
ABSTRACT
The compositions and inosinic acid contents of ATP-related compounds in various processed fish meat products were examined in this study. Moreover, the relationships between these variables and the heating processes used on the products were investigated. Dried and salted fish meat products manufactured immediately after manufacturing, were found to maintain some inosinic acid contents, whereas those stored for a long period of time had low inosinic acid contents. It was observed that the inosine content in processed fish meat products manufactured without the heat treatment was decreased during the storage at 5°C. Therefore, the low inosinic acid content in such processed fish meat products was considered to be because the maintenance of inosine monophosphate (IMP)-degrading enzyme activity, resulting in the decomposition of inosinic acid even under refrigerated storage. The inosine content of unheated water extract decreased faster than that of fish meat product, and the effect of heating temperature could be observed more clearly. The results suggested that IMP-degrading enzymes in club mackerel, Atlantic bluefin tuna, and Japanese jack mackerel were deactivated by heating at 70°C or higher, while those in fine patterned puffer and half-smooth golden pufferfish were deactivated with heating at 60°C or higher. Since IMP decomposition was suppressed incrementally with heating, it appears that the multiple enzymes involved in the decomposition of inosinic acid might have varying thermal stabilities.
Introduction
Fish and shellfish have high water contents, and as such, they decay at room temperature in a relatively short period. Dried fish meat products, whose water contents are reduced by drying in order to enhance preservation of the product, include plain-dried fish, dried and salted fish, boiled and dried fish, fushi products, and smoked fish. The wide variety of available dried fish meat products is a result of the various types of fish and shellfish that can be used as raw materials. In particular, dried and salted fish, where preservability is enhanced by immersion in salt followed by completion of a drying process, is a simple fish meat product.[Citation1–Citation3]) The water activity of dried and salted fish is low so that the growth of microorganisms and the enzyme activity are effectively suppressed, thus enhancing the potential storage time of these products.[Citation4,Citation5]) In Japan, dried and salted products are important, accounting for about half of the country’s dried fish meat products, with an annual production of around 13.9 million tons.[Citation6]) The umami components of fish meat include inosinic acid or inosine monophosphate (IMP), amino acids such as glutamic acid, and lipids.[Citation7–Citation9]) This study focused on one of these components, IMP. IMP is a degradation product of the initial material, adenosine triphosphate (ATP). The ATP-related compounds in fish meat decompose in the order of ATP, adenosine diphosphate (ADP), adenosine monophosphate (AMP), inosine monophosphate (IMP), inosine (HxR), and hypoxanthine (Hx). Although most of the compounds are present as ATP in live muscles, they begin to decompose after death. In fish, only a small amount of the compounds remain as ADP and AMP, with most of them accumulating as IMP.[Citation10,Citation11]) While IMP is stable during heating, studies of several fish species have confirmed that IMP is decomposed into inosine (HxR) as it is dephosphorylated by enzymes in fish meat such as 5ʹ-nucleotidase and acid phosphatase.[Citation12–Citation16]) It has been reported that the purification of IMP-degrading enzymes varies among Alaska pollack,[Citation14]) chicken grunt,[Citation15,Citation16]) and carp.[Citation17]) Further, IMP-degrading enzyme activity varies between carp dark muscles and ordinary muscles.[Citation18])
It is important to determine the properties of the endogenous IMP-degrading enzymes as these have implications for the manufacturing and long-term storage of processed fish meat products containing high IMP contents. Of the various types of dried fish meat products, those plain-dried and those dried and salted, which are simple and easy manufacturing processes, do not usually undergo heating processes. Even if the fish meat is processed with a maximum IMP count, it is expected that the enzymes work to decompose the IMP during storage.
