Abstract
Many volatiles in fresh tea leaf are in the forms bound with glycosides and they are released by action of ß–primeverosidase and ß-glucosidase during tea processing. Effect of ultra-violet B on release of volatiles in fresh tea leaves of two tea cultivars, “Maoxie” and “Anhui-9,” were investigated. Types of volatile and their concentration increased when ultra-violet B was applied at 3.0 μmol m−2 s−1 for 2 h and then decreased as the irradiation was extended to 6 h. The relation of expression of ß–primeverosidase and ß-glucosidase genes to the release of volatiles were also discussed.
INTRODUCTION
Aroma is an important determinant of sensory quality of tea and wine.[Citation1–3] There are more than 500 tea aromatic volatiles detected in tea, including monoterpene and aromatic alcohols which are considered to have great contribution to specific floral aroma of tea. Citation[4] The composition and concentration of tea volatiles depended on many factors including tea cultivars, fertilizing, harvesting season, pruning and processing, which resulted in diversities of tea flavor.Citation5–7
Most of the aromatic volatiles are in the form of glycosides in fresh tea leaves such as ß-primeverosides,Citation[8–Citation11] β-D-glucopyranside,Citation[12] vicianoside,Citation[13] and acuminoside,Citation[10,Citation14] which are called, bound aromatic precursors. The bound glycosides precursors varied with tea cultivars, climate and maturity of tea shoots.Citation[15] Aromatic volatiles are released from the precursors by action of endogenous glycosidases during tea manufacture.Citation[16] Glycosidases such as ß–primeverosidase (TBP) and ß-glucosidase (TBG) were confirmed to be two important enzymes hydrolyzing the aromatic precursors to liberate volatiles,Citation[16,Citation17] and the genes encoding the two enzymes were isolated from tea.Citation[12,Citation18] The glycosidase activities reached high level during withering process but decreased after rolling and hydrolysis of glycosides mainly occurred during the stage of rolling.Citation[16] During tea manufacture, many tea volatiles were formed and the up-regulated transcripts for encoding various proteins homologous to stress response proteins occurred in the gene expression profiles.Citation[19] During oolong tea (a semi-fermented tea) manufacturing process, fresh tea leaves are solarized in the sunshine for tens of minute to promote the release of volatiles.Citation[16] A special kind of oolong tea “Oriental Beauty” was prepared using tea leaves infested by an insect “green leafhopper” and it had a unique aroma similar to ripe fruit and honey.Citation[19,Citation20,Citation21] This method is a process that utilizes plant defense response to elevate the production of volatile compounds.Citation[19] Although there have been reports published on the constituents of tea aroma, the formation of tea flavor still conceals many secrets.Citation[22]
Ultraviolet B (UV-B), a radiation with wave length 280–320 nm, was confirmed to have important effects on expression of many genes related to plant secondary metabolites, including carotenoid,Citation23–25 glycyrrhizin,Citation[26] flavonoids,Citation[27] and tea catechins,Citation[28] but reports about effect of UV-B on formation of tea volatiles are not readily available. This study was set to investigate the effects of UV-B on release of tea volatiles and expression of TBG and TBP genes. It will be propitious to understand the mechanism of flavor elevation by solarization treatment during oolong tea processing and to develop new methods to improve tea flavor.
MATERIALS AND METHODS
Plant Materials
Tea shoots (5 kg) with two leaves and a bud were harvested from bushes of Camellia sinensis cultivars “Maoxie” and “Anhui-9” grown on experimental farm of Zhejiang University Tea Research Institute (Hangzhou, China). To avoid the influence of sunlight, the tea shoots were harvested at 6 am.
UV-B Irradiation
Tea shoots (500 g each sample) were irradiated by UV-B with fluence 3.0 μmol m−2 s−1 from four BLE-IT158 lamps (Spectronics Corporation Westbury, NY, USA) for 2 h and 6 h respectively. Samples (200mg each) for RNA extraction and gene expression test were taken from the irradiated tea shoots and stored at −70°C. The remaining leaves were fixed by steam for 30 s and then dried at 80°C overnight. The dried leaves were ground to particle size 0.45 mm for extracting volatiles. The tea shoots for control were kept under dark condition and sampled at 0, 2, and 6 h, respectively. The sampled shoots were treated as discussed above.
