
ABSTRACT
In alphabetic languages, print consistently elicits enhanced, left-lateralized N170 responses in the event-related potential compared to control stimuli. In the current study, we adopted a cross-linguistic design to investigate N170 tuning to logographic Chinese and to pinyin, an auxiliary phonetic system in Chinese. The results demonstrated that logographic characters elicit a left-lateralized print-tuning effect in Chinese readers only. Crucially, the observed tuning effect is clearly driven by script familiarity, rather than by differences in visual features between print and control stimuli. This can be concluded because Dutch participants who viewed the same set of stimuli showed a bilateral topography instead. For pinyin, the left-hemispheric modulation was absent in both language groups, presumably because long strings of pinyin are unfamiliar to both groups. Because grapheme-to-phoneme conversion does not exist in logographic Chinese, our results tend to suggest a visual familiarity rather than a grapheme-to-phoneme mapping account of the print N170.
Introduction
Skilled reading depends on fast, automatized recognition of print (words or pseudowords). Converging evidence from neuroimaging studies suggests that the visual decoding of print is accomplished in the left ventral occipito-temporal cortex (Dehaene & Cohen, Citation2011; Dehaene, Cohen, Sigman, & Vinckier, Citation2005). This particular patch of the visual cortex has been labelled the visual word form system (VWFS; Cohen et al., Citation2000), as it is consistently activated for visually presented words (for a review, see Wandell, Citation2011). The VWFS appears to be tuned to print processing: In response to written words, fluent adult readers activate fast, specialized visual processes in the VWFS, enabling efficient categorization and recognition of the stimuli (Brem et al., Citation2006; Dehaene et al., Citation2005; van der Mark et al., Citation2009; Vinckier et al., Citation2007).
A robust neurophysiological marker for such transient visual specialization processes is the N170 component of the event-related potential (ERP), an occipito-temporal negativity peaking between 140 and 220 ms after stimulus onset. The N170 is consistently found to be larger for print than for visually matched control stimuli, even in implicit reading tasks (Bentin, Mouchetant-Rostaing, Giard, Echallier, & Pernier, Citation1999; Maurer, Brandeis, & McCandliss, Citation2005; Maurer, Brem, Bucher, & Brandeis, Citation2005; Schendan, Ganis, & Kutas, Citation1998), suggesting that such enhanced response is largely automatic. The N170 tuning effect for print is believed to develop with increasing exposure to written language (Brem et al., Citation2010; Maurer et al., Citation2007; Maurer et al., Citation2006), as the N170 component has long been known to be shaped by individual visual expertise in many domains. For example, bird experts showed larger N170 to bird than to dog stimuli, whereas dog experts demonstrated the opposite pattern (Tanaka & Curran, Citation2001). Similarly, familiarity with human faces leads to an enlarged N170 for face stimuli over visual baseline (Gauthier, Curran, Curby, & Collins, Citation2003; Latinus & Taylor, Citation2006). In contrast to the object and the face N170, which typically show a bilateral pattern, the print N170 is lateralized to the left hemisphere (Brandeis, Lehmann, Michel, & Mingronem, Citation1995; Brem et al., Citation2005; Rossion, Joyce, Cottrell, & Tarr, Citation2003). Furthermore, source localization studies have identified activity in the left VWFS as its metabolic correlate (Bentin et al., Citation1999; Helenius, Tarkiainen, Cornelissen, Hansen, & Salmelin, Citation1999; Provobio & Zani, 2003), suggesting that the print N170 forms an independent category that is specific to language, rather than being a variant of the object N170.
So far, most ERP studies on N170 tuning for print have focused on alphabetic languages, and the few conducted in the context of nonalphabetic languages have yielded inconsistent results. Although enlarged N170 responses to print (relative to control stimuli) have been generally reported, for example, for Japanese (Koyama, Kakigi, Hoshiyama, & Kitamura, Citation1998; Kuriki, Hirata, Fujimaki, & Kobayashi, Citation1996; Maurer, Zevin, & McCandliss, Citation2008; Shirahama, Ohta, Takashima, Matsushima, & Okubo, Citation2004), Korean (Kim, Yoon, & Park, Citation2004), and Chinese (Cao, Li, Zhao, Lin, & Weng, Citation2011; Lin et al., Citation2011; Wong, Gauthier, Woroch, DeBuse, & Curran, Citation2005; Yum, Su, & Law, Citation2015; Zhang et al., Citation2011; Zhao, Li, Cao, He, & Weng, Citation2012), the lateralization pattern of such effect remains obscure. On the one hand, left-hemispheric dominance has been found in Japanese (Kuriki et al., Citation1996) and Chinese (Cao et al., Citation2011; Lin et al., Citation2011; Wong et al., Citation2005; Yum et al., Citation2015; Zhao et al., Citation2012) readers. Notably in a cross-linguistic study by Maurer et al. (Citation2008), native Japanese readers, but not English-only readers, showed left-lateralized N170 responses to logographic Japanese script, indicating that the left-hemispheric modulation of the print N170 indexes reading expertise with a particular writing system. On the other hand, bilateral N170 responses have also been observed in Korean readers (who could read Chinese) viewing Chinese characters (Kim et al., Citation2004), native Chinese readers viewing Chinese characters (Zhang et al., Citation2011), and Japanese readers viewing Kana scripts (Koyama et al., Citation1998). Clearly, additional studies are needed to investigate N170 tuning for print in nonalphabetic languages, especially its lateralization pattern.
