
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
Phenolic compounds and carotenoids profile of Amazonian Eryngium foetidum leaves and the quenching ability of their hydrophilic extract against singlet oxygen (1O2) were determined. Chlorogenic acid (4327 μg/g, dry basis, d.b.) was the major phenolic compound in the leaves at very high concentrations, while lutein (205 μg/g, d.b.) and β-carotene (161 μg/g, d.b.) were the major carotenoids. The extract of E. foetidum leaves was able to scavenge DPPH● (91.6% at 5 mg/mL), ABTS●+ (15.77 μM Trolox equivalent/g extract) and it exhibited high efficiency to protect tryptophan against 1O2, with IC50 at 343 μg/mL and 78% of protection at the highest tested concentration (625 μg/mL). Therefore, E. foetidum leaves can be exploited as an accessible natural source of bioactive compounds with antioxidant properties to be used by the food or pharmaceutical industries.
Introduction
Most of the natural vegetal resources found in Amazonia are exceptionally rich in bioactive compounds with antioxidant properties.[Citation1–Citation3] Given this scenario, E. foetidum, a green leafy native of the Amazon region and Central America, is known as “chicory” in the Amazonian States of Brazil, has been included among the main non-conventional cultivated vegetables[Citation4] and it has attracting the attention of researchers for its versatility as both phytotherapeutic plant and spice herb in Amazonian dishes.[Citation5,Citation6]
The chemical evaluation of E. foetidum leaves indicated a high concentration of phenolic compounds (total tannins and total flavonoids, as determined by spectrophotometry), presence of saponins, carotenoids, ascorbic acid, various triterpenoids, as well as antioxidant and anti-inflammatory activities.[Citation5–Citation7] However, as far as we are concerned, there is no data in the literature describing the composition of individual phenolic compounds and carotenoids from E. foetidum leaves.
The importance of describing the composition of bioactive compounds in vegetables has been widely reported, due to the association between their ingestion and the decrease of the risk of developing chronic degenerative diseases, such as arteriosclerosis, cataracts, macular degeneration, cancer and others.[Citation8,Citation9]
Bioactive compounds, such as phenolic compounds and carotenoids, play important role in human health due to modulatory effects on physiological or cellular activities. As examples, these compounds were reported to inhibit the induction of several proinflammatory cytokines and the impairment of endogenous antioxidant enzymatic systems during inflammation,[Citation10] and they are frequently associated with changes in the expression of genes implicated in cancer development, transcription, apoptosis, xenobiotic metabolism, inflammation, redox process, among others.[Citation11] Furthermore, one of the main biological actions of these compounds is related to their antioxidant capacity to scavenge reactive species, such as reactive oxygen (ROS) and nitrogen (RNS) species. ROS and RNS exert normal physiological functions in the human organism; however, the overproduction of these species can damage cellular components, such as proteins, lipids and DNA, inducing the increased development of chronic degenerative diseases and premature aging.[Citation9]
Considering that E. foetidum leaves are widely used in traditional dishes in the Amazonia, the knowledge concerning their bioactive compound profiles and the associated antioxidant capacity are of paramount importance to support any claimed potential health benefits. Therefore, this study was designed to carry out the identification and quantification of phenolic compounds and carotenoids, by HPLC-DAD-MS/MS, in E. foetidum leaves cultivated in the Amazonia, as well as to evaluate the in vitro antioxidant potential of their hydrophilic extract against singlet oxygen (1O2) and DPPH (2,2-diphenyl-1-picrylhydrazyl) and ABTS [2,2ʹ-azino-bis-(3-ethylbenzothiazoline-6-sulfonic acid] free radicals.
Material and methods
Chemicals
Quercetin, all-trans-β-carotene, gallic acid, chlorogenic acid, ethanol, methanol, methyl tert-butyl ether (MTBE), 2,2-diphenyl-1-picrylhydrazyl (DPPH), and 2,2ʹ-azino-bis-(3-ethylbenzothiazoline-6-sulfonic acid (ABTS), trolox (6-hydroxy-2,5,7,8-tetramethylchroman-2-carboxylic acid), potassium persulfate, methylene blue (MB), L-tryptophan, acetonitrile, formic acid, and all other analytical grade salts and solvents were purchased from Sigma-Aldrich (St. Louis, USA). Ultrapure water was obtained from the Pro arium® system (Sartorius, Germany). For chromatographic analysis, samples and solvents were filtered using 0.22 and 0.45 μm Millipore membranes, respectively (Billerica, MA, USA).
Samples
Fifty plants were collected in ten rows of a producing property in Santa Izabel, Pará State, Brazil (Latitude: −1.29938, Longitude: −48.161, 1°17ʹ58”South, 48°9ʹ40”West), in July 2017. The E. foetidum leaves (approximately 500 g) were washed with distilled water and dried at 25°C/2 h. After that, part of the material was used to the proximate composition characterization and another one was freeze-dried (Liotop, L101, São Paulo, Brazil) at −55°C, in a continuous vacuum for 24 h and used for the bioactive compound determination and the assessment of the in vitro antioxidant capacity. The freeze-dried leaves were grinded, vacuum packed in plastic bags and stored under free-light conditions at −18°C until analysis.
Proximate composition
Analysis of moisture, ashes, lipids and protein (conversion factor of 5.75 of total nitrogen to total protein for plants) were determined according to AOAC.[Citation12] Carbohydrate were calculated by difference [Total carbohydrates = 100–(%moisture+% ashes+% total proteins+% total lipids)]. Total energetic value was calculated according to the specific Atwater conversion fator for vegetables, as follows: total energetic value (kcal/100 g) = (protein×2.44 kcal/g)+(lipid×8.37 kcal/g)+(total carbohydrate×3.57 kcal/g).[Citation13] The experiments were carried out in triplicate and expressed in g/100 g (%) (fresh weight, f.w.).
