diatoms, including the benthic raphid diatoms Seminavis robusta (Chepurnov et al., Citation2002) and Pinnularia cf. gibba (Poulíčková et al., Citation2007), but different patterns have been reported within raphid diatoms (reviewed by Chepurnov et al., Citation2004). As an example, only one uninucleate gamete is formed in each gametangium of the benthic freshwater genus Sellaphora (Mann, Citation1989), while two uninucleate gametes are produced in each gametangium of Pseudo-nitzschia pungens (Chepurnov et al., Citation2005). Unfortunately, we could not follow gamete conjugation in F. kerguelensis in real time. This species grows at low temperature and observation of live material in light microscopy can be carried out for only very limited periods to avoid cell death. Therefore we cannot confirm that conjugation occurs between the gametes of the two adjoined gametangia or if gametes are released in the medium and conjugate with gametes produced by different gametangia. Nevertheless, the binucleate gametes were almost always attached to the gametangial thecae and very few rounded protoplasts were observed free in the culture medium. This suggests that conjugation occurs between gametes of adjoined gametangia.
Auxospore formation and development
In samples incubated with PDMPO – a stain that binds to newly deposited silica – we observed four-nucleate zygotes surrounded by thin fluorescent ‘platelets’, which corresponded to the thin scales visible on the incunabula at the ends of the auxospores in SEM preparations. The presence of slightly silicified scales surrounding the auxospore is a common feature of centric diatoms (Kaczmarska et al., Citation2001), but thin, lightly silicified scales have been described also in some pennate diatoms (e.g. Idei et al., Citation2013). Within the Bacillariaceae, scales are present in Nitzschia longissima, where they occur on the surface of gametes of both mating types and, following auxospore elongation, remain at the tip of the caps and/or along the auxospore surface (Kaczmarska et al., Citation2007); and in Nitzschia inconspicua, where circular scales are present in the incunabula surrounding the zygotes and again persist in the caps at the ends of the expanding auxospores (Mann et al., Citation2013). Thin scales have been recorded also on the walls of passive gametes and on the auxospore caps of Pseudo-nitzschia multiseries (Kaczmarska et al., Citation2000), but have not been reported in other Pseudo-nitzschia species that have been investigated in SEM so far (e.g. Amato et al., Citation2005; Chepurnov et al., Citation2005; D'Alelio et al., Citation2009a). Information on the ultrastructure of the longitudinal perizonium and the modality and timing of synthesis is limited due to the fact that this thin structure is almost invisible in light microscopy. Incubation of F. kerguelensis auxospores with PDMPO showed that the longitudinal perizonium is deposited at the end of auxospore elongation. The fact that during the incubation period only a part of the perizonium became visible, suggests that this structure is composed by longitudinal bands that are deposited in sequence. The longitudinal perizonium can be observed in SEM preparations in which the outer, transverse perizonium has become broken. In Nitzschia inconspicua, it has been possible to determine that the longitudinal perizonium has an asymmetrical structure and is constituted by five bands comprising one wider primary band, two narrower secondary bands on one side of the primary band, and two extremely narrow ones on the other side (Mann et al., Citation2013).