It is interesting that such studies may provide a window into the underlying mechanism of the print N170. Despite the consensus that the N170 print-tuning effect reflects extensive experience with visual words in one’s own language, the exact level at which the tuning takes place is still hotly debated. Although some studies relate the print N170 to grapheme-to-phoneme conversion processes underpinning reading in alphabetic languages (Maurer & McCandliss, Citation2007; Proverbio & Zani, Citation2003), others focus on low-level, perceptual aspects of print processing, attributing the observed tuning effect to visual familiarity with a particular script (Wong et al., Citation2005). In alphabetic languages, the transparent orthography–phonology relationship renders it difficult to disentangle “pure” visual aspects of print processing from evoked phonological processes. In this light, nonalphabetic languages, in which the mapping between sound and image is relatively arbitrary, may offer a unique opportunity to unravel the nature of print tuning.
In the present study, we thus examine N170 tuning for print in Chinese, a logographic script in which the primary unit (i.e., character) represents meaning rather than sound. In Chinese, although each character is realized phonologically as a syllable, constituent phonemes of the syllable are not atomistically represented within the corresponding character (McBride-Chang & Chen, Citation2003). Thus grapheme-to-phoneme conversion processes are absent in reading Chinese logographic script (Chen, Allport, & Marshall, Citation1996; Perfetti & Zhang, Citation1995). This renders Chinese particularly suitable for studying the underlying mechanism of print tuning: If the enhanced and left-lateralized print N170 is triggered by grapheme-to-phoneme conversion, such effect should be weak or even nonexistent in logographic Chinese; in contrast, if the print N170 is engendered by visual familiarity with a particular script, it should be observed for Chinese, as well. In addition to the logographic script, a phonetic pinyin system is used as an auxiliary means to code Chinese speech in written form. By adopting the Latin alphabet and additional tone-marking diacritics, pinyin facilitates the transcription of Chinese characters into the Latin alphabet. Although never used in formal texts as an independent script, pinyin is widely taught at the beginning of reading education to facilitate learning of the pronunciations of characters. It is interesting that pinyin maps letters to phonemes, as do alphabetic scripts. Such characteristics render pinyin an apt tool for studying the processing of sound-based scripts in Chinese readers. It is worth noting that because phonics instruction with pinyin ceases at the end of Grade 3 (i.e., for 10-year-olds), it is uncommon for Chinese adults to read pinyin, especially in long strings (more details are provided in the Discussion section).
Several print-tuning studies have been done in Chinese (Cao et al., Citation2011; Lin et al., Citation2011; Wong et al., Citation2005; Zhang et al., Citation2011; Zhao et al., Citation2012). In addition to the mixed results regarding lateralization (just discussed), existing studies suffer from confounds of low-level stimulus features. As mentioned in Maurer et al. (Citation2008), a common difficulty confronting print-tuning studies is to match control stimuli (e.g., symbols) with print for low-level visual features. This poses particular difficulties to studies with Chinese, because logographic characters, with their uniquely square, compact structure, differ from control stimuli in many aspects other than wordness (the status of being a word); these include visual complexity, string length, spatial structure, and so on. It is not yet clear which features are most critical to control for, and tests of visual similarity are rarely reported. It is thus possible that the N170 tuning effects, if observed at all, are (partially) caused by differences in low-level visual features between control and print stimuli and thus do not necessarily reflect reading expertise with the script per se. Yet another limitation with existing print-tuning studies in Chinese is that most of them have focused on single characters, whereas the majority of Chinese words are composed of two or more characters (Wood & Connelly, Citation2009). Last but not least, no study so far has examined the N170 tuning effect for pinyin.
Motivated by such a scenario, in the current study we investigate N170 tuning to logographic Chinese characters and to phonetic pinyin. To control for confounds of low-level stimulus features, we included a group of native Dutch readers as control participants. The Dutch participants were completely ignorant of logographic Chinese characters. Furthermore, although they might be familiar with individual pinyin graphemes, which resemble letters of the Latin alphabet used in Dutch, they are expected to be unfamiliar with long strings of pinyin, because spelling rules and orthographic patterns are radically different between Chinese and Dutch. During the experiment, native and naïve readers of Chinese performed a repetition detection task with Chinese words (composed of two to four characters), pinyin, and symbol strings. Because the two groups of participants viewed the same set of stimuli, any difference in their patterns of tuning, as indexed by the contrast between print (character or pinyin) and control (symbol) stimuli, must be attributed to their differential experience with the Chinese script rather than to low-level stimulus features. Our primary objective is to investigate whether enhanced, left-lateralized N170 response for print, which has consistently been found in alphabetic languages, can also be observed in logographic Chinese and phonetic pinyin script. Implications for the underlying mechanism of specialized visual processing for print are also addressed.
Methods
Participants
Native Chinese readers (n = 19) and native Dutch readers who could not read Chinese (n = 19) participated in the study. The two language groups were matched for age (Chinese: 23.7 years, SD = 2.4, range = 20.5–30.3; Dutch: 23.8 years, SD = 3.3, range = 20.3–33.5; p > .9) and gender ratio (Chinese: six male, 13 female; Dutch: seven male, 12 female). Both the Chinese and the Dutch participants were graduate or undergraduate students and were tested in the same ERP lab located in the Netherlands. All Chinese participants spoke fluent English but no other alphabetic languages; two of them had a rudimentary knowledge of Dutch. The Dutch participants were selected on the basis of their complete ignorance of Chinese language and its orthography. All participants were neurologically healthy, were right-handed, and had normal or corrected-to-normal vision. Written informed consent was gained from each participant prior to inclusion. Ethical approval was waived for the current study because the participants were healthy adults.
