
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
Our objective was to evaluate the interaction of supplemental essential oils (EO) and monensin (MON) on performance and digestion. Two levels of EO (0 vs 0.011% Xtract®) and two levels of MON (0 vs 33 mg/kg) were evaluated in a 2 × 2 factorial arrangement. Treatment effects on 216-d growth performance were evaluated in 96 Holstein steers. Supplemental MON enhanced (P ≤ 0.05) ADG, DMI and gain efficiency. Without EO, MON improved (P < 0.05) gain efficiency and dietary NE. However, in diets with both EO and MON, gain efficiency and dietary NE were not different from non-supplemented diets (interaction, P < 0.10). Treatment effects on characteristics of digestion were evaluated using four cannulated steers. Ruminal digestion of OM and N were greater for MON alone. Ruminal digestion of OM and N were similar for EO-alone vs MON-alone, but not by the combination of the two (interaction P ≤ 0.02). Supplemental MON enhanced (P ≤ 0.02) total tract digestion of OM and N. We conclude that supplemental EO-alone or MON-alone may enhance gain efficiency and NE value of the diet. However, effects are negated when fed in combination. Treatment effects on performance are consistent with digestion measures.
1. Introduction
Ionophores are antibiotics extensively included in diets for feedlot cattle in the USA, largely due to their beneficial effects on gain efficiency (Goodrich et al. Citation1984). This effect is partially attributable to selective action on the gram positive bacteria, potentially decreasing ruminal acetate:propionate molar ratio and methane energy losses. Additionally, ionophores may enhance metabolizable protein (Russell and Strobel Citation1988; Zinn et al. Citation1994), and reduce maintenance energy requirements (Zinn Citation1987). Monensin is among the ionophores most commonly used in the growing-finishing diets for feedlot cattle (Samuelson et al. Citation2016). The present ban on ionophore supplementation within the European Union (European Commission Citation2003; Directive 1831/2003/CEE), and the potential trend elsewhere, has led the quest for ‘organic’ alternatives to conventional antibiotic supplements. Among those considered are the plant extracted essential oils. As with the ionophores, essential oil supplementation may likewise reduce acetate:propionate molar ratio, methane production and ruminal protein degradation (Cobellis et al. Citation2016). However, the broader, often non-specific antimicrobial activities within the rumen (Calsamiglia et al. Citation2007; Benchaar et al. Citation2008; Patra Citation2011; Khiaosa-ard and Zebeli Citation2013; Cobellis et al. Citation2016) may also lead to reduced ruminal digestion and growth performance. Studies evaluating effects of essential oils supplementation on growth performance of feedlot cattle are limited and inconclusive. Araujo et al. (Citation2019) observed similar growth performance of feedlot cattle when supplemented with essential oils alone or monensin alone. Enhancements were additive when the two were fed in combination. In contrast, Geraci et al. (Citation2012) observed greater ADG in steers supplemented with essential oils than for steers supplemented with monensin. Souza et al. (Citation2019) observed greater ADG and DMI of feedlot heifers supplemented with an essential oil blend than heifers receiving rosemary essential oil alone.
2. Material and methods
All procedures involving animal care and management were in accordance with and approved by the University of California, Davis, Animal Use and Care Committee (Protocol # 20548, 20615).
2.1. Experiment 1
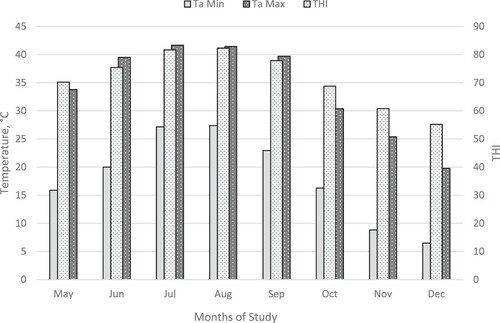
Weather measurements were obtained every 30 min from an on-site weather station throughout the experimental period. Temperature humidity index (THI) was calculated using the following formula: THI = 0.81 × T + RH (T−14.40) + 46.40 (Mader et al. Citation2006). The THI during this experiment is shown in . In accordance with nominal coding (Normal THI < 74; Alert 75 < THI < 78; Danger 79 < THI < 83; and Emergency THI > 84; Mader et al. Citation2006), cattle experienced ‘alert’ or greater ambient conditions during months of June through September.
Figure 1. Minimum and maximum daily temperature and temperature humidity index (THI) during the study.
