Abstract
The effects of space-travel-relevant radiation doses (gamma radiation) on structure, function, and antioxidant properties of select wheat cultivars (Triticum aestivum L.) were investigated. Following radiation treatment, proteins were analyzed using SDS-PAGE, mixograph, and atomic force microscopy methods. Starch granule damage and pasting curve functionality were evaluated; lipid oxidation was determined using a thiobarbituric acid reactive substances method; and antioxidant capacity was measured using 2,2-diphenyl-1-picrylhydrazyl analyses. Increasing radiation exposure resulted in significant differences between wheat cultivars. Therefore, low-dose gamma-radiation affects wheat, and differences exist between cultivars, so careful consideration is needed when selecting wheat for use in elevated radiation conditions. The Apogee cultivar is likely a better NASA candidate crop than Perigee; however, Yecora Rojo, Parshall, and/or Yavaros 79 cultivars may be more stable to radiation and provide better food functionality traits. Ultimately, wheat should be protected from radiation exposure during space travel, if possible, to maintain quality.
INTRODUCTION
Food quality and stability are extremely important for an extended mission in outer space. During previous missions to the moon and the space station, prepackaged stabilized food has been consumed. These missions have relied upon a limited amount of processed food along with re-supply options to sustain the crew. For a mission to Mars or beyond, a large self-sustaining food system must be developed in tandem with extending the shelf-life of prepackaged foods. For a mission to Mars, a combination of prepackaged foods, bulk ingredients, and crops will be used to provide a well-balanced diet.[Citation1,Citation2] Bulk food items may be sent to the surface ahead of or along with the crew to establish a short-term food system while housing is being constructed and crops are being grown. Once a base is established, a Martian-grown vegan diet will be the main source of food due to the distance and high costs of a continuous re-supply alternative and the mission objective of maintaining an enclosed bioregenerative habitat. Prepackaged foods will then be used for the return trip to Earth.[Citation1]
With missions planned beyond low Earth orbit and away from the protection of the Earth's magnetic field, the threat of radiation to the crew and food system is greatly increased. The proposed mission to Mars includes 6–8 months in transit each way as well as 600 days on the surface of Mars.[Citation1] Such a mission is estimated to encounter radiation levels on the order of 1–3 sieverts[Citation3] (1 seivert or Sv, the unit for measuring radiation exposure, is equivalent to 1 gray or Gy gamma radiation). The unpredictability of cosmic ray bursts also has the potential of increasing radiation exposure by up to 50 Sv per event. At these radiation levels, there are few reports of radiation effects on food. The contribution of expected deep space radiation and unpredictable cosmic ray bursts causes the total expected radiation amount, or mission-relevant dose, to vary considerably from 1 to perhaps several hundred Gy. Single radiation doses (500–5,000 Gy) have been used to prevent mold and fungal growth on wheat, but the quality of the wheat was not examined.[Citation4,Citation5] The goal of the NASA foods group is to ensure food safety, quality, and nutritional value throughout every mission. Therefore, the effect of mission-relevant radiation doses on foods is important.
Wheat has been designated as an important advanced life support candidate crop for a NASA mission to Mars due to its ability to be utilized in various recipes as well as its nutritional characteristics.[Citation6] As part of integrating the food system with the other life support systems, Apogee and Perigee super-dwarf wheat cultivars were developed to provide the maximum wheat berry production with a minimum amount of waste. These cultivars grow to roughly a quarter of the height of their terrestrial counterparts while producing the same amount of food product.[Citation7,Citation8] While growth conditions have been characterized, further analysis of these cultivars must be conducted to characterize the protein, lipid, and starch functionalities, antioxidant capacities, and food uses.[Citation9,Citation10] Furthermore, studies must be conducted after exposure to radiation to determine how these characteristics could be affected by radiation that will be encountered in space. Because wheat could be shipped as a bulk ingredient, effects of radiation on a variety of wheats used for food production are also important to characterize. The objective of this study was to determine the effects of gamma radiation exposure, including space-relevant low doses, on quality and functional properties of select wheat cultivars.