Determination of Volatiles
Volatiles were prepared using the ground samples (15 g) by simultaneous distillation extraction (SDE) method as a previous paperCitation[29] and determined by gas chromatography-mass spectrometry (GC-MS). The GC-MS was carried out on Aglient model HP6890/5973 GC-MS (Agilent Technologies, Palo Alto, CA, USA) equipped with a mass selective detector (Model HP 5973, Hewlett Packard, CA, USA) under following conditions: sample injection volume 2 μL, HP-INNOWax capillary column (30 m × 0.32 mm × 0.5 μm i.d.), column temperature programmed to hold at 50°C for 5 min, then increase to 230°C at 3°C min−1, finally hold at 230°C for 10 min, carrier gas (helium, 99.999%) flow rate 1.0 ml min−1, injector temperature 230°C, ion source temperature 230°C, EI ionization style. The MS was scanned at 70 eV over 10–400 a.m.u. The volatiles were identified by comparison with the spectral library of the GC-MS. The level of the detected volatile was expressed as the ratio of its peak area to the area of internal standard (ethyl caprate, 0.2 μg μl−1).
Expression Study on TBP and TBG Genes
Total RNA was extracted using TRIzol reagent (Invitrogen Life Technologies, Carlsbad, CA, USA) as manufacturer's manual and its concentration was determined according to the absorbance at 260 nm and 280 nm on GeneQuant Pro DNR/DNA calculator (Biochrom Ltd, Cambridge, UK). First strand cDNA was synthesized from 1 μg of total RNA and 0.5 μg of oligo (dT) primer using D6110A PrimeScriptTM First Strand cDNA Synthesis Kit (TaKaRa Biotechnology Co., Ltd., Dalian City, China) according to the product manual. The cDNA was used to test the expression profiles of TBG and TBP genes by polymerase chain reaction (PCR) method.Citation[18]
Primers for amplifying TBG and TBP genes were designed by software Lasergene Primer Select (DNA Star, Madison, WI, USA) based on the corresponding sequences published on the GenBank (). Total volume of PCR system was 25 μL, including 1μL cDNA, 2.5μL 10-time PCR buffer (50 mmol L−1 KCl, 10 mmol L−1 Tris–HCl, 0.1g L−1 gelatin, pH 8.3), 2 μL MgCl2 (25 mmol L−1), 0.5μL dNTPs (200 μmol L−1), 0.5μL primer (0.5 μmol L−1), 0.4μL of Taq enzyme (2 units, Bio Basic Inc., East Markham, Ontario, Canada), and 18.1μL H2O. The PCR was carried out on a PTC 200 Thermocyler (MJ Research, Waltham, MA, USA) as follows: 1 cycle of 94°C for 5 min, followed by 25 cycles of 94°C for 1 min, 54°C for 1 min and 72°C for 1 min, and finally 1 cycle of 72°C for 5 min for strand extension. The PCR products were fractionated on agarose gel (15 g L−1) and stained with ethidium bromide. To confirm the identity of the gene amplified fragments, the PCR products were cloned into pGEM®-T Easy Vector System I (Promega Corporation, Madison, WI, USA) and sequenced according to dideoxynucleotide chain termination method on an ABI 3130 Prism Sequencer (Applied Biosystems, Foster City, CA,USA). The experiments in the present study were carried out in duplicate.
Table 1 Primers for amplifying TBG and TBP genes. a
RESULTS AND DISCUSSION
Effect of UV-B Irradiation on Release of Volatiles
Composition and concentration of volatiles varied with tea cultivars. The number of volatiles detected in freshly picked shoots (0 h) of cultivars “Maoxie” and “Anhui-9” were 14 and 34 respectively (–). The top five abundant volatiles in the freshly picked shoots were 2-hexenal, cis-linalool, α-Methyl-α-(4-methyl-3-pentenyl)-oxiranemethanol and trans-Linalool in cultivar “Maoxie” (), but cis-linalool, dehydrolinalool, 2,4-heptadienal, nerolidol, and β-ionone in cultivar “Anhui-9” (). When the tea shoots were irradiated by UV-B for 2 h, the number of volatiles increased from 14 to 33 in “Maoxie” and from 34 to 37 in “Anhui-9” respectively. The volatiles in shoots kept in dark condition for 2 h were 18 in “Maoxie” and 34 in “Anhui-9.” However, when the UV-B irradiation was extended to 6 h, the number of volatiles increased to 46 in “Maoxie,” but decreased to 34 in “Anhui-9.” The abundant volatiles in the freshly picked tea shoots decreased greatly after UV-B irradiation and storage in the dark. After 6 h of UV-B irradiation, the top five abundant volatiles were 2-hexenal, α-farnesene, indolizine, nerolidol, and cis-linalool in cultivar “Maoxie,” but nerolidol, cis-linalool, dehydrolinalool, diisobutyl phthalate, and β-ionone in cultivar “Anhui-9.” Total concentration of volatiles was the highest when UV-B was applied for 2 h. When the UV-B irradiation was extended to 6 h, total concentration of volatiles decreased and it showed a similar trend in the two cultivars (–). This suggests that appropriate dosage of UV-B stimulated the release of volatiles from the bounded volatile precursors but excessive UV-B might promote the degradation of volatiles. During the process “withering-rolling-fermentation-drying” of black tea manufacturing, the enzyme activities of TGP and TBG remained at a high level during withering but decreased drastically after rolling.Citation[16] The decrease in total volatile concentration after extended UV-B irradiation might be related to the decrease in the activities of TGP and TBG.