Bioactive compounds and antioxidant capacity
Equipment
The identification of carotenoids and phenolic compounds in the samples was carried out in a Shimadzu HPLC (Prominence UFLC model, Kyoto, Japan) equipped with a binary pump (LC-20AD), a degasser unit (DGU-20A3R), an automatic injector (SIL-20AHT), an oven (CTO-20A), a DAD detector (SPD-M20A) and connected in series to a MS from Bruker Daltonics (Amazon speed ETD, Bremen, Germany) with an ion-trap as the m/z analyzer and atmospheric pressure chemical ionization (APCI) and electrospray (ESI) and as the ionization sources for carotenoids and phenolic compounds, respectively. The quantification of phenolic compounds and carotenoids was carried out in an Agilent HPLC (Agilent 1260 Infinity model, Santa Clara, CA, USA) equipped with a quaternary pump (G1311C), an automatic injector (G7129), an oven (G1316A) and a DAD detector (G1328C).
Bioactive compounds
The carotenoids were extracted from 0.5 mg of freeze-dried leaves and the final extract was solubilized in MTBE and filtered immediately before injecting into the HPLC systems. The carotenoids were separated on a C30 YMC column (5 μm, 250 mm × 4.6 mm) at 29°C, with a linear gradient of methanol (solvent A) and MTBE (solvent B) at 0.9 mL/min.[Citation14] The UV-Visible spectra were recorded between 200 and 600 nm and the chromatograms were processed at 450 nm. The column eluent was directed to the APCI interface, the MS spectra were obtained after ionization in the positive ion mode, with a scan interval at m/z 100 to 800, and the MS parameters were set as described by Chisté and Mercadante.[Citation14] The carotenoids were identified according to the following combined information: elution order and retention time at C30 column, co-elution with authentic standards, UV-visible [λmax, spectral thin structure (%III/II) and cis peak intensity (%AB/AII)] and MS spectra, in comparison to the analyzed standards and data available in the literature.[Citation2,Citation3,Citation14-Citation18] The identification of cis isomers of carotenoids was carried out based on the observed decrease in %III/II values and increase in %AB/AII values (≈ 7–11% = 9-cis, ≈ 45% = 13-cis and ≈ 56% = 15-cis carotenoid).[Citation17]
The carotenoids were quantified using β-carotene as the external standard at concentrations varying from 3.12 to 100 μg/mL [R2 = 0.99, limit of detection (LOD) = 0.97 μg/mL and limit of quantification (LOQ) = 2.96 μg/mL and lutein [R2 = 0.98, LOD = 0.20 μg/mL and LOQ = 0.62 μg/mL]. The LOD and LOQ were calculated using the parameters of the analytical curves (standard deviation and slope).[Citation19] For the calculation of the vitamin A content, the NAS-IOM conversion factor was used, considering 12 μg of all-trans-β-carotene in the extract corresponding to 1 μg of retinol activity equivalent (RAE), and the activity was of 100% for all-trans-β-carotene.[Citation20] The carotenoid contents were expressed in μg/g (dry basis, d.b.), considering three independent extraction procedures (n = 3).
The phenolic compounds were extracted from the freeze-dried leaves (0.5 g) with a methanol/water solution (80:20 v/v), were separated on a C18 Synergi Hydro column (Phenomenex, 4 μm, 250 ×4.6 mm), set at 29°C. The UV-visible spectra were obtained between 200 and 600 nm, and the chromatograms were processed at 270, 320 and 360 nm. The MS spectra were obtained at m/z from 100 to 1000 after ionization in an ESI source.[Citation14] The phenolic compounds were identified based on the following data: elution order and retention times, UV-visible and MS spectra features in comparison with authentic standards (data not shown) and analyzed under the same conditions and data available in the literature.[Citation1,Citation8,Citation14] Quantitation was carried out by external standards with six-point analytical curves (3.12–100 μg/mL, in duplicate): quercetin (360 nm, R2 = 0.99, LOD = 0.15 μg/mL and LOQ = 0.62 μg/mL), kaempferol (360 nm, R2 = 0.99, LOD = 0.15 μg/mL and LOQ = 0.62 μg/mL) and chlorogenic acid (320 nm, R2 = 0.99, LOD = 0.15 μg/mL and LOQ = 0.62 μg/mL). The contents were expressed in μg/g of freeze-dried leaves (d.b.), considering three independent extraction procedures (n = 3).
In vitro antioxidant capacity
The extracts of E. foetidum leaves used for the antioxidant capacity assays were obtained according to the same procedure described for phenolic compounds identification.
Trolox equivalent antioxidant capacity (TEAC) assay was determined according to a procedure proposed by Re et al.,[Citation21] with modifications. Aliquots of 30 μL of E. foetidum leaf extracts reacted with ABTS●+ and the absorbance was read at 734 nm after 6 min. The antioxidant capacity was calculated using analytical curves (duplicate) of trolox at concentrations varying from 102 to 2048 μM and the results were expressed in μM Trolox equivalent (TE)/g leaves (f.w.).
The DPPH assay was carried out according to the procedure described by Cheng et al.,[Citation22] by monitoring the reaction between the E. foetidum leaf extracts and DPPH radical at 515 nm after 27 min. The antioxidant capacity was calculated according to EquationEq. 1Equation 1
Equation 1 , and the results were expressed as % of inhibition of the DPPH● (d.b.).
where, A1: absorbance before reaction (DPPH● + methanol), A0: absorbance after reaction (DPPH + extract).
The quenching ability of E. foetidum leaf extract against singlet oxygen (1O2) was carried out according to the procedure described by Siqueira et al.,[Citation23] The extracts (125 to 600 μg/mL) and quercetin standard (positive control) (0.78 to 4.75 μg/mL) were tested at four concentrations. The kinetic data obtained from the decay intensity of TRP absorbance at 219 nm were fitted to a first order reaction (EquationEq. 2)Equation 2
Equation 2 using Origin Pro 8 software (OriginLab Corporation, Northampton, MA) and the rate constants were calculated (EquationEq. 3)
Equation 3
Equation 3 . The percentage of protection of TRP that the E. foetidum extracts (EXT) or quercetin exhibited against 1O2 was calculated through EquationEq. 4
Equation 4
Equation 4 .
where Y is the absorbance of TRP; Y ∞ is the absorbance of TRP in infinite time; A is a preexisting factor; k is a pseudo-first order rate constant; x is the reaction time; t1/2 is the half-life time (min); is the observed pseudo-first order rate constant fitted to the TRP decay curve (obtained in the blank experiment); and
is the observed pseudo-first order rate constant fitted to the TRP decay curve in the presence of the E. foetidum extract (EXT) or quercetin standard.