MATERIALS AND METHODS
Wheat Samples
Wheat samples (Triticum aestivum L.) of Apogee, Perigee, Yecora Rojo, Parshall, and Yavaros 79 cultivars were used. Apogee and Perigee are super-dwarf cultivars and were originally obtained from Utah State University.[Citation7,Citation8] Berries from these cultivars were grown in a greenhouse at Purdue University in three lots. Parshall, a hard red spring wheat, was obtained from Foundation Seed Company (Fargo, ND, USA). Yecora Rojo, a hard red spring wheat, and Yavaros 79, a durum wheat, were purchased from Foundation Seed Company (Davis, CA, USA). Triplicate samples of each of these “terrestrial” wheats were purchased. Wheat samples were milled twice using a Nutrimill grinder (Nutrimill Home Mill, Salt Lake City, UT, USA) and sifted through a sieve with a screen opening diameter of 1 mm. Flour samples were then adjusted to 14% moisture for all experiments.
Reagents
The following supplies and chemicals were obtained from the indicated sources: acetic acid, ethanol, methanol, petroleum ether, and sulfuric acid from Mallinckrodt (Paris, KY, USA); Coomassie brilliant blue R-250, 2-mercaptoethanol, sodium acetate, and sodium dodecasulfate from Bio-Rad (Hercules, CA, USA); 2,2-diphenyl-1-picrylhydrazyl, gliadin, sodium borate, sodium chloride, thiobarbituric acid, and trichloroacetic acid from Sigma-Aldrich (St. Louis, MO, USA).
Radiation Treatment
Radiation treatments were performed using a Cobalt 60 (Co60) Gamma Cell 220 (Nordion International, Ottawa, Ontario, Canada) at Purdue University with a dose rate of approximately 10 Gy per min. Dose was calculated from a known decay rate of Co60 that varied during the course of the experiments, but averaged approximately 0.20 Gy per sec. Radiation doses of 1, 3, 10, and 100 Gy were used on wheat berries. An additional dose of 10 kGy was used on wheat berries prior to SDS-PAGE analysis. Approximately 100 grams of wheat berries from each cultivar were placed in Ziploc® bags and irradiated. Additionally, purified wheat gliadin was exposed to 10 kGy for an atomic force microscopy (AFM) study. All samples were analyzed within 2 days of radiation treatment unless otherwise noted.
Protein Analyses
Proteins (albumins, globulins, gliadins, glutenins) were isolated from ground wheat flour using a method adapted from Hamaker and others.[Citation11] Proteins were fractionated into three tubes based on solubility differences: albumins and globulins were isolated in 0.9M NaCl, gliadins were isolated in 70% ethanol, and glutenins were isolated from a borate solution containing SDS detergent. SDS-PAGE was performed to separate proteins on the basis of size and enabled the visualization of the gliadin and glutenin fragments on the gels. Once isolated, the proteins were run on a 10–20% criterion gradient gel at 200 V for 1 h. A broad range molecular weight marker (7.1‐209 kDa) was run along with the proteins for size comparison (Bio-Rad, Hercules, CA, USA). Gels were stained and destained to observe protein fragments.[Citation11]
Mixograph studies (National Manufacturing, Lincoln, NE, USA) were used to indirectly determine gluten strength by measuring the amount of force necessary to mix dough as water was added. Thirty grams of flour were adjusted to 14% moisture and added to the mixograph mixing bowl. Twenty grams of water were initially added to a flour sample, and additions of water were made as needed until comparable graphs could be produced as described in American Association of Cereal Chemists (AACC) Method 54–40.[Citation12]
For AFM studies, purified gliadin samples (Sigma-Aldrich, St. Louis, MO, USA) were further purified following the method published by Makarenko and others.[Citation13] One gram of gliadin was suspended in an Erlenmeyer flask with 25 mL of 70% ethanol and stirred for 3 h. The proteins in the supernatant fraction obtained after centrifugation of the extract at 6000 × g for 15 min[Citation13] were freeze-dried using a Freezemobile 5SL (VirTis, Inc., Gardiner, NY, USA) and the purified gliadin was resuspended in 0.1 N acetic acid. A 1 mL aliquot of the gliadin suspension was dispensed into each of six Eppendorf tubes. Three of these Eppendorf tubes were then exposed to 10 kGy of radiation, and the other three were used as controls. A 10−4 dilution was then made, and 10 μL of each sample were dispensed onto a diamond surface and allowed to dry in a desiccator over desiccant overnight at room temperature. Samples were then imaged using an Asylum Research MFP-3D-Bio atomic force microscope (Santa Barbara, CA, USA) and AC160 probes (Olympus Corp., Tokyo, Japan). These silicon probes have rectangular cantilevers with an average spring constant of 42 N/m and tetrahedral shaped tips with a tip radius of less than 10 nm. The scan rate was 0.8 Hz, while the scan line and scan points were both set to 256. Height images based on piezoelectric voltage were modified using a zero-order flattening function and 3D rendered. Images of the irradiated and non-irradiated samples were collected in triplicate, and each sample was also examined in triplicate.
