
ABSTRACT
Polymethoxyflavones (PMFs) are dietary bioactive phytochemicals, naturally present in all forms of citrus or plants. For this updated review, the specialized databases of PubMed and Web of Science were researched using keywords such as “polymethoxy flavones,” “pharmacology,” “molecular mechanisms,” “signaling pathways,” and “therapeutic applications.” The most prominent citrus PMFs with proven pharmacological effects are tangeretin, nobiletin, 5′-demethylnobiletin, tetramethyl-o-scutellarein, pentamethoxyflavone, tetramethyl-o-isoscutellarein, and sinensetine. The results of in vivo and in vitro preclinical studies of PMFs have highlighted molecular mechanisms and signaling pathways for anti-inflammatory, anti-cancer, chemopreventive, antidiabetic, anti-obesity, hepatoprotective and neuroprotective effects in neurodegenerative diseases. The results of the studies support the traditional effects of PMFs and therefore it is necessary that these natural compounds be further investigated in clinical trials. PMFs should be designed and formulated in nanoformulations that increase their bioavailability and transport them precisely to the target in order to increase their therapeutic effects.
Introduction
In the current age, dietary bioactive compounds are gaining popularity fast because of their positive and beneficial effects body in treating different diseases. [Citation1]] Flavonoids are phytochemicals recognized for their anti-inflammatory, anti-diabetic, antioxidant, anti-hypertensive properties.[Citation2] Tangerine, sweet orange, and disagreeable orange strips commonly contain citrus flavonoids. They can be polymethoxyflavones (PMFs), also known as glycosides, or O-methylated aglycones of flavones.[Citation3] The superfamily of flavonoids, which includes anthoxanthins (such as flavones and flavonols), flavanones, flavanonols, flavans, and anthocyanidins, includes polymethoxyflavones (PMFs). Citrus reticulata “Chachi” (CRC)‘s is a rich source of PMFs.[Citation4] PMFs are flavones with several methoxy (-O-CH3) groups attached to their primary benzo-pyrone structure. PMFs contain nobiletin (20.98%), tetramethyl-o-isoscutellarein (1.23%), sinensetin (0.35%), 5-demethylnobiletin (3.52%), tangeretin (33.87%), sinensetin (0.35%), and pentamethoxyflavone (1.08%) had very low aqueous permeability and solubility. However, all PMFs showed high permeability. 5-hydroxylated PMFs and 5-acetyl PMFs had the greatest permeability as compared to other PMFs compounds. Different scientific studies showed that all PMFs compounds have a high absorption rate as well as permeability and solubility.[Citation5] It is naturally present in all form of citrus fruits or plants. Its presence also found herbs Nicotiana plumbaginifolia Vivane known for the treatment of toothache, wounds, rheumatic swelling and cuts in Bangladesh.[Citation6] Similarly, it is present in the plant Ageratum conyzoides L. extensively used against antinociceptive, anti-inflammatory, and antibacterial properties.[Citation7]
The PMFs exhibited anti-inflammatory, anticarcinogenic, chemopreventive, sedative, and anti-depressant properties in both vivo and in vitro studies. The biological activities of PMF compounds were investigated in many studies. In addition, they play a nominal role in regulating the signaling cascade, protein activity, and gene transcription.[Citation7] PMFs are hydrophobic particles with less water solubility, just like oil on the body or surrounding areas. Generally, PMFs assimilated quickly during the digestive process. One investigation in rodents showed that combined use of nobiletin or tangeretin, showed that tangeretin’s bio-retention was significantly lower than nobiletin’s. The two main metabolites of tangeretin, glucuronic and aglycone metabolites, with the highest individual convergences.[Citation8]
The following three segments are known as bioavailability, which is defined as the portion of oral organization that eventually reaches the basic course: the amount of bioactive mixtures delivered into the small digestive system and becoming bio-accessible for the resulting bio-assimilation, the amount of bioactive mixtures ingested through the gut, and the amount of bioactive compounds maintained by the primary pass digestion.[Citation9,Citation10] PMFs’ weak solvency restricts their bioavailability in the framework of the human gut. While the elevated capacity to burn calories rate influences the amount of PMFs entering the human gastrointestinal tract, which ultimately compromise bioavailability and diminishes the remedial doses they can accomplish in the objective organs.[Citation11,Citation12] Chemical alteration is used to increase the bioavailability of PMFs. 5-acetyloxy-6,7,8,3,4-pentamethoxyflavone reduced triacylglycerol, whereas nanoencapsulation improved safety concerns. In previous studies, it was demonstrated that lipophilic bio-actives could be dissolved in oil transporters to increase bioavailability. Surface-active substances provided during lipid breakdown, and free unsaturated fats that contributed to the grip and absorption of nutrients through the gut cell dividers, were attributed to this effect.[Citation13] Specialists have found to utilize emulsions to work on the bioavailability of PMFs.[Citation14,Citation15]
Comparing the 2.6-overlap to the tangeretin oil suspension.[Citation16] In vivo mice concentrate additionally affirmed revealed as compared to an unformulated oil suspension. Different examinations additionally demonstrated tangeretin’s bio-viability against colorectal illness.[Citation17,Citation18] This review offers a thorough summary of traditional medicinal uses of PMFs, their chemical classification, their bioavailability, updated molecular mechanisms of action from recent preclinical pharmacological studies, and toxicological data. As a novelty, new perspectives on the nano-pharmaceutical formulations of PMFs are described. So, key data for future clinical trials and the development of new therapeutic agents are highlighted.
Search strategy
Specialised databases like Web of Science and PubMed were researched using keywords such as “polymethoxyflavones,” “pharmacology,” “molecular mechanisms,” “signaling pathways,” “therapeutic applications,” “anticancer,” “anti-inflammatory,” “antidiabetic,” neuroprotective “,” antiobesity “,” antihypertensive “, “pharmaceutical nano-formulations.” The articles written in English were selected, abstracts, letters to editor or comments were not taken into account.
Inclusion Criteria:
Investigations with natural PMFs and their derivatives.
Studies that emphasized the cellular mechanisms and signaling pathways at the target.
Studies that had animals or single/multiple cell lines as an experimental model.
Exclusion Criteria:
Studies without mentioning the content of PMFs.
Studies with flavonoids other than polymethoxylated flavones.
Paper titles which do not meet the inclusion criteria.
Research with homeopathic remedies.