Cardinal points in the life cycle of F. kerguelensis
The first evidence for sexual reproduction in F. kerguelensis came from a record of auxospores in phytoplankton samples collected in the Southern Ocean (Assmy et al., Citation2006). In the natural samples, auxospores of various sizes (i.e. in different stages of development) were recorded and they were often still connected to gametangial thecae 10–31 µm in length. The overall size in the natural population was 12–90 µm (P. Assmy unpublished data) and the fact that the gametangial thecae detected were only 10–31 µm long suggests that the cell size window for sexualization is close to that range. In the crosses carried out in the present study, most of the parental strains we tested had average lengths of ≤ 25 µm with one exception, which was strain L9C3 with an average length of 36 µm. This strain underwent sexual reproduction when crossed with compatible strains, showing that at least some F. kerguelensis cells can be sexualized up to 36 µm. Our current observations provide additional information on the size range of initial cells, which – in culture – were 78.4–100.8 µm in length; this is a slightly wider range than was recorded in natural samples by Assmy et al. (Citation2006), who reported 76–90 µm. The fact that initial cells are often attached to the theca of the gametangia in F. kerguelensis makes it easy to link the sizes of these two cardinal points or stages (sensu Geitler, Citation1932) in the life cycles; our data suggest that there is no clear relationship between their size in F. kerguelensis (see ). The few experimental studies addressing this question have provided contrasting results: a direct correlation between the size of parental strains and initial cells has been recorded in some species but not in others (Davidovich, Citation1994; Edlund & Bixby, Citation2001). Interestingly, different size ranges of gametangial and initial cells have been recorded between populations of the same species, suggesting that this trait might be used for their differentiation (Edlund & Bixby, Citation2001). Davidovich (Citation1994) investigated the possible effect of different irradiances and day-lengths on the size of initial cells but could not detect any significant relationship; he also showed that auxospore development was possible when crosses between opposite mating types were made in darkness. These results led him to conclude that the elongation capacity of the auxospore depends on increase in the volume of the vacuole, which is independent from photosynthesis and thus from the duration and intensity of the irradiance provided.
The length of a species’ sexual cycle is determined by the size of the initial cells, the rate of cell size reduction, and the size at which descendant cells become sexually inducible. In turn, the rate of cell size reduction depends on both the rate of cell division and the amount by which cells decrease in size at each division. In Pseudo-nitzschia delicatissima it has been shown that larger cells have the highest rates of cell size decrease per division, and this parameter progressively reduces as the average cell size of the strain diminishes (Amato et al., Citation2005). A similar trend has been detected in P. multistriata (D'Alelio et al., Citation2009a). Moreover, both of these Pseudo-nitzschia species show differences in their maximum growth rates during progression through their vegetative life phase, with larger cells showing lower growth rates. The data obtained for F. kerguelensis in the present study showed – at least for cells ≤ 50 µm in length – a direct relationship between cell size and the monthly reduction rates for strains grown under the same experimental conditions; this suggests either that smaller cells have lower division rates or that they possess a mechanism to limit cell size reduction at each division. We could estimate growth rates and thus cell size reduction per division only for seven strains ranging in size between 30 and 68 µm and no significant differences in cell size reduction per division were observed between the different strains. These results and the considerable variability detected in cell size reduction rates of larger strains suggest the presence of considerable intraspecific variability in growth rate, so that it will be necessary to examine a higher number of strains, covering the whole cell size range, in order to gain better insights into the mechanism and speed of cell size reduction in F. kerguelensis.
Fragilariopsis kerguelensis as a palaeo-proxy: a life cycle perspective
Valve morphometrics of Fragilariopsis kerguelensis have been suggested to have potential for reconstructing palaeo-oceanographic conditions in the Southern Ocean, based on the fact that the average frustule size varies amongst sites characterized by different productivity regimes (Cortese & Gersonde, Citation2007; Crosta, Citation2009; Cortese et al., Citation2012). Cortese & Gersonde (Citation2007) analysed the valve length and valve area of F. kerguelensis frustules in surface sediments from the Pacific and Atlantic sectors of the Southern Ocean (providing a spatial gradient) and one piston core (giving a temporal gradient) near the Antarctic Polar Front. Average valve size was larger in the proximity of the APF, characterized by high productivity, while it was smaller in the less productive zones to the north and south of it. In sediment cores, larger cells were found in layers from the glacial period compared with the interglacials. The authors related the presence of F. kerguelensis populations with larger average cell size to alleviation of iron limitation during iron-replete glacial conditions (Cortese & Gersonde, Citation2007). The possible link between glacial–interglacial changes in the average cell size of F. kerguelensis and iron availability (cells with a larger valvar area occurring in glacial periods characterized by a higher Fe deposition from atmospheric dust) was further confirmed by Cortese et al. (Citation2012). The authors also reported a larger valvar area for F. kerguelensis inside fertilized patches of two iron-fertilization experiments. Furthermore, analysis of Holocene core samples collected in the Eastern Antarctic Continental Shelf, an area not limited by iron, has shown that F. kerguelensis valves were longer and more abundant during the warmer Mid-Holocene period and shorter and less abundant during the colder Late-Holocene (Crosta, Citation2009). In this case, differences in the average population cell size were linked to changes in the length of the growth season, sea surface temperature and ice coverage.