Lipid Analysis
Wheat oil was isolated using the Soxhlet extraction method described in AACC Method 30–25.[Citation12] Approximately thirty grams of wheat flour were placed into extraction thimbles and placed under a continuous drip of petroleum ether for 16 h. After that time, samples were placed under a nitrogen evaporator to remove excess petroleum ether. Oil was then transferred to small test tubes and held at 65°C for one week before being tested using the 2-thiobarbituric acid reactive substances (TBARS) test.[Citation14] Ten mL of TBARS reagent was vortexed for 30 sec with 2 μL of oil and heated for one hour at 95°C. The sample was then centrifuged for 10 min at 10,000 × g and the absorbance was read at 532 nm (Beckman Coulter, DU 800, Fullerton, CA, USA).
Starch Analyses
The pasting curve procedure mixed 1 g of starch isolated from wheat flour and 10 g of water into the vessel of a Rapid Viscoanalyzer (Newport Scientific, Warriewood, Australia) with a rotating spindle at 50°C for 1 min followed by 95°C for 5 min.[Citation15] Starch damage analysis was performed using a starch damage kit (Megazyme Starch Damage Kit, Wicklow, Ireland) following AACC Method 76–31.[Citation12] Damaged starch granules were hydrated and hydrolyzed with fungal alpha-amylase to give near complete solubilization of damaged granules with minimum breakdown of undamaged granules. The reaction was terminated with the addition of dilute sulfuric acid, and aliquots were treated with excess levels of purified amyloglucosidase to give complete degradation of starch-derived dextrins to glucose. The glucose was then spectrophotometrically measured at 510 nm (Beckman Coulter, DU 800, Fullerton, CA, USA) with a high purity glucose oxidase/peroxidase reagent mixture.
Antioxidant Analysis
The stable free radical 2,2-diphenyl-1-picrylhydrazyl (DPPH) method was used to determine the antioxidant capacity of wheat.[Citation16] For DPPH analyses following the method described by Miller et al.,[Citation16] 50 mL of 100 μM DPPH in 70% methanol were combined with 0.10 g of wheat flour prepared as described above, the samples were held at 37°C for 4 h, and absorbance readings were taken at 510 nm using a Beckman DU 640 UV/Vis Spectrophotometer (Beckman Coulter Ltd., Bedfordshire, UK).
Statistical Analysis
Experiments were performed in triplicate. Three lots of Apogee and Perigee were grown and analyzed, and three bags each of Yecora Rojo, Parshall, and Yavaros 79 were purchased and tested. These measures were taken to prevent subsampling from the sample source. The data were analyzed using SAS® 9.2 software (SAS Institute, Cary, NC, USA). The general linear model procedure was applied and Duncan's multiple range test was used to compare the mean values at P < 0.05.