Traditional uses and clinical applications of pmfs
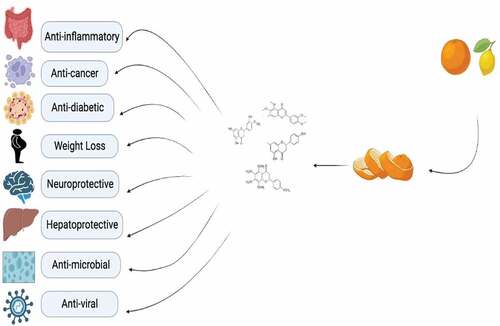
Strips, particularly matured citrus strips, which have long been utilized as a traditional Chinese remedy, are rife with PMFs to treat a variety of conditions, including ringworm, back pain, skin irritation, and diseases with anti-cancerous, anti-inflammatory, and anti-atherogenic effects.[Citation19] The health benefits of polymethoxyflavones were shown in . The mice study was the one that first revealed the anti-obesity effects of Chenpi separate. They claimed that the effects of a high-fat diet on weight, hepatic steatosis, and diabetes symptoms were all lessened following oral chenpi extract administration.[Citation20] Polymethoxyflavones (PMFs), which are substantial elements in citrus strips and have attracted noteworthy attention due to their anti-hypertensive, anti-carcinogenic, and antiatherogenic activities, have been identified by phytochemical analyses of citrus species.[Citation21] Numerous studies revealed that PMFs may show obvious antinociceptive, moderating, anticarcinogenic, malignant growth chemo preventive, narcotic, stimulant, anxiolytic, as well as benzodiazepine restricting effects in vivo or in vitro. They also exhibit astonishing oral bioavailability.[Citation22]
Figure 1. Health benefits of polymethoxyflavones.
An annual herb from Bangladesh called Nicotiana plumbaginifolia Vivane is used to cure cuts, wounds, toothaches, and rheumatic pain. Therapeutic studies revealed that the leaves of this plant had important anxiolytic and pain-relieving properties. According to a phytochemical analysis of the plant’s leaves. In addition to the labyrinth test on mice, Acetic acid-induced writhing on the hot plate, tail drenching, formalin-prompted nociception, and other nociceptive situations, the antinociceptive characteristics of PMFs from this plant have been investigated for the carrageenan-actuated paw edema test, and neuro-pharmacological impact in open field. A key component of logical and test pharmacology, along with the introduction of novel therapeutic experts, is the test concentration alongside deciphering the mechanism of activities of the trial drugs or mixes in the living framework. In this investigation, there isn’t really any concrete evidence linking narcotic, In the anxiolytic activity of the included PMFs, ATP-sensitive K+ directly interacts with the antinociceptive and benzodiazepine framework.[Citation6]
Phytochemistry of pmfs
The word “PMF” refers to flavones that have multiple methoxy groups on their fundamental benzo-pyrone skeleton, which is typically composed of 15 carbons (C6-C3-C6).[Citation23] Different derivatives of polymethoxyflavones were presented in .
Table 1. Derivatives of Polymethoxyflavones.
Anticancer activity
A key phase in the early transition of cells toward the malignant stage is unchecked cell proliferation.[Citation26] A major feature of cell growth is the G1, S, G2, and M stages of the cell cycle. G0 is the phase in which cells do not grow, G1 is the proliferating phase in which cells prepare for DNA synthesis acquis. In phase S the DNA is synthesized, in phase G2 the cell prepares for mitosis and phase M is the phase of mitosis.[Citation27,Citation28] Different stages of cancer initiation, development, and progression have been associated with PMFs.[Citation29] PMFs have a major role in arresting cell cycle on G1, S and G2 phases brings apoptosis by increasing the production of tenin and phosphatase homolog cancer suppressor and decrease the cyclin D and cdc-2- protein expressions.[Citation30] Nobiletin is one of the promising PMFs compounds which has significant chemoprotective and anti-cancer activities in vivo and vitro studies, not only nobiletin but also its derivatives include 3′-demethylnobiletin, 4′-demethylnobiletin, 3′,4′ -didemethylnobiletin 5′demethylnobiletin. Nobiletin suppressed the proliferation stage by halting the G2 and M phases of the cell cycle by increasing Bax and p53 protein and decreasing Bcl-2. It also helped in reducing Reduced Expression of E2 promoter-binding factor 1, p-Akt, cyclin D1, cyclin D2, cyclin D4, cyclin D4, p-p38, and cyclin D1 and V Phosphorylated-Jun N-terminal kinase protein results in glioma U87 and Hs683 cells have more viable cells.[Citation31] In a recent study, Nobiletin prevents glioma cells from proliferating and migrating by blocking the protein kinase pathways. The anti-cancer activities PMFs has been demonstrated in colon carcinogenesis animal model, inhibit the proliferation of the cell, induce apoptosis and reduce the formation of tumor.[Citation32] Nobiletin’s effects on colorectal cancer have also been shown by a study in which inhibited cell invasion and migration and suppressed the growth factor A and angiopoietin-2 to block angiogenesis in colorectal cancer. Besides, nobiletin inhibited cell proliferation, migration and angiogenesis by suppression of Nuclear factor-κB. Nobiletin induces apoptosis in MCF-7 cell, cleaved caspase-3, and p53. Nobiletin downregulated the protein matrix metalloproteinase-2 and inhibited cell migration and matrix metalloproteinase-9.[Citation33]
PMFs greatly inhibit the growth of colorectal tumors. Reduction in BaP mutagenic metabolite and DNA was identified within the PMFs-treated groups. Nobiletin, a citrus flavonoid, was given orally along with atorvastatin at a concentration of 0.02%, and the combined effect was far more effective at inhibiting colon carcinogenesis in rats than either substance alone at greater concentrations. This medication significantly reduced colon cancer risk by drastically altering important signaling proteins.[Citation34] Nobiletin, sinensetin, tangeretin, and scutellarein have a significant role in colorectal cancer model is used to study cell proliferation and stemness. It aids by reducing PROM1 and LGR5 expression as well as colony formation. Scutella rein tetramethylether altered the expression of the mesenchymal/metastatic transitional markers CDH1, ZEB1, and SNAI1 while downregulating the expression of PROM1 and SNAI1 with nobiletin.[Citation35] 5-acetyloxy tetramethoxyflavone is a tangeretin derivative more bioavailable and potent for suppression of cell lines from breast and leukemia cancer proliferative activity. It also showed significant results in non-small cell via postponing G2/M-phase and obstructing in related cell proliferation with CDC25c and cdc2. It also apoptosis in the cell with the Caspases are activated, and Survivin, Bcl-2, and XIAP are downregulated due to the cytosolic release of cytochrome c and the mitochondrial membrane potential being disturbed. Autophagosome formation activated autophagy, elevated LC3-II levels, and it lowered the signaling pathways of phosphatidylinositol 3-kinase/AKT/mTOR. In an animal study, polymethoxyflavones significantly slowed the progression of tumor growth in a mouse model without adverse effect.[Citation36] In a concentration-dependent way, caustic in inhibits in vitro carcinogenesis and the features of cancer stem cells, reduces the capacity of second-generation spheres from SCLC H446 cells to form colonies, and downregulates uPAR and CD133. Additionally, casticin dose-dependently decreased p FoxO3a expression and increased AMPK and ACC phosphorylation levels. Small interfering RNAs were used to shut down AMPK, attenuating the aforementioned effects. It also showed similar behaviors to those that counteracted casticin’s effects on in vitro carcinogenesis and CSC characteristics.[Citation37]