In this study, the amount of ATP-related compounds in commercially available dried and salted fish and fushi products were firstly investigated. Then, the effects of the storage period and the presence or absence of a heating process on the IMP content in each product were examined. Dried and salted products were manufactured and the behavior of IMP during manufacturing and storage was examined. All of the ATP-related compounds and IMP-degrading enzymes contained in fish meat are water-soluble components. Therefore, if these components are transformed into water extracts, the behaviors of IMP and IMP-degrading enzymes in fish meat would be easier to confirm. Thus, according to the method of Tomioka et al.,[Citation12]), water extracts of several types of fish meat were prepared and the thermal stabilities of their IMP-degrading enzymes were examined.
Experimental method
Experimental samples
The following processed fish meat products (half-dried products) were purchased from Karato Market in Shimonoseki, Yamaguchi Prefecture: club mackerel (Scomber japonicas), half-smooth golden pufferfish, tilefish, yellow croaker (Larimichthys polyactis), blackthroat seaperch (Doederleinia berycoides), brown barracuda (Sphyraena pinguis), Japanese Spanish mackerel (Scomberomorus niphonius), two types of Japanese jack mackerel (Trachurus japonicus), Japanese armorhead (Pentaceros japonicus), golden threadfin bream (Nemipterus virgatus), Okhotsk atka mackerel (Pleurogrammus azonus), bensasi goatfish (Upeneus japonicus), righteye flounder, flathead, chicken grunt (Parapristipoma trilineatum), and Japanese sardine (Sardinops melanostictus). Boiled and half-dried skipjack tuna (Katsuwonus pelamis) products were also purchased from the same market ().
Table 1. Details of the processed fish products investigated in this study.
The following fresh fish were purchased from the same market: club mackerel, Atlantic bluefin tuna (thunnus thynnus), Japanese jack mackerel, fine patterned puffer (Takifugu flavipterus), and half-smooth golden pufferfish (Lagocephalus wheeleri).
Manufacturing and storage of dried and salted Japanese jack mackerel
After removing the head of the fresh Japanese jack mackerel, the belly was split to remove the internal organs. Dried and salted products were manufactured by immersing the fish in 5% (w/v) saline for 30 minutes and then drying it overnight using a fish cooling dryer (FV50 W-30 H, Fujiki Industries, Ltd.). The manufactured dried and salted products were stored at 5°C.
Preparation and heat treatment of fish meat water extracts
The fish meat water extracts were prepared using a modified version of the method described by Tomioka et al.[Citation12]) The method was as follows: 300 g of ordinary muscles from each fish species (club mackerel, Atlantic bluefin tuna, Japanese jack mackerel, fine-patterned puffer, and half-smooth golden pufferfish) were obtained. Then, 300 mL of distilled water was added to each muscle sample and the mixture was homogenized using a homogenizer (PH91 model, SMT Co., Ltd.) at 20,000 rpm for 9 min. The resulting samples were then centrifuged (centrifuge: 5500 model from Kubota Corporation) at 8,590 x g for 10 minutes to obtain a supernatant. The supernatant was filtered through gauze to a constant volume of 600 mL and was then dispensed into six 100 mL glass beakers. One of the beakers was not heated while the other five were heated in a constant-temperature bath (T-104NA model, Thomas Kagaku Co., Ltd.) at 40°C, 50°C, 60°C, 70°C, or 80°C for 20 minutes, and then immediately cooled in ice water and stored at 5°C in a low-temperature refrigerator/freezer (SC-DF25 model, Twinbird Corporation).
Extraction of ATP-related compounds
ATP-related compounds were extracted according to the method of Maeda et al.[Citation19]) as follows: muscle samples of approximately 5 g were obtained. Then, 15 mL of 10% perchloric acid water solution was added and the mixture was pulverized using a polytron homogenizer (PT2100 model, Kinematica). The resulting sample was then filtered using filter paper (No. 2 filter paper from Toyo Roshi; 110 mm diameter). Next, 10 mL of 5% perchloric acid water solution was added to the residue on the filter paper; the two were mixed well and the solution was left to stand for approximately 30 seconds before it was filtered again. This procedure was repeated twice. After adjusting the pH level of the filtrate to approximately 6.4 using 10NKOH and 1NKOH, the filtrate was filtered and distilled water was added to a constant volume of 50 mL to obtain the extract.