Table 2 Effect of UV-B on release of volatiles in tea cultivar Maoxie.a
Table 3 Effect of UV-B on release of volatiles in tea cultivar Anhui-9.a
Aromatic volatiles in made tea were released from their bound precursors such as glycosides in fresh tea leaves during tea processing.Citation[8,Citation9,Citation11,Citation30] The present study showed that the concentrations of volatiles which were abundant in freshly picked tea shoots decreased under both dark and UV-B irradiation conditions though the number of volatiles increased (–). This means that degradation of the initial volatiles and release of new volatiles took place simultaneously. Total concentration of volatiles depended on balance between the rates of degradation and release. At early stage of UV-B treatment, the release of volatiles was more quickly than the degradation because of abundance of bound volatile precursors. At late stage, however, it changed the other way round because of depletion of volatile precursors. This explains why the abundance of volatiles changed after UV-B treatment and total concentration of volatiles at 2 h UV-B treatment was higher than at 6 h.
Expression Patterns of TBG and TBP Genes
The sequencing results showed that the PCR fragments amplified from primers based on TBG and TBP genes were 232 bp and 265 bp, respectively. The sequence alignment showed that the obtained TBG sequence had 98% identity with the published ß-glucosidase gene (GenBank accession No. AF537127) and TBP sequence had 99% identity with the published ß-primeverosidase gene (GenBank accession No. AB088027) in tea plant. This suggests that the PCR fragments were from ß-glucosidase gene and ß-primeverosidase gene respectively.
Gene expression profiles showed that expressions of TBG and TBP genes were weak in freshly picked tea shoots of cultivar “Maoxie” (A). Expressions of the two genes were enhanced when tea shoots was irradiated by UV-B for 2 h and then weakened as UV-B irradiation was extended to 6 h (A). The expression of TBG was also accelerated in darkness for 2 h though it was weaker than the corresponding UV-B treatment. TBP gene showed a similar trend in both the cultivars, but TBG exhibited strong expression in the freshly picked shoot of cultivar “Anhui-9” (B). The strongest expression of TBG in “Anhui-9” was observed in 2h UV-B treatment and 6 h dark control (B). The study showed that appropriate dosage of UV-B radiation accelerated the expression of genes TBG and TBP though initial expression levels of the two genes differed between the tea cultivars. Excessive UV-B radiation suppressed the expression of the two genes.
Figure 1 Effect of UV-B on expression of TBG and TBP genes in tea shoots. A: Cultivar Maoxie; B: cultivar Anhui-9; TBG: β-glucosidase gene; and TBP: β-primeverosidase gene. 1: Freshly picked tea shoot (0 h); 2: Under dark condition for 2 h; 3: UV-B irradiation for 2 h; 4: Under dark condition for 6 h; 5: UV-B irradiation for 6 h.