RESULTS AND DISCUSSION
Protein Analysis
Previous studies using wheat exposed to elevated radiation (50–100 kGy) found degradation,[Citation17] specific and nonspecific amino acid damage,[Citation18] hydrogen bond disruption,[Citation19] and cleavage of disulfide bonds.[Citation20] In this study, wheat proteins were examined following lower radiation treatments of 0–100 Gy, which are more representative of NASA mission-relevant radiation doses than the previous studies. SDS-PAGE banding patterns and intensities were not consistent between cultivars following radiation exposure (a–). The Perigee cultivar had a decrease in protein concentration on the SDS-PAGE as radiation doses increased (, compare lanes 12 and 16) while the Apogee did not (). The terrestrial cultivars (represented by Yavaros 79 in ) displayed no changes in banding pattern or intensity following radiation exposure up to 100 Gy. Concentrations of albumin and globulin proteins in Apogee and Perigee were higher than levels observed in the terrestrial cultivars. Proximate analysis of these cultivars revealed that the protein concentrations in Apogee (18.4%) and Perigee (17.9%) were higher than Yecora Rojo (13.1%), Yavaros 79 (13.7%), and Parshall (16.9%).[Citation10]
Figure 1 SDS-PAGE of (a) Perigee, (b) Apogee, and (c) Yavaros 79 at various gamma radiation doses ranging from 0 to 100 Gy (A&G signifies albumin and globulin protein).
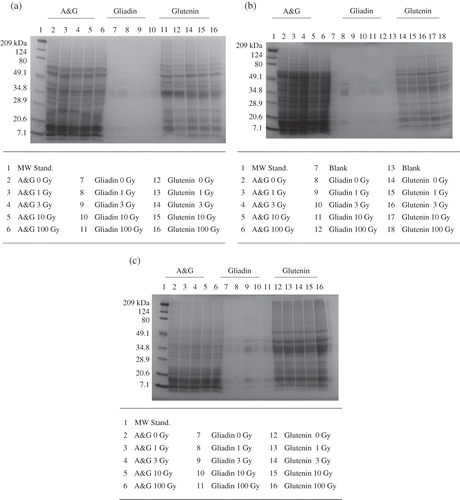
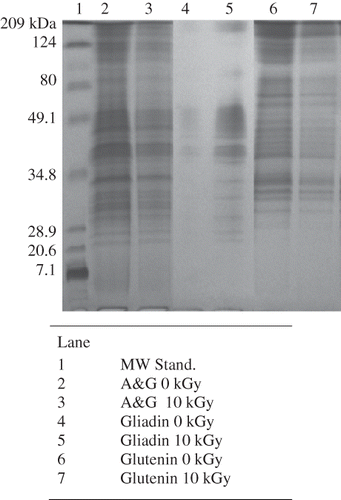
Maire and others[Citation21] investigated proteins exposed to high doses of radiation (0–400 kGy) and concluded that proteins contain sites that are susceptible to radiation-induced breakage. These fragile sites were located close to the C-terminal portion of the protein, and the susceptibility of proteins to radiation-induced fragmentation occurred independent of their size. Although the radiation treatments used in this study (1–100 Gy) were much lower than those used by Maire and others,[Citation21] radiation effects on wheat proteins were noticeable (). A dramatic reduction in the quantity of protein present in SDS-PAGE of wheat proteins exposed to 10 kGy gamma radiation () is likely a result of fragmentation similar to results observed in the Maire and others study.[Citation21]
Figure 2 SDS-PAGE of Yecora Rojo exposed to 10 kGy of gamma radiation (A&G signifies albumin and globulin protein).
Protein-protein interactions were evaluated using a mixograph,[Citation12] and peak dough development times are reported in . To facilitate data interpretation, Bruckner and others[Citation22] found that a longer time to peak dough development indicated a stronger gluten interaction and better bread quality. No differences were found between the control and 1 Gy samples; however, peak dough development times significantly decreased between 1 Gy and 3 Gy, and again between 3 Gy and 10 Gy. At doses of 10 Gy, the proteins reached their peak interaction strength more quickly than the control for all cultivars tested, indicating poorer gluten strength. This coincides with previous studies,[Citation9,Citation23] which found that low levels of radiation treatments (10–50 Gy) on medium wheat (11–13% protein) produced bread with volume and overall bread quality equal to or slightly better than those of the control flour. Additionally, Apogee had the significantly highest peak dough development time at all radiation doses except 10 Gy, at which point it had equivalent times to all other cultivars ().