Materials
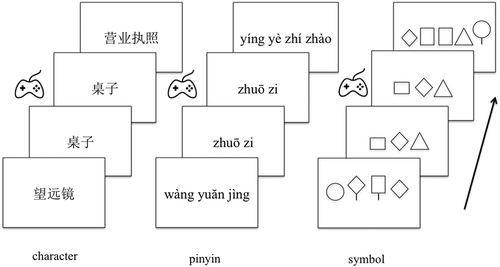
The participants performed a one-back repetition detection task with three conditions: Chinese words (in logographic characters), pinyin, and symbol strings. The Chinese words were high-frequency nouns consisting of two to four characters (average occurrences per million: 23.48; Cai & Brysbaert, Citation2010), extending 9.7–19.1 cm (M = 16.7 cm, SD = 2.5). The pinyin strings were phonetic transcriptions of the items in the character condition, extending 15–27.6 cm (M = 20.2 cm, SD = 3.4). The symbol strings consisted of three to five geometric symbols, similar to those used in previous studies (Brem et al., Citation2005; Maurer et al., Citation2007), extending 8.6–16.2 cm (M = 13.1 cm, SD = 2.2). The N170 is an automatic response to visual stimulation that can be elicited irrespective of the subjects’ attention or behavioural task. The purpose of having a task was to ensure that the participants viewed the stimuli.
The stimuli were shown in black on a white background in the centre of the screen. For each condition, the stimuli were divided into two blocks, each consisting of 48 items (i.e., 96 items in total); for each block, eight immediate repetitions were pseudo-randomly presented as targets (17%), avoiding two immediate repetitions in a row. The items were shown for 700 ms, followed by an interstimulus interval of 1,000 ms. The participants were seated 1.2 m away from the computer screen and were instructed to press a joystick button as quickly as possible after immediate stimulus repetition (see ).
Figure 1. Stimuli and experimental procedure. Stimulus duration: 700 ms; ISI: 1000 ms.
ERP recording and analysis
The electroencephalogram (EEG) was recorded on an ASA–Lab system, using a 64-channel WaveGuard cap with sintered Ag/AgCl electrodes (ANT Neuro Inc., Enschede, the Netherlands). Additional electrodes were used to record horizontal (electrodes positioned at the outer canthus of each eye) and vertical (electrodes positioned above and below the left eye) eye movements. Impedance was kept below 10kΩ. The EEG was digitized online at a sampling rate of 512 Hz and was referenced to the average of all channels.
For offline analyses, the acquired EEG data were corrected for eye blinks and horizontal eye movements using principal component analysis (Ille, Patrick, & Scherg, Citation2002). The corrected file was subsequently band-pass filtered (0.5–40 Hz) and segmented into trials of 1,625 ms duration, including a 125-ms baseline. Trials with extreme amplitudes (larger than ± 100 μV) were rejected, and the remaining trials were baseline corrected before averaging. Individual ERPs were then computed by averaging separately for each condition, including only nontarget trials. Target trials were excluded from further analyses because they were likely to be contaminated by finger movements involved in button pressing. Next grand averages of the condition ERPs were computed separately for each language group. For further statistical analysis, three pairs of occipito-temporal channels (i.e., P7/P8, O1/O2, PO7/PO8) were selected according to the maximum of the potential distribution on topographic maps (see ). Peak amplitudes of the N170 component at selected channels were determined by detecting the amplitude of the negative peak between 110 and 220 ms after stimulus onset. For most of the participants, there was only one negative peak; in case there was more than one peak, we selected the first negative peak following P1. Mean N170 peak amplitude for each hemisphere was computed by averaging across selected channels per hemisphere.
Figure 2. Topographic maps for the character - symbol difference (left) and the pinyin - symbol difference (right), separately for the Chinese (upper) and the Dutch (lower) group.

Statistical analysis
Behavioural data
To assess task involvement of the participants, accuracy and reaction time of their behavioural responses were each analysed using a repeated measures analysis of variance (ANOVA) with the within-subject factor of condition (character, pinyin or symbol) and the between-subject factor of language group (Chinese vs. Dutch). Because the distributions of accuracy scores and reaction times were significantly skewed, transformations were applied (arcsine and logarithmic transformation, respectively; Winer, Brown, & Michels, Citation1991), which normalized the distributions.
ERP data
To investigate N170 tuning to logographic Chinese characters and to phonetic pinyin, two repeated measures ANOVAs were performed with within-subject factors of condition (character vs. symbol or pinyin vs. symbol) and hemisphere (left vs. right) and the between-subject factor of language group (Chinese vs. Dutch).
Results
Behavioural data
Accuracy and reaction time measures are summarized in . In general, repetitions of characters and pinyin were more accurately detected than repetitions of symbol strings: condition, F(2, 72) = 6.02, p < .01. This pattern was more pronounced in the Chinese participants, presumably because they benefited from their knowledge of the words represented by these character strings (Condition × Language Group), F(2, 72) = 8.76, p < .001. Post hoc pairwise comparisons showed that the Chinese participants were more accurate in the character condition than in both other conditions (p < .05); the Dutch participants were most accurate in the pinyin condition (p < .05).
Table 1. Behavioural results: Accuracy and reaction time for the repetition detection task.