Ninety-six Holstein steer calves were used to evaluate treatment effects on growth performance and dietary net energy. Treatments consisted of steam-flaked corn-based growing-finishing diets supplemented (DM basis) with two levels of a blend of phytomolecules (0 vs 0.011% Xtract® Ruminant 7065, coated extract blend containing 17% eugenol, 11% cinnamaldehyde, and 7% capsaicin oleoresin.; Pancosma, Geneva, Switzerland) and two levels of monensin (0 vs 30 mg/kg (Rumensin-90), Elanco, Greenfield, IN, USA) in 2 × 2 factorial arrangement. Composition of experimental diets are shown in . Based on final 196-d DMI of calf-fed Holstein steers from a prior study (Ortiz et al. Citation2016) fed diets similar to that of the present study, it was expected that DMI of steers during the course of the study would average approximately 9.1 kg/d. Accordingly, the level of inclusion of Xtract® Ruminant 7065 was determined to provide an average of 1 g/hd/d. However, actual DMI average 8.4 kg/d. Consequently, average intake of Xtract® Ruminant 7065 averaged 0.93 g/hd/d.
Table 1. Experimental diets, Trials 1 and 2.Table Footnotea
Steers used in the study originated from Tulare, California, arriving at the University of California Desert Research Center, Holtville, on January 23, 2018, with an arrival weight of 127 ± 9 kg. The trial initiated on May 15, 2018, following a 112-d receiving period. Upon initiation of the study, steers were individually weighed, injected with 500,000 IU vitamin A (Vital EA-D, Stuart Products, Bedford, TX) and implanted with Revalor-S (Intervet, Millsboro, DE). Steers were sorted by weight into four weight groupings and then randomly assigned within groupings to 16 pens (6 steers/pen). Diets were prepared at weekly intervals and stored in plywood boxes located in front of each pen. Steers were allowed ad libitum access to water and dietary treatments. Fresh feed was provided twice daily. Steers were reimplanted with Revalor-S on September 4, 2018 (d 112 of the study).
Energy gain (EG, Mcal/d) was calculated by the equation (NRC Citation1984): EG = (ADG1.097) (0.0557W.75). Maintenance energy (EM) was calculated by the equation: EM = (0.086) (W.75) (Fox and Black Citation1984; NRC Citation1988). From the derived estimates of energy required for maintenance and gain, the NEm and NEg values of the diet were obtained by means of the quadratic formula:
where a = (−0.41) (EM); b = [(0.877) (EM)] + [(0.41) (DMI)] + EG; and c = [(−0.877) (DMI)], and NEg = [(0.877) (NEm)] – 0.41 (Zinn and Shen Citation1998). For calculating steer performance, live weights were reduced 4% to account for digestive tract fill. The trial was analysed as a randomized complete block design experiment with a 2 × 2 factorial arrangement of treatments, considering pen as experimental unit. The following statistical model was used:
where μ is the general mean, αi is the effect of the factor A, βj is the effect of the factor B, (αβ)ij is the effect of the interaction of the factor A with the factor B, and ϵijk represents the residual error. Treatment main effects and interactions were tested by means of orthogonal contrasts. Significance was declared at P ≤ 0.05; a tendency was considered at P > 0.05 and P ≤ 0.10. (Statistix 10, Analytical Software, Tallahassee, FL).
2.2. Experiment 2
Four Holstein steers (189 ± 13 kg) with cannulas in the rumen and proximal duodenum were used in a 4 × 4 Latin square experiment. Treatments were the same as in Trial 1. A single basal diet was prepared, with the incorporation of 0.3% chromic oxide as a digesta marker. Corresponding amounts of Xtract (120 mg/kg diet DM) and monensin (33.5 mg/kg diet DM) were top-dress on the basal diet at time of feeding. To avoid refusals, DMI was restricted to 2.2% of live weight. Diets were fed at 0800 and 2000 daily. Experimental periods consisted of a 17-d diet adjustment period followed by a 4-d collection period. During the collection period duodenal and fecal samples were taken from all steers, twice daily as follows: d 1, 0750 and 1350; d 2, 0900 and 1500; d 3, 1050 and 1650; and d 4, 1200 and 1800. Individual samples consisted of approximately 700 ml duodenal chime, 200 g (wet basis) fecal material, and 100 mL ruminal fluid (obtained from each steer via the ruminal cannula).
Ruminal fluid pH was determined on freshly collected samples strained through 4 layers of cheesecloth. Freshly prepared 25% (wt/vol) m-phosphoric acid (2 mL) was added to 8 mL of the strained ruminal fluid. Samples were centrifuged (17,000 × g for 10 min) and supernatant fluid stored at −20°C for analysis of VFA concentrations (gas chromatography; Zinn Citation1988). Duodenal and fecal samples from each steer within each collection period were composited (equal weight basis) for analysis. Upon completion of the trial, ruminal fluid was obtained from all steers and composited for isolation of ruminal bacteria via differential centrifugation (Bergen et al. Citation1968).