Results and discussion
Proximate composition of Eryngium foetidum leaves
The main chemical constituents of the E. foetidum leaves showed a moisture content of ≈ 87%, similar to those reported for E. foetidum leaves cultivated in Asia[Citation6] and Alta Verapaz, in Guatemala[Citation24] with values varying from 86 to 89%. The lipid content (2.8%) and protein content (1.99%) were approximately 10 and 3 fold higher, respectively, than those reported for the leaves of E. foetidum from Andaman and Nicobar Islands.[Citation6] The values of carbohydrates were 1.5 times higher than those found for the leaves of E. foetidum from Alta Verapaz (Guatemala).[Citation24] Regarding the energetic value, the leaves of E. foetidum exhibited 52.23 (± 5.46) kcal/100 g.
Carotenoid profile of Eryngium foetidum leaves
The HPLC-DAD-APCI-MSn analysis allowed the identification and quantification of nine carotenoids. The MS/MS experiment confirmed the assignment of the protonated molecule ([M +H]+) of all carotenoids identified through the expected fragments from the polyene chain of carotenoids and functional groups, along with the UV-Vis spectra (, ). Peak 1 was tentatively identified as a mix of all-trans-violaxanthin + 9-cis-neoxanthin due to the presence of a protonated molecule ([M +H]+) at m/z 601 and fragments at m/z 583, 565, and 547 corresponding to neutral losses of one, two and three OH groups (−18 u), respectively, along with a fragment at m/z 509, showing the neutral loss of a toluene moiety ([M +H-92]+). In addition, other fragments were detected at m/z 393, resulting from the cleavage of the allylic double bond to the allenic carbon (9-cis-neoxanthin), as well as the fragment at m/z 221 corresponding to the presence of a β-ring epoxy substituent with a OH group.[Citation15,Citation17,Citation18] This same carotenoid mixture was previously reported in Brazilian loquat.[Citation16]
Table 1. Chromatographic, UV-visible, mass spectroscopy characteristics and content of carotenoids from Eryngium foetidum leaves, obtained by HPLC-DAD-APCI-MS/MS.
Figure 1. HPLC-DAD chromatogram of the carotenoids of Amazonian Eryngium foetidum L. leaves. Peak identification: 1 = Mix of all-trans-violaxanthin+9-cis-neoxanthin; 2 = all-trans-Luteoxanthin; 3 = Mix of cis-lutein + epoxy-carotenoid; 4 = all-trans-Lutein; 5 = all-trans-Zeaxanthin; 6 = 9-cis-Lutein; 7 = 13-cis-β-Carotene; 8 = all-trans-β-Carotene; 9 = 9-cis-β-Carotene.
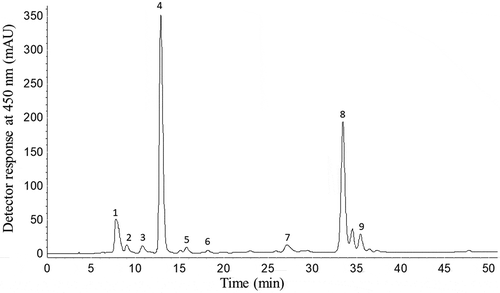
Peak 2 was identified as all-trans-luteoxanthin due to the λmax at 421 nm and the high %III/II value (100%), which are characteristic of this epoxycarotenoid, and also due to the [M +H]+ at m/z 601, and the fragment at m/z 583 [M +H-18]+, as well as the m/z 221, similar to that reported for Amazonian fruits.[Citation18]
Peak 3 was identified as a mixture of cis-lutein + epoxy-carotenoid because two protonated molecules were detected, one at m/z 585 and another at m/z 569. The m/z 585 showed fragments at m/z 567, 549 and 531, corresponding to consecutive losses of one, two and three hydroxyl groups, respectively, at m/z 493 [M +H-92]+ corresponding to the neutral loss of toluene, at m/z 475 [M +H-18-92] + and at m/z 221. The [M +H]+ at m/z 569 showed the following characteristic lutein fragments: m/z 551 and m/z 533 due to the removal of one and two water molecules, respectively, together with the fragment at m/z 477, corresponding to the loss of toluene ([M +H-92]+), m/z 463 [M +H-106]+ corresponding to loss of a xylene group, as well as fragment at m/z 459 [M +H-92], resulting from the loss of toluene and one water molecule. These fragmentation patterns were also reported in Couepia bracteosa fruit[Citation2,Citation3] and in Dovyalis and tamarillo fruits.[Citation17]
Peaks 4 and 5 were tentatively identified as all-trans-lutein and all-trans-zeaxanthin, respectively. Both the compounds are isomers and, therefore, present the same [M +H]+ at m/z 569. Lutein has a β-ring and a ε-ring and zeaxanthin two β-rings. Thus, one of the OH groups of lutein are allylic to the double bond in the ε-ring and not conjugated to the polyene chain, resulting in ten conjugated double bonds (c.d.w.). On the other hand, zeaxanthin the two β-rings double bonds are conjugated to the polyene chain and, consequently, the chromophore shows 11 c.d.w. Based on these characteristics, it is possible to identify these two compounds by the UV-visible and MS spectra features. As expected, λmax values of lutein were lower than those of zeaxanthin, the MS spectrum of all-trans-zeaxanthin showed a more intense [M +H]+ at m/z 569 compared to the m/z 551 [M +H-18]+ fragment, while the opposite was observed for lutein, as previously reported.[Citation14,Citation17,Citation18]
Peak 6 presented the same MS spectra features observed to all-trans-lutein and was identified as 9-cis-lutein, as it UV-visible spectra was similar to the all-trans-carotenoid, but taking into account the presence of the cis peak at 332 nm. Peaks 7, 8 and 9 showed the same [M +H]+ at m/z 537 with a characteristic neutral loss of toluene at m/z 444 [M-92]+. These peaks were identified as 13-cis-β-carotene, all-trans-β-carotene and 9-cis-β-carotene, respectively, based on their UV-visible characteristic differentiation as the increased %AB/AII values (≈ 7–11% = 9-cis; ≈ 45% = 13-cis-carotenoid). Peaks 7 and 9 were characterized as cis-isomers considering the presence of the cis peak at the 336–337 nm. The assignment of cis-isomers also considered that the fine spectral structure (%III/II) decreased and the intensity of the cis peak (%AB/AII) increased as the cis-double bond is approaching the center of the molecule.[Citation2] The identification of all-trans-β-carotene was positively confirmed by co-elution and comparison of the UV-Visible and MS spectra with an authentic standard.
