Abstract
Potato cooking was analysed on the basis of kinetic theory of potato softening. The application of the theory makes it possible to connect cooking time, i.e., time of cooking prior to the start of disintegration of the potato, with some terms of the kinetic theory, mainly kinetic coefficient and critical concentration of the intracellular bonds. Simple models were developed for description of the disintegration part in terms of the cooking part parameters. Two years of data from three different varieties, in one case in six different cultivation modes, shows that deep discrepancy exists between parameters for the non-salad varieties, which correlated with each other, and the parameters obtained for the salad variety Nicola. Nicola differed from the other analysed varieties mainly much lower cooking sensitivity to starch content. This difference was the main source of the observed loss of correlation among the different cooking parameters.
INTRODUCTION
The phenomenon of “sloughing”, the flaking and disintegration of the outer layers of potato tubers cooked in water, is considered to be one of the principal characteristics of potato texture.[Citation1] Potatoes are classified into different cooking types that are part of the potato texture profile. Potato texture results from many factors, including starch content, cell size, cell-wall composition, starch swelling pressure and breakdown of the cell-wall middle lamella during cooking.[Citation2,Citation3],[Citation4] Among these factors, tuber density and the degree of disintegration after cooking are often correlated, particularly within one cultivar. However, contradictory results have also been reported regarding the relationship between specific gravity and sloughing.[Citation1]
Different modifications of sloughing tests based on measurements of the degree of cell separation were used.[Citation4,Citation5,Citation6][Citation7,Citation8] Direct methods for assessment of sloughing are mostly based on CPM (cooked potato mass) tests. This group of tests used to be referred to as CPW (cooked potato weight) tests. During the CPM test,[Citation9] potato flakes are cooked in a stirred water bath then decanted and sluiced down on a sieve with 2 mm mesh. The mass of the remaining cooked potato tissue on the sieve is recorded and the characteristic time CT100 (the maximum cooking time up to which the initial 100 g potato mass remains on the sieve) is determined by interpolation as a measure for the resistance to sloughing.
The CPM test was modified into the CPEM (cooked potato effective mass) sloughing method in our previous paper.[Citation10] The CPEM method involves cooking the potato flakes on the sieve in a stirred water bath and determining their effective mass periodically during cooking. The effective mass is the difference between the real mass of the cooked specimen and the mass of the actual cooking medium of the same volume. The method was set up in such a way that temperature and density variations in both the tested sample and the cooking medium during a test did not affect results. The CPEM method enables us to determine cooking properties of small samples consisting of only a few tubers. Two main parts are distinguished (): the cooking part and the breaking part, from which two main parameters were derived. The cooking part represents the part of test in which no sample disintegration is observed. In contrast to the cooking part, the sample disintegration is characteristic for the breaking part. The cooking time (CT) corresponds to the transition from the cooking to the breaking part. In the breaking part, the initial part was fitted linearly with time and the absolute value of the obtained slope was called the slope of the breaking part (SBP).
Figure 1 Scheme of the CPEM cooking curve: CT (cooking time) defined as intersection of the IEM (initial effective mass) and the linear approximation of the breaking part by excluding its last part, SBP (slope of the breaking part) derived as the slope of the approximation with opposite sign.
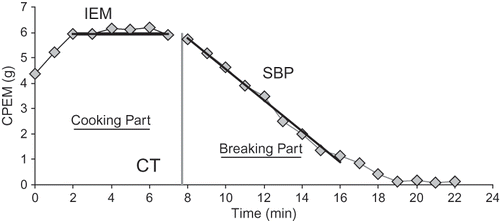
The CPEM sloughing method was applied to cooking properties assessment of potato tubers with different density, i.e., with different content of starch for three potato varieties (Nicola, Agria, and Saturna) in the other previous paper.[Citation11] It was shown that both the basic cooking parameters: CT and SBP are described in frames of the individual variety as linear functions of tuber density.