The synergistic effects of PMFs and paclitaxel (PTX) a highly effective substance used to treat cancer were also observed on lung cancer cells. A significant reduction in cell viability and increasing of apoptosis was noticed in the CL1–5 cells via the caspase pathway.[Citation38] Sudachitinis a polymethoxyflavone which has significant role apoptosis. A recent study demonstrated the role of PMFs on keratinocyte cells, sudachitin activates the p38MAPK and inhibit the ERK1/2, while on the other hand nobiletin, activated ERK1/2. Furthermore, sudachitin blocked EGF-induced cell migration.[Citation39] PMFs also have positive role malignancy suppression, it neutralizes the pH of cell organelles by ammonium chloride, restore melanogenesis, degrade the tyrosinase in lysosomes and inhibit melanogenesis by the acidification of the cells.[Citation40]
One of the deadliest subtypes of thyroid cancer, anaplastic thyroid carcinoma has a high fatality rate. No significant cure has been found for this and it needs new therapeutic strategies. Nobiletin decreases significantly thyroid cancer and its effect has been sowed the same effects like the as other conventional cytotoxic drugs for example cisplatin. It has also been shown that cisplatin associated with PMFs is more effective compared to a drug administered alone.[Citation41]
PMFs have been shown to decrease the rate of LNCaP cells of the prostate cancer, as it has been seen in inhibiting Apoptosis was concurrently induced by the development of LNCaP cells (GI50 14.6 M). By demethylating the p21 promoter region, it increased the expression of the p21 gene at the genetic level and decreased the DNMT 3B and HDACs 1, 2, through epigenetic controls.[Citation42] Another study showed that PMFs showed significant therapeutic values in metastatic prostate cancer through cellular apoptosis. Casticin reduced cell mobility, supported the prevention of cell invasion and migration and diminished MMP2/9‘s gelatinolytic activities. Casticin, however, increased E-cadherin levels after 48 hours after therapy while decreasing them at 24 hours.[Citation43]
Anti-inflammatory effects
Inflammation is the immune-mediated response of the body to internal injuries or external factors and its types are acute and chronic.[Citation44,Citation45] The acute phase starts with including infection, injuries, immune or toxins reactions and it develops rapidly but chronic inflammation lasts for a long time and causes by the acute inflammation, autoimmune conditions, different infections, and settings that are pathogenic.[Citation46,Citation47] A relationship between chronic inflammation and a number of illnesses, including neurodegenerative, metabolic, and cardiovascular conditions.[Citation33] PMFs trigger the antioxidant-mediated response that activates superoxide dismutase, glutathione peroxidases, catalase, and phase II detoxifying enzymes which eliminates toxic reactive intermediates generated by xenobiotic metabolism.[Citation48] Nitric oxide, Prostaglandin E2, interleukin-1 beta, and interleukin-6, as well as tumor necrosis factor-alpha production were all reduced. Additionally, it inhibits and reverses the production of cyclooxygenase-2, TNF-, IL-1, IL-6, and also LPS-induced nitric oxide synthase.[Citation49]
Trimethylamine-N-oxide, which plays a role in vascular inflammation, was also reduced by PMFs and tea, as were other inflammatory markers. Tea and PMFs also significantly decreased the VCAM-1 and the TNF- and E-selectin mRNA.[Citation50] PMFs alleviated lipopolysaccharide whereas they had no effect on TLR1, TLR2, TLR4, and TLR6, they increased inducible nitric oxide synthase and IL-6.[Citation51] Faqueti et al.[Citation7] demonstrated the antinociceptive and anti-inflammatory properties of standardized polymethoxyflavone extract. After oral administration there has been observed an improvement Interleukin-1beta, prostaglandin E2, and the proinflammatory cytokines tumor necrosis factor-alpha and formalin acutely produced nocifens behavior in mice.[Citation7]
Tangeretin is the most effective polymethoxyflavones compound with anti-inflammatory effect. Tangeretin increased heme oxygenase 1 and NAD(P)H quinone oxidoreductase 1 expression, according to research done by Liang et al..[Citation52] Additionally, tangeretin stimulated nuclear factor-like 2 translocations, mitogen-activated protein kinase phosphorylation, and luciferase activation that is reliant on the antioxidant-responsive element.[Citation52]
Hagenlocher et al.[Citation53] examined how nobiletin/tangeretin affected the activation of human intestinal mast cells by LPS and IgE. Nobiletin or tangeretin used in the treatment of human intestinal tissue before stimulation of inflammation through LPS/sCD14 or IgE-dependently. Phosphorylation of ERK1/2 and pro-inflammatory cytokine expression and degranulation were observed. Nobiletin/tangeretin inhibited the CCL3, CCL4, CXCL8, and IL-1β and showed different reaction like nobiletin BETA-Hexosaminidase and cysteinyl LTC4 release was reduced, but tangeretin did not respond. But tangeretin only reduced and inhibited −1 expression while in the case of NF-κB or MEK-inhibitor, both respond in the same way by showing the effect on producing mediators. Nobiletin and tangeretin dramatically reduced the phosphorylation of ERK1/2.[Citation53]
Crohn’s disease and ulcerative colitis as the two main types of chronic inflammation in the gastrointestinal tract, has increased recently. IBD-related ulcers cause severe diarrhea, life-threatening colon cancer, and severe abdominal pain.[Citation54] It has been demonstrated that PMFs, particularly nobiletin and tangeretin, have suppressive effects on a number of pro-inflammatory cytokines, including important mediators related to IBD. It has been demonstrated that nobiletin inhibits the COX-2 and iNOS, two proteins. Additionally, it reduces the release of proinflammatory cytokines as well as mediators like NO and PGE2. After receiving nobiletin, IL-10-/- mice had significantly fewer colitis symptoms and lived longer. In IL-10-/- mice treated with nobiletin, mast cell number and degranulation were reduced, and nobiletin also reduced the fibrotic marker of collagen deposition. Nobiletin therapy decreased the expression of the pro-inflammatory cytokines IL-6, TNF, and CCL2 as well as collagen expression in hiFB cells.[Citation53]