During vegetative growth the average cell size of diatoms decreases while its variance increases (according to the MacDonald–Pfitzer rule; Chepurnov et al., Citation2004). Cell size decrease – in the absence of sexual reproduction – is proportional to the number of cell divisions that have occurred. The faster cells can divide, the faster they become smaller. Sexual reproduction, however, circumvents the progressive size diminution of the population by producing large initial cells. Thus, the average cell size of a diatom population is the integrated result of intrinsic life cycle traits and the response to proximate environmental factors. Iron availability, or the availability of other nutrients, such as silicate required for the synthesis of the frustules, might in fact influence valve morphometrics or the size of initial cells. Laboratory experiments have shown that Pseudo-nitzschia species can change their valve aspect ratio, increasing their transapical axis, when acclimated to iron-limited culture conditions (Marchetti & Harrison, Citation2007). If F. kerguelensis is found to have a similar response, it might help explain the differences in valve area recorded under iron-replete and -deplete conditions (Marchetti & Cassar, Citation2009). However, when comparing differences in cell size between natural populations, one should not only consider the average cell size/valvar area but the whole cell-size distribution. A higher average value can, in fact, be due to a relatively rapid response to specific environmental conditions that prompted the variation of the cell aspect ratio or to the presence of a higher proportion of larger cells produced following sexual reproduction. The plots of frequency distributions of valve length of F. kerguelensis reported by Crosta (Citation2009) show the presence of large cells in the same size range as the initial cells reported in the present study, thus supporting the idea that sexual reproduction may have been occurring in the natural populations whose frustules were subsequently deposited in the sediments. However, the only direct evidence of the occurrence of sexual reproduction in natural populations of F. kerguelensis was provided by Assmy et al. (Citation2006), who found auxospores, accounting for 0.03–0.4% of the total cell number during an iron-fertilization experiment. The highest percentages of auxospores were recorded in the fertilized patch, suggesting that iron-replete conditions might favour the onset of the sexual phase and the consequent production of large-sized cells. Furthermore, our data show considerable length variation amongst initial cells, suggesting genetic differences amongst strains, which opens the possibility of rapid environmental selection for bigger or smaller initial cells, and consequently for the average cell size of the whole population over time.
The possibility of manipulating the life cycle of F. kerguelensis in laboratory conditions, demonstrated here, opens the possibility of investigating the extent to which the various phases of the life cycle of this diatom are regulated by specific environmental conditions and assessing the plasticity and intraspecific diversity of different life cycle traits. This approach, coupled with a better understanding of the level at which sediment records reflect water column events, and the analysis of cell size distributions with appropriate measurement strategies, will help in assessing the possible use of F. kerguelensis parameters as a proxy for palaeoecological conditions and improve our understanding of the mechanisms through which life history traits have evolved in planktonic diatoms and contributed to their ecological success.
Supplementary information
The following supplementary material is available for this article, accessible via the Supplementary Content tab on the article’s online page at http://dx.doi.org/10.1080/09670262.2013.849360
Supplementary file 1. ITS alignment, in FASTA format. Position 1 constitutes the 5′-end of ITS-1 and position 776 the 3′-end of ITS-2; ITS-1 spans positions 1–309; 5.8S rDNA, 310-481; and ITS-2, 482–776. Note that all ITS sequences, except the one of strain 4-20, are incomplete at the 5′-end and that the ITS sequences of strains MM-E13C5 and MM-E13D2 are incomplete at the 3′-end.
Supplementary Table 1. Fragilariopsis kerguelensis strains used in this study, including GenBank accessions.