Cooking softening can be described in terms either by the chemical kinetics[Citation12] or in terms of changes in the tuber texture.[Citation13] In the first case, the simplest kinetic equation of the first order can be applied:
Parameter cu in EquationEqs. (2a) and Equation(2b) represents the asymptotic bond concentration at infinite time; it could be understood as the remained bond concentration after thermal processing. In this paper the results obtained previously in the CPEM potato tests are analysed using a simple theory based on the cooking kinetics.
RESULTS AND DISCUSSION
Cooking Part
The Cooking Part of a CPEM curve is accompanied by molecular bond destruction in the whole volume of the cooked tuber tissue. This process can be simply described by EquationEq. (1) with kinetic coefficient k dependent on the cooking temperature. The higher the kinetic coefficient indicates the quicker the cooking process. Texture changes during cooking can be described by the same formulas as (1a) and (1b), in which the bond concentration is replaced by some important texture parameter, mostly modulus of elasticity.[Citation16,Citation17,Citation18]
The cooking part ends at cooking time CT when the cell wall bond concentration reaches the critical value cCT . This state can be also expressed by relative critical bond concentration ccr = cCT/c0 . Using this definition, CT is expressed from EquationEq. (1b) as:
The critical relative bond concentration represents the portion of the initial bond concentration c0 that was not destroyed in the cooking part. EquationEq. (3) is the equation connecting three important cooking parameters: relative critical bond concentration ccr, cooking time CT and kinetic coefficient k (relatively independent on potato density[Citation16]). The cooking time can be determined by the CPEM method (e.g., ) and k can be estimated from texture-time plots obtained during potato cooking[Citation13,Citation16]. shows that experimental CT values decrease approximately linearly with increasing potato density within all the tested varieties. The differences between the individual plots are given by the coefficients aCT and b in the linear regression:
Figure 2 Cooking time CT plotted against density. Coefficients in EquationEq. (4b) see . Nicola (R2 = 0.683), Saturna (R2 = 0.746), Agria1 (R2 = 0.916), data from growing year 2005 in Hejlová and Blahovec.[Citation23]
![Figure 2 Cooking time CT plotted against density. Coefficients in EquationEq. (4b) see Table 1. Nicola (R2 = 0.683), Saturna (R2 = 0.746), Agria1 (R2 = 0.916), data from growing year 2005 in Hejlová and Blahovec.[Citation23]](/cms/asset/803a5fff-64db-463e-9a7a-97984a87a764/ljfp_a_316479_o_f0002g.gif)
Table 1 Quantities measured and calculated (CAL) in the simple kinetic model
The CT sensitivity to density is expressed mainly by the parameter b that takes part also in the modified relation between CT and density ρ[Citation11]:
The relative critical bond concentrations were calculated from our experimental data () using EquationEq. (3). The kinetic coefficients were approximated from our previous experiments in which the elastic module was measured during potato cooking—see . Results of the calculations for three potato densities are also given in . They confirm that in the salad variety Nicola the intermolecular bonds are less sensitive to cell wall tensions caused by starch swelling: at higher tuber densities we obtained lower relative critical bond concentrations in Nicola than in the other two varieties. The role of the partial parameters aCT0 and b in critical bond concentration can be assessed by putting EquationEq. (4b) into EquationEq. (3):
The curvilinear character of the ccr dependence on density has a special polynomial character, in which the product kb plays an important role. The higher the value of kb, the steeper and more curved the ccr -ρ dependence. The examples of the calculated ccr are given in . The kb value in Nicola is less than half of the corresponding values obtained in the other two varieties. Parameter b termed as CT-sensitivity to starch represents a relatively stable variety characteristic[Citation11],[[Citation21]. On the other hand, the initial value of the ccr (in relation to the density ρ 0 ) is equal to the first term in EquationEq. (5b): exp(-kaCT0) that is determined by product of k and aCT0 .