In a recent study, the therapeutic effects of PMFs on mice with endotoxic shock caused by lipopolysaccharide (LPS) were also revealed. It was clearly shown that pretreatment with nobiletin reduces the levels of the late-phase mediator HMGB1 in blood and tissues of the kidney, lung, and liver. According to the histopathological investigation, NOB treatment considerably lessened the tissue damage brought on by LPS. Additionally, NOB inhibits nuclear factor-kappa B activity.[Citation36] Another study revealed that not only did it block nitric oxide synthesis caused by LPS in a dose-dependent manner, but it also markedly reduced the expression of pro-inflammatory mediators, such as IL-6, PGE2, and IL-1, and prevented nuclear translocation of NF-B and AP-1.[Citation55]
PMFs have been widely used as a hepatoprotective agent all around the world by reducing the oxidative status, LDL receptor expression and activity are increased by lowering LDL absorption in macrophages. A study discovered the protective effects of PMFs 50% extracted against hepatotoxicity caused by Carbon tetrachloride. The 50 and 200 mg dose of PFMs in both acute and chronic male mice showed significant alteration in hepatic enzymes level and by increasing antioxidant enzymes and lowering lipid peroxidation levels, the state of oxidative stress was regained. Additionally, cytoprotective signals as well as nuclear factor E2-related factor 2 were also increased.[Citation56,Citation57]
As in lipopolysaccharide-stimulated microglia, tangeretin has been demonstrated to have a significant role in anti-inflammatory responses and protective benefits. Tangeretin prevented LPS from increasing nitric oxide, tumor necrosis factor-alpha, interleukin-6, and IL-1 production. Matrix metalloproteinase-3 and MMP-8 protein levels were reduced, whereas tissue inhibitor of metalloproteinase-2 expression was increased. This increased expression inhibited nuclear Factor-B.[Citation58] By measuring the activity of the NF-B protein signaling pathway and mitogen-activated protein kinase protein, a study investigated the inhibitory mechanism of tangeretin. Tangeretin also reduced IB- and IKK-stimulated phosphorylation in response to LPS, in addition to inhibiting ERK, N-terminal Kinase phosphorylation.[Citation59]
Another study showed the protective effects against inflammation of PMFs and hydroxylated PMFs. PMFs had lower cytotoxic effects compared to MTTs verified by the MTT method. On the other hand, the comparison of two monomers of PMFs, such as 3,5,6,7,8,3 ′, 4′-heptamethoxyflavone, showed less cytotoxic effects than 3,5,6,7,3′, 4′ -hexamethoxyflavone. Low concentrations (10 μg/mL) of OPE enriched with Hepta MF, PMF and OH-PMF suppressed the expression of the group of genes responsible for the inflammatory response. Significantly, a dose of 250 mg/kg orally reduced inflammation with no side effects and reduced paw edema.[Citation60]
PMFs demonstrated to suppress the activation of inflammatory mediators associated with the toxicity. In contrast to cadmium-induced cardiotoxicity, tangeretin’s effects are explained by Zhang et al.[Citation61] via lowering oxidative stress and pro-inflammatory mediators. When compared to the negative control group, the cadmium levels were considerably lower following tangeritin treatment. Heart function markers significantly decreased in the tangeritin-treated rats as compared to the negative control group. In the cardiac tissues of the tangeritin-treated group, HO-1 and Nrf-2 expression was markedly higher than in the untreated group.[Citation61]
Smoking, arsenic elements are exogenous toxicants triggers the oxidative stress initiate many human diseases and becomes the reason for cancer development.[Citation62] In response to transcription factor NF-E2 counteract the oxidative stress induced by toxicants.[Citation63] PMFs are significant Nrf2 activators. It was demonstrated by a study observed the PMFs induced cNrf2-dependent protection against smoking and arsenic-induced cytotoxicity in lungs epithelial cells.[Citation64,Citation65] A study by Lakshami et al.[Citation66]has shown good antioxidant activity of tangeretin against 7,12-dimethylbenzanthracene -induced renal damage in rats with breast tumors. Tangeretin was administered orally for four weeks and significantly reduced the levels of lipid peroxides, markers of DNA damage, inflammatory cytokines and stabilized Nrf2/Keap1 expression.[Citation66]
Anti-diabetes properties
High blood glucose levels are a defining characteristic of diabetes, a chronic illness. One of the most prevalent chronic illnesses is it, currently affecting an impressive number of people around the world; it can occur at any age and, if not treated in time and correctly, causes serious complications: cardiovascular diseases; eye damage; renal impairment; nerve damage.[Citation26] A healthy lifestyle, a correct allopathic treatment and a complementary treatment with natural compounds with real therapeutic properties proven by scientific studies can control the increased value of serum glycemia.[Citation67] The effect of chenpi which is a citrus plant Citrus reticulata Blanco’s dry peel extract on treating metabolic including diabetes, hypertension, cardiovascular disease etc. oral dosage of Chenpi extract in diet for 15 weeks reduced obesity, hepatic steatosis, and diabetes symptoms via activating protein kinase in adipose tissue. That were induced by HFD in rats. The results demonstrated the positive effects in the treatment of type 2 diabetes.[Citation20] PMFs helps reduce obesity and increases insulin sensitivity It was also noted in a study by Ochiai et al.[Citation68] that black ginger extract containing several PMFs displayed PPAR ligand-binding potential in vitro, irrespective of adiponectin secretion. The black ginger extract containing extract PMFs represses accumulation of fat in adipose tissues, liver, and muscles. While in another study there were no differences in plasma triacylglycerol levels between the groups of black ginger and black ginger extract containing PMFs during the lipids administration test. Tangeretin has a preventive role in diabetes by improving lipid profile, reducing oxidative stress and proinflammatory cytokines. A study explored the defensive nature of tangeretin. Diabetes reversed in diabetic rats by improvement in plasma and cardiac lipid profiles. By giving diabetic rats tangeretin orally for 30 days, the drug altered the activity of lipid metabolic enzymes increased GLUT4 expression in the heart and liver. By boosting the activities of antioxidant enzymes, inflammatory cytokines and cardiac marker enzymes were reduced in the plasma and heart of diabetic rats.[Citation68]