Table 1 Peak dough development times for wheat flours from select cultivars exposed to various radiation doses (0–100 Gy)
Commercially, food products may be irradiated with doses up to 10 kGy to control pathogenic organisms, prevent mold growth, or limit insect infestation.[Citation24] Maire and others[Citation21] found that a 10 kGy dose fragmented wheat proteins. To visualize radiation effects on protein physical structures, atomic force microscopy was performed on gliadin exposed to 0 and 10 kGy (). The surface of gliadin in its native state is hydrophobic.[Citation25] When exposed to a hydrophilic surface such as the diamond used in this study, the gliadin aggregates and repels itself from the surface. This is evident by the 4-fold increase in the thickness of the gliadin layer (12 nm versus 3 nm high in the irradiated gliadin image) and the presence of fewer, larger pockets in the non-irradiated gliadin image (). Upon exposure of protein to radiation, increased availability of hydrophilic amino acids occurs.[Citation26] This phenomenon explains why after exposure to 10 kGy, the gliadin protein is attracted to, instead of repulsed by, the diamond surface and a more compact adhesion occurs. All the proteins isolated from wheat berries were influenced by 10 kGy of radiation as evident by fainter banding in Further AFM analyses on additional wheat proteins exposed to select radiation doses could be used to determine a threshold radiation level for observed protein structural differences.
Figure 3 AFM images of (a) non-irradiated gliadin and (b) irradiated (10 kGy) gliadin at 500 nm showing changes in surface topography.

Lipid Oxidation
Lipid oxidation increased with increasing radiation exposure for Apogee, Perigee, and Parshall (), the cultivars in this study with the highest levels of lipids and crude ash.[Citation10] Lipid oxidation significantly increased in Yavaros 79 from 0 to 1 Gy, but increasing radiation beyond 1 Gy, up to 100 Gy, had no further effect on lipid oxidation in this cultivar (). In Apogee, oxidation increased 52% from 0 to 1 Gy, 99.3% from 0 to 10 Gy, and 342.8% from 0 to 100 Gy. Lipid oxidation in Perigee increased 61.3% from 0 to 1 Gy, 252.3% from 0 to 10 Gy, and 497% from 0 to 100 Gy. Perigee had the largest increase in lipid oxidation from 0 to 100 Gy, while Yecora Rojo had the smallest increase (18.5%) over the same radiation range. Gruiz and Kiss[Citation27] found that increased lipid oxidation in wheat can be caused by increasing radiation doses (5–50 kGy). Gogolewski and others[Citation28] found that irradiation of wheat at 2.3–20 kGy led to both an increase in peroxide content and acceleration of the rate of tocopherol decomposition with increasing radiation dosage. They also reported that autoxidation of irradiated wheat oil led to a quicker end of shelf-life. Our study found that lower radiation doses of 1–100 Gy also have an effect on the amount of lipid oxidation in wheat berries. Lipid oxidation impacts the palatability and decreases the shelf-life of foods. Therefore, products which utilize wheat as an ingredient could be affected by space-relevant radiation doses, especially over a prolonged storage period.
Table 2 Lipid oxidation, determined by 2-thiobarbituric acid reactive substances test and reported as TEP equivalence, in wheat berries from select cultivars following exposure to elevated radiation (0–100 Gy)
Starch Analysis
Higher doses of radiation (1–5 kGy) have been shown to affect the starch structure[Citation29] and sensory characteristics[Citation30] of wheat, but the effects of lower radiation doses on wheat starch damage and functionality have not been reported. All wheat cultivars used in this study had significant decreases in peak swelling and retrogradation viscosities as radiation doses increased from 0 to 100 Gy ( and ), but increased damage to starch granules was not observed (). The Yecora Rojo starch control had the highest peak swelling and retrogradation viscosities at 433 cP and 442 cP, respectively, while the Yavaros 79 had the lowest viscosities across all radiation treatments, starting with the control at 56 and 53 cP, respectively ( and ). The peak swelling viscosities of all cultivars significantly decreased from 0 to 100 Gy (Apogee by 23.5%; Perigee by 31.3%; Yecora Rojo by 75.3%; Parshall by 34.5%; and Yavaros 79 by 71.4%). Retrogradation viscosities also decreased across this range of radiation exposure (Apogee by 49.5%; Perigee by 23.5%; Yecora Rojo by 76.2%; Parshall by 37.2%; and Yavaros 79 by 88.6%). No significant differences in peak swelling viscosity were found between 0 and 1 Gy, and only Perigee starch was significantly different than the control at 3 Gy (). The retrogradation viscosities for all starches except starch isolated from Perigee were significantly lower following exposure to 3 Gy of gamma radiation compared to the controls (). Radiation, therefore, directly affects the pasting profiles of wheat starch, even at space-relevant 1 or 3 Gy doses depending on the wheat cultivar.