The expression for CT-starch sensitivity b can be derived if k is independent on ρ as follows:
EquationEq. (6a) shows that x/c0 is proportional to the relative critical bond concentration with the proportionality constant kb described previously. Using EquationEq. (5a), EquationEq. (6a) can be rewritten into the following form:
The importance of the product kb for calculating the dependence of critical bond concentration on density dcCT /dρ is clearly expressed in EquationEq. (6b). Numerical results for x/c0 are given in . This quantity increases with increasing density and is much lower for Nicola than for the other two varieties. The formulas (6), (6a), and (6b) show that CT-starch sensitivity b increases with increasing slope of critical bond concentration dcCT /dρ. The higher the dcCT /dρ indicates the higher the CT-starch sensitivity b. The role of the initial bond concentration c0 in a raw tuber is given by formulas (6), (6a), and (6b), expressing changes caused by different cultivation & growing conditions. These conditions influence mainly the parameter aCT0 with a very limited influence on CT-starch sensitivity b[Citation21] so that the c0 can enter into EquationEqs. (5b) and Equation(6b) only by means of the member exp(-kaCT0 ). Cooking part theory for a more sophisticated reversible kinetic model is described in the Appendix. The reversible kinetic model is used for discussion of the experimental values in the Applications.
Breaking Part
The initial slope of the breaking part, SBP (see ), can be expressed as the initial rate of depression of the effective potato mass at the time of cooking in a CPEM test t = CT in the following manner:
The first term in EquationEq. (7) expresses change of the effective potato mass inside the sieve basket due to changes of the cell wall bond concentration close to its critical value cCT . It is determined by getting the potato fragments through the sieve meshes. It could be estimated on the base of its anticipated asymptotic values[Citation22] :
As one of suitable functions with the required asymptotic properties was analysed the generalized power function:
For constant K, k, and n Π(0, 1), the SBP behaves as a decreasing (exponential) function of the cooking time CT. The density derivative of EquationEq. (10) then can be expressed using EquationEq. (4a) as:
Notes to the Materials and CPEM Method
This part contains the most important information on previous experiments (materials, method, and main results in CPEM tests) and applications to the model. The potatoes were harvested in October 2005 and kept in cold store (at temperature 6°C and 95% humidity) until testing in November and December 2005 (Agria) and in January and February 2006 (Nicola and Saturna). Potato sample for a CPEM test was prepared from three or four tubers of approximately the same size and mean density (determined by weighing the tubers in air and in tap water by considering the water temperature). Rectangular slices 10 × 10 × 1.5 mm were cut mainly from the inner parenchyma, immediately before testing washed in a sieve (15 seconds) and dried (3–5 minutes) leaving on a textile tissue at room temperature. The sample, 100 g of washed and dried potato flakes, was then tested by the CPEM method.[Citation10]
Main CPEM Results
Both CPEM parameters are analysed as functions of potato tissue density. The dependence of cooking time CT on density is described by linear regression equations in individual potato groups creating the linear model of cooking stage [see (4a,b) and ]. Similarly, the dependence of SBP on density is characterised in linear model of disintegration stage ().