ATP-related compounds using high-performance liquid chromatography
ATP-related compounds were determined using a modified HPLC-based method.[Citation20]) After filtering the extract described above using a syringe filter with a pore diameter of 0.45 μm (Sartorius), it was placed in a vial and analyzed using an HPLC system consisting of an L-7100 pump, an L-7610 degasser, an L-7200 auto-sampler, an L-7300 column oven, an L-7400 UV detector, and a D-7500 chromatography data processor (Hitachi, Ltd.). The HPLC conditions were as follows: flow rate 1.0 mL/min, column temperature 35°C, analysis volume 10 μL, and detection wavelength 260 nm. The column used in the system was an Inertsil ODS-4 from GL Sciences (column size: 5 μm 4.6 × 150 mm).
Results
IMP contents of commercially available processed fish meat products
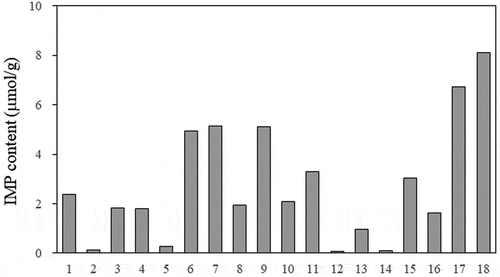
The ratios of ATP-related compounds in a total of 18 dried and salted products and fushi products () are shown in and the IMP contents are shown in . The total amount of ATP-related compounds in each product was in the range of 5 to 13 μmol/g (data not shown). The ratios of ATP, ADP, and AMP in the compounds were low, and most were accounted for by IMP, HxR, and Hx. IMP contents ranged from 0.08 to 8 μmol/g, the ratio of IMP to ATP-related compounds ranged from 1% to 68%, and the K value ranged from 27% to 92%. Thus, great differences were observed among the products. When individual products were compared, the IMP content varied greatly depending on the fish species and the manufacturing methods used for processing the fish meat products; for example, the IMP content was high in fushi products and low in pufferfish products. Fushi products, in particular, had the highest IMP content of all products. The results also indicated that the IMP content in the dried and salted products varied depending on their storage period. For example, products such as Japanese jack mackerel (No. 9) and Japanese sardine (No. 17), when measured 1 dayafter manufacturing, maintained relatively high IMP contents, while half-smooth golden pufferfish (No. 2), blackthroat seaperch (No. 5), and Okhotsk atka mackerel (No. 12) products that had been manufactured several months earlier exhibited extremely low IMP contents.
Figure 1. The ratios of ATP-related compounds in various dried and salted fish and fushi products. The sample numbers are listed in.
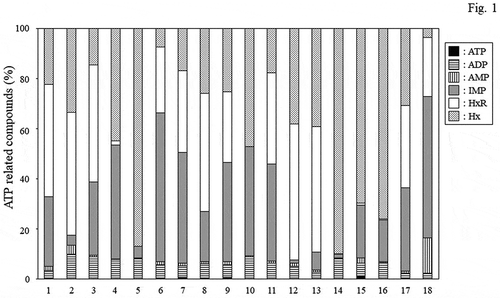
Figure 2. IMP contents in various half-dried fish and fushi products. The sample numbers are listed in.