The major carotenoids were all-trans-lutein (37%), and all-trans-β-carotene (29%), the other seven carotenoids represented approximately 25% of the total sum. The total sum of the carotenoids identified from the leaves of E. foetidum () was which was high when compared to other plants, such as Brassica oleracea, broccoli (B. oleracea L. var. Italica) (222.8–367.6 μg/g d.b.) and cauliflower (B. oleracea L. var. Botrytis) (36.5–146.8 μg/g d.b.).[Citation25] The vitamin A value found in E. foetidum leaves was 16.76 μg RAE/g (d.b.). For vitamin A activity, it should be remembered that a carotenoid must have at least one unsubstituted β-ionone ring attached to the polyene chain with at least eleven carbons. According to these structural requirements, among the nine identified carotenoids, only β-carotene (100% conversion) and its isomers (50% conversion) showed vitamin A activity. Therefore, the main contribution to the total vitamin A value was from β-carotene (86.5%).
Scientific studies have shown a positive correlation between high fruit and vegetable intake, especially rich in lutein and zeaxanthin, with a reduced risk of macular degeneration.[Citation26,Citation27] Other benefits are also associated with the ingestion of these pigments, such as reduction of the risk of cataract, protective effect against atherosclerosis, prevention of the onset and progression of some types of cancer and other chronic degenerative diseases.[Citation28] Lutein and zeaxanthin contents reported here were higher than those reported previously for India mustard (lutein; 53.8 ± 4.2; zeaxanthin; 0.8 ± 0.1), brocoli (lutein; 1.4 ± 0.2; zeaxanthin; not detected), spinach (lutein; 43.7 ± 3.8; zeaxanthin; 0.7 ± 0.1), watercress (lutein; 42.8 ± 4.1; zeaxanthin; 0.4 ± 0.1), endive (lutein; 34.2 ± 4.0; zeaxanthin; 0.4 ± 0.1), lettuce (lutein; 2.0 ± 0.1; zeaxanthin; 0.1 ± 0.1) and cabbage (lutein; 2.5 ± 0.1; zeaxanthin; 0.1 ± 0.1).[Citation29] Importantly, the values indicated by Britton and Khachik,[Citation30] to classify individual carotenoid levels as high (500–2000 μg/100 g or 5–20 μg/g) and very high (> 2000 μg/100 g or > 20 μg/g) suggested that E. foetidum leaves can be considered as a very high source of lutein and β-carotene and a high source of zeaxanthin. Furthermore, based on the daily intake ranges of carotenoids reported by Meléndez-Martínez,[Citation31] the intake of 30 g of E. foetidum fresh leaves correspond to dietary intake of 7% and 156% of β-carotene and lutein, respectively.
Phenolic compounds profile of Eryngium foetidum leaves
Regarding the profile of phenolic compounds, the HPLC-DAD-ESI-MSn allowed the identification and quantification of six phenolic compounds (, ). Peak 1 was identified as chlorogenic acid since it exhibited a deprotonated molecule ([M-H]−) at m/z 353 and an intense fragment at m/z 191 ([M-H-162]−) corresponding to the loss of a caffeoyl moiety releasing quinic acid. Chlorogenic acid was the main phenolic compound found in leaf extract of E. foetidum accounting for 43.28% of the total sum, and was positively confirmed based on the same UV-visible features and MS fragmentation pattern observed for the authentic standard. The content of the chlorogenic acid found in this study () was 9 times higher than that found in artichoke leaves (Cynara cardunculus var. Scolymus Hayek L.) (481 μg/g, d.b.).[Citation32]
Table 2. Chromatographic, spectroscopic characteristics and content of phenolic compounds from Eryngium foetidum leaves, obtained by HPLC-DAD-ESI-MSn.
Figure 2. HPLC-DAD chromatogram of phenolic compounds of Eryngium foetidum L. leaves. Chromatographic conditions: see text. Peak characterization is given in . Peak identification: 1: Chlorogenic acid; 2: Feruloylquinic acid; 3: Quercetin glucuronide; 4: Luteolin hexoside; 5: Luteolin glucuronide; 6: Ferulic acid derivative.