In terms of reaction time, the Dutch participants generally responded faster than the Chinese participants: language group, F(1, 36) = 6.93, p < .05. In addition, there was a significant main effect of condition, F(2, 72) = 20.89, p < .001, and a significant interaction effect of Condition × Language Group, F(2, 72) = 4.64, p < .05. Post hoc tests per language group showed that the Chinese participants responded significantly slower in the pinyin than in the character or the symbol string condition (p < .001), whereas the Dutch participants responded the fastest in the symbol string condition (p < .05). Overall, the high accuracy and speed indicated that participants were attending to the stimuli during the experiment.
ERP data
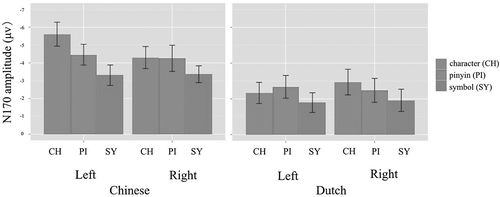
displays the average ERPs recorded over the occipito-temporal region. In both language groups, a negative peak around 170 ms was clearly visible for all stimulus types and all electrodes. Topographic maps for the tuning effects (i.e., character/pinyin–symbol contrast) are presented in . shows the N170 peak amplitudes for each language group, hemisphere and condition, averaged over the three electrodes selected per hemisphere.
Figure 3. Average ERPs at occipito-temporal channels.
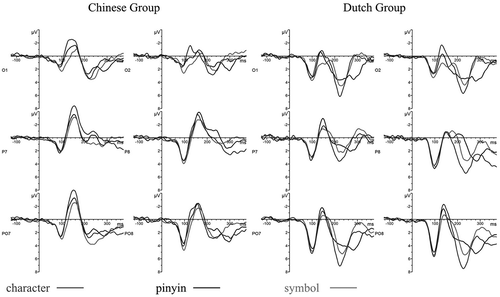
Figure 4. Mean N170 amplitude by group, hemisphere and condition. Error bars indicate standard deviations.
N170 tuning to logographic Chinese characters: Character versus symbol
An overall analysis of variance was performed on the combined data of both groups with condition (character vs. symbol), hemisphere (left vs. right), and language group (Chinese vs. Dutch) as independent variables and the amplitude of brain responses as dependent variable. In general, characters evoked larger N170 responses than symbol strings: condition, F(1, 36) = 23.16, p < .001. In addition, Chinese participants showed larger N170 than Dutch participants: language group, F(1, 36) = 6.54, p < .05. Crucially, although the overall size of the character–symbol N170 amplitude difference did not differ between the two language groups (Condition × Language Group), F(1, 36) = 2.83, p > .1, the lateralization pattern of this difference did (Language Group × Condition × Hemisphere), F(1, 36) = 13.71, p < .001.
Planned comparisons were performed separately for each language group, with condition and hemisphere as independent variables and the amplitude of brain responses as a dependent variable. The analyses revealed that in Chinese participants, the character–symbol N170 amplitude difference was highly significant—condition, F(1, 18) = 15.32, p < .001—and that this difference was strongly left lateralized (Condition × Hemisphere), F(1, 18) = 14.87, p < .001. Dutch participants, who had no experience with written Chinese, also responded more strongly to Chinese characters than to symbol strings, although with lower significance, condition, F(1, 18) = 7.86, p < .05. Different from Chinese participants, Dutch participants showed a similar character–symbol N170 amplitude difference for both hemispheres (Condition × Hemisphere), F(1, 18) = 1.88, p > .1.
N170 tuning to phonetic pinyin script: Pinyin versus symbol
An overall ANOVA was performed on the combined data of both groups with condition (pinyin vs. symbol), hemisphere (left vs. right), and language group (Chinese vs. Dutch) as independent variables and the amplitude of brain responses as a dependent variable. In general, the N170 response was stronger for pinyin than for symbol strings—condition, F(1, 36) = 13.85, p < .001—and stronger in Chinese participants than in Dutch participants: language group, F(1, 36) = 4.71, p < .05. There was no significant interaction effect between condition, language group, or hemisphere. Separate group analyses revealed that both language groups showed an enlarged N170 response to pinyin compared with symbol strings—condition: Chinese group, F(1, 18) = 12.33, p < .01; Dutch group: F(1, 18) = 3.86, p < .05. Neither group showed a lateralization effect for the pinyin–symbol difference in N170 amplitude (Condition × Hemisphere), Chinese group: F(1, 18) = 0.37, p > .5; Dutch group: F(1, 18) = 0.53, p > .4).
Discussion
The current study aimed to investigate specialized visual brain processes engaged in reading Chinese, a logographic script that links characters with meaning rather than with sound. For the first time, we adopted a cross-linguistic design to study N170 tuning for print in Chinese, in which native Chinese readers and Dutch readers who were completely ignorant of written Chinese detected immediate repetitions in sequences of Chinese words, pinyin, and symbol strings. Because the two groups were exposed to the same set of stimuli, any difference in their patterns of tuning must be attributed to their differential experience with the Chinese script rather than to differences in visual features between print and control stimuli. In this way, we ruled out potential confounds of low-level stimulus features, which have posed a persistent challenge to print-tuning studies in Chinese. With such confounds properly controlled, we were able to increase the visual complexity of the character stimuli, that is, to include Chinese words composed of multiple characters. Furthermore, we extended the current understanding of the print-tuning phenomenon in Chinese by including pinyin, an auxiliary phonetic system for transcribing logographic Chinese. The outcome of this research was also expected to provide insights into the underlying mechanisms of specialized visual processing for print. In sum, the results demonstrated that both groups responded more strongly to characters than to symbols, but the character–symbol difference was significantly left lateralized in the Chinese group only; the Dutch group showed a bilateral topography instead. With respect to pinyin, both Chinese and Dutch participants showed larger N170 responses to pinyin than to symbol strings; the left-hemispheric modulation, however, was absent in both groups.