Figure 3 SBP plotted against density. Nicola (SBP = 0.00395ρ − 3.958, R2 = 0.435), Saturna (SBP = 0.01018ρ −10.575, R2 = 0.400), Agria1 (SBP = 0.01972 ρ −20.885, R2 = 0.443), data from growing year 2005 in Hejlová and Blahovec.[Citation23]
![Figure 3 SBP plotted against density. Nicola (SBP = 0.00395ρ − 3.958, R2 = 0.435), Saturna (SBP = 0.01018ρ −10.575, R2 = 0.400), Agria1 (SBP = 0.01972 ρ −20.885, R2 = 0.443), data from growing year 2005 in Hejlová and Blahovec.[Citation23]](/cms/asset/0aae55c3-2a66-49a3-bd7b-54a79b76bd07/ljfp_a_316479_o_f0003g.gif)
In the transformed regression equation (4b), ρ 0 is an approximation of a fictive tuber density without starch derived from Scheele´s empirical formulas measuring the relationship among potato tuber density, dry matter content, and starch content.[Citation19,Citation20] Hence, the intercept aCT0 represents the fictive cooking time required for sloughing of potato tissue with zero starch content and is associated with the cell wall properties. The regression coefficient b was termed as CT- sensitivity to starch content.[Citation11] It represents a new cooking parameter typical for variety and relatively independent on cultivation and growing conditions.[Citation11,Citation21]
Data Application and Discussion
The SBP estimation was written as product of two components in EquationEq. (10): The model component ke–kCT (1–n) is derived from the cooking part and its kinetic model and and a suitable n-value[Citation22]. The other component Kc0 represents an additional mass factor, which can be estimated from EquationEq. (10). Statistical analysis of our experimental data in [22] indicates optimum n-values in the range 0.2<n<0.4, for which the Kc0 values are close to 10 g, i.e. 10% of the initial mass of the sample in a CPEM test, and weakly correlated with tissue density (). Then the main source of SBP dependence on density () laid in the dependence of its model component SBP/Kc0 on density in individual tested groups (see and[Citation22]). Hence the parameter n and the factor Kc0 could express some inner disintegration relation associated with experiment conditions and relatively independent on the tested variety.[Citation22]
Table 2 Product Kc 0 calculated from EquationEq. (10) using experimental CT and SBP data and k-values from , parameter n = 0.2
Figure 4 Model component SBP/(Kc0) calculated using EquationEq. (10) and plotted against density. Nicola (SBP/(Kc0) = 0.00043ρ − 0.4328, R2 = 0.709), Saturna (SBP/(Kc0) = 0.00421ρ − 4.5324, R2 = 0.768), Agria1 (SBP/(Kc0) = 0.00336ρ − 3.5875, R2 = 0.963), CT data from and Hejlová and Blahovec.[Citation22]
![Figure 4 Model component SBP/(Kc0) calculated using EquationEq. (10) and plotted against density. Nicola (SBP/(Kc0) = 0.00043ρ − 0.4328, R2 = 0.709), Saturna (SBP/(Kc0) = 0.00421ρ − 4.5324, R2 = 0.768), Agria1 (SBP/(Kc0) = 0.00336ρ − 3.5875, R2 = 0.963), CT data from Fig. 2 and Hejlová and Blahovec.[Citation22]](/cms/asset/cb966bad-ef45-47d8-ad5c-8307a3673f76/ljfp_a_316479_o_f0004g.gif)
The parameter b in Eq. (4) is a relatively well-defined variety parameter nearly independent on cultivation conditions. The correlation matrix in , based on two years data for 8 different potato variants,[Citation23] confirms this clearly. It seems that this fact has roots in the loss of correlation between aCT0 and b, which is displayed in . It illustrates the real state well: there exists a relatively tight increasing relationship between the parameters b and aCT0 for all the tests on varieties Agria and Saturna, but the results on variety Nicola, even if different in two years, differ in both years significantly from results obtained for Agria and Saturna. It seems that cooking of the salad varieties is controlled by another mechanism than cooking of the other varieties. This difference represents the main source of low correlations in . The CT-starch sensitivity b is given by EquationEq. (6); it can be rewritten into the following form:
Figure 5 Parameter b plotted against parameter aCT0 from the EquationEq. (4b). Two years data of potato cooking tests[Citation23]. The plots for Agria and Saturna are approximated by the common linear equation: b = 0.0114 aCT0 − 0.0798, R2 = 0.875.