PMFs also have protective effects on diabetic retinopathy caused prolonged hyperglycemia in diabetes mellitus. After 48 hours of receiving streptozotocin injections, animals whose blood glucose levels were more than 350 mg/dL were given nobiletin for four weeks. Blood retinal barrier permeability was quantified at the end of eight weeks. Nobiletin reduced permeability enhanced the thickness of the retinal layer and the Blood-Retinal Barrier.[Citation69]
Guo et al.[Citation70] revealed that chenpi extract is enriched 5-OH PMFs- and is most active to prevent obesity, hyperglycemia, insulin resistance, hypercholesterolemia and fatty liver AMPK pathway. Castin is a 5-OH PMF derivative has been investigated in lipid model, have one extra hydroxyl group at which counterpart, 5-OH nobiletin. It reduces intracellular lipid accumulation in differentiated adipocytes. Casticin has an additional hydroxyl group at the 3” position compared to its counterpart, 5-OH nobiletin, which results from adipogenesis” direct suppression of cell growth and lipogenesis. Additionally, compared to 5-OH nobiletin, casticin has a better efficacy at decreasing cholesterol levels and a different mechanism of action, demonstrating the structure-activity link of 5-OH PMFs in the suppression of adipogenesis. In the case of Oral use of chenpi extract reduces hepatic fat accumulation, body weight increase, and type II diabetes symptoms when treating obesity and type II diabetes in case of treatment of NAFLD and NASH, it significantly reduced the hepatic steatosis, inflammation and fibrosis.[Citation70] Effect of orange peel extract in type 2 diabetes. It reduces inflammation. OPE lowered blood sugar levels and was much better tolerated than metformin, an oral atidiabetes drug.[Citation71]
Nobiletin’s effects on cultured human pancreatic islets were examined in a study. Different nobiletin doses were applied to isolated human islets, which were then cultivated for 24 and 72 hours. Apoptosis markers were assessed and then measured. Through decreased HIF-1 and ROS production, apoptosis suppression, and increased islet activity, nobiletin therapies greatly increased the islets’ longevity. Nobiletin’s ability to protect the islets. Therefore, the pre-transplantation period during which nobiletin was administered is beneficial to create live and functional islets for clinical transplantation.[Citation72]
It has been demonstrated that nobiletin suppresses hepatic lipogenesis by AMP-activated protein kinase. According to study findings, nobiletin significantly reduced hepatic lipid buildup brought on by high glucose in HepG2 cells. Additionally, it decreased the protein production of lipogenic factors. Compound C, an AMPK inhibitor, pretreatment eliminated nobiletin’s effects on SREBP-1c and FAS expression.[Citation73]
By reducing hepatic fat accumulation in a mouse model of obesity and diabetes brought on by the HFD, oral treatment of N-phenylpiperazine successfully reduces body weight gain and type II diabetes symptoms. Additionally, oral Niclosamide ethanolamine therapy directly inhibits hepatic stellate cell activation, preventing and reversing CCl4-induced fibrosis in mice. Overall, hepatic steatosis and the stimulation of hepatic stellate cells are the targets of NEN-induced mitochondrial uncoupling, which explains its therapeutic promise for treating NASH.[Citation70]
Anti-obesity effects
Obesity is a becoming one of the major health problem all around the world due to the energy disparity Energy is stored as fat when the amount of energy consumed exceeds the amount of energy expended. Both intra-abdominal and subcutaneous adipose tissue can be deposited.[Citation26,Citation28] In vitro and in vivo studies have demonstrated the ability of PMFs to control lipid metabolism, prevent obesity, and treat type 2 diabetes.[Citation74] Dysfunctions of lipid and lipoprotein metabolism cause various severe complications such as heart disease, hypertension, obesity and diabetes (Salehi et al.,2020). To test its impact on obesity and NAFLD, Pan et al.[Citation75] devised a supplement made of citrus PMFs, green tea extract, and lychee extract. Both weight growth and the size of fat cells were dramatically reduced. Additionally, it was shown that the liver and adipose tissue both had downregulated levels of Sterol regulatory element binding protein 1, AMP-activated protein kinase signaling, and peroxisome proliferator activated receptor. 5-demethylated polymethoxyflavones rather caused a reduction in activation of AMP-activated protein kinase related to lipid accumulation and by controlling the lipogenic genes side by side down-regulation of adipogenic transcription factors.[Citation76] Hydroxymethoxyflavone significantly showed not the only reduction in body weight but also decreased the serum levels of liver function test as well as downregulated the hepetic genes at the genetic level, besides they also lowered the inflammatory response of body by upregulating and controlling the genes involved in the oxidation of fatty acids and energy.[Citation74] Hydroxylated polymethoxyflavones also activate the protein kinase signaling in 3T3-L1 adipocytes.[Citation77] Nobiletin and 5-demethylnobiletin have shown to be more significant in reducing intra-abdominal fat, body weight, plasma and liver TG levels, and plasma cholesterol both in vivo and in vitro.[Citation78]
Cardio protective effect
Arterial hypertension leads to further complications of liver, kidney, stroke, heart and spleen, which disturbs the balance of the body.[Citation79] Natural bioactive compounds help in restoring the immunological functions of the body by regulating the visceral functions, which helps in treating hypertension.[Citation80] The prophylactic effects of PMFs were determined by Li et al.[Citation33] on hypertensive rats induced by Nω-nitro-L-arginine. Hexamethoxyflavone in hypertensive rats, heme oxygenase-1, endothelial nitric oxide synthase mRNA expression, neuronal nitric oxide synthase level, adrenomedullin, receptor activity modifying protein 2, adrenomedullin, and similarly inducible nitric oxide synthase expression were represented by sinensetin, tetramethyl-O-isoscutellarein, tetramethyl-o-scut.[Citation33] Sinensetin promotes the peroxisome proliferator-activated receptor, the sterol regulatory element-binding protein 1c, and the CCAAT/enhancer-binding protein. It has been seen in the expression of C/EBP and the activation of the protein that binds to cAMP-responsive elements. According to Kang et al.[Citation81] 3T3-L1 preadipocytes treated with sinensetin had higher levels of intracellular cAMP and protein kinase A activation.