IMP contents in half-dried Japanese jack mackerel products
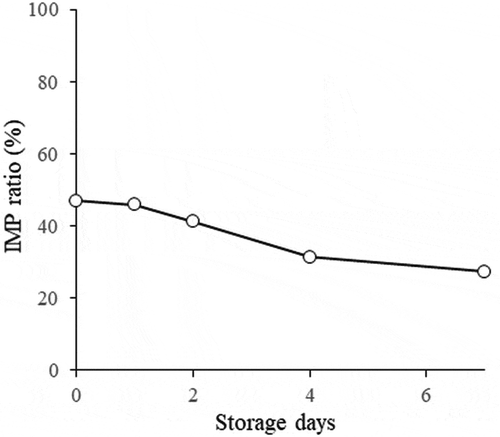
The IMP ratios of the dried and salted Japanese jack mackerel products, that were manufactured as part of this study and were stored at 5°C for 7 days after manufacturing, are shown in . The IMP ratio before salting was 48%. The IMP ratio after salting was 49%, and after drying, the IMP ratio was 48%. The results indicate that the IMP ratio was almost constant within one day of manufacturing, which is a typical period for manufacturing of dried and salted products (data not shown). The IMP ratio subsequently declined gradually during storage at 5°C: to 46% on day 1 of storage, 41% on day 2, 32% on day 4, and 26% on day 7 of storage.
Figure 3. Changes in the IMP ratios of half-dried Japanese jack mackerel products during storage for 7 days at 5°C.
Changes in the IMP ratio in water extracts of various fish muscles
After adding equal parts of water to the five ordinary fish muscles, the mixtures were homogenized and centrifuged to obtain supernatants. These supernatants were used to examine the thermal stabilities of the IMP-degrading enzymes. After processing water extracts of club mackerel without heat treatment or with heat treatment at 40°C to 80°C for 20 minutes, the extracts were stored at 5°C and the IMP ratios were measured each day until day 7. The results are shown in . The IMP ratio at the start of storage was about 80% in both the unheated sample and the samples heated at 40°C to 80°C for 20 minutes. In the unheated sample, the IMP ratio declined during storage at 5°C, down to 25% on day 7 of storage. A similar trend was observed in the sample processed with 40°C heat; the ratio declined to 29% on day 7 of storage. Heating at 50°C or higher suppressed this reduction in the IMP ratio with an increasing effect observed with increasing temperatures. Specifically, the IMP ratio declined to 46% on day 7 of storage with 50°C heating, 67% with 60°C heating, 76% with 70°C heating, and 77% with 80°C heating. Thus, heating at 70°C or higher maintained the IMP ratio that was observed at the beginning of storage.
Figure 4. Changes in the IMP ratios of water extracts of club mackerel (A), Atlantic bluefin tuna (B), Japanese jack mackerel (C), fine-patterned puffer (D), and half-smooth golden pufferfish (E) stored for seven days.
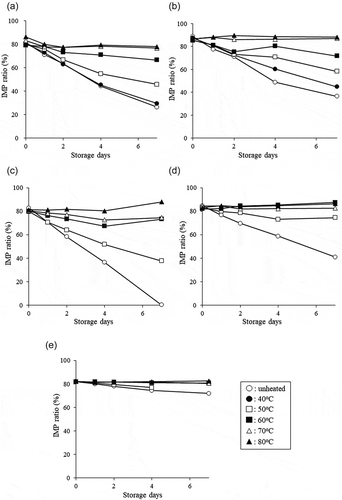
Changes in the IMP ratios in the water extracts of Atlantic bluefin tuna are shown in . Although the IMP ratios in all samples at the beginning of storage were about 90%, they declined in the unheated sample, down to 36% on day 7 of storage. The heated extracts exhibited suppressed IMP ratio reduction during storage, with stronger effects evident with increasing heating temperature. Specifically, the IMP ratio was 45% on day 7 of storage with 40°C heating, 58% with 50°C heating, 72% with 60°C heating, 87% with 70°C heating, and 88% with 80°C heating. Thus, consistent with the findings in club mackerel, heating at 70°C or higher maintained the ratio that was observed at the beginning of storage.