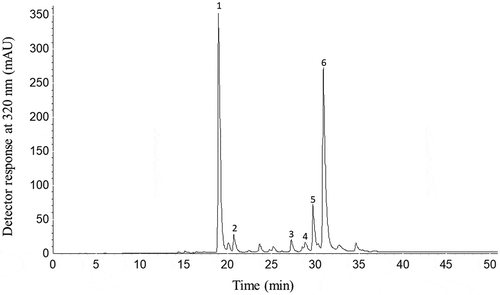
Peak 2 was tentatively identified as feruloylquinic acid, with [M-H]− at m/z 367 and a MS2 fragment at m/z 193 [M-H-174]−, corresponding to ferulic acid after losing the quinic acid attached to the molecule. This fragmentation pattern corresponds to those previously described for feruloylquinic acid found in artichoke extracts.[Citation33]
Peak 3 was identified as quercetin-glucuronide with [M-H]− at m/z 477 and MS2 fragment at m/z 301 [M-H-176]− showing the quercetin molecule after the removal of a glucuronic acid unit. The fragments of quercetin (m/z 301) in the MS3 experiment was the same as reported in studies where quercetin-glucuronide was identified.[Citation1,Citation8]
Peaks 4 and 5 showed [M-H]− at m/z 447 and 461, respectively, and were identified as luteolin derivatives according to their UV-Visible and MS spectra and data available in the literature.[Citation8,Citation33] Peak 4 presented an intense fragment at m/z 285 ([M-H-162]−) in the MS2 spectrum, corresponding to the loss of an hexose unit and was identified as a luteolin hexoside. Peak 5 was identified as luteolin glucuronide due to the same fragment at m/z 285 ([M-H-176]−) in the MS2 spectrum, which corresponds to the loss of one glucuronic acid unit. Both the peaks showed the same fragmentation pattern for luteolin (m/z 285) in the MS3 spectra, as compared to previous data.[Citation8,Citation33] The intake of luteolin and apigenin (flavones) in the diet is usually lower than that of flavonols, such as quercetin and kaempferol, since they occur in significant concentrations in few foods. A luteolin was identified in artichoke leaves with a concentration ranging from 53 (± 1) a 69 (± 2) μg/g d.b.,[Citation32] whose values are lower than those reported in this study for leaves of E. foetidum.
Peak 6 exhibited the [M-H]− at m/z 359 with an MS2 fragment at m/z 165 ([M-H-194]−) corresponding to the loss of ferulic acid, being tentatively identified as a ferulic acid derivative. The MS characteristics of this compound were similar to those previously reported for ferulic acid derivatives in strawberry (Fragaria ananassa).[Citation34]
The content of phenolic compounds found in this study was higher than previously reported for leaves of E. foetidum from the Andaman and Nicobar Islands (India) whose value of 283.10 μg/g (d.b.) (35 times less) was determined by spectrophotometry.[Citation6] When compared to other plants, such as artichoke leaves (Cynara cardunculus var. Scolymus Hayek L.), the phenolic content of E. foetidum leaves was approximately 7 times higher (1387 μg/g, d.b.).[Citation32] The average consumption of total phenolic compounds and, more specifically phenolic acids, by the Brazilian population was reported to be 460.15 and 314 mg/day, respectively.[Citation35] Based on this information, the ingestion of 30 g of E. foetidum leaves can contribute with ≈10.4% and 8.5% of the total phenolic compounds and phenolic acids, respectively.
Antioxidant capacity of Eryngium foetidum leaf extract
The methanolic extract of E. foetidum were able to scavenge both ABST and DPPH radicals. However, the leaf extract of E. foetidum showed low efficiency in removing ABTS●+ (15.77 ± 0.92 μM Trolox/g, FW) when compared to Arrabidaea chica leaves (86.81 TE/g, FW).[Citation23] This is the first time that the scavenging capacity of E. foetidum leaf extract against ABTS●+ was reported. The methanolic extract of E. foetidum leaves may also be able to capture DPPH● with inhibition percentage of 91.60 ± 0.45% at the tested concentration of 0.06 μg/g, a value similar to that found for the extracts of E. foetidum leaves previously reported by Singh et al.,[Citation7] (92.7%), but superior to the authors of a previous study (73.93%).[Citation6]
ABTS●+ and DPPH● are stable, synthetic and non-biological free radicals; however, due to their chemical stability, sensitivity and operational ease in antioxidant capacity protocols, they are widely used in determining the in vitro antioxidant potential of isolated compounds or plant extracts from food samples.[Citation36]
Regarding the 1O2-quenching capacity, the extract of E. foetidum leaves, at the studied concentrations, was efficient to inhibit tryptophan degradation, as induced by the photosensitization reaction in the presence of methylene blue, showing excellent fitting to first order reaction, with a high determination coefficient (R2 = 0.99) (). According to Figure 3b, 1O2 was inhibited in a concentration-dependent manner and the extract of E. foetidum leaves presented an IC50 at 343.32 μg/mL with high percentage of protection (77.50%) at the highest tested concentration (625 μg/mL). Quercetin (positive control) showed higher 1O2-quenching ability (IC50 of 1.9 μg/mL) than the extracts of E. foetidum leaves. The extract of E. foetidum leaves was more effective against 1O2 than fruit extracts of Terminalia chebula (IC50 = 424.50 μg/ml) and Emblica officinalis (IC50 of 490.42 μg/ml).[Citation37] However, it was less efficient than the artichoke extracts (IC50 = 20.5–30.1 μg/ml)[Citation33] and for Caryocar villosum fruit extracts (IC50 = 156 μg/ml).[Citation38]
Figure 3. (a) Decay of the intensity of the absorbance of tryptophan in the presence of the methylene blue and extract of the Eryngium foetidum L. leaves and (b) percentage of inhibition of the1O2 as a function of the concentration of the extract of the Eryngium foetidum L. leaves. The experiments were carried out in four concentrations and n = 3 (Mean ± standard deviation).
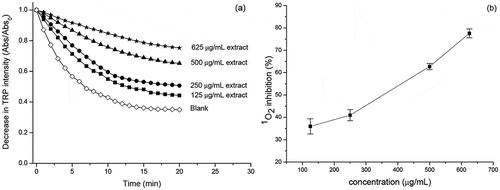
1O2 is an important ROS with high reactivity that can be found both in human body and in food systems. The formation of 1O2 can generate other physiological reactive species, such as alkyl (R•), alkoxyl (RO•), organic hydroperoxides (ROOH), superoxide anion (O2•-), hydrogen peroxide (H2O2), hydroxyl radical (•OH) and peroxynitrite (ONOO−), and these ROS are often responsible for several biological deleterious effects.[Citation39,Citation40] Therefore, the investigation of plant extracts, such as E. foetidum, with high contents of bioactive compounds and high ability to inhibit 1O2 is highly desirable to support and stimulate deeper studies aiming the inhibition of the oxidation of biomolecules present in both the human body and food systems.