Supplementary Table 2. Nucleotide differences between the ITS sequences of F. kerguelensis strain 4-20 (sequence EF660061) and those of the strains used in this study.
Supplementary Figure 1. Map of the Atlantic Sector of the Southern Ocean with the sampling stations at which Fragilariopsis kerguelensis strains were collected.
Supplementary Figure 2. Size spectra of seven clonal strains of Fragilariopsis kerguelensis measured at time 0 and after 7 months.
Supplementary material
Download Zip (2.3 MB)Acknowledgements
Part of this work was funded by the German Science Foundation (grant no. DE 1455/2‐1) granted to N.F. and part from the PhD thesis of E.S. supported by Stazione Zoologica Anton Dohrn (SZN). The research of P.A. at SZN was supported by the European Community – Research Infrastructure Action under the FP7 ‘Capacities’ Specific Programme ASSEMBLE (grant no. 227799). The authors wish to thank Carmen Minucci for strain maintenance at SZN.
Related Research Data
References
- Amato, A., Orsini, L., D'Alelio, D. & Montresor, M. (2005). Life cycle, size reduction patterns, and ultrastructure of the pennate planktonic diatom Pseudo-nitzschia delicatissima (Bacillariophyceae). Journal of Phycology, 41: 542–556.
- Amato, A., Kooistra, W.H.C.F., Levialdi Ghiron, J.H., Mann, D.G., Pröschold, T. & Montresor, M. (2007). Reproductive isolation among sympatric cryptic species in marine diatoms. Protist, 158: 193–207.
- Assmy, P., Henjes, J., Smetacek, V. & Montresor, M. (2006). Auxospore formation in the silica-sinking oceanic diatom Fragilariopsis kerguelensis (Bacillariophyceae). Journal of Phycology, 42: 1002–1006.
- Assmy, P., Henjes, J., Klaas, C. & Smetacek, V. (2007). Mechanisms determining species dominance in a phytoplankton bloom induced by the iron fertilization experiment EisenEx in the Southern Ocean. Deep Sea Research Part I: Oceanographic Research Papers, 54: 340–362.
- Brzezinski, M.A. (1985). The Si: C: N ratio of marine diatoms: interespecific variability and the effect of some environmental variables. Journal of Phycology, 21: 347–357.
- Cefarelli, A.O., Ferrario, M.E., Almandoz, G.O., Atencio, A.G., Akselman, R. & Vernet, M. (2010). Diversity of the diatom genus Fragilariopsis in the Argentine Sea and Antarctic waters: morphology, distribution and abundance. Polar Biology, 33: 1463–1484.
- Chepurnov, V.A., Mann, D.G., Vyverman, W., Sabbe, K. & Danielidis, D.B. (2002). Sexual reproduction, mating system, and protoplast dynamics of Seminavis (Bacillariophyceae). Journal of Phycology, 38: 1004–1019.
- Chepurnov, V.A., Mann, D.G., Sabbe, K. & Vyverman, W. (2004). Experimental studies on sexual reproduction in diatoms. International Review of Cytology, 237: 91–154.
- Chepurnov, V.A., Mann, D.G., Sabbe, K., Vannerum, K., Casteleyn, G., Verleyen, E., Peperzak, L. & Vyverman, W. (2005). Sexual reproduction, mating system, chloroplast dynamics and abrupt cell size reduction in Pseudo-nitzschia pungens from the North Sea (Bacillariophyta). European Journal of Phycology, 40: 379–395.
- Cortese, G. & Gersonde, R. (2007). Morphometric variability in the diatom Fragilariopsis kerguelensis: implications for Southern Ocean paleoceanography. Earth and Planetary Science Letters, 257: 526–544.
- Cortese, G., Gersonde, R., Maschner, K. & Medley, P. (2012). Glacial-interglacial size variability in the diatom Fragilariopsis kerguelensis: possible iron/dust controls? Paleoceanography, 27: 1–14.
- Crawford, R.M. (1995). The role of sex in the sedimentation of a marine diatom bloom. Limnology and Oceanography, 40: 200–204.