Table 3 Peak swelling viscosity of wheat starch from various cultivars exposed to 0–100 Gy of radiation
Table 4 Retrogradation viscosity of wheat flour from various cultivars exposed to 0–100 Gy gamma radiation
Table 5 Percent starch damage of select wheat cultivars at mission relevant doses of radiation (0–100 Gy) as described by AACC method 76–31
It appears that low doses of radiation weaken ( and ), but do not consistently damage (), the wheat starch granule. Starch from Perigee was significantly more damaged than starch from all other cultivars, across all radiation treatments up to 100 Gy (). Additionally, only starch from Perigee was significantly more damaged following the 100 Gy radiation treatment than its control. These higher levels of starch damage in Perigee could indicate that Perigee has a weaker starch granule structure than the other wheat cultivars examined.
Antioxidant Capacity
Differences in antioxidant capacity are evident between cultivars and radiation treatments (). For Apogee and Perigee cultivars, trolox equivalence significantly decreased as radiation exposure increased. The antioxidant capacities of Yecora Rojo, Parshall, and Yavaros 79 cultivars were not as affected by radiation. From 0 to 100 Gy, the antioxidant capacities of the cultivars decreased as follows: Apogee by 61.5 %; Perigee by 66.5%; and Parshall by 19.3%. No significant differences in antioxidant capacity were found between 0 and 100 Gy for Yecora Rojo or Yavaros 79 cultivars, and Yecora Rojo had the significantly lowest antioxidant capacity among the control samples. In un-irradiated wheats, Perigee had the highest antioxidant capacity, almost double that of the other cultivars (). At both 10 Gy and 100 Gy treatments, Yavaros 79 had the highest antioxidant capacity.
Table 6 Antioxidant capacity, determined by DPPH analysis and reported as trolox equivalence, of wheat berries from select cultivars following exposure to elevated radiation (0–100 Gy)
The precise mechanism for the deterioration of antioxidant activity upon increasing radiation exposure for Apogee, Perigee, and Parshall cultivars was not determined. Ferulic acid is the most prevalent antioxidant in wheat and is primarily located in the cell walls of the bran tissue in the form of diferulic linkages.[Citation31] Radiation has been used as a source for cross-linking components of the cell wall, and a similar mechanism may be occurring in the cell wall of wheat to decrease accessibility to the antioxidants. Ferulic, vanillic, and p-coumaric acids are the major phenolics in wheat bran extracts, along with other free phenolics including caffeic, chlorogenic, gentisic, syringic, and p-hydroxybenzoic acids.[Citation32] It has also been noted that phenolic acids are concentrated in the bran and aleurone fractions of wheat.[Citation33] Apogee, Perigee, and Parshall have the three smallest berry sizes and therefore highest surface area per gram of wheat of the five cultivars tested.[Citation10] The surface to volume ratios of these cultivars are higher than the cultivars with more stable antioxidant capacities, suggesting that increased radiation exposure of the bran and aleurone fractions increases the amount of antioxidant degradation.
CONCLUSION
Expected Mars-mission radiation doses (1–100 Gy) affect the protein, lipid, starch, and antioxidant characteristics of wheat berries, and effects are different between wheat cultivars (Apogee, Perigee, Parshall, Yavaros 79, and Yecora Rojo). Further studies are needed to optimize selection of wheat cultivars for growth in space or use in bulk food systems based on performance following radiation exposure, or a system to protect wheat from radiation exposure should be employed. Enough biomass should be generated for each cultivar and radiation treatment to enable baking studies to determine if radiation-induced effects are detectable in food production and by a sensory panel. Of the two super-dwarf cultivars used, Apogee had a more stable lipid and antioxidant profile and more stable starch granules than Perigee. The protein profile of Apogee remained stable with increasing radiation while the protein in Perigee degraded. Therefore, Apogee is likely a better NASA candidate crop than Perigee; however, terrestrial wheat cultivars may be more stable to radiation and provide better food functionality traits than either Apogee or Perigee.