Print tuning to logographic Chinese characters
Our analyses revealed that, similar to alphabetic words, logographic Chinese characters evoked a larger N170 response than control stimuli (i.e., symbol strings), especially in the left hemisphere. This adds to existing evidence of enhanced and left-lateralized N170 responses for individual Chinese characters (Cao et al., Citation2011; Lin et al., Citation2011; Wong et al., Citation2005; Yum et al., Citation2015); further, we expand upon previous findings by showing that such tuning effect extends to whole words composed of multiple characters. Crucially, our results clearly demonstrate that the left-lateralized N170 print-tuning effect is driven by expertise with a particular script, rather than by differences in visual features between print and control stimuli. We came to this conclusion because the Dutch participants, who were exposed to the same task and stimuli, showed a bilateral instead of left-lateralized character–symbol difference in N170 amplitude.
Although the left-lateralization of the print N170 is well-established in alphabetic languages, it is less so in logographic Chinese. Both left-lateralized and bilateral topographies have been reported (Cao et al., Citation2011; Kim et al., Citation2004; Lin et al., Citation2011; Maurer et al., Citation2008; Wong et al., Citation2005; Zhang et al., Citation2011). The inconsistency is likely caused by differences in task demand across studies. For example, Kim and colleagues (Citation2004) found bilateral N170 responses to Chinese characters using a semantic decision task, which has been known to engage right-hemisphere processing (Bookheimer, Citation2002). On the other hand, left-lateralized N170 topography is more common in implicit reading tasks, such as repetition detection (e.g., Maurer et al., Citation2008; the present study) or colour-matching tasks (e.g., Lin et al., Citation2011).
Notably the Dutch participants, who had no experience with written Chinese, also responded more strongly to Chinese characters than to symbol strings, presumably because the former were visually more complex than the latter. This between-condition difference, which indexes N170 tuning effect for logographic Chinese, was significant in both language groups. This is consistent with previous findings of enhanced N1 responses for long stimuli as compared to short stimuli (Wong et al., Citation2005; Wydell, Vuorinen, Helenius, & Salmelin, Citation2003; Xue, Jiang, Chen, & Dong, Citation2008). As noted by Xue et al. (Citation2008), visual complexity strongly modulates N1 amplitude so that longer, more complex stimuli evoke larger N1 responses; this is especially true when the stimuli are visually unfamiliar (e.g., Chinese logographic characters and symbol strings for Dutch participants in the current study).
To summarize, although both language groups showed larger N170 responses to characters than to symbol strings, the lateralization pattern of this tuning effect differed significantly between native and naïve readers of Chinese, as demonstrated by a left-lateralized topography in Chinese participants and a bilateral topography in Dutch participants. Taken together, our results suggest that enhanced N170 responses to print compared to visual baseline may be (partially) due to visual complexity, whereas the left-lateralization pattern of the print N170 is clearly driven by reading expertise with a particular script. This aligns with consistently reported findings of left-lateralized print N170 in alphabetic languages, suggesting that the left-hemispheric modulation is a characterizing feature of the print N170 across languages and that similar visual specialization processes underpin reading in both alphabetic and logographic scripts. Because the current study focused specifically on visual tuning for print in Chinese, we did not include Dutch words. To explicitly test the universality of the print N170, further studies are needed to investigate N170 tuning to logographic and alphabetic stimuli within the same groups of participants.
Print tuning to phonetic pinyin script
With regard to the N170 print-tuning effect for pinyin, our analyses revealed that both Chinese and Dutch participants showed enhanced, bilateral N170 responses to pinyin compared to symbol strings. Such enhanced responses presumably reflect script familiarity with pinyin: For Chinese participants, pinyin is an auxiliary system for transcribing logographic characters; for Dutch participants, pinyin resembles the Latin alphabet except for the usage of tone-marking diacritics. However, the possibility cannot be ruled out that the observed tuning effects (in both groups) were partially triggered by low-level stimulus features. As just discussed, visual complexity strongly influences N1 amplitude. Following this line of reasoning, enhanced N170 responses to pinyin strings may be attributed to the fact that they were longer than the symbol strings (average string length: pinyin = 20.2 cm, symbol = 13.1 cm).
Because pinyin maps letters to phonemes as do alphabetic scripts, one would expect the N170 tuning effect for pinyin to be left lateralized. Our results, however, demonstrated a bilateral topography of the pinyin–symbol difference in both groups. The lack of left-hemispheric modulation in the Chinese participants may reflect the fact that they are less familiar with pinyin than with the logographic script. As a method to facilitate associations between characters and spoken words, pinyin is presented only at the initial stage of reading education to denote the pronunciation of the corresponding character; at the age of 10, children start to read solely in the logographic script. For adult readers, pinyin serves primarily as one of the input methods for entering Chinese characters into computers using alphabetic keyboards (except for that, pinyin is used only to spell Chinese geographic and personal names in foreign publications). Due to widely implemented associative relation functions, however, input efficiency is greatly improved, and one seldom needs to enter the entire pinyin string for multicharacter words; in most cases, one merely enters (part of) the pinyin for the first character and the rest of the word will be displayed automatically (among several alternatives) on the screen. It is therefore uncommon for Chinese adults to process pinyin in long strings. Similarly for the Dutch participants, though individual pinyin graphemes can be read in analogy to Latin alphabets, the long strings pose a challenge for recognition. Because the N170 print-tuning effect is specific to strings of letters and is typically not evoked by individual letters (Maurer et al., Citation2005b; Pernet et al., Citation2003), the lack of visual familiarity for pinyin strings may account for the absence of left-lateralized N170 responses for pinyin in both Chinese and Dutch participants.