- Crosta, X. (2009). Holocene size variations in two diatom species off East Antarctica: productivity vs environmental conditions. Deep-Sea Research Part I Oceanographic Research Papers, 56: 1983–1993.
- D'Alelio, D., Amato, A., Luedeking, A. & Montresor, M. (2009a). Sexual and vegetative phases in the planktonic diatom Pseudo-nitzschia multistriata. Harmful Algae, 8: 225–232.
- D’Alelio, D., Amato, A., Kooistra, W.H.C.F., Procaccini, G., Casotti, R. & Montresor, M. (2009b). Internal Transcribed Spacer polymorphism in Pseudo-nitzschia multistriata (Bacillariophyceae) in the Gulf of Naples: recent divergence or intraspecific hybridization? Protist, 160: 9–20.
- D’Alelio, D., Ribera D'Alcalà, M., Dubroca, L., Sarno, D., Zingone, A. & Montresor, M. (2010). The time for sex: a biennial life cycle in a marine planktonic diatom. Limnology and Oceanography, 55: 106–114.
- Davidovich, N.A. (1994). Factors controlling the size of initial cells in diatoms. Russian Journal of Plant Physiology, 41: 220–224.
- Davidovich, N.A. & Bates, S.S. (1998). Sexual reproduction in the pennate diatoms Pseudo-nitzschia multiseries and P. pseudodelicatissima (Bacillariophyceae). Journal of Phycology, 34: 126–137.
- Edlund, M.B. & Bixby, R.J. (2001). Intra- and inter-specific differences in gametangial and initial cell size in diatoms. In Proceedings of the 16th International Diatom Symposium 2000 (Economou-Amilli, A., editor), 169–190. University of Athens, Athens.
- Evans, K.M., Wortley, A.H., Simpson, G.E., Chepurnov, V.A. & Mann, D.G. (2008). A molecular systematic approach to explore diversity within the Sellaphora pupula species complex (Bacillariophyta). Journal of Phycology, 44: 215–231.
- Gall, M.P., Boyd, P.W., Hall, J., Safi, K.A. & Chang, H. (2001). Phytoplankton processes. Part 1: Community structure during the Southern Ocean Iron RElease Experiment (SOIREE). Deep Sea Research Part II: Topical Studies in Oceanography, 48: 2551–2570.
- Geitler, L. (1932). Der Formwechsel der pennaten Diatomeen (Kieselalgen). Archiv für Protistenkude, 78: 1–226.
- Geitler., L. (1973). Auxosporenbidung und Systematik bei pennaten Diatomeen und die Zytologie von Cocconeis-Sippen. Österreichische Botanische Zeitschrift, 122: 299–321.
- Guillard, R.L. & Ryther, J.H. (1962). Studies of marine planktonic diatoms. I. Cyclotella nana Hustedt, and Detonula confervacea (Cleve) Gran. Canadian Journal of Microbiology, 8: 229–239.
- Hamm, C.E., Merkel, R., Springer, O., Jurkojc, P., Maier, C., Prechtel, K. & Smetacek, V. (2003). Architecture and material properties of diatom shell provide effective mechanical protection. Nature, 421: 841–843.
- Hamsher, S.E., Evans, K.M., Mann, D.G., Poulícková, A. & Saunders, G.W. (2011). Barcoding diatoms: exploring alternatives to COI-5P. Protist, 162: 405–422.
- Hart, T.J. (1942). Phytoplankton periodicity in Antarctic surface waters. Discovery Reports, 21: 263–348.
- Hasle., G.R. (1965). Nitzschia and Fragilariopsis species studied in the light and electron microscopes. III. The genus Fragilariopsis. Skrifter utgitt av det Norske Videnskaps-Akademi I Oslo: I Matematisk-Naturvideskapeling Klasse, 21: 1–49.
- Hasle, G.R. (1968). Observations on the marine diatom Fragilariopsis kerguelensis (O’Meara) Hust. in the scanning electron microscope. Nytt Magasin for Botanikk, 15: 205–208.