The high antioxidant potential of the E. foetidum leaf extract may be mainly attributed to the high contents of phenolic compounds in the hydrophilic extract (), which corroborates to the fact that the antioxidant action of phenolic compounds from plants is associated to the inhibition of the oxidizing effect of reactive species by delaying chain reactions and/or scavenging free radicals.[Citation9,Citation33] Chlorogenic acid was the major phenolic compound found in the extract of E. foetidum leaves, and epidemiological studies showed a positive association between the consumption of foods containing chlorogenic acid and health benefits.[Citation41–Citation43] These combined information makes Amazonian E. foetidum leaves an interesting plant source for research of food supplementation aiming increasing human health.
Importantly, the biological properties of chlorogenic acid, or any bioactive compound, depend on its bioavailability in the gut, which can be affected by several factors, such as the bioaccessibility of each compound, bioactive compounds–gut microbiota interactions, different mechanisms of absorption of water-soluble and lipid-soluble molecules, interactions with other nutrients, among others.[Citation44] Pharmacokinetic studies involving chlorogenic acid and its metabolites suggest that they bound to human serum albumin and/or may be sequestered in tissues to exhibit its biological activity.[Citation43]
The relevance of researches on bioactive constituents from plants is fully justified by the association that fruits, vegetables, and food legumes are therapeutic agents useful for treating a wide range of human disorders.[Citation45] Regarding the application on food industries, the safety concerns regarding some synthetic antioxidants could be relieved with the use of natural plant extracts with high contents of bioactive compounds at lower concentrations. Thus, E. foetidum leaves can be seen as a dietary source of natural antioxidants with potential health benefits whose consumption should be stimulated.
Conclusion
All-trans-lutein and all-trans-β-carotene were the major carotenoids in E. foetidum leaves, while chlorogenic acid and a ferulic acid derivative were the major phenolic compounds. Amazonian E. foetidum leaves, besides its traditional use in cooking and folk medicine, may represent a valuable source of bioactive compounds with antioxidant properties, with undeniable nutraceutical value and great potential to be explored and used in the food, cosmetic and pharmaceutical industries. Furthermore, future systematic studies based on cellular antioxidant capacity assays should be applied to understand the protective effects of the bioactive compounds of this plant species on different oxidative/nitrosative stress pathways.
Acknowledgments
The authors acknowledge CNPq (Conselho Nacional de Desenvolvimento Cientifico e Tecnológico, Brazil, Project 428403/2016-6), FAPESPA (Fundação Amazônia de Amparo a Estudos e Pesquisas, Belém, PA, Brazil, grant number ICAAF 103/2014) and Federal University of Pará (UFPA) through PROPESP/UFPA for the financial support. D. S. Leitão was supported by CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, - Brasilia, DF, Brazil – Finance Code 001). Adriana Z. Mercadante thanks FAPESP (Fundação de Amparo à Pesquisa do Estado de São Paulo, São Paulo, Brazil, Projects 2013/07914-8 and 2018/23752-1) for the financial support.
Additional information
Funding
References
- Ribeiro, A. B.; Chisté, R. C.; Freitas, M.; Da Silva, A. F.; Visentainer, J. V.; Fernandes, E. Psidium Cattleianum Fruit Extracts are Efficient in Vitro Scavengers of Physiologically Relevant Reactive Oxygen and Nitrogen Species. J. Agric. Food Chem. 2014, 165, 140–148. DOI: 10.1016/j.foodchem.2014.05.079.
- Berto, A.; Ribeiro, A. B.; de Souza, N. E.; Fernandes, E.; Chisté, R. C. Bioactive Compounds and Scavenging Capacity of Pulp, Peel and Seed Extracts of the Amazonian Fruit Quararibea Cordata against ROS and RNS. Food Res. Int. 2015a, 77, 236–243. DOI: 10.1016/j.foodres.2015.06.018.
- Berto, A.; Ribeiro, A. B.; Sentandreu, E.; de Souza, N. E.; Mercadante, A. Z.; Chisté, R. C.; Fernandes, E. The Seed of the Amazonian Fruit Couepia Bracteosa Exhibits Higher Scavenging Capacity against ROS and RNS than Its Shell and Pulp Extracts. Food Funct. 2015b, 6(9), 3081–3090. DOI: 10.1039/c5fo00722d.
- MAPA – Ministério da Agricultura, Pecuária e Abastecimento. 2010. Manual de hortaliças não-convencionais. MAPA/ACS: Brasília, p. 92. http://www.agricultura.gov.br/arq_editor/file/vegetal/Qualidade/Qualidade%20dos%20Alimentos/manual%20 hortali%C3%A7as_WEB_F.pdf. (accessed Feb 10, 2018).
- García, M. D.; Saenz, M. T.; Gomez, M. A.; Fernandez, M. A. Topical Antiinflammatory Activity of Phytosterols Isolated from Eryngium Foetidum on Chronic and Acute Inflammation Models. Phytotherapy Research. Int. J. Devoted Pharmacol. Toxicol. Eval. Nat. Product Derivatives. 1999, 13(1), 78–80.
- Singh, S.; Singh, D. R.; Salim, K. M.; Srivastava, A.; Singh, L. B.; Srivastava, R. C. Estimation of Proximate Composition, Micronutrients and Phytochemical Compounds in Traditional Vegetables from Andaman and Nicobar Islands. Int. J. Food Sci. Nutr. 2011, 62(7), 765–773. DOI: 10.3109/09637486.2011.585961.
- Singh, S.; Singh, D. R.; Banu, S.; Salim, K. M. Determination of Bioactives and Antioxidant Activity in Eryngium Foetidum L.: A Traditional Culinary and Medicinal Herb. Proc. National Acad. Sci. India Sec. B - Biolog. Sci. 2013, 83(3), 453–460. DOI: 10.1007/s40011-012-0141-y.