Underlying mechanism of N170 tuning for print
The underlying mechanism of the print-tuning phenomenon is still hotly debated. Whereas some researchers attribute the tuning effect to grapheme-to-phoneme conversion processes, others highlight the role of visual familiarity with the script. In the current study, we found enhanced and left-lateralized N170 responses for Chinese, a logographic script in which grapheme-to-phoneme conversion rules do not apply, thus adding to the evidence for the perceptual familiarity account. In support of this idea, the left-hemispheric modulation of the print N170 was absent for pinyin, a phonetic script that engages grapheme-to-phoneme conversion but is encountered far less frequently than the logographic script by adult readers.
We acknowledge, however, that visual word recognition in Chinese may be (partially) modulated by phonological activation in a broader sense, if not by grapheme-to-phoneme conversion in particular. According to the universal phonological principle theory (Perfetti & Harris, Citation2013; Perfetti, Zhang, & Berent, Citation1992), all reading engages phonology at the earliest moment and the smallest unit allowed by the writing system. A similar account was proposed by the grain size theory (Ziegler & Goswami, Citation2005), that is, reading procedures assemble phonology according to the grain size of orthography. Specifically, the small grain size of alphabets allows a phoneme-level assembly of phonology, whereas the large grain size of Chinese logographs supports syllable-level assembly. Under these universal hypotheses, reading in both alphabetic and logographic scripts engages phonology; what differs between them is the way phonology is implemented into the writing system (Perfetti, Cao, & Booth, Citation2013). Supporting this view, there is good evidence that phonology is activated when Chinese characters are read for meaning (see Perfetti, Liu, & Tan, Citation2005, for a review). Following this line of reasoning, phonology may very well influence visual word recognition in Chinese, thus playing a role in shaping the left-lateralized print N170 for logographic characters.
Conclusion
To sum up, fluent adult readers of Chinese showed enhanced, left-lateralized N170 responses to logographic characters compared to visual baseline (i.e., symbols). The observed tuning effect was clearly driven by native readers’ familiarity with the script rather than by differences in visual features between print and baseline. This can be concluded because such pattern was absent in control participants viewing the same set of stimuli. This aligns with consistently reported findings of left-lateralized print N170 in alphabetic languages, suggesting that the left-hemispheric modulation is a characterizing feature of the print N170 across languages. On the other hand, the left-hemispheric modulation was absent for the N170 tuning effect for pinyin, possibly because the long strings of pinyin were visually unfamiliar to both groups of participants. Taken together, our findings point to a perceptual origin of the N170 print-tuning effect, that is, it is visual expertise with a particular script that leads to enhanced and left-lateralized N170 response for print. We acknowledge, however, that phonological activation in general, if not grapheme-phoneme conversion in particular, may also contribute to the N170 print-tuning effect in Chinese.
Funding
This work was funded by the Erasmus Mundus PhD Program IDEALAB (University of Groningen, University of Potsdam, Macquarie University, Newcastle University, and University of Trento).
Additional information
Funding
References
- Bentin, S., Mouchetant-Rostaing, Y., Giard, M., Echallier, J., & Pernier, J. (1999). ERP manifestations of processing printed words at different psycholinguistic levels: Time course and scalp distribution. Journal of Cognitive Neuroscience, 3, 235–260. doi:10.1162/089892999563373
- Bookheimer, S. (2002). Functional MRI of language: New approaches to understanding the cortical organization of semantic processing. Annual Review of Neuroscience, 25, 151–188. doi:10.1146/annurev.neuro.25.112701.142946
- Brandeis, D., Lehmann, D., Michel, C., & Mingronem, W. (1995). Mapping event-related brain potential microstates to setence endings. Brain Topography, 8(2), 145–159. doi:10.1007/BF01199778
- Brem, S., Bach, S., Kucian, K., Guttorm, T., Martin, E., Lyytinen, H., & Richardson, U. (2010). Brain sensitivity to print emerges when children learn letter-speech sound correspondences. Proceedings of the National Academy of Science, USA, 103, 7939–7944. doi:10.1073/pnas.0904402107
- Brem, S., Bucher, K., Halder, P., Summers, P., Dietrich, T., Martin, E., & Brandeis, D. (2006). Evidence for developmental changes in the visual word processing network beyond adolescence. NeuroImage, 29, 822–837. doi:10.1016/j.neuroimage.2005.09.023