Type 1 diabetes mellitus increased the state of oxidative stress linked with the activation of c-Jun Nuclear Kinase in the body leads to can cardiac hypertrophy. Oral treatment with naringin on rats for 56 days expressively reversed the state of lipid peroxidation, proteins oxidation, oxidative stress.[Citation82] A study was conducted by Jenie and her fellows showed that only small concentration of tangeretin can lessen the H2O2-induced oxidative stress on human umbilical vein endothelial cells.[Citation83] Another recent study investigated the antihypertensive role of naringin on 5 groups of rats. The hemodynamic and left ventricular function and biochemical parameters and histological were assessed in the kidney. At 15, 30, and 45 minutes after the clamp was removed, it was found that the renal artery occlusion group’s hemodynamic parameters had dramatically increased.[Citation84]
Neuroprotective effect
Neurodegenerative diseases are chronic, incurable disorders that cause nerve cells to gradually deteriorate or even die. They interfere with mental or physical function.[Citation63] The most prevalent neurodegenerative illnesses are Alzheimer’s and Parkinson’s, which are characterized by intracellular neurofibrillary tangles and extracellular amyloid plaques that cause nerve cell death.[Citation67,Citation85] The prime target in neuropathological disorders are the production of amyloid- proteins is catalyzed by the one of the major steps in the development of Alzheimer’s disease.[Citation86] The strongest evidence is available suggested that PMFs exhibits strong neuroprotective, anti-inflammatory and anti-dementia effects in vivo and vitro studies by inhibiting functions of BACE1 that could lower Aβ production and improving the regulation of signals at the molecular level through interaction with mitochondria.[Citation87] The amyloid-peptide (A), the defining feature of The oxidative and inflammatory pathways that Alzheimer’s disease sets off lead to the loss of neurons. Nobiletin is a powerful inhibitor of A. By reversing aberrant alterations in intracellular oxidative stress, nuclear morphology, and apoptotic caspase activity, nobiletin greatly reduced A-mediated cell death. Nobiletin significantly reduced the synthesis of tumor necrosis factor, interleukin-1, nitric oxide, and prostaglandin E2 in response to A stimulation in terms of anti-inflammatory reactions. Additionally, nobiletin reduced the expression of cyclooxygenase-2 and inducible NO synthase when A was stimulated, which was linked to the inhibition of nuclear Factor-B p65 and phosphorylation of its inhibitor, I-B-. It’s interesting to note that nobiletin reduced c-Jun N-terminal kinase and p38 expression without influencing the activation of extracellular signal-regulated kinase ½. A study conducted by Nakajima in[Citation88] on demonstrated nobiletin for 3 months improved the short-term and recognition memory in 3×Tg-AD mice as it reduces the level of soluble A1–40 in the brain as soluble A1–40 has been shown to demonstrate to induce synaptic dysfunction and memory impairment PMFs becomes a novel drug for the treatment and prevention of AD. As a result of its ability to reduce inflammation, oxidative stress, and apoptosis, nobiletin may be a promising candidate for the treatment of Alzheimer’s disease.[Citation89] By measuring the activity of the NF-B protein signaling pathway and mitogen-activated protein kinase protein, a study investigated by Shu et al.[Citation59] the inhibitory mechanism of tangeretin. Tangeretin also reduced IB- and IKK-stimulated phosphorylation in response to LPS, in addition to inhibiting ERK, N-terminal Kinase phosphorylation. Tangeretin suppresses the unnecessary microglial activation associated with the neurotoxicity. It diminished the production of nitric oxide (NO), tumor necrosis factor-alpha, prostaglandin E2, interleukin-1 beta, and interleukin-6. Moreover, it subdued the expression of LPS-induced nitric oxide synthase, TNF-α, IL-1β, and IL-6 and cyclooxygenase-2 in microglial cells. The pharmacological action of PMFs on AMP/PKA/ERK/CREB signaling has been shown to improve short and learning memory. Kawahata et al.[Citation90] found that sinensetin amplified the transcription to a greater extent as compare to nobiletin. Furthermore, he identified that tangeretin, sinensetin, 6-demethoxynobiletin, and 6-demethoxytangeretin were also identified as bioactive substances. In animal models of Alzheimer’s disease, nobiletin has been demonstrated to reduce oxidative stress, tau hyperphosphorylation, and other pathogenic characteristics of the illnesses, such as cognitive impairments. Additionally, nobiletin reduced the cognitive and motor impairments in PD animal models.[Citation91]
Hepatoprotective effect
The liver is the largest organ of the body involved majorly in metabolism, energy production, detoxification, storage and activation of vitamins and minerals. Certain factors lead to liver damage, oxidative stress is one of them which activates the Kupffer cells, macrophages, increase the release of damaging pro-inflammatory mediators that harm the liver such as tumor necrosis factor-α.[Citation92] Chronic inflammation brought on by the ongoing liver damage progresses into fibrosis, cirrhosis, and ultimately end-stage liver disease. PMFs have been widely used because of their hepatoprotective effects all around the world by reducing the oxidative status, reducing the uptake of low-density lipoprotein improve the production of and activity of LDL receptors in macrophages.[Citation92] Nobiletin is the most effective among all PMFs in the treatment of the liver diseases. A study conducted by Kim et al., discovered the protective effects of 50% PMFs extracted from Citrus aurantium L. extract against hepatotoxicity caused by CCl4. Both acute and chronic male mice showed a substantial change in hepatic enzyme levels after receiving the 50 and 200 mg dose of PFMs. The oxidative stress status was restored by strengthening antioxidant enzymes and lowering levels of lipid peroxidation. Additionally, cytoprotective signals were also increased.[Citation56] Another study showed the pharmacological effect of 50 and 100 mg/kg Citrus aurantium L. extract (CAE) derived 27% PMFs nobiletin on high fat-induced animal model developing NAFLD for 8 weeks to induce obesity. All of the animals were put to death after 8 weeks of therapy, and the liver and plasma tissues were examined histopathologically and biochemically. According to the findings, HFD-induced NAFLD in groups treated with CAE is dramatically reduced, along with the blood lipid index level and liver histology.[Citation93] Expression of the genes for CD36, scavenger receptor A, and lectin-like oxidized LDL receptor 1 and activity were all decreased by PMFs. HepG2 increased LDL receptor activity and transcription and decreased the A crucial enzyme in the hepatic triacylglycerol biosynthetic process is diacylglycerol acyltransferase 2, or acyl CoA. This was at least partially done through steroid-response element-binding protein 2 activation. The research also demonstrated that nobiletin’s 5-demethyl form is highly effective at preventing foam cell production and the conversion of monocytes to macrophages.[Citation94]
Other biological properties
Antimicrobial
Flavonoids are universal in photosynthesizing cells and are normally found in natural product, nuts, seeds, vegetables, stems, blossoms, wine, tea, and nectar. Arrangements with these mixes as the essential physiologically dynamic components have been used to treat human ailments for a very long period. This category of everyday things is increasingly becoming a topic with antifungal, antiviral, and antibacterial action.[Citation95] Additionally, a few investigations have exhibited cooperative energy between dynamic flavonoids just as among flavonoids and existing chemotherapeutics. Certain examinations have analyzed the connection between flavonoid construction and antibacterial, in the same way various examination bunches have tried to clarify the antibacterial instruments of flavonoids. The action of quercetin, for instance, has been unquestionably somewhat credited to hindrance of DNA gyrase and that licochalcones an and C restrain energy digestion. Different flavonoids whose components of activity have been explored incorporate robinetin, apigenin, myricetin, rutin, galangin. These mixtures address novel leads, and future examinations might permit the improvement of a pharmacologically satisfactory antimicrobial specialist or class of specialists.[Citation96]