![Figure 5 Parameter b plotted against parameter aCT0 from the EquationEq. (4b). Two years data of potato cooking tests[Citation23]. The plots for Agria and Saturna are approximated by the common linear equation: b = 0.0114 aCT0 − 0.0798, R2 = 0.875.](/cms/asset/a516ebab-15e7-475b-b62b-9fede7415f3d/ljfp_a_316479_o_f0005g.gif)
Table 3 Correlation matrix of parameters describing dependence of sloughing on tuber density
The application of the more sophisticated partly reversed kinetics of potato cooking developed in the appendix is limited by a lack of suitable data describing the further parameter k2 - see EquationEqs. (A1) and Equation(A2). This model could be preferred for ccr approaching to cu/c0 , i.e. as is clear from EquationEq. (A2a) for ccr being a little bigger than ratio k2/k. The potatoes are well cooked at relative bond concentrations 0.1[Citation16] so that partly reversed kinetics can lead to important differences in comparison to the simple model at k2/k values higher than 0.1. Using of k2/k = 0.1 and density 1090 kg m−3 in EquationEq. (A3) leads to ccr values 0.196 for Agria and 0.171 for Nicola. At lower densities the obtained ccr values decrease consequently (see ). At these conditions the partly reversible model can give results significantly different from the simple one. Similarly as in EquationEq. (6) the CT-starch sensitivity b can be expressed in a partly reversible model in the following:
Figure 6 Derivation x/c0 calculated from Eqs. (14b) and (A3) plotted against ratio k2/k. Data for Agria and Nicola at constant density 1090 kg.m−3 from .
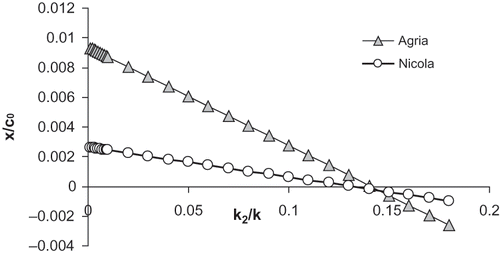
Our analysis based on the potato cooking kinetics showed that the variety susceptibility to sloughing is simply expressed via CT-starch sensitivity b that has roots in i/ dependence of critical bond concentration on density, ii/ initial critical bond concentration, and possibly iii) some details of more complicated kinetics of softening that are not yet fully clear.
CONCLUSIONS
Potato cooking is controlled by sensitivity of the intercellular bonds to the stresses produced by starch swelling. The simple kinetic model shows that cooking time is predetermined by two main parameters: kinetic coefficient of the tissue softening on one hand and the critical bond concentration in the intercellular space at the stage of starting disintegration on the other. Potatoes, in which the process of softening is slower, i.e. kinetic coefficient is of a lower value, and/or in which disintegration starts at lower bond concentrations in the intracellular spaces, the susceptibility to sloughing is lower than in the more susceptible varieties. Some role is played also by the initial bond concentration as a source of cultivation induced differences. The important role in potato sloughing can be also played by details of more complicated but still not fully known kinetics of softening. Knowing the main parameters of cooking, i.e. kinetic coefficient of the potato softening and the relative critical concentration of the intracellular bonds, the main parameters of disintegration can be estimated using the model proposed. There are some principal differences between most of potato varieties and the salad varieties consisting in much lower cooking sensitivity of salad varieties to the starch content. This difference is a source of reduction of the correlations among different cooking parameters that are usually observed with the other varieties than with the salad ones.