- Brem, S., Lang-Dullenkopf, A., Maurer, U., Halder, P., Bucher, K., & Brandeis, D. (2005). Neurophysiological signs of rapidly emerging visual expertise for symbol strings. NeuroReport, 16(1), 45–48. doi:10.1097/00001756-200501190-00011
- Cai, Q., & Brysbaert, M. (2010). SUBTLEX-CH: Chinese word and character frequencies based on film subtitles. Plos ONE, 5(6), e10729. doi:10.1371/journal.pone.0010729
- Cao, X., Li, S., Zhao, J., Lin, S., & Weng, X. (2011). Left-lateralized early neurophysiological response for Chinese characters in young primary school children. Neuroscience Letters, 492, 165–169. doi:10.1016/j.neulet.2011.02.002
- Chen, Y., Allport, D., & Marshall, J. (1996). What are the functional orthographic units in Chinese word recognition: The stroke or the stroke pattern? Quarterly Journal of Experimental Psychology Section A: Human Experimental Psychology, 49, 1024–1043. doi:10.1080/713755668
- Cohen, L., Dehaene, S., Naccache, L., Lehericy, S., Dehaene-Lambertz, G., Henaff, M.-A., & Michel, F. (2000). The visual word form area: Spatial and temporal characterization of an initial stage of reading in normal subjects and posterior split-brain patients. Brain, 123, 291–307. doi:10.1093/brain/123.2.291
- Dehaene, S., & Cohen, L. (2011). The unique role of the visual word form area in reading. Trends in Cognitive Sciences, 15(6), 254–262. doi:10.1016/j.tics.2011.04.003
- Dehaene, S., Cohen, L., Sigman, M., & Vinckier, F. (2005). The neural code for written words: A proposal. Trends in Cognitive Sciences, 9(7), 335–341. doi:10.1016/j.tics.2005.05.004
- Gauthier, I., Curran, T., Curby, K., & Collins, D. (2003). Perceptual interference supports a non-modular account of face processing. Nature Neuroscience, 6(4), 428–432. doi:10.1038/nn1029
- Helenius, P., Tarkiainen, A., Cornelissen, P., Hansen, P. C., & Salmelin, R. (1999). Dissociation of normal feature analysis and deficient processing of letter-strings in dyslexic adults. Cerebral Cortex, 9, 476–483. doi:10.1093/cercor/9.5.476
- Ille, N., Patrick, B., & Scherg, M. (2002). Artifact correction of the ongoing EEG using spatial filters based on artifact and brain signal topographies. Journal of Clinical Neurophysiology, 19(2), 113–124. doi:10.1097/00004691-200203000-00002
- Kim, K., Yoon, H., & Park, H. (2004). Spatiotemporal brain activation pattern during word/picture perception by native Koreans. NeuroReport, 15, 1099–1103. doi:10.1097/00001756-200405190-00003
- Koyama, S., Kakigi, R., Hoshiyama, M., & Kitamura, Y. (1998). Reading of Japanese Kanji (morphograms) and Kana (syllabograms): A magnetoencephalographic study. Neuropsychologia, 36, 83–98. doi:10.1016/S0028-3932(97)00097-3
- Kuriki, S., Hirata, Y., Fujimaki, N., & Kobayashi, T. (1996). Magnetoencephalographic study on the cerebral neural activities related to the processing of visually presented characters. Cognitive Brain Research, 4, 185–199. doi:10.1016/S0926-6410(96)00030-4
- Latinus, M., & Taylor, M. (2006). Face processing stages: Impact of difficulty and the separation of effects. Brain Research, 1123(1), 179–187. doi:10.1016/j.brainres.2006.09.031
- Lin, S., Chen, H., Zhao, J., Li, S., He, S., & Weng, X. (2011). Left-lateralized N170 response to unpronounceable pseudo but not false Chinese characters - The key role of orthography. Neuroscience, 190, 200–206. doi:10.1016/j.neuroscience.2011.05.071
- Maurer, U., Brandeis, D., & McCandliss, B. (2005). Fast, visual specialization for reading in English revealed by the topography of the N170 ERP response. Behavioral and Brain Functions, 1, 13. doi:10.1186/1744-9081-1-13
- Maurer, U., Brem, S., Bucher, K., & Brandeis, D. (2005). Emerging neurophysiological specialization for letter strings. Journal of Cognitive Neuroscience, 17, 1532–1552. doi:10.1162/089892905774597218
- Maurer, U., Brem, S., Bucher, K., Kranz, F., Benz, R., Steinhausen, H.-C., & Brandeis, D. (2007). Impaired tuning of a fast occipito-temporal response for print in dyslexic children learning to read. Brain, 130, 3200–3210. doi:10.1093/brain/awm193
- Maurer, U., Brem, S., Kranz, F., Bucher, K., Benz, R., Halder, P., & Brandeis, D. (2006). Coarse neural tuning for print peaks when children learn to read. NeuroImage, 33, 749–758. doi:10.1016/j.neuroimage.2006.06.025
- Maurer, U., & McCandliss, B. (2007). The development of visual expertise for words: The contribution of electrophysiology. In E. L. Grigorenko & A. J. Naples (Eds.), Single-word reading: Biological and behavioral perspectives (pp. 43–64). Mahwah, NJ: Erlbaum.
- Maurer, U., Zevin, J., & McCandliss, B. (2008). Left-lateralized N170 effects of visual expertise in reading: Evidence from Japanese syllabic and logographic scripts. Journal of Cognitive Neuroscience, 20, 1878–1891. doi:10.1162/jocn.2008.20125
- McBride-Chang, C., & Chen, H. (2003). Reading development in Chinese Children. Westport, CT: Praeger.