IMP ratio measurements in the water extracts of Japanese jack mackerel are shown in . Although the IMP ratios in all samples at the beginning of storage were about 80%, they declined in the unheated sample, down to 0% on day 7 of storage. In the heated extracts, there was suppression of this IMP ratio reduction during storage, with stronger effects associated with increasing heating temperatures. Specifically, the IMP ratio was 38% on day 7 of storage with 50°C heating, 73% with 60°C heating, 74% with 70°C heating, and 88% with 80°C heating. Thus, heating at 80°C or higher maintained the ratio that was observed at the beginning of storage.
IMP ratio measurements in the water extracts of fine-patterned puffer are shown in . Although the IMP ratios in all samples at the beginning of storage were 83–85%, they declined in the unheated sample, down to 41% on day 7 of storage. Heating suppressed this IMP ratio reduction, down to 75% with 50°C heating. However, there was no apparent reduction with heating at 60°C or higher.
IMP ratio measurements in water extracts of half-smooth golden pufferfish are shown in . The IMP ratios in all samples at the beginning of storage were about 80%. In the unheated sample, the IMP ratio declined during storage, down to 72% on day 7 of storage. Heating suppressed this reduction in the IMP ratio, down to 77% with 50°C heating on day 4 of storage. However, there was no apparent reduction in samples heated at 60°C or higher.
Discussion
The focus of this study was IMP, one of the umami components of processed fish meat products. Firstly, how much IMP is contained in processed fish meat products that are available on the market was investigated and from this, relationships between manufacturing processes and storage periods were inferred. Then, the thermal stabilities of the IMP-degrading enzymes that are involved in the decomposition of IMP were analyzed in each fish species.
The IMP content in commercially available dried and salted fish and fushi products was measured to assess the current status of these commercial products. After the death of fish, ATP-related compounds are degraded by muscle-inherent enzymes. In many fish species, IMP accumulates over a certain period of time.[Citation10,Citation11]) In all of the dried and salted products investigated in this study, the ratios of ATP, ADP, and AMP in the compounds were low, and most were accounted for by IMP, HxR, and Hx. Likewise, the IMP ratios and contents were low in many products ( and ). The results also indicated that the composition of IMP, HxR, and Hx varied depending on the product. This is likely because the activity of the IMP-degrading enzymes and the storage period vary among fish species.[Citation12])
Both the IMP ratios and contents were higher in the fushi product (No. 18) than in the dried and salted products. In the initial manufacturing stages of fushi products, skipjack tuna is boiled and goes through various steps such as fermentation and drying, before being processed into katsuobushi (dried bonito).[Citation21]) Katsuobushi is a product containing a large amount of IMP[Citation22]) and is typically used as an ingredient in dashi (soup stock). The higher IMP ratios and contents in fushi products are likely attributed to the heating process during the initial manufacturing stages, which deactivated the IMP-degrading enzyme, leading to maintenance of a high IMP content.
When the IMP contents of two dried and salted products of Japanese jack mackerel (No. 8 and No. 9) that had been stored for different numbers of days after manufacturing was compared, a higher IMP content was observed in the product with a shorter storage period. However, it was likely that this result was the result of a difference in the initial freshness of the fish. Therefore, dried and salted Japanese jack mackerel products was manufactured for traceable histories of handling, and then the IMP contents in the products were tracked during processing and storage (). Although there was no noticeable reduction in the IMP ratio within 1 day of product processing, on day 7 of storage, the IMP ratio declined to about half its level on the first day of storage. This confirms that IMP decomposes even during refrigerated storage.
Based on these results, it is evident that IMP in processed fish meat products manufactured without a heating treatment, such as dried and salted fish meat products, decomposes gradually, even if they are stored at low temperatures. The IMP decomposition rate was also found to vary depending on the fish species used as the raw material. Therefore, it was considered important to understand the properties of the IMP-degrading enzymes in each raw material.