- Pereira, C.; Barros, L.; Carvalho, A. M.; Santos-Buelga, C.; Ferreira, I. C. F. R. Infusions of Artichoke and Milk Thistle Represent a Good Source of Phenolic Acids and Flavonoids. Food Funct. 2014, 6(1), 56–62. DOI: 10.1039/c4fo00834k.
- Rojas, J.; Buitrago, A. Antioxidant Activity of Phenolic Compounds Biosynthesized by Plants and Its Relationship with Prevention of Neurodegenerative Diseases. Bio. Compounds. 2019, 3–31. DOI: 10.1016/b978-0-12-814774-0.00001-3.
- Ribeiro, D.; Freitas, M.; Lima, J. L. F. C.; Fernandes, E. Proinflammatory Pathways: The Modulation by Flavonoids. Med. Res. Rev. 2015, 35(5), 1–57. DOI: 10.1002/med.21347.
- Pokimica, B.; García-Conesa, M. T. Critical Evaluation of Gene Expression Changes in Human Tissues in Response to Supplementation with Dietary Bioactive Compounds: Moving Towards Better-quality Studies. Nutrients. 2018, 10, 807. DOI: 10.3390/nu10070807.
- AOAC (Association of Analytical Chemists). Official Methods of Analysis, 17th ed.; AOAC International: Gaithersburg, MD, USA, 2000.
- FAO/WHO. 2003. Food Energy—methods of Analysis and Conversion Factors. Report of a Technical Workshop. FAO Food and Nutrition Paper No. 77 Food and Agriculture Organization of the United Nations, Rome http://www.fao.org/3/Y5022E/y5022e00 (accessed Dec 20, 2018).
- Chisté, R. C.; Mercadante, A. Z. Identification and Quantification, by HPLC-DAD-MS/MS, of Carotenoids and Phenolic Compounds from the Amazonian Fruit Caryocar Villosum. J. Agric. Food Chem. 2012, 60(23), 5884–5892. DOI: 10.1021/jf301904f.
- Faria, A. F.; De Rosso, V. V.; Mercadante, A. Z. Carotenoid Composition of Jackfruit (Artocarpus Heterophyllus), Determined by HPLC-PDA-MS/MS. Plant Foods Human Nutr. 2009a, 64(2), 108–115. DOI: 10.1007/s11130-009-0111-6.
- Faria, A. F.; Hasegawa, P. N.; Chagas, E. A.; Pio, R.; Purgatto, E.; Mercadante, A. Z. Cultivar Influence on Carotenoid Composition of Loquats from Brazil. J. Food Compost. Anal. 2009b, 22(3), 196–203. DOI: 10.1016/j.jfca.2008.10.014.
- De Rosso, V. V.; Mercadante, A. Z. HPLC-PDA-MS/MS of Anthocyanins and Carotenoids from Dovyalis and Tamarillo Fruits. J. Agric. Food Chem. 2007a, 55(22), 9135–9141. DOI: 10.1021/jf071316u.
- De Rosso, V. V.; Mercadante, A. Z. Identification and Quantification of Carotenoids, by HPLC-PDA-MS/MS, from Amazonian Fruits. J. Agric. Food Chem. 2007b, 55(13), 5062–5072. DOI: 10.1021/jf0705421.
- ICH, 2005. International Conference on Harmonization of Technical Requirements for the Registration of Pharmaceuticals for Human Use (ICH). Guidance for industry, Q2B validation of analytical procedures: Methodology. Geneva: ICH Secretariat c/o IFPMA7–10.
- NAS-IOM. Dietary Reference Intakes for Vitamin A, Vitamin K, Arsenic, Boron, Chromium, Copper, Iodine, Iron, Manganese, Molybdenum, Nickel, Silicon, Vanadium, and Zinc; National Academy Press 92: WA, 2001.
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant Activity Applying an Improved ABTS Radical Cation Decolorization Assay. Free Radical Biol. Med. 1999, 26(9–10), 1231–1237. DOI: 10.1016/S0891-5849(98)00315-3.
- Cheng, Z.; Moore, J.; Yu, L. High-throughput Relative DPPH Radical Scavenging Capacity Assay. J. Agric. Food Chem. 2006, 54(20), 7429–7436. DOI: 10.1021/jf0611668.
- Siqueira, F. C.; Leitão, D. S. T. C.; Mercadante, A. Z.; Chisté, R. C.; Lopes, A. Profile of Phenolic Compounds and Carotenoids of Arrabidaea Chica Leaves and the in Vitro Singlet Oxygen Quenching Capacity of Their Hydrophilic Extract. Food Res. Int. 2019, 126, 108597. DOI: 10.1016/j.foodres.2019.108597.
- Booth, S.; Bressani, R.; Johns, T. Nutrient Content of Selected Indigenous Leafy Vegetables Consumed by the Kekchi People of Alta Verapaz, Guatemala. J. Food Compost. Anal. 1992, 5(1), 25–34. DOI: 10.1016/0889-1575(92)90005-5.
- Guzman, I.; Yousef, G. G.; Brown, A. F. Simultaneous Extraction and Quantitation of Carotenoids, Chlorophylls, and Tocopherols in Brassica Vegetables. J. Agric. Food Chem. 2012, 60(29), 7238–7244. DOI: 10.1021/jf302475d.
- Eisenhauer, B.; Natoli, S.; Liew, G.; Flood, V. M. Lutein and Zeaxanthin — Food Sources, Bioavailability and Dietary Variety in Age‐related Macular Degeneration Protection. Nutrients. 2017, 9(2). DOI: 10.3390/nu9020120.
- Liu, R.; Wang, T.; Zhang, B.; Qin, L.; Wu, C.; Li, Q.; Ma, L. Lutein and Zeaxanthin Supplementation and Association with Visual Function in Age-related Macular Degeneration. Invest. Ophthalmol. Visual Sci. 2015, 56(1), 252–258. DOI: 10.1167/iovs.14-15553.
- Calvo, M. M.;. Lutein: A Valuable Ingredient of Fruit and Vegetables. Crit. Rev. Food Sci. Nutr. 2005, 45(7–8), 671–696. DOI: 10.1080/10408690590957034.