- Hasle, G.R. & Syvertsen, E.E. (1997). Marine diatoms. In Identifying marine phytoplankton (Tomas, C.R., editor), 5–385. Academic Press, San Diego, CA.
- Hoffmann, L.J., Peeken, I. & Lochte, K. (2007). Effects of iron on the elemental stoichiometry during EIFEX and in the diatoms Fragilariopsis kerguelensis and Chaetoceros dichaeta. Biogeosciences, 4: 569–579.
- Holtermann, K.E., Bates, S.S., Trainer, V.L., Odell, A. & Armbrust, E.V. (2010). Mass sexual reproduction in the toxigenic diatoms Pseudo-nitzschia australis and P. pungens (Bacillariophyceae) on the Washington coast. Journal of Phycology, 46: 41–52.
- Hustedt, F. (1958). Diatomeen aus der Antarktis und dem Südatlantik. Deutsche Antarktische Expedition 1938/39, 2: 103–191.
- Idei, M., Sato, S., Watanabe, T., Nagumo, T. & Mann, D.G. (2013). Sexual reproduction and auxospore structure in Diploneis papula (Bacillariophyta). Phycologia, 52: 295–308.
- Jewson., D.H. (1992). Size reduction, reproductive strategy and the life strategy of a centric diatom. Philosophical Transactions of the Royal Society of London, Series B, 336: 191–213.
- Kaczmarska, I., Bates, S.S., Ehrman, J.M. & Léger, C. (2000). Fine structure of the gamete, auxospore and initial cell in the pennate diatom Pseudo-nitzschia multiseries (Bacillariophyta). Nova Hedwigia, 71: 337–357.
- Kaczmarska, I., Ehrman, J.M. & Bates, S.S. (2001). A review of auxospore structure, ontogeny and diatom phylogeny. In Proceedings of the 16th International Diatom Symposium 2000 (Economou-Amilli, A., editor), 153–167. Amvrosius Press, Athens.
- Kaczmarska, I., Davidovich, N.A. & Ehrman, J.M. (2007). Sex cells and reproduction in the diatom Nitzschia longissima (Bacillariophyta): discovery of siliceous scales in gamete cell walls and novel elements of the perizonium. Phycologia, 46: 726–737.
- Kaczmarska, I., Poulíčková, A., Sato, S., Edlund, M.B., Idei, M., Watanabe, T. & Mann, D.G. (2013). Proposals for a terminology for diatom sexual reproduction, auxospores and resting stages. Diatom Research, 28: 263–294.
- Kooistra, W.H.C.F., De Stefano, M., Medlin, L.K. & Mann, D.G. (2003). The phylogenetic position of Toxarium, a pennate-like lineage within centric diatoms (Bacillariophyceae). Journal of Phycology, 39: 185–197.
- Leblanc, K. & Hutchins, D.A. (2005). New applications of a biogenic silica deposition fluorophore in the study of oceanic diatoms. Limnology and Oceanography – Methods, 3: 462–476.
- Levialdi Ghiron, J.H., Amato, A., Montresor, M. & Kooistra, W.C.H.F. (2008). Plastid inheritance in the planktonic raphid pennate diatom Pseudo-nitzschia delicatissima (Bacillariophyceae). Protist, 159: 91–98.
- Lundholm, N. & Hasle, G.R. (2010). Fragilariopsis (Bacillariophyceae) of the Northern Hemisphere – morphology, taxonomy, phylogeny and distribution, with a description of F. pacifica sp. nov. Phycologia, 49: 438–460.
- Lundholm, N., Daugbjerg, N. & Moestrup, Ø. (2002). Phylogeny of the Bacillariaceae with emphasis on the genus Pseudo-nitzschia (Bacillariophyceae) based on partial LSU rDNA. European Journal of Phycology, 37: 115–134.
- Mann, D.G. (1986). Methods of sexual reproduction in Nitzschia: systematic and evolutionary implications. Diatom Research, 1: 193–203.
- Mann, D.G. (1988). Why didn't Lund see sex in Asterionella? A discussion of the diatom life cycle in nature. In Algae and the aquatic environment (Round, F.E., editor), 385–412. Biopress, Bristol.