The outcomes permit the researchers to set up connections between antimicrobial impact of the mixtures and film constructions of these microorganisms. A more significant outcome was achieved when flavonoid was used in combined against E. coli than against S. aureus. because of the presence of porins in the external layer of G – microscopic organisms. The substance that served as an enhancer works by removing the charges on the amino acids in the porins and creating a pathway for other substances to enter the bacterial cell through diffusion.[Citation97]
Seven pure flavonoids were segregated and recognized from five moss species. The antibacterial activities of some of these flavonoids against E. aerogenes, Enterobacter cloaceae, and Pseudomonas aeruginosa have been demonstrated by Basile et al..[Citation98] These flavonoids may be crucial instruments in antibacterial efforts because of their antibacterial range, which is primarily active against Gram negative bacterial strains that cause serious opportunistic infections and are resistant to conventional antibacterial therapy due to their antibacterial range principally dynamic against Gram negative bacterial strains for serious pioneering diseases, these flavonoids might be significant devices in antibacterial methodologies. The liposomes dramatically boosted fluorescence polarization.[Citation99] Such increases were observed in both liposomes when two fluorescence probes were used to test the fluidity of the membrane. This suggests that sophora flavanone G decreases the fluidity of the membranes. Naringenin also had a membrane impact, although it required concentrations greater than 2.5 g/ml to significantly reduce membrane fluidity. It is thought that sophoraflavanone G inhibits bacterial growth by decreasing the permeability of cellular membranes.[Citation100]
Sophoraflavanone G isolated from Sophora exigua showed significant antibacterial actions recommending that sophoraflavanone G suppressed the flow in the external and internal layers of membrane. The impact of Galangin against Staphylococcus aureus was observed via patterns of potassium loss from suspensions of bacterial cells. The MICs flavonol galangin was 50 μg/mL, the bactericidal anti-bacterial Against Staphylococcus aureus NCTC 6571, novobiocin and the bactericidal anti-infection penicillin G were used.[Citation101]
Penicillin G showed a 6% increase in potassium loss while Novobiocin had no effect on potassium loss showed that PMFS play a significant role against Staphylococcus aureus cells losing potassium, which might destroy cytoplasmic layer or create autolysis and weakens cell wall.[Citation102] Leguminosae and their antibacterial actions against methicillin-safe Staphylococcus aureus plays a major role in destruction of MRSA action and that replacement with a specific aliphatic gathering at the 6-or 8-position likewise improves the movement.[Citation103,Citation104] Two bioflavonoids, 5,7-dihydroxy-4′,6,8-trimethoxyflavone and 5,6-dihydroxy-4′,7,8-trimethoxyflavone, were tested for their antimicrobial effects against Candida albicans, Escherichia coli, Bacillus subtilis, Staphylococcus aureus, Alternaria solani, and Salmonella typhimurium, among other microbial strains, compound 1 demonstrated bactericidal Compound 2 was found to efficiently kill B. subtilis by cell lysis, while compound 1 showed efficacy against E. coli and S. aureus. Intensities 1 and 2 each had a limiting effect on the expansion of A. C. Solani and albicans.[Citation105] SEM analysis of the treated cells was used to focus the effects of flavonoids on cellular architecture and glucose metabolism. This analysis also looked at the specific actions of each pathway’s main catalysts. At sublethal doses, they reduced phosphofructokinase and isocitrate activity while enhancing the mobility of gluconeogenic fructose bisphosphatase., quercetin and kaempferol has been found to balance to induce alterations in the atomic factor kappa B pathway, cyclooxygenase-2, responsive C-protein articulation, and inducible nitric oxide synthase Quercetin or kaempferol in the human hepatocyte-derived cell line or a cytokine mixture were incubated with Chang Liver. On the levels of the proteins iNOS, COX-2, and CRP, kaempferol had a dose-dependent effect. The two flavonoids altogether restrained iNOS, COX-2, and CRP mRNA levels.[Citation106] It was believed that Pmfs action involves inhibiting NF-B activation and the ensuing upregulation of the pro-inflammatory genes through the inhibitory actions of quercetin and kaempferol on NF-B activation, protein concentration of the phosphorylated version of the inhibitor I-B, and IKK. Our results further imply that the minor structural differences between the two drugs account for variations in their inhibitory potency.[Citation107,Citation108]
Antiviral
An extract of Eclipta alba had the ability to inhibit HCV replicase, which decreased the viral protein synthesis and HCV RNA titer. Two flavonoid substances showed a dose-dependent suppression of HCV replication in vitro when they were evaluated separately.[Citation109] The viral protease. Non-structural Protein 3 is inhibited by quercetin, which has antiviral effects against HCV by reducing HCV replication. This mechanism has broad genotypic activity against different HCV strains and is associated with a high level of Barrier against resistance mechanisms.[Citation110] Flavonoids such as luteolin, naringenin, diosmetin, and apigenin all have stronger affinity for the HCV NS5B protein than the antiviral medication sofosbuvir.[Citation111]
Regardless of the HBV genotype, epigallocatechin gallate, the primary catechin derived from tea, prevented HBV from entering hepatocellular cell lines from cell through a process that led to the polypeptide sodium taurocholate cotransporter of HBV receptor plasma being endocytosed in a clathrin-dependent manner. Membrane, then its protein is broken down.[Citation112]
Vitexin obtained from the Flos Trollii plant exerts its effect on the H1N1 influenza virus by blocking the Toll Like receptor 3 and Toll Like receptor 7 signaling pathways, partially inhibiting and increasing the molecular Toll like receptor.[Citation112] TLR is a transmembrane glycoprotein, the preferred target of various viruses. TLR activates the core factor signaling pathway of the activated B-cell Capalic Chain Enhancer (NFkB) or through the regulation of interferon to produce pro-inflammatory cytokines. Some viruses enter host cells, but once they invade, the virus can inhibit the production of cytokines that affect the immune response. Viral infection can trigger an inflammatory process characterized by the overproduction of nitric oxide and interleukin.[Citation112] Various flavonoids increase the production of interferon to fight viral infections. The antiviral effect of Scutellaria baicalensis Georgi is related to its activation.[Citation22] The natural extract of Tetrastigma hemsleyanm contains a variety of flavonoids, has different antiviral effects and different antiviral effects. By reducing the number of influenza-induced plaques in infected MadinDarby canine kidney cells. Houttuynia cordata’s flavonoid quercetin rhamnoside exhibits anti-influenza A/WS/33 action and lessens virus-mediated Hic cell pathology through direct interaction with virus particles. In addition, the same author also showed that after 6 days of influenza virus infection, quercetin rhamnoside has anti-influenza activity in mice at a dose of 6.25 mg/kg/day.[Citation111,Citation113]