ACKNOWLEDGMENTS
This research was partially funded by NASA grant NAG5-12686. The authors would like to acknowledge Dr. Cary Mitchell and Dr. Gioia Massa for their assistance in cultivating the wheat crops in the greenhouse at Purdue University. They also wish to thank Dr. Bruce Bugbee for providing wheat berries needed for production of Apogee and Perigee biomass Dr. Bruce Hamaker for use of the cereal lab facilities and equipment.
REFERENCES
- Perchonok , M.H. 2003 . Advanced food technology workshop report–Volumes I, II , Houston , TX : National Aeronautics and Space Administration .
- Perchonok, M.H. NASA packaged food systems. World Food Science, Vol. 2, 2007 http://www.iufost.org/publications/world_of_food_science/ (http://www.iufost.org/publications/world_of_food_science/) (Accessed: 24 September 2009 ).
- Eckart , P. 1994 . Life Support & Biospherics , Munchen , , Germany : Herbert Utz Publishers .
- Milner , M. 1957 . Some effects of gamma irradiation on the biochemical storage and breadmaking properties of wheat . Cereal Science Today , 2 : 130 – 137 .
- Hartung , T.E. , Bullerma , L.B. , Arnold , R.G. and Heidelba , N.D. 1973 . Application of low-dose irradiation to a fresh bread system for space flights . Journal of Food Science , 38 ( 1 ) : 129 – 132 .
- Henninger , D.L. and Escobedo , V.M. 1998 . Baseline crops for advanced life support program , Houston , TX : National Aeronautics and Space Administration .
- Bugbee, B. Historical summary of the selection of USU-Perigee. 1999 http://www.usu.edu/cpl/Progression.pdf (http://www.usu.edu/cpl/Progression.pdf) (Accessed: 28 July 2008 ).
- Bugbee , B. , Koerner , G. , Albrechtsen , R. , Dewey , W. and Clawson , S. 1996 . ‘USU-Apogee’ Wheat Registration. ,
- Gandolph, J.; Shand, A.; Stoklosa, A.; Ma, A.; Weiss, I.; Alexander, D.; Perchonok. M.H.; Mauer, L.J. Foods for a mission to Mars: Investigations of low-dose gamma radiation effects. World Food Science, Vol. 2, 2007 http://www.iufost.org/publications/world_of_food_science/ (http://www.iufost.org/publications/world_of_food_science/) (Accessed: 24 September 2009 ).
- Stoklosa , A.M. , Weiss , I. , Bugbee , B. , Perchonok , M.H. and Mauer , L.J. 2011 . Composition and functional properties of Apogee and Perigee compared to common terrestrial wheat cultivars . International Journal of Food Properties , 14 : 996 – 1006 .
- Hamaker , B.R. , Mohamed , A.A. , Habben , J.E. , Huang , C.P. and Larkins , B.A. 1995 . Efficient procedure for extracting maize and sorghum kernel proteins reveals higher prolamin contents than the conventional method . Cereal Chemistry , 72 ( 6 ) : 583 – 588 .
- American Association of Cereal Chemists . 1995 . Approved methods of the AACC , 9th , St Paul , MN : AACC .
- Makarenko , S. , Trufanov , V. and Putilina , T. 2002 . Infrared spectroscopic study of the secondary structure of wheat, rye, and barley prolamins . Russian Journal of Plant Physiology , 49 ( 3 ) : 326 – 331 .
- Kulisic , T. , Radonic , A. , Katalinic , V. and Milos , M. 2004 . Use of different methods for testing antioxidative activity of oregano essential oil . Journal of Food Chemistry , 85 : 633 – 640 .