- Mann, D.G. (1989). The diatom genus Sellaphora: separation from Navicula. British Phycological Journal, 24: 1–20.
- Mann, D.G. & Stickle, A.J. (1989). Meiosis, nuclear cyclosis, and auxospore formation in Navicula sensu stricto (Bacillariophyceae). British Phycological Journal, 24: 167–181.
- Mann, D.G., Sato, S., Rovira, L. & Trobajo, R. (2013). Paedogamy and auxosporulation in Nitzschia sect. Lanceolatae (Bacillariophyta). Phycologia, 52: 204–220.
- Marchetti, A. & Cassar, N. (2009). Diatom elemental and morphological changes in response to iron limitation: a brief review with potential paleoceanographic applications. Geobiology, 7: 419–431.
- Marchetti, A. & Harrison, P.J. (2007). Coupled changes in the cell morphology and elemental (C, N, and Si) composition of the pennate diatom Pseudo-nitzschia due to iron deficiency. Limnology and Oceanography, 52: 2270–2284.
- Moniz, M.B.J. & Kaczmarska, I. (2010). Barcoding of diatoms: nuclear encoded ITS revisited. Protist, 161: 7–34.
- Poulíčková, A., Mayama, S., Chepurnov, V.A. & Mann, D.G. (2007). Heterothallic auxosporulation, incunabula and perizonium in Pinnularia (Bacillariophyceae). European Journal of Phycology, 42: 367–390.
- Quijano-Scheggia, S.I., Garcés, E., Lundholm, N., Moestrup, O., Andree, K. & Camp, J. (2009). Morphology, physiology, molecular phylogeny and sexual compatibility of the cryptic Pseudo-nitzschia delicatissima complex (Bacillariophyta), including the description of P. arenysensis sp. nov. Phycologia, 48: 492–509..
- Rambaut, A. (2002). Sequence Alignment Editor v2.0a11. Available at http://tree.bio.ed.ac.uk/software/seal/
- Round, F.E., Crawford, R.M. & Mann, D.G. (1990). The diatoms. Biology and morphology of the genera. Cambridge University Press, Cambridge.
- Sarno, D., Kooistra, W.C.H.F., Balzano, S., Hargraves, P.E. & Zingone, A. (2007). Diversity in the genus Skeletonema (Bacillariophyceae): III. Phylogenetic position and morphological variability of Skeletonema costatum and Skeletonema grevillei, with the description of Skeletonema ardens sp. nov. Journal of Phycology, 43: 156–170.
- Sarno, D., Zingone, A. & Montresor, M. (2010). A massive and simultaneous sex event of two Pseudo-nitzschia species. Deep Sea Research Part II: Topical Studies in Oceanography, 57: 248–255.
- Smetacek, V., Assmy, P. & Henjes, J. (2004). The role of grazing in structuring Southern Ocean pelagic ecosystems and biogeochemical cycles. Antarctic Science, 16: 541–558.
- Treguer, P., Nelson, D.M., Bennekom, A.J.V., Demaster, D.J., Leynaert, A. & Queguiner, B. (1995). The silica balance in the world ocean: a reestimate. Science, 268: 375–379.
- Trobajo, R., Clavero, E., Chepurnov, V.A., Sabbe, K., Mann, D.G., Ishihara, S. & Cox, E.J. (2009). Morphological, genetic and mating diversity within the widespread bioindicator Nitzschia palea (Bacillariophyceae). Phycologia, 48: 443–459.
- Trobajo, R., Rovira, L., Ector, L., Wetzel, C.E., Kelly, M. & Mann, D.G. (2013). Morphology and identity of some ecologically important small Nitzschia species. Diatom Research, 28: 37–59.
- Zielinski, U. & Gersonde, R. (1997). Diatom distribution in Southern Ocean surface sediments (Atlantic sector): implications for paleoenvironmental reconstructions. Palaeogeography, Palaeoclimatology, Palaeoecology, 129: 213–250.