Generally, enzyme activity is low at refrigeration temperatures, and increases with rises in temperature. When the temperature exceeds the thermal denaturation temperatures of enzymes, enzyme activity is lowered and eventually deactivated. Therefore, the heating temperatures at which the degrading enzymes were deactivated were examined using water extracts of highly fresh catch of five fish species. A sufficient amount of IMP was contained in the water extract from each fish species for the purpose of this study. When the water extracts were processed without heating and were stored at 5°C, the IMP contents declined in the extracts from all fish species. In Japanese jack mackerel, which exhibited the greatest IMP ratio reduction among all the fish species, the 80% IMP ratio at the beginning of storage declined to 0% on day 7, whereas in half-smooth golden pufferfish, which exhibited the smallest IMP ratio reduction, the 80% IMP ratio at the beginning of storage declined to 70% on day 7. These results also suggested that the activity and optimum temperature of the IMP-degrading enzymes varied noticeably among the fish species. Of note, when the water extracts were processed with heating, the decomposition of IMP was suppressed as the heating temperature rose. When club mackerel, Atlantic bluefin tuna, and Japanese jack mackerel were heated at 70°C or higher, the decomposition of IMP was suspended. Comparison of the water extract and the processed Japanese jack mackerel product (, ) revealed that even though the IMP decomposition rate in the fish muscles (product) was slower than that in the water extract, decomposition was still occurring even with storage at 5°C.
For the pufferfish (globefish) species, two fresh catch of fine-patterned puffers and half-smooth golden pufferfish were compared each other. With heating at 60°C or higher, the two types of pufferfish maintained the same IMP ratio as was observed at the beginning of storage (). Since the decomposition of IMP in both types of pufferfish was suppressed with heating temperatures of 60°C, which is lower than that required to suppress IMP decomposition in other fish species, it appears that the thermal stabilities of the IMP-degrading enzymes in these fish were low. In particular, in an unheated sample of half-smooth golden pufferfish, IMP decomposition was significantly low. There are many possible reasons for this finding, including the low activity of the IMP-degrading enzymes in half-smooth golden pufferfish, low enzyme activity at a storage temperature of 5°C, and low enzyme content. Future studies should investigate this further.
In actual processed fish meat products, the pH level and salt content vary depending on the freshness of raw fish and how the fish is seasoned/prepared. It has been reported that IMP-degrading enzymes are affected by the presence of salt[Citation23–Citation25]) and by the pH level,[Citation26]) where the addition of salt is effective in suppressing IMP decomposition. However, it is not possible to completely suspend the decomposition of IMP with salt. This study also confirmed that IMP in processed fish meat products, such as dried and salted fish products manufactured without heating treatment, decomposes even during refrigerated storage. IMP-degrading enzymes show varying thermal stabilities depending on the fish species. In the various types of fish meat examined in this study, heating at 60–70°C deactivated the IMP-degrading enzymes, suspending the decomposition of IMP. These findings are considered useful for IMP content management in various processed fish meat products.
Conclusion
The IMP contents of processed fish meat products available on the market were firstly investigated. The results showed that the IMP contents in dried and salted products manufactured without the use of heating processes declined in long-term storage. This suggests that processed fish meat products manufactured without the use of heating processes maintain the activity of IMP-degrading enzymes, resulting in decomposition of inosinic acid even during refrigerated storage. Since heated products were found to contain high IMP contents, it appears that heating deactivated the IMP-degrading enzymes, preventing the decomposition of IMP. The results also suggested that IMP-degrading activity is species-specific. Therefore, water extracts containing IMP and IMP-degrading enzymes were prepared from several types of fish meat in order to examine the thermal stabilities of the IMP-degrading enzymes. The results showed that the thermal stabilities of the degrading enzymes were species-specific; the enzymes in club mackerel, Atlantic bluefin tuna, and Japanese jack mackerel were deactivated by heating processes at 70°C or higher, while those in pufferfish (globefish) species were deactivated by heating processes at 60°C or higher.
Acknowledgments
We would like to thank the late Prof. Yutaka Fukuda for drafting this study. We would also like to thank Scientific Language (www.scientific-language.co.jp) for the English language review.