Feed, duodenal fluid and fecal samples were subjected to the following analysis: DM (oven drying at 105 °C until no further weight loss; method 930.15; AOAC Citation2000); ash (method 942.05; AOAC Citation2000), Kjeldahl N (method 984.13; AOAC Citation2000); aNDFom [Van Soest et al. (Citation1991), corrected for NDF-ash, incorporating heat stable α-amylase (Ankom FAA, Ankom Technology, Macedon, NY) at 1 mL per 100 mL of NDF solution]; chromic oxide (Hill and Anderson Citation1958); and starch (Zinn Citation1990). Duodenal samples were analysed for ammonia N (method 941.04; AOAC Citation2000) and purines (Zinn and Owens Citation1986). Duodenal flow and fecal excretion of DM were calculated based on marker ratio, using chromic oxide. Microbial organic matter (MOM) and microbial N (MN) leaving the abomasum was calculated using purines as a microbial marker (Zinn and Owens Citation1986). Organic matter (OM) fermented in the rumen is considered equal to OM intake minus the difference between the amount of total OM reaching the duodenum and MOM reaching the duodenum. Feed N escape to the small intestine is considered equal to total N leaving the abomasum minus ammonia-N, MN and endogenous N (0.195 × BW0.75; Ørskov et al. Citation1986). Methane production was estimated based on the theoretical fermentation balance for observed molar distribution of VFA and OM fermented in the rumen (Wolin Citation1960).
The trial was analysed as a balanced 4 × 4 Latin square with a 2 × 2 factorial arrangement of treatments according to the following statistical model:
where Yijk is the response variable, µ is the common experimental effect, Ai is the steer effect, Pj is the period effect, Tk is the treatment effect and Eijk is the residual error. Treatment main effects and interaction were tested by means of orthogonal contrasts. Treatment effects on ruminal pH and VFA were analysed using linear mixed model for analysis of repeated measures as follows:
where Yij(k)l is the treatment variable, µ is the common experimental effect, Ai is the random steer effect, Pj is the fixed period effect; Tk is the fixed treatment effect, Hl is the fixed hour effect; (TH)kl is the fixed interaction effect of treatment with hour; Eij(k) is the random effect of interaction of steer with period within treatment; and Eijl(k) is the residual error (SAS Inst. Inc., Cary, NC; Version 9.1). Covariance structures for pH and VFA (selected for minimum AIC, AICC and BIC), were heterogeneous Toeplitz and heterogeneous AR(1), respectively.
3. Results
3.1. Experiment 1
Treatment effects on cattle growth-performance are shown in . Supplemental monensin enhanced ADG during the initial 112-d period (main effect, P = 0.05) and across the entire 216 d of the trial (main effect, P = 0.04) by 6.3 and 4.3%, respectively. During the final 104-d period there was a tendency (P = 0.07) for a treatment interaction. In diets without EO monensin improved ADG; however in diets with EO, supplemental monensin depressed ADG.
Table 2. Treatment effects on growth performance.
Consistent with differences in ADG, supplemental monensin enhanced DMI during the initial 112-d period (main effect, P = 0.01) and across the entire 216-d trial (main effect, P = 0.03). During the final 104-d period there was a treatment interaction on DMI (P = 0.05).
There were no treatment effects (P > 0.10) on gain efficiency during the initial 112-d and final 104-d period. Although, across the entire 216-d trial there was a tendency for treatment interactions on both gain efficiency (P = 0.08) and estimated dietary NE (P = 0.09). In diets without EO, monensin improved gain efficiency and dietary NE. However, in diets with EO supplemental monensin depressed gain efficiency and estimated dietary NE.
3.2. Experiment 2
Treatment effects on characteristics of ruminal and total tract digestion are shown in . There was a tendency (P = 0.09) for treatment interaction on estimated flow of microbial N to the small intestine. In diets without EO, monensin increased estimated flow of microbial N to the small intestine. However, in diets with EO, monensin supplementation reduced microbial N flow to the small intestine. This increase in net synthesis was consistent with greater ruminal digestion of OM (interaction, P = 0.01) for monensin alone, and the effect of EO-alone was not different than MON-alone. However, ruminal OM was not enhanced by the combination of the two. Consistent with increased ruminal OM digestion, ruminal digestion of dietary N digestion was greater (interaction, P = 0.02) for MON-alone, or EO-alone versus the non-supplemented control diet. But feed N digestion was not improved with the combination of MON with EO.
Table 3. Treatment effects on characteristics of digestion.
There was a tendency (interaction, P = 0.06) for treatment interaction on ruminal NDF digestion. In diets without EO, monensin improved ruminal NDF digestion. Whereas, in diets with EO, supplemental monensin depressed ruminal NDF digestion. Total tract NDF digestion was greater (interaction, P = 0.05) for MON-alone than for the combination of MON with EO.
Both supplemental monensin and EO enhanced total tract OM digestion. However, the effect was not additive (interaction, P = 0.06). Supplemental monensin enhanced (main effect, P ≤ 0.01) post ruminal and total tract apparent N digestion (5.4 and 6.5%, respectively).
Treatment effects on ruminal fermentation characteristics are shown in . Treatment effects on ruminal pH at different hours of the day are shown in . There was a treatment by time interaction (P = 0.014). Ruminal pH was lower for monensin alone, or EO alone, than for the combination of the two at 6:00 h and 16:00 h. At 10:30 h ruminal pH was greater for non-supplemented steers than for steers supplemented with monensin, EO, or the combination of monensin and EO.