- Grant , L.A. 1998 . Effects of starch isolation, drying, and grinding techniques on its gelatinization and retrogradation properties . Cereal Chemistry , 75 ( 5 ) : 590 – 594 .
- Miller , H.E. , Rigelhof , F. , Marquart , L. , Prakash , A. and Kanter , M. 2000 . Whole-grain products and antioxidants . Cereal Foods World , 45 ( 2 ) : 59 – 62 .
- Deschreider , A.R. June 1966 . “ Action of the gamma rays on the constituent elements of the wheat flour ” . In Proceedings of International Atomic Energy Agency Symposium, IAEA/SM-73/4 June , 6 – 10 . Karlsruhe , , Germany
- Patten , R.A. and Gordy , W. 1964 . Further studies of radiation effects on the proteins and their constituents . Radiation Research , 22 : 29 – 34 .
- Williams , J.F. and Hunt , J.W. 1963 . Molecular lesions produced in ribonucleases by gamma rays . Nature , 200 : 779 – 785 .
- Doguchi , M. 1969 . Effects of gamma irradiation on wheat gluten . Agriculture and Biological Chemistry , 33 : 1769 – 1774 .
- Maire , M.I. , Thauvette , L. , Foresta , B. , Viel , A. , Beauregard , G. and Potier , M. 1990 . Effects of ionizing radiations on proteins . Biochemical Journal , 267 ( 2 ) : 431 – 439 .
- Bruckner , P.L. , Habernicht , D. , Carlson , G.R. , Wichman , D.M. and Talbert , L.E. 2001 . Comparative bread quality of white flour and whole grain flour for hard red spring and winter wheat . Crop Science , 41 ( 6 ) : 1917 – 1920 .
- Paredes , O.L. and Covarrubias , M.M. 1984 . Influence of gamma radiation on the rheological and functional properties of bread wheats . International Journal of Food Technology , 19 ( 2 ) : 225 – 231 .
- IAEA . October 12–16 1998 . Report on the Joint AAEA/FAO/IAEA Regional Workshop on Present Status and Guidelines for Preparing Harmonized Legislation on Food Irradiation in the Near East , October 12–16 , Tunis : Tunisia .
- Arangoa , M.A. , Ponchel , G. , Orecchioni , A.M. , Renedo , M.J. , Duchene , D. and Irache , J.M. 2000 . Bioadhesive potential of gliadin nanoparticulate systems . European Journal of Pharmaceutical Science , 11 ( 4 ) : 333 – 341 .
- Matloubi , H. , Aflaki , F. and Hadjiezadegan , M. 2004 . Effect of gamma-irradiation on amino acids content of baby food protein . Journal of Food Composition and Analysis , 17 ( 2 ) : 133 – 139 .
- Gruiz , K. and Kiss , I. 1987 . Effect of ionizing radiation on the lipids in frozen poultry . Acta Alimentaria , 16 ( 2 ) : 111 – 127 .
- Gogolewski , M. , Bartowiak , F. and Stepniak , A. 1999 . The effects of ionizing radiation on the quality of selected vegetable oils . Bromatology I Chemistry and Toxicology , 32 ( 1 ) : 21 – 26 .
- Fenno , C.W. 1985 . Use of specially treated starches in bread baking . Dissertation Abstracts International , 46 ( 1 ) : 13
- Brownell , L.E. , Harlin , H.A. and Nehemias , J.V. 1955 . A preliminary study of the effect of gamma radiation on the baking quality of flour . Food Technology , 9 : 620 – 623 .
- Bunzel , M. , Ralph , J. , Marita , J.M. , Hatfield , R.D. and Steinhart , H. 2001 . Diferulates as structural components in soluble and insoluble cereal dietary fiber . Journal of Science and Food Agriculture , 81 ( 7 ) : 653 – 660 .
- Onyeneho , S.N. and Hettiarachchy , N.S. 1992 . Antioxidant activity of durum wheat bran . Journal of Agricultural and Food Chemistry , 40 : 1496 – 1500 .
- Graf , E. 1992 . Antioxidant potential of ferulic acid . Free Radical Biology and Medicine , 13 : 435 – 448 .