References
- Sampels, S.;. The Effects of Processing Technologies and Preparation on the Final Quality of Fish Products. Trends Food Sci. Technol. 2015, 44(2), 131–146. DOI: 10.1016/j.tifs.2015.04.003.
- Bellagha, S.; Sahli, A.; Farhat, A.; Kechaou, N.; Glenza, A. A Studies on Salting and Drying of Sardine (Sardinella Aurita): Experimental Kinetics and Modelling. J. Food Eng. 2007, 78, 947–952. DOI: 10.1016/j.jfoodeng.2005.12.008.
- Jian., L.; Li, Q.; Zhang., L.; Zhang, J.; Dong, Z.; Feng, L.; Luo, Y. Changes in Quality of Rainbow Trout (Oncorhynchus Mykiss) Fillets Preserved with Salt and Sugar at Low Concentrations and Stored at 4°C. Int. J. Food Prop. 2017, 20(10), 2286–2298. DOI: 10.1080/10942912.2016.1236272.
- Guizani, N.; Al-Shoukri, A. O.; Mothershaw, A.; Rahman, M. S. Effects Of Salting And Drying On Shark (Carcharhinus sorrah) Meat Quality Characteristics. Dry. Technol. 2008, 26(6), 705–713. DOI: 10.1080/07373930802046294.
- Oliveira, H.; Pedro, S.; Nunes, M. L.; Costa, R.; Vaz-Pires, P. Processing of Salted Cod (Gadus Spp.): A Review. Compr. Rev. Food Sci. 2012, 11, 546–564. DOI: 10.1111/j.1541-4337.2012.00202.x.
- Ministry of Agriculture, Forestry and Fisheries. Statistics of Distribution of Fishery Production in Japan; Statistics Dept. of the Ministry of Agriculture: Tokyo, 2018.
- Yamaguchi, S.;. The Synergistic Taste Effect of Monosodium Glutamate and Disodium 5ʹ-inosinate. J. Food Sci. 1967, 32(4), 473–478. DOI: 10.1111/j.1365-2621.1967.tb09715.x.
- Otsuka, S.;. Umami in Japan, Korea, and Southeast Asia. Food Rev. Int. 1998, 14(2–3), 247–256. DOI: 10.1080/87559129809541159.
- Hajeb, P.; Jinap, S. Umami Taste Components and Their Sources in Asian Foods. Crit. Rev. Food Sci. Nutr. 2015, 55(6), 778–791. DOI: 10.1080/10408398.2012.678422.
- Li, K.; Luo, Y.; Shen, H. Postmorten Changes of Crucian Carp (Carassius Auratus) during Storage in Ice. Int. J. Food Prop. 2015, 18, 205–212. DOI: 10.1080/10942912.2011.614986.
- Scherer, R.; Augusti, P. R.; Bochi, V. C.; Steffens, C.; Fries, L. L. M.; Daniel, A. P.; Kubota, E. H.; Neto, J. R.; Emanuelli, T. Chemical and Microbiological Quality of Grass Carp (Ctenopharyngodon Idella) Slaughtered by Different Methods. Food Chem. 2006, 99, 136–142. DOI: 10.1016/j.foodchem.2005.06.048.
- Tomioka, K.; Endo, K. K Value-increasing Rates and IMP-degrading Activities in Various Fish Muscles. Bull. Japan. Soc. Sci. Fish. 1984, 50, 889–892. DOI: 10.2331/suisan.50.889.
- Tomioka, K.; Endo, K. Properties of 5ʹ-nucleotidase from Carp Muscle. Bull. Japan. Soc. Sci. Fish. 1984, 50, 1739–1744. DOI: 10.2331/suisan.50.1739.
- Tarr, H. L. A.; Gardner, L. J.; Ingram., P. Pacific Cod Muscle 5′‐Nucleotidase. J. Food Sci. 1969, 34, 637–640. DOI: 10.1111/j.1365-2621.1969.tb12110.x.