Figure 2. Treatment effects on pH during a 12-h feeding interval. Designations: -MON = no monensin; +MON = with monensin; -EO = no essential oils; +EO = with essential oils. There were treatment by time interactions (P < 0.05). At 06:00 h and 16:00 h ruminal pH was lower for monensin alone, or EO alone, than for the combination. At 10:30 h ruminal pH was greater for non-supplemented steers than for steers supplemented with monensin, EO, or the combination of monensin and EO.
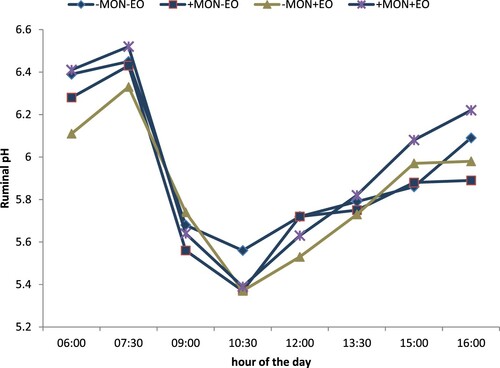
Table 4. Treatment effects on ruminal pH, VFA, and estimated methane for 12-h feeding interval.
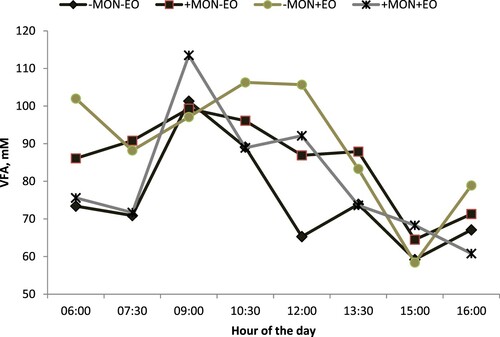
Treatment effects on ruminal VFA concentration at different hours of the day are shown in . There were treatment by time interactions (P < 0.05). At 06:00 h and 16:00 h ruminal pH was lower for monensin alone, or EO alone, than for the combination. At 10:30 h ruminal pH was greater for non-supplemented steers than for steers supplemented with monensin, EO, or the combination of monensin and EO.
Figure 3. Treatment effects on total VFA during a 12-h feeding interval. Designations: -MON = no monensin; +MON = with monensin; -EO = no essential oils; +EO = with essential oils. There was interaction (MONxEO) effect at 06:00 h (P < 0.01); 7:30 h (P = 0.03) and 10:30 h (P < 0.01). The interaction of treatment with time was not appreciable (P = 0.32).
There were no treatment effects (P > 0.10) on ruminal acetate and propionate molar proportions or estimated methane production (mole methane/mole glucose equivalent fermented). Molar proportions of butyrate and isovalerate tended to be greater (interaction, P < 0.08) for monensin alone, or EO alone, than for the combination of the two.
4. Discussion
4.1. Experiment 1
The influence of monensin supplementation on growth performance of feedlot cattle has been inconsistent. In the present study, monensin improved ADG during the first 112- d period and across the entire 216-d study. Barreras et al. (Citation2013) observed improved gain efficiency and dietary NE in beef heifers fed a steam-flaked corn-based finishing diet supplemented with monensin. Montano et al. (Citation2015) observed improved ADG, gain efficiency and estimated dietary NE in steers fed a steam-flaked corn-based finishing diet.
The increase in DMI with monensin supplementation contrasts with the generalized expectation that the ionophore decreases DMI of feedlot cattle (Duffield et al. Citation2012). However, in those instances where monensin supplementation increased ADG, as in the present study, DMI was not depressed (Felix and Loerch Citation2011; Barreras et al. Citation2013; Thompson et al. Citation2016). In the present study, increase in DMI and associated increase in ADG with monensin supplementation may be at least partially attributable to ambient thermal conditions. Cattle were exposed to caloric stress for approximately 100 d of the 216-d study. Barreras et al. (Citation2013) observed reduced DMI of feedlot cattle supplemented with monensin when cattle were not exposed to caloric stress. However, when exposed to caloric stress (THI values greater than 79), supplemental monensin did not reduce DMI.
In the present study, monensin supplementation tended to improve gain efficiency and estimated dietary NE. Although, in numerous other studies with feedlot cattle fed steam flaked corn-based diets, monensin supplementation did not enhance ADG or gain efficiency (Zinn and Borques Citation1993; Zinn et al. Citation1994; Depenbusch et al. Citation2008; Salinas-Chavira et al. Citation2009). Again, these differences in response to supplemental monensin may be associated with ambient conditions. In the present study cattle were exposed to caloric stress during initial 112-d period of the study (), when the monensin effect was most evident.