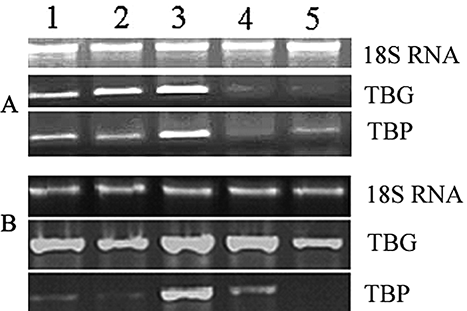
The bound precursors of the alcoholic aromatic volatiles in tea leaf were glycosides and most of them were β-primeverosides.Citation[8,Citation9,Citation10,Citation12,Citation30] The aroma precursors of linalool oxides and (z)-3-hexenol were exceptionally isolated as β-D-glucopyranosides.Citation[10] Hydrolysis of the glycosides mainly occurred during the stage of rolling when the glycosidase activities remained at a high level.Citation[16] TBG and TBP are genes encoding ß-glucosidase and ß-primeverosidase. The enhancement in expression of TBG and TBP genes by 2 h UV-B irradiation suggested that the biosynthesis of ß-glucosidase and ß-primeverosidase were promoted, which in turn accelerated the hydrolysis of the bound volatile precursors and resulted in the accumulation of free volatiles. The oolong tea “Oriental Beauty” with a unique aroma was prepared using tea leaves infested by insect “green leafhopper.”Citation[19] Traditionally, freshly picked tea leaf was solarized for ten minutes to increase tea aroma level. These processes elevate the production of volatile compounds through plant defense response.Citation[19] Recent studies have demonstrated that in plants, the changes induced as a consequence of UV damage was similar to those elicited in response to insect damage and the common signals, such as jasmonic acid, play important roles in the mechanisms by which plants respond to UV-B.Citation[31] The expression elevation of TBG and TBP genes might be the response of tea leaf to UV-B. The released volatiles may be also the signal compounds in the pathway of response of tea shoot to UV-B damage. It is known that UV-B is a part of solar radiation. That may be the reason why solarization of tea leaf was carried out in conventional oolong tea processing.
The number of volatiles in freshly picked leaf of cultivar “Anhui-9” was much more than that in cultivar “Maoxie,” but total concentration of volatiles was just the opposite (–). The expression level of TBG gene in freshly picked leaf of “Anhui-9” was higher than that of “Maoxie” (), suggesting that the basic level of ß-glucosidase in fresh leaf of “Anhui-9” was higher. This implies that hydrolysis of glucoside volatile precursors was more active in freshly picked leaf of “Anhui-9” than that of “Maoxie,” leading to more volatiles to be released by UV-B from glucoside bound precursors in freshly picked leaf of “Anhui-9.” However, β-primeverosides were major aroma precursorsdoubtCitation8–10,Citation30] and ß-primeverosidase was a key enzyme involved in tea aroma formation.Citation[12,Citation16] When the expression of TBP gene was induced by irradiation of UV-B for 2 h (), the number and total concentration of volatiles in cultivar “Maoxie” were higher than cultivar “Anhui-9” (–). This suggested that cultivar “Maoxie” had higher level of volatile precursors in primeveroside bound form than cultivar “Anhui-9.” In china, cultivar “Maoxie” is used for processing oolong tea, a kind of semi-fermented tea with strong aroma, while cultivar “Anhui-9” is for processing green tea. This suggested that the number and concentration of volatiles depends on the level of bound volatile precursors and activity of hydrolases. The level of volatile precursors, especially the primeveroside bound ones, is more important factor influencing the total concentration of aroma volatiles in made tea. This mechanism of volatile release was enzymatic one. It is different from the volatile formation in coffee processing in which thermal reactions took place.Citation[32]
CONCLUSION
The bound volatile precursors in fresh tea leaf were hydrolyzed and then released by action of ß-primeverosidase and ß-glucosidase during tea processing. Appropriate dosage of ultra-violet B (UV-B) could stimulate the expression of ß-primeverosidase and ß-glucosidase genes, resulting in the release of volatiles in tea leaf. The effects of UV-B on release of volatiles depended on UV-B dosage and tea cultivar. The number and total concentration of released volatiles increased when the fresh tea leaves were irradiated by UV-B with fluence 3.0 μmol m−2 s−1 for 2 h and then decreased as the UV-B irradiation was extended to 6 h. The volatiles induced by UV-B were more in the cultivar with weak expression of TBG gene than the cultivar with strong expression of TBG gene. The study could be interesting for developing new method to improve aromatic flavor of tea.
ACKNOWLEDGMENT
The authors gratefully acknowledge the Ministry of Agriculture of China for financial grant (Agricultural Commonweal Project No. 3–35).
Related Research Data
REFERENCES
- Liang , Y.R. , Zhang , L.Y. and Lu , J.L. 2005 . A study on chemical estimation of pu-erh tea quality . Journal of the Science of Food and Agriculture , 85 : 381 – 390 .