- Murillo, E.; Meléndez-Martínez, A. J.; Portugal, F. Screening of Vegetables and Fruits from Panama for Rich Sources of Lutein and Zeaxanthin. J. Agric. Food Chem. 2010, 122(1), 167–172. DOI: 10.1016/j.foodchem.2010.02.034.
- Britton, G.; Khachik, F. Carotenoids in Food. In Carotenoids: Nutrition and Health; Britton, G., Liaaen-Jensen, S., Pfander, H., Eds.; Birkhauser: Basel, 2009; Vol. 5, pp 45–66.
- Meléndez‐Martínez, A. J.;. An Overview of Carotenoids, Apocarotenoids and Vitamin A in Agro‐Food, Nutrition, Health and Disease. Mol. Nutr. Food Res. 2019, 63(15), 1801045. DOI: 10.1002/mnfr.201801045.
- Nouraei, S.; Rahimmalek, M.; Saeidi, G. Variation in Polyphenolic Composition, Antioxidants and Physiological Characteristics of Globe Artichoke (Cynara Cardunculus Var. Scolymus Hayek L.) As Affected by Drought Stress. Sci. Hortic. 2018, 233(October 2017), 378–385. DOI: 10.1016/j.scienta.2017.12.060.
- Pistón, M.; Machado, I.; Branco, C. S.; Cesio, V.; Heinzen, H.; Ribeiro, D.; Freitas, M. Infusion, Decoction and Hydroalcoholic Extracts of Leaves from Artichoke (Cynara Cardunculus L. Subsp. Cardunculus) are Effective Scavengers of Physiologically Relevant ROS and RNS. Food Res. Int. 2014, 64, 150–156. DOI: 10.1016/j.foodres.2014.05.078.
- Aaby, K.; Ekeberg, D.; Skrede, G. Characterization of Phenolic Compounds in Strawberry (Fragaria X Ananassa) Fruits by Different HPLC Detectors and Contribution of Individual Compounds to Total Antioxidant Capacity. J. Agric. Food Chem. 2007, 55(11), 4395–4406. DOI: 10.1021/jf0702592.
- Corrêa, V. G.; Tureck, C.; Locateli, G.; Peralta, R. M.; Koehnlein, E. A. Estimate of Consumption of Phenolic Compounds by Brazilian Population. Revista De Nutrição. 2015, 28(2), 185–196. DOI: 10.1590/1415-52732015000200007.
- Huang, D.; Ou, B.; Prior, R. L. The Chemistry behind Antioxidant Capacity Assays. J. Agric. Food Chem. 2005, 53, 1841–1856. DOI: 10.1021/jf030723c.
- Hazra, B.; Sarkar, R.; Biswas, S.; Mandal, N. Comparative Study of the Antioxidant and Reactive Oxygen Species Scavenging Properties in the Extracts of the Fruits of Terminalia Chebula, Terminalia Belerica and Emblica Officinalis. BCM Complementary Altern. Med. 2010, 10, 20.
- Chisté, R. C.; Freitas, M.; Mercadante, A. Z.; Fernandes, E. The Potential of Extracts of Caryocar Villosum Pulp to Scavenge Reactive Oxygen and Nitrogen Species. J. Agric. Food Chem. 2012, 135(3), 1740–1749. DOI: 10.1016/j.foodchem.2012.06.027.
- Blázquez-Castro, A.; Breitenbach, T.; Ogilby, P. R. Singlet Oxygen and ROS in a New Light: Low-dose Subcellular Photodynamic Treatment Enhances Proliferation at the Single Cell Level. Photochem. Photobiol. Sci. 2014, 13(9), 1235–1240. DOI: 10.1039/c4pp00113c.
- Carrasco, E.; Calvo, M. I.; Blázquez-Castro, A.; Vecchio, D.; Zamarrón, A.; de Almeida, I. J. D.; Espada, J. Photoactivation of ROS Production in Situ Transiently Activates Cell Proliferation in Mouse Skin and in the Hair Follicle Stem Cell Niche Promoting Hair Growth and Wound Healing. J. Invest. Dermatol. 2015, 135(11), 2611–2622. DOI: 10.1038/jid.2015.248.
- Abu-Amsha, R.; Croft, K. D.; Puddey, I. B.; Proudfoot, J. M.; Beilin, L. J. Phenolic Content of Various Beverages Determines the Extent of Inhibition of Human Serum and Low-Density Lipoprotein Oxidationin Vitro: Identification and Mechanism of Action of Some Cinnamic Acid Derivatives from Red Wine. Clinical Sci. 1996, 91(4), 449–458. DOI: 10.1042/cs0910449.
- Natella, F.; Nardini, M.; Belelli, F.; Pignatelli, P.; Di Santo, S.; Ghiselli, A.; … Scaccini, C. Effect of Coffee Drinking on Platelets: Inhibition of Aggregation and Phenols Incorporation. Br. J. Nutr. 2008, 100(6), 1276. DOI: 10.1017/S0007114508981459.
- Clifford, M. N.; Kerimi, A.; Williamson, G. Bioavailability and Metabolism of Chlorogenic Acids (Acyl‐quinic Acids) in Humans. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1299–1352. DOI: 10.1111/1541-4337.12518.
- Rein, M. J.; Renouf, M.; Cruz-Hernandes, C.; Actis-Goretta, L.; Thakkar, S. K.; Pinto, M. S. Bioavailability of Bioactive Food Compounds: A Challenging Journey to Bioefficacy. Br. J. Clin. Pharmacol. 2012, 75(3), 588–602. DOI: 10.1111/j.1365-2125.2012.04425.x.
- Zhu, F.; Du, B.; Xu, B. Anti-inflammatory Effects of Phytochemicals from Fruits, Vegetables, and Food Legumes: A Review. Crit. Rev. Food Sci. Nutr. 2018, 58(8), 1260–1270. DOI: 10.1080/10408398.2016.1251390.