Consistent with the present study, differences in gain efficiency between supplemental monensin and EO have not been appreciable (Benchaar et al. Citation2006; Devant et al. Citation2007; Meyer et al. Citation2009; Yang et al. Citation2010). However, Ornaghi et al. (Citation2017) observed increased ADG and DMI of finishing bulls supplemented with clove or cinnamon essential oils. Souza et al. (Citation2019) observed greater ADG, DMI and gain efficiency of feedlot heifers supplemented with a blend of eugenol, thymol, vanillin, rosemary and clove essential oils vs non-supplemented heifers. Likewise, An et al. (Citation2020) observed that EO supplementations (same blend as used in the present study) enhanced growth performance, nutrient digestion, immune response, antioxidant status of feedlot lambs.
The apparent negative interactions of the combination of essential oils and monensin on gain efficiency and estimated dietary NE in present study are in agreement with digestion and ruminal fermentation characteristics. This may be indicative of similarities between MON and EO in their mode of action (as likewise, growth performance was not enhanced with the combination of the two additives). In contrast, Araujo et al. (Citation2019) observed that whereas gain efficiency was similar for feedlot steers receiving monensin alone and essential oils alone (different blend than that used in the present study), the combination of the two had an additive effect. Authors did not report digestion or rumen fermentation characteristics.
4.2. Experiment 2
The greater ruminal OM digestion for monensin alone (interaction effect) contrasts with Zinn (Citation1987) who did not observe an influence of supplemental monensin on ruminal OM digestion. In several instances supplemental monensin decreased ruminal OM digestion (Zinn and Borques Citation1993; Zinn et al. Citation1994; Surber and Bowman Citation1998; Salinas-Chavira et al. Citation2009; Montano et al. Citation2015) and feed nitrogen degradation (Poos et al. Citation1979; Zinn and Borques Citation1993; Zinn et al. Citation1994; Surber and Bowman Citation1998; Montano et al. Citation2015). Likewise, the positive effect of monensin on post ruminal and total tract apparent N digestion (treatment main effect) of cattle receiving finishing diets is not consistent with earlier studies (Zinn et al. Citation1994; Salinas-Chavira et al. Citation2009; Montano et al. Citation2015).
In the present study, monensin alone tended to increase microbial N flow to the small intestine. In prior studies the effect of supplemental monensin on flow of microbial N to the small intestine was non-appreciable (Zinn and Borques Citation1993; Surber and Bowman Citation1998) or even negative (Poos et al. Citation1979; Zinn et al. Citation1994; Montano et al. Citation2015). These contrasting effects are related to differing effects of supplemental monensin on ruminal OM digestion. In the present study, monensin supplementation did not affect ruminal microbial efficiency (g microbial N/kg OM fermented). The increase in passage of microbial N to the small intestine is consistent with the increased ruminal OM digestion.
The influence of supplemental monensin on fibre digestion has been variable. In the present study, monensin alone tended to increase ruminal and total tract NDF digestion. In some studies the influence of supplementation on NDF digestion has not been appreciable (Zinn and Borques Citation1993; Zinn et al. Citation1994). Whereas, in other studies, monensin reduced NDF digestion (Salinas-Chavira et al. Citation2009; Montano et al. Citation2015).
In the present study, essential oils supplementation without monensin, tended to increase flow of microbial N to the small intestine and total tract OM digestion. Ruminal digestion of OM and dietary N were not different for supplemental EO-alone and MON-alone, but they were not improved with the combination of the two additives. Essential oils supplementation without monensin, tended to increase ruminal NDF digestion. Yang et al. (Citation2010) observed that supplementation with essential oil blend (cinnamaldehyde-eugenol and capsicum) decreased ruminal NDF and CP digestion, but increased flow of microbial N flow to the small intestine. Cardozo et al. (Citation2005) observed decreased in vitro ammonia N concentration with capsicum and cinnamaldehyde supplementation (speculating that the response was due to decreased dietary protein deamination). In contrast, they observed increased ruminal ammonia N concentration with eugenol supplementation. Cardozo et al. (Citation2006) observed decreased in vitro ammonia N concentration with a blend of cinnamaldehyde and eugenol. However, they did not observe an effect of added capsicum on in vitro ammonia N concentration. Geraci et al. (Citation2012) observed decreased ruminal ammonia concentrations in cattle supplemented with a blend of cinnamaldehyde and eugenol vs cattle supplemented with monensin.
The lower ruminal pH and increased total VFA concentration observed with monensin alone may reflect the increased ruminal OM digestion. Likewise, Montano et al. (Citation2015) observed changes in ruminal VFA concentration and pH with monensin supplementation were consistent with changes in ruminal OM digestion. However, Zinn et al. (Citation1994) observed that whereas monensin supplementation tended to decrease ruminal pH, ruminal VFA concentration was not affected. In other studies (Salinas-Chavira et al. Citation2009), supplemental monensin did not affect ruminal pH or total VFA concentrations.