NOMENCLATURE
| aCT (min) | = |
constant in the linear regression relation between CT and density |
| aCT0 (min) | = |
hypothetical CT value for potato tissue with zero starch content |
| b (min.m3.kg−1) | = |
slope in the linear regression relation between CT and density, CT-sensitivity to density |
| c (m−3) | = |
cell wall bond concentration |
| ccr = cCT/c0 (−) | = |
critical relative cell wall bond concentration |
| cCT (m−3) | = |
critical cell wall bond concentration (corresponding to the cooking time CT) |
| cu (m−3) | = |
asymptotic cell wall bond concentration in partly reversible model |
| c0 (m−3) | = |
initial cell wall bond concentration in the raw tuber |
| CT (min) | = |
“cooking time”, time of cooking in CPEM test just before indication of the disintegration process |
| CV | = |
coefficient of variations |
| CPEM | = |
“cooked potato effective mass”, modified CPM test[Citation10],[Citation11] |
| CPM | = |
“cooked potato mass”, official test of potato sloughing[Citation9] |
| CPW | = |
“cooked potato weight”, term commonly used for CPM tests[Citation9] |
| k (min−1) | = |
kinetic coefficient, in partly reversible model k = k1 + k2 |
| K (g.m3) | = |
parameter of EquationEq. (9), defined by EquationEq. (8b) |
| k1 (min−1) | = |
main kinetic coefficient in the partly reversible model |
| k2 (min−1) | = |
kinetic coefficient in partly reversible model that is responsible for reversibility |
| MV | = |
mean value |
| m (g) | = |
effective sample mass |
| RC (m3.min−1) | = |
slope in the linear regression relation between SBP and density |
| R2 (−) | = |
coefficient of determination |
| SBP (g.min−1) | = |
“slope of the breaking part”, initial slope (absolute value of regression coefficient) of the breaking part in a CPEM test |
| SD | = |
standard deviation |
| x = dcCT /dρ (kg−1) | = |
density derivative of cell wall bond concentration at cooking time CT |
| ρ (kg.m−3) | = |
density |
| ρ 0 = 1005.5 (kg.m−3) | = |
hypothetical potato density corresponding to zero starch content (estimation based on the Von Scheele's empirical formulas) |
ACKNOWLEDGMENT
The research was supported in part by the Research Intention MSM 6046070905 (Czech Republic).
REFERENCES
- Warren , D.S. and Woodman , J.S. 1974 . The texture of cooked potatoes: A review . Journal of the Science of Food and Agriculture , 25 : 129 – 138 .
- Hoff , J.E. 1973 . Chemical and physical basis of texture in horticultural products . HortScience , 8 ( 2 ) : 108 – 110 .
- Jarvis , M.C. and Duncan , H.J. 1992 . The textural analysis of cooked potato. 2. Swelling pressure of starch during gelatinisation . Potato Research , 35 : 93 – 102 .
- Marle , J.T. van , van , Dijk, C. , Voragen , A.G.J. and Biekman , E.S.A. 1994 . Comparison of the cooking behaviour of the potato cultivars Nicola and Irene with respect to pectin breakdown and the transfer of ions . Potato Research , 37 : 183 – 195 .
- Jarvis , M.C. and Duncan , H.J. 1992a 1992 . The textural analysis of cooked potato. 1. Physical principles of the separate measurement of softness and dryness . Potato Research , 35 : 83 – 91 .
- Jarvis , M.C. and Duncan , H.J. 1992 . The textural analysis of cooked potato. 3. Simple methods for determining texture . Potato Research , 35 : 103 – 109 .
- Matsuura-Endo , C. , Ohara-Takada , A. , Yamauchi , H. , Mori , M. and Fujikawa , S. 2002 . Disintegration differences in cooked potatoes from three Japanese cultivars: Comparison of starch distribution within one tuber and morphology of tissue . Food Science and Technology Research , 8 ( 3 ) : 252 – 256 .
- Matsuura-Endo , C. , Ohara-Takada , A. , Yamauchi , H. , Mori , M. and Ishibashi , K. 2002 . Disintegration differences in cooked potatoes from three Japanese cultivars: Comparison of the properties of isolated starch, degree of cell separation with EDTA, and contents of calcium and galacturonic acid . Food Science and Technology Research , 8 ( 4 ) : 323 – 327 .
- Anonymous . 1977 . Methods of assessment for potatoes and potato products . EAPR. Pudoc , : 48 Wageningen
- Hejlová , A. , Blahovec , J. and Vacek , J. 2006 . Modified test for potato sloughing assessment . Journal of Food Engineering , 77 : 411 – 415 .