- Perfetti, C., Cao, F., & Booth, J. (2013). Specialization and universals in the development of reading skill: How Chinese research informs a universal science of reading. Scientific Studies of Reading, 17(1), 5–21. doi:10.1080/10888438.2012.689786
- Perfetti, C., & Harris, L. (2013). Universal reading processes are modulated by language and writing system. Language Learning and Development, 9, 296–316. doi:10.1080/15475441.2013.813828
- Perfetti, C., Liu, Y., & Tan, L. (2005). The lexical constituency model: Some implications of research on Chinese for general theories of reading. Psychological Review, 12(11), 43–59. doi:10.1037/0033-295X.112.1.43
- Perfetti, C., & Zhang, S. (1995). Very early phonological activation in Chinese reading. Journal of Experimental Psychology: Learning, Memory, and Cognition, 21, 24–33.
- Perfetti, C., Zhang, S., & Berent, I. (1992). Reading in English and Chinese: Evidence for a “universal” phonological principle. In R. Frost & L. Katz (Eds.), Orthography, phonology, morphology, and meaning (pp. 227–248). Amsterdam, the Netherlands: North-Holland.
- Pernet, C., Basan, S., Doyon, B., Cardebat, D., Démonet, J. F., & Celsis, P. (2003). Neural timing of visual implicit categorization. Brain Research, Cognitive Brain Research, 17, 327–338. doi:10.1016/S0926-6410(03)00134-4
- Proverbio, A. M., & Zani, A. (2003). Time course of brain activation during graphemic/phonologic processing in reading: An ERP study. Brain and Language, 87, 412–420. doi:10.1016/S0093-934X(03)00139-1
- Rossion, B., Joyce, C., Cottrell, G., & Tarr, M. (2003). Early lateralization and orientation tuning for face, word, and object processing in the visual cortex. NeuroImage, 20(1), 1609–1624. doi:10.1016/j.neuroimage.2003.07.010
- Schendan, H., Ganis, G., & Kutas, M. (1998). Neurophysiological evidence for visual perceptual categorization of words and faces within 150 ms. Psychophysiology, 35(3), 240–251. doi:10.1111/psyp.1998.35.issue-3
- Shirahama, Y., Ohta, K., Takashima, A., Matsushima, E., & Okubo, Y. (2004). Magnetic brain activity elicited by visually presented symbols and Japanese characters. NeuroReport, 15, 771–775. doi:10.1097/00001756-200404090-00006
- Tanaka, J., & Curran, T. (2001). A neural basis for expert object recognition. Psychological Science, 12(1), 43–47. doi:10.1111/1467-9280.00308
- van der Mark, S., Bucher, K., Maurer, U., Schulz, E., Brem, S., Buckelmuller, J., … Brandeis, D. (2009). Children with dyslexia lack multiple specializations along the visual word-form (VWF) system. NeuroImage, 47, 1940–1949. doi:10.1016/j.neuroimage.2009.05.021
- Vinckier, F., Dehaene, S., Jobert, A., Dubus, J., Sigman, M., & Cohen, L. (2007). Hierarchical coding of letter strings in the ventral stream: Dissecting the inner organization of the visual word-form system. Neuron, 55, 143–156. doi:10.1016/j.neuron.2007.05.031
- Wandell, B. (2011). The neurobiological basis of seeing words. Annals of the New York Academy of Sciences, 1224, 63–80. doi:10.1111/j.1749-6632.2010.05954.x
- Winer, B. J., Brown, D. R., & Michels, K. M. (1991). Statistical principles in experimental design. New York, NY: McGraw-Hill.
- Wong, A., Gauthier, I., Woroch, B., DeBuse, C., & Curran, T. (2005). An early electrophysiological response associated with expertise in letter perception. Cognitive, Affective & Behavioral Neuroscience, 5, 306–318. doi:10.3758/CABN.5.3.306
- Wood, C., & Connelly, V. (2009). Contemporary perspectives on reading and spelling. New York, NY: Routledge.
- Wydell, T. N., Vuorinen, T., Helenius, P., & Salmelin, R. (2003). Neural correlates of letter-string length and lexicality during reading in a regular orthography. Journal of Cognitive Neuroscience, 15(7), 1052–1062. doi:10.1162/089892903770007434
- Xue, G., Jiang, T., Chen, C., & Dong, Q. (2008). Language experience shapes early electrophysiological responses to visual stimuli: The effect of writing system, stimulus length, and presentation duration. NeuroImage, 39, 2025–2037. doi:10.1016/j.neuroimage.2007.10.021
- Yum, Y. N., Su, I.-F., & Law, S.-P. (2015). Early effects of radical position legality in Chinese: An ERP study. Scientific Studies of Reading, 19, 456–467. doi:10.1080/10888438.2015.1081204
- Zhang, M., Jiang, T., Mei, L., Yang, H., Chen, C., Xue, G., & Dong, Q. (2011). It’s a word: Early electrophysiological response to the character likeness of pictographs. Psychophysiology, 48, 950–959. doi:10.1111/psyp.2011.48.issue-7
- Zhao, J., Li, S., Cao, X., He, S., & Weng, X. (2012). Selectivity of N170 in the left hemisphere as an electrophysiological marker for expertise in reading Chinese. Neuroscience Bulletin, 28(5), 577–584. doi:10.1007/s12264-012-1274-y
- Ziegler, J. C., & Goswami, U. (2005). Reading acquisition, developmental dyslexia, and skilled reading across languages: A psycholinguistic grain size theory. Psychological Bulletin, 131(1), 3–29. doi:10.1037/0033-2909.131.1.3