In the present study monensin supplementation did not affect molar proportions of acetate and propionate or estimated methane production. Improved growth performance of feedlot cattle with supplemental monensin has been attributed to increased propionate, and decreased acetate and butyrate, and methane energy loss (Richardson et al. Citation1979). However, this response has been quite variable. Whereas in some studies (Rogers et al. Citation1991; Zinn and Borques Citation1993; Surber and Bowman Citation1998; Guan et al. Citation2006) supplemental monensin was associated with decreased acetate:propionate molar ratios, in other studies (Zinn Citation1987; Zinn et al. Citation1994; Salinas-Chavira et al. Citation2009; Felix and Loerch Citation2011; Montano et al. Citation2015) ruminal VFA molar proportions were not affected. Observed changes in ruminal isovalerate molar proportion with supplemental monensin alone, are consistent with the observed increase in ruminal N digestion.
Essential oils supplementation without monensin decreased ruminal pH. Meyer et al. (Citation2009) did not observe and influence of either essential oils (blend of thymol, eugenol, vanillin, guaiacol, and limonene) or monensin on ruminal pH of feedlot steers.
Total ruminal VFA concentration was greater for essential oils alone or monensin alone than the combination of the two. Geraci et al. (Citation2012) reported numerically reduced ruminal pH and increased total VFA for an essential oil blend (mixture of cinnamaldehyde, eugenol and capsicum oleoresin) than monensin.
In the present study, supplemental essential oils did not influence on molar proportions of acetate and propionate or estimated methane production. Meyer et al. (Citation2009) observed similar molar proportion of acetate, but numerically greater molar proportion of propionate, and decreased acetate:propionate ratio with essential oils treatments. Yang et al. (Citation2010) observed decreased acetate and acetate:propionate ratio with supplemental eugenol. Geraci et al. (Citation2012) did not observe differences between supplemental monensin and essential oils on VFA molar rations.
Molar proportions of butyrate and isovalerate were greater for EO alone or monensin alone than the combination of the two. Observed changes in isovalerate are consistent with treatment effects on ruminal N digestion, as isovalerate is a product of ruminal branched chain amino acid degradation (Geraci et al. Citation2012; Salinas-Chavira et al. Citation2016).
The combination of supplemental monensin plus EO reduced ruminal OM digestion and total VFA. These effects are consistent with observed increase in ruminal pH. Further research is warranted to elucidate the basis for this response.
Treatment interactions on digestion and ruminal fermentation parallel observed effects on growth performance. In diets without EO, monensin improved rumen digestion of OM, feed N and total VFA. Whereas, in diets with EO, supplemental monensin had opposite effect.
5. Conclusion
Supplemental monensin increased ADG and DMI. Supplementation with either EO alone, or monensin alone tended to enhance both gain efficiency and the estimated NE value of the diet. However, these effects were negated (negative associative effect) when the two were fed in combination. Treatment effects on growth performance were consistent with corresponding effects on measures of digestive function.
Disclosure statement
No potential conflict of interest was reported by the author(s).
References
- An XP, Wang Y, Wang RF, Hao XR, Hu YC, Guo T, Zhang J, Wang WW, Shi XY, Han S, Qi JW. 2020. Effects of a blend of cinnamaldehyde, eugenol and capsicum oleoresin (CEC) on growth performance, nutrient digestibility, immune response and antioxidant status of growing ewes. Livest Sci. 234:1–6.
- AOAC. 2000. Official methods of analysis of AOAC international. 17th ed. Assoc. Off. Anal. Chem.; Gaithersburg, MD.
- Araujo RC, Daley DR, Goodall SR, Jalali S, Bisneto OG, Budde AM, Wagner JJ, Engle TE. 2019. Effects of a microencapsulated blend of essential oils supplemented alone or in combination with monensin on performance and carcass characteristics of growing and finishing beef steers. Appl Anim Sci. 35:177–184.
- Barreras A, Castro-Pérez BI, López-Soto MA, Torrentera NG, Montaño MF, Estrada-Angulo A, Ríos FG, Dávila-Ramos H, Plascencia A, Zinn RA. 2013. Influence of ionophore supplementation on growth performance, dietary energetics and carcass characteristics in finishing cattle during period of heat stress. Asian Australas J Anim Sci. 26:1553–1561.
- Benchaar C, Calsamiglia S, Chaves AV, Fraser GR, Colombatto D, McAllister TA, Beauchemin KA. 2008. A review of plant-derived essential oils in ruminant nutrition and production. Anim Feed Sci Technol. 145:209–228.
- Benchaar C, Duynisveld JL, Charmley E. 2006. Effects of monensin and increasing dose levels of a mixture of essential oil compounds and intake, digestion and growth performance of beef cattle. Can J Anim Sci. 86:91–96.
- Bergen WG, Purser DB, Cline JH. 1968. Effect of ration on the nutritive quality of rumen microbial protein. J Anim Sci. 27:1497–1501.
- Calsamiglia S, Busquet M, Cardozo PW, Castillejos L, Ferret A. 2007. Essential oils as modifiers of rumen microbial fermentation. J Dairy Sci. 90:2580–2595.