- Blahovec , J. and Hejlová , A. 2006 . Role of tuber density in potato sloughing . Journal of Texture Studies , 37 : 165 – 178 .
- Daniels , F. and Alberty , R.A. 1955 . Physical Chemistry , 650 New York : John Wiley & Sons, Inc .
- Huang , Y.T. and Bourne , M.C. 1983 . Kinetics of thermal softening of vegetable . Journal of Texture Studies , 14 : 1 – 9 .
- Pravisani , C.I. , Califano , A.N. and Calvelo , A. 1985 . Kinetics of starch gelatinization in Potato . Journal of Food Science , 50 : 657 – 660 .
- Ramesh , M.N. , Sathyanarayana , K. and Girish , A.B. 1996 . Kinetic modelling of vegetable cooking at 100 C . Journal of Texture Studies , 27 : 257 – 264 .
- Blahovec , J. and Esmir , A.A.A. 2001 . Precise study of cooked potato texture . Journal of Texture Studies , 32 : 165 – 184 .
- Solomon , W.K. and Jindal , V.K. 2003 . Comparison of mechanical tests for evaluating textural changes in potatoes during thermal softening . Journal of Texture Studies , 33 : 529 – 542 .
- Solomon , W.K. and Jindal , V.K. 2006 . Comparison of axial and radial compression tests for determining elasticity modulus of potatoes . International Journal of Food Properties , 9 : 855 – 862 .
- Scheele , C. von , Svensson , G. and Rasmussen , J. 1937 . Die Bestimmung des Stärkegehaltes und der Trockensubstanz der Kartoffel mit Hilfe des spezifischen Gewichts . Die Landwirtschaftlichen Versuchs-Stationen , 7 : 67 – 96 .
- Es , A. Van and Hartmans , K.J. 1987 . “ Structure and chemical composition of the potato ” . In Storage of potatoes , Edited by: Rastovski , A. Es , A. van . 39 – 47 . Pudoc Wageningen : Centre for Agricultural Publishing and Documentation .
- Hejlová , A. and Blahovec , J. 2007 . Role of cultivation conditions in potato sloughing as indicated by CPEM method . Plant, Soil and Environment , 53 ( 9 ) : 403 – 412 .
- Hejlová , A. and Blahovec , J. A Simple Model of the Breaking Part in CPEM Tests . Proceedings of the International Conference Trends in Agricultural Engineering TAE 2007 . Sept 12–14 2007 . Prague , , Czech Republic : Czech University of Life Sciences Prague . ISBN 978-80-213-1668-3
- Hejlová , A. and Blahovec , J. 2007 . Sloughing in potatoes induced by tuber density and affected by variety . Czech Journal of Food Science , 26 ( 1 ) : 41 – 50 .
APPENDIX: TWO MEMBER KINETIC THEORY OF COOKING PART
Instead of EquationEq. (1a) the kinetics of softening could be also described by the following two member linear kinetic equation:
This equation is an example of reversible kinetic equations[Citation12] describing a superposition of the original (k1 ) and the reversible (k2 ) processes. Even if in our case the process of cooking could be understood as rather irreversible one, the reverse part in EquationEq. (A1), i.e. k2(c0 – c) can be used to description of the stress induced process activated during cooking new and new intercellular bonds that did not originally participate in the product cohesion. The starch swelling stresses seem to play some important role in processes of this kind.
The solution of EquationEq. (A1) can be written easily in the following form:
The relative critical bond concentration ccr defined for simple kinetic equation in EquationEq. (3) is then expressed in the following form:
The parameter k = k1 + k2 plays in EquationEq. (A4) a similar role as the kinetic coefficient k in EquationEq. (5b). The relative critical bond concentration contains two parts in EquationEq. (A4): the first one independent on cooking parameters and proportional to kinetic parameter k2 and the second one that is sensitive to potato density and determined mainly by the product kb similarly as in EquationEq. (5b). When k2 = 0 then EquationEq. (A4) has the same form as EquationEq. (5b].