By preventing the viral nonstructural protein 1‘s Asn130 glycosylation, the flavonoids Sanggenon O, EGCG, and Chamaejasmin can prevent the replication of the dengue virus. A flavonoid called begalin was discovered in Scutellaria baicalensis Georgi. It has virucidal effect on the extracellular virus particles of Dengue fever virus 2.[Citation56] Studies on the antiviral effects of flavonoids, quercetin and rosin have shown that riboflavin can inhibit the molecular replication mechanism of dengue virus and enterovirus A71. In addition, another antiviral mechanism used by pinocembrine is to attack the molecular replication mechanism of Zika virus. Have its own genome in the host cell. This flavanone works by inhibiting the production of viral RNA and the synthesis of coat protein during the post-entry process of the Zika virus replication cycle.[Citation114,Citation115]
Interestingly, the flavonoids quercetin and kaempferol are among the several phytochemicals that moringa trees offer, and their leaf extract can be utilized as a preventive or therapeutic treatment for HSV1. Extract from Moringa oleifera Lam considerably decreased plaque formation brought on by strains of phosphonoacetate-resistant HSV, thymidine kinase-deficient HSV, and wild-type HSV. Additionally, the HSV1 virus is resistant to the extract from erythrina, which is mostly attributed to vitexin’s antiviral activity, which is a result of the interaction between flavonoids and the HSV1-binding pocket. is associated and ensuing thymidine kinase release.[Citation116] It’s interesting to note that Melissa officinalis’ apigenin-7O-glucoside suppressed virus multiplication, which decreased the quantity of RRV-induced plaques in infected MA104 cells. Aspalathus linearis luteolin and vitexin also showed antiviral action.[Citation117,Citation118] Tangeretina and Nobiletina are two polymethoxy flavonoids extracted from cross-linked citrus chacha fruit with anti-RSV properties. Hesperetin inhibits and has a dose-dependent inhibitory effect on RSV-induced plaque formation in HEp2 cells. The phosphoprotein RSV (Protein P) that binds to the viral genome to form a complete nucleocapsid. The antiviral effect of this secondary plant material is similar to the common antiviral medication ribavirin.[Citation119]
Anti-osteoporosis (prevention of bone loss)
The number of elderly patients is increasing, and it is anticipated that one resorption disorders will become more common. These diseases include osteoporosis and periodontal disease. Bone remodeling and bone mass are controlled by the equilibrium between osteoclast bone resorption and osteoblast bone synthesis. The primary bone-resorbing cells, which differ from the macrophage monocyte cell line, are called osteoclasts. The relationship between the activator of NF-B ligand and Osteoclast differentiation requires (RANKL) and RANK.[Citation120]
When bone resorption agents such lipopolysaccharide (LPS) are present, osteoblasts respond by expressing RANKL on their cell surfaces.[Citation121] Interleukin (IL) 1 is also present, and osteoclast precursor cells express RANK. Osteoblasts are the principal producers of prostaglandin (PG) E2, which is a potent inducer of inflammatory bone resorption. It is well known that LPS and IL1 stimulate osteoblasts to produce PGE2 by increasing the membrane-bound form of PGE synthase 1 and cyclooxygenase 2 mRNA and PGE2 induces the surface osteoblasts’ expression of RANKL.[Citation122] PGE2 is known to play a significant EP4 receptors on osteoblasts are able to detect this participation in inflammatory bone resorption, which causes them to produce RANKL, which causes osteoclast differentiation.[Citation123,Citation124]
The most prevalent condition affecting bones is osteoporosis, and postmenopausal women who have low estrogen levels may experience substantial bone loss.[Citation125] Mice that have undergone ovariectomies are frequently utilized as postmenopausal osteoporosis animal models. Due to the lack of estrogen in the femur, OVX mice suffered severe bone loss. Inflammatory cytokines such as IL1 may be related to bone loss.[Citation126] Periodontal disease is an inflammatory bone disease. The breakdown of alveolar bone results from the advancement of periodontal disease. The membrane of in gram-negative bacteria, the pathogenesis is aided of teeth around inflammation. Periodontitis transmits a similar signal to the receptor (TLR) 4 In the synthesis of PGE2 mediated by mPGES1, it has been shown to be important for LPS-mediated bone loss in periodontal disease. The relationship between OVX-induced bone loss and local inflammatory bone destruction has been extensively studied in periodontal models. Mice’s alveolar bone suffered substantial bone loss as a result of the OVX and LPS treatments. It has been found that elderly patients receive treatment for two bone diseases at the same time.[Citation125]
PMFs, which have a broad spectrum of biological processes oranges and hesperetin have 6 or 5 methoxy groups and show different activities such as anti-tumor, anti-inflammatory and anti-obesity. Heptamethoxy flavone regulates inflammation and The behavior of the central nervous system. By attenuating IKK-mediated NFκB activation in vitro and restoring femoral loss in OVX mice, IL1 induces osteoclast differentiation. In a mouse model of periodontitis, local injection of chuanxifen and hesperetin can inhibit LPS-induced alveolar bone loss. The HMF model also inhibits osteoclast differentiation and LPS in-induced bone resorption. Recent reports indicate that a PMF mixture composed of Ligusticum chuanxiong, oranges, tetramethoxy flavonoids and HMF extracted from orange peel can prevent ultraviolet rays.[Citation126]
Skin injury
PMF strains composed of nobiletin, Tangeretin, TMF and HMF, and confirmed that PMF protective action against osteoclasts and bone resorption.[Citation127] A mixture of PMF orange peel extracts composed of citrus, tangeretin and HMF uses UVB to inhibit COX2-can be activated to induce expression the receptor is activated by peroxisome proliferation and matrix metalloproteinase 1 by inhibiting the phosphorylated expression of c-jun N-terminal kinase in the human HaCaT keratinocyte line. The PMF mixture is also inhibited the production of melanin in the human melanoma cell line MH3KO.[Citation128,Citation129] Tangeretin’s potency was lower than that of other compounds, whereas that of the PMF mixture was comparable to that of nobiletin and HMF. According to a study, nobiletin and HMF suppress the formation of mature osteoclasts, but IL1 and LPS stimulate osteoclast differentiation via activating the transcription factors NFATc1 and RANKL in osteoblasts and osteoclast progenitor cells, respectively. In addition, in a mouse model of periodontitis, Nob showed a stronger protective effect on bones than tangeretin.[Citation130]
In order to compare the effectiveness of PMF between nob and tan a mixture of 10 µg/ml PMF plus added to a co-culture POB and BMC for osteoclast differentiation caused by IL1. PMF prevented IL-1-induced osteoclast development at this insufficient dosage. while nob and tan inhibited. Therefore, PMF is more effective than nob and tan. The combination of PMFs included nobiletin, tangeretin, HMF, and TMF, these components have a synergistic effect by inhibiting the production of osteoclasts. In mice lacking estrogen, oral PMF (5 mg/day/mouse) dramatically reversed the loss of trabecular bone. In vitro, mixed PMF reduced osteoclast development and bone resorption activity brought on by IL1, and in vivo, it reversed bone loss in OVX animals. In sham mice, PMF administration improved the BMD measured by DEXA in the center of the femur, showing only cortical bone although the volume of trabecular bone measured by μCT was not affected by PMF. The bone formation between trabecular bone and cortical bone is different.[Citation131] PMF (5 mg/day/mouse) can More research is required to substantiate the notion that this process can induce bone growth in the cortical bone. Another study found that administering Nob intraperitoneal to OVX mice might restore their lost femurs, but oral PMF is sufficient to restore bone mass.[Citation132] PMF inhibited the production of LPS-induced chemicals that cause inflammation, such as IL6, TNF, and PGE2. PMF does not affect the uterine weight of OVX mice, and oral PMF can help treat postmenopausal osteoporosis.[Citation133]
Skin antiaging
UV rays promote the development of the skin’s matrix metalloproteinase, which are responsible for deep wrinkles. These wrinkles are brought on by the disruption of the extracellular matrix components like collagen fibers.[Citation134] PMF mixture reduces the UVB