- Cardozo PW, Calsamiglia S, Ferret A, Kamel C. 2005. Screening for the effects of natural plant extracts at different pH on in vitro rumen microbial fermentation of a high-concentrate diet for beef cattle. J Anim Sci. 83:2572–2579.
- Cardozo PW, Calsamiglia S, Ferret A, Kamel C. 2006. Effects of alfalfa extract, anise, capsicum and a mixture of cinnamaldehyde and eugenol on ruminal fermentation and protein degradation in beef heifers fed a high concentrate diet. J Anim Sci. 84:2801–2808.
- Cobellis G, Trabalza-Marinucci M, Yu ZT. 2016. Critical evaluation of essential oils as rumen modifiers in ruminant nutrition: A review. Sci Total Environ. 545:556–568.
- Depenbusch BE, Drouillard JS, Loe ER, Higgins JJ, Corrigan ME, Quinn MJ. 2008. Efficacy of monensin and tylosin in finishing diets based on steam-flaked corn with and without corn wet distillers grains with solubles. J Anim Sci. 86:2270–2276.
- Devant M, Anglada A, Bach A. 2007. Effects of plant extract supplementation on rumen fermentation and metabolism in young Holstein bulls consuming high levels of concentrate. Anim Feed Sci Technol. 137:46–57.
- Duffield TF, Merrill JK, Bagg RN. 2012. Meta-analysis of the effects of monensin in beef cattle on feed efficiency, body weight gain, and dry matter intake. J Anim Sci. 90:4583–4592.
- European Commission. 2003. Regulation (EC) No. 1831/2003 of the European Parliament and of the Council of 22 September 2003 on Additives for Use in Animal Nutrition. Off. J. Eur. Union L:L268/29–L268/43.
- Felix TL, Loerch SC. 2011. Effects of haylage and monensin supplementation on performance, carcass characteristics, and ruminal metabolism of feedlot cattle fed diets containing 60% dried distillers grains. J Anim Sci. 89:2614–2623.
- Fox DG, Black JR. 1984. A system for predicting body composition and performance of growing cattle. J. Anim Sci. 58:725–739. doi:10.2527/jas1984.583725xm.
- Geraci JI, Garciarena AD, Gagliostro GA, Beauchemin KA, Colombatto D. 2012. Plant extracts containing cinnamaldehyde, eugenol and capsicum oleoresin added to feedlot cattle diets: ruminal environment, short term intake pattern and animal performance. Anim Feed Sci Technol. 176:123–130.
- Goodrich RD, Garrett JE, Gast DR, Kirick MA, Larson DA, Meiske JC. 1984. Influence of monensin on the performance of cattle. J Anim Sci. 58:1484–1498.
- Guan H, Wittenberg KM, Ominski KH, Krause DO. 2006. Efficacy of ionophores in cattle diets for mitigation of enteric methane. J Anim Sci. 84:1896–1906.
- Hill FW, Anderson DL. 1958. Comparison of metabolizable energy and productive energy determinations with growing chicks. J Nutr. 64:587–603.
- Khiaosa-ard R, Zebeli Q. 2013. Meta-analysis of the effects of essential oils and their bioactive compounds on rumen fermentation characteristics and feed efficiency in ruminants. J Anim Sci. 91:1819–1830.
- Mader TL, Davis MS, Brown-Brandl T. 2006. Environmental factors influencing heat stress in feedlot cattle. J Anim Sci. 84:712–719.
- Meyer NF, Erickson GE, Klopfenstein TJ, Greenquist MA, Luebbe MK, Williams P, Engstrom MA. 2009. Effect of essential oils, tylosin, and monensin on finishing steer performance, carcass characteristics, liver abscesses, ruminal fermentation, and digestibility. J Anim Sci. 87:2346–2354.
- Montano MF, Manriquez OM, Salinas-Chavira J, Torrentera N, Zinn RA. 2015. Effects of monensin and virginiamycin supplementation in finishing diets with distiller dried grains plus solubles on growth performance and digestive function of steers. J Appl Anim Res. 43:417–425.
- NRC. 1984. Nutrient Requirements of Beef cattle, 6th rev. ed. Washington, DC: National Academy Press.
- NRC. 1988. Nutrient requirement of dairy cattle, 6th Rev. Ed. Washington, DC: National Academy Press.
- NRC. 1996. Nutrient Requirements of Beef cattle, 7th rev. ed. Washington, DC: National Academy Press.
- Ornaghi MG, Passetti RAC, Torrecilhas JA, Mottin C, Vital ACP, Guerrero A, Sañudo C, del Mar Campo M, Prado IN. 2017. Essential oils in the diet of young bulls: effect on animal performance, digestibility, temperament, feeding behaviour and carcass characteristics. Anim Feed Sci Technol. 234:274–283.