- Marseno, D. W.; Hori, K.; Miyazawa, K. Purification and Properties of Membrane-bound 5ʹ-nucleotidase from Black Rockfish (Sebastes Inermis) Muscle. J. Agri. Food Chem. 1993, 41, 863–869. DOI: 10.1021/jf00030a004.
- Marseno, D. W.; Hori, K.; Miyazawa, K. Purification and Properties of Cytosol 5ʹ-nucleotidase from Black Rockfish (Sebastes Inermis) Muscle. J. Agri. Food Chem. 1993, 41, 1208–1212. DOI: 10.1021/jf00032a008.
- Tomioka, K.; Endo, K. Purification of 5ʹ-nucleotidase from Carp Muscle. Bull. Japan. Soc. Sci. Fish. 1984, 50, 1077–1081. DOI: 10.2331/suisan.50.1077.
- Li, Q.; Li, D. P.; Qin, N.; Hong, H.; Luo, Y. K. Comparative Studies of Quality Changes in White and Dark Muscles from Common Carp (Cyprinus Carpio) during Refrigerated (4°C) Storage. Int. J. Food Sci. Tech. 2016, 51, 1130–1139. DOI: 10.1111/ijfs.13090.
- Maeda, T.; Yuki, A.; Sakurai, H.; Watanabe, K.; Itoh, N.; Inui, E.; Seike, K.; Mizukami, Y.; Fukuda, Y.; Harada, K. Alcohol Brine Freezing of Japanese Horse Mackerel (Trachurus Japonicas) for Raw Consumption. Trans. of the JSRAE. 2007, 24, 323–330.
- Veciana-Nogues, M. T.; Izquierdo-Pulido, M.; Vidal-Carou, M. C. Determination of ATP Related Compounds in Fresh and Canned Tuna Fish by HPLC. Food Chem. 1997, 59(3), 467–472. DOI: 10.1016/S0308-8146(96)00243-9.
- Kunimoto, M.; Kaminishi, Y.; Minami, K.; Hatano, M. Lipase and Phospholipase Production by Aspergillus Repens Utilized in Molding of ‘Katsuobushi’ Processing. Fish. Sci.1996, 62, 594–599.
- Delay, E. R.; Weaver, B.; Lane, D. R.; Kondoh, T. Dried Bonito Dashi: Contributions of Mineral Salts and Organic Acids to the Taste of Dashi. Physiol. Behav. 2019, 199, 127–136. DOI: 10.1016/j.physbeh.2018.11.016.
- Tomioka, K.; Endo, K. Effect of Sodium Chloride and Glycerine on Activities of Enzymes Docomposing 5ʹ-inosinic Acid in Fish Muscle. Bull. Japan. Soc. Sci. Fish. 1988, 54, 1947–1951. DOI: 10.2331/suisan.54.1947.
- Seki, H.; Osako, K.; Hamada-Sato, N. Effect of Bitterns and Constituent Salts on the Inosinate Phosphatase Activity in Horse Mackerel Muscle. Int. Food Res. J. 2014, 21, 2215–2220.
- Seki, H.; Hamada-Sato, N. Effect of Various Salts on Inosinic Acid-degrading Enzyme Activity in White and Dark Muscle of the Pacific Saury. Fish. Sci. 2015, 81(2), 365–371. DOI: 10.1007/s12562-014-0840-9.
- Yada, O.; Tsuchimoto, M.; Wang, Q.; Apablaza, P. A. G.; Jabarsyah, A.; Tachibana, K. Influence of Interposition of Pink Muscle Fiber into Dorsal Ordinary Muscle on 5ʹ-IMP Degrading Actively in Various Fish Species. Fish. Sci. 2001, 67, 948–955. DOI: 10.1046/j.1444-2906.2001.00321.x.