- Liang , Y.R. , Lu , J.L. , Zhang , L.Y. , Wu , S. and Wu , Y. 2003 . Estimation of black tea quality by analysis of chemical composition and colour difference of tea infusions . Food Chemistry , 80 : 283 – 290 .
- Elmaci , Y. , Yildirim , H.K. , Yuecel , U. , Ova , G. and Altug , T. 2007 . Descriptive profiling of flavor attributes of white wines from different grape varieties . International Journal of Food Properties , 10 : 651 – 659 .
- Wan , X.C. 2003 . Tea Biochemistry , 3rd , 40 – 43 . Beijing, , China : China Agricultural Press .
- Kumazawa , K. , Kubota , K. and Masuda , H. 2005 . Influence of manufacturing conditions and crop season on the formation of 4-mercapto-4-methyl-2-pentanone in Japanese green tea (sen-cha) . Journal of Agricultural and Food Chemistry , 53 : 5390 – 5396 .
- Ramarethinam , S. , Latha , K. and Rajalakshmi , N. 2002 . Use of a fungal lipase for enhancement of aroma in black tea . Food Science and Technology Research , 8 : 328 – 332 .
- Ravichandran , R. 2004 . The impact of pruning and time from pruning on quality and aroma constituents of black tea . Food Chemistry , 84 : 7 – 11 .
- Guo , W. , Sakata , K. , Watanabe , N. , Nakajima , R. , Yagi , A. , Ina , K. and Luo , S. 1993 . Geranyl 6-O-ß-D-xylopyranosyl-ß-D-glucopyranoside isolated as an aroma precursor from tea leaves for oolong tea . Phytochemistry , 33 : 1373 – 1375 .
- Guo , W. , Hosoi , R. , Sakata , K. , Watanabe , N. , Yagi , A. , Ina , K. and Luo , S. 1994 . (S)-Linalyl, 2-phenylethyl, and benzyl disaccharide glycosides isolated as aroma precursors from oolong tea leaves . Bioscience Biotechnology and Biochemistry , 58 : 1532 – 1534 .
- Moon , J. H. , Watanabe , N. , Ijima , Y. , Yagi , A. , Ina , K. and Sakata , K. 1996 . cis- and trans-Linalool 3,7-oxides and methyl salicylate glycosides and (z)-3-hexenyl ß-d-glucopyranoside as aroma precursors from tea leaves for oolong tea . Bioscience Biotechnology and Biochemistry , 60 : 1815 – 1819 .
- Nishikitani , M. , Wang , D. , Kubota , K. , Kobayashi , A. and Sugawara , F. 1999 . (z)-3- Hexenyl and trans-linalool 3,7-oxide β-primeverosides isolated as aroma precursors from leaves of a green tea cultivar . Bioscience Biotechnology and Biochemistry , 63 : 1631 – 1633 .
- Ma , S.J. , Watanabe , N. , Yagi , A. and Sakata , K. 2001 . The (3R,9R)-3-hydroxy-ß-ionol disaccharide glycoside is an aroma precursor in tea leaves . Phytochemistry , 56 : 819 – 825 .
- Nishikitani , M. , Kikue , K. , Kobayashi , A. and Sugawara , F. 1996 . Geranyl 6-O-R-L- arabinopyranosyl-α-D-glucopyranoside isolated as an aroma precursor from leaves of a green tea cultivar . Bioscience Biotechnology and Biochemistry , 60 : 929 – 931 .
- Mizutani , M. , Nakanishi , H. , Ema , J.I. , Ma , S.J. , Noguchi , E. , Inohara-Ochiai , M. , Fukuchi-Mizutani , M. , Nakao , M. and Sakata , K. 2002 . Cloning of ß-primeverosidase from tea leaves, a key enzyme in tea aroma formation . Plant Physiology , 130 : 2164 – 2176 .
- Saikia , P. and Mahanta , P.K. 2002 . Specific fluctuations in the composition of lipoxygenase- and glycosidase-generated flavors in some cultivated teas of Assam . Journal of Agricultural and Food Chemistry , 50 : 7691 – 7699 .
- Wang , D. M. , Kurasawa , E. , Yamaguchi , Y. , Kubota , K. and Kobayashi , A. 2001 . Analysis of glycosidically bound aroma precursors in tea leaves. 2. Changes in glycoside contents and glycosidase activities in tea leaves during the black tea manufacturing process . Journal of Agricultural and Food Chemistry , 49 : 1900 – 1903 .
- Ma , S.J. , Mizutani , M. , Hiratake , J. , Hayashi , K. , Yagi , K. , Watanabe , N. and Sakata , K. 2001 . Substrate specificity of beta-primeverosidase, a key enzyme in aroma formation during oolong tea and black tea manufacturing . Bioscience Biotechnology and Biochemistry , 65 : 2719 – 2729 .
- Zhao , L. , Chen , L. , Wang , X.C. and Yao , M.Z. Quantitative detection of β-glucosidase and β-primeverosidase gene expressions in different leaves of tea plant (Camellia sinensis) by Real-time PCR analysis . Journal of Tea Science 2006 , 26 11 – 16 . (in Chinese)
- Cho , J.Y. , Mizutani , M. , Shimizu , B.I. , Kinoshita , T. , Ogura , M. , Tokoro , K. , Lin , M.L. and Sakata , K. 2007 . Chemical profiling and gene expression profiling during the manufacturing process of Taiwan oolong tea “oriental beauty.” . Bioscience Biotechnology and Biochemistry , 71 : 1476 – 1486 .
- Ahmad , S. , Vashney , A.K. and Srivasta , P.K. 2005 . Quality attributes of fruit bar made from papaya and tomato by incorporating hydrocolloids . International Journal of Food Properties , 8 : 89 – 99 .
- Subramanian , R. , Hebbar , H.U. and Rastogi , N.K. 2005 . Processing of honey: A review . International Journal of Food Properties , 8 : 89 – 99 .
- Naef , R. , Jaquier , A. , Velluz , A. and Maurer , B. 2006 . New constituents related to 3-methyl-2,4-nonanedione identified in green tea . Journal of Agricultural and Food Chemistry , 54 : 9201 – 9205 .
- Steel , C.C. and Keller , M. 2000 . Influence of UV-B irradiation on the carotenoid content of Vitis vinifera tissues . Biochemical Society Transactions , 28 : 883 – 885 .
- Caldwell , C.R. and Britz , S.J. 2006 . Effect of supplemental ultraviolet radiation on the carotenoid and chlorophyll composition of green house-grown leaf lettuce (Lactuca sativa L.) cultivars . Journal of Food Composition and Analysis , 19 : 637 – 644 .
- Ebrahim , M.K.H. 2004 . Stress/tolerance responses of two cotton cultivars exposed to ultraviolet-A (366nm): photosynthetic performance and some chemical constituents . Agrochimica , 48 : 177 – 191 .
- Afreen , F. , Zobayed , S.M.A. and Kozai , T. 2005 . Spectral quality and UV-B stress stimulate glycyrrhizin concentration of Glycyrrhiza uralensis in hydroponic and pot system . Plant Physiology and Biochemistry , 43 : 1074 – 1081 .
- Ruhland , C.T. , Xiong , F.S. , Clark , W.D. and Day , T.A. 2005 . The influence of ultraviolet-B radiation on growth, hydroxycinnamic acids and flavonoids of Deschampsia antarctica during springtime ozone depletion in Antarctica . Photochemistry and Photobiology , 81 : 1086 – 1093 .
- Mamati , G.E. , Liang , Y.R. and Lu , J.L. 2006 . Expression of basic genes involved in tea polyphenol synthesis in relation to accumulation of catechins and total tea polyphenols . Journal of the Science of Food and Agriculture , 86 : 459 – 464 .
- Liang , Y.R. , Wu , Y. , Lu , J.L. and Zhang , L.Y. 2007 . Application of chemical composition and infusion colour difference analysis to quality estimation of jasmine-scented tea . International Journal of Food Science and Technology , 42 : 459 – 468 .
- Moon , J.H. , Watanabe , N. , Sakata , K. , Yagi , A. , Ina , K. and Luo , S. 1994 . trans- and cis-Linalool 3,6-oxides 6-O-ß-D-xylopyranosyl-ß-D-glucopyranosides isolated as aroma precursors from leaves for oolong tea . Bioscience Biotechnology and Biochemistry , 58 : 1742 – 1744 .
- Foggo , A. , Higgins , S. , Wargent , J.J. and Coleman , R.A. 2007 . Tri-trophic consequences of UV-B exposure: plants, herbivores and parasitoids . Oecologia , 154 : 505 – 512 .
- Strezov , V. and Evans , T.J. 2005 . Thermal Analysis of the Reactions and kinetics of green coffee during roasting . International Journal of Food Properties , 8 : 101 – 111 .