- Ørskov ER, MacLeod NA, Kyle DJ. 1986. Flow of nitrogen from the rumen and abomasum in cattle and sheep given protein-free nutrients by intragastric infusion. Br J Nutr. 56:241–248.
- Ortiz J, Montaño M, Plascencia A, Salinas J, Zinn RA. 2016. Influence of kaolinite clay supplementation on growth performance and digestive function in finishing calf-fed Holstein steers. Asian australas. J Anim Sci. 29:1569–1575.
- Patra AK. 2011. Effects of essential oils on rumen fermentation, microbial ecology and ruminant production. Asian J Anim Vet Adv. 6:416–428.
- Poos MI, Hanson TL, Klopfenstein TJ. 1979. Monensin effects on diet digestibility, ruminal protein bypass and microbial protein synthesis. J Anim Sci. 48:1516–1524.
- Richardson LF, Raun AP, Potter EI, Cooley CO, Rathmacher RP. 1979. Effect of monensin on rumen fermentation in vitro and in vivo. J Anim Sci. 43:657–664.
- Rogers M, Jounany JP, Thivend P, Fontenot JP. 1991. Comparative effects of feeding and duodenal infusion of monensin on digestion in sheep. Can J Anim Sci. 71:1125–1133.
- Russell JB, Strobel HJ. 1988. Effects of additives on in vitro ruminal fermentation: a comparison of monensin and bacitracin, another gram-positive antibiotic. J Anim Sci. 66:552–558.
- Salinas-Chavira J, Barreras A, Plascencia A, Montano MF, Navarrete JD, Torrentera N, Zinn RA. 2016. Influence of protein nutrition and virginiamycin supplementation on feedlot growth performance and digestive function of calf-fed Holstein steers. J Anim Sci. 94:4276–4286.
- Salinas-Chavira J, Lenin J, Ponce E, Sanchez U, Torrentera N, Zinn RA. 2009. Comparative effects of virginiamycin supplementation on characteristics of growth-performance, dietary energetics, and digestion of calf-fed Holstein steers. J Anim Sci. 87:4101–4108.
- Samuelson KL, Hubbert ME, Galyean ML, Loest CA. 2016. Nutritional recommendations of feedlot consulting nutritionists: The 2015 New Mexico State and Texas Tech University survey. J Anim Sci. 94:2648–2663.
- Souza KAD, Monteschio JDO, Mottin C, Ramos TR, Pinto LADM, Eiras CE, Guerrero A, Prado IND. 2019. Effects of diet supplementation with clove and rosemary essential oils and protected oils (eugenol, thymol and vanillin) on animal performance, carcass characteristics, digestibility, and ingestive behavior activities for Nellore heifers finished in feedlot. Livest Sci. 220:190–195.
- Surber LMM, Bowman JGP. 1998. Monensin effects on digestion of corn or barley high-concentrate diets. J Anim Sci. 76:1945–1954.
- Thompson AJ, Smith ZKF, Corbin MJ, Harper LB, Johnson BJ. 2016. Ionophore strategy affects growth performance and carcass characteristics in feedlot steers. J Anim Sci. 94:5341–5349.
- Van Soest PJ, Robertson B, Lewis BA. 1991. Methods for dietary fiber, neutral detergent fiber, and non-starch polysaccharides in relation to animal nutrition. J Dairy Sci. 74:3583–3597.
- Wolin MJ. 1960. A theoretical rumen fermentation balance. J Dairy Sci. 43:1452–1459.
- Yang WZ, Benchaar C, Ametaj BN, Beauchemin KA. 2010. Dose response to eugenol supplementation in growing beef cattle: ruminal fermentation and intestinal digestion. Anim Feed Sci Technol. 158:57–64.
- Zinn RA. 1987. Influence of lasalocid and monensin plus tylosin on comparative feeding value of steam-flaked versus dry-rolled corn diets for feedlot cattle. J Anim Sci. 65:256–266.
- Zinn RA. 1988. Comparative feeding value of supplemental fat in finishing diets for feedlot steers supplemented with and without monensin. J Anim Sci. 66:213–227.
- Zinn RA. 1990. Influence of flake density on the comparative feeding value of steam-flaked corn for feedlot cattle. J Anim Sci. 68:767–775.
- Zinn RA, Borques JL. 1993. Influence of sodium bicarbonate and monensin on utilization of a fat-supplemented, high energy growing-finishing diet by feedlot cattle. J Anim Sci. 71:18–25.
- Zinn RA, Owens FN. 1986. A rapid procedure for purine measurement and its use for estimating net ruminal protein synthesis. Can J Anim Sci. 66:157–166.
- Zinn RA, Plascencia A, Barajas R. 1994. Performance and digestive function interaction of forage level and monensin in diets for feedlot cattle on growth. J Anim Sci. 72:2209–2215.
- Zinn RA, Shen Y. 1998. An evaluation of ruminally degradable intake protein and metabolizable amino acid requirements of feedlot calves. J Anim Sci. 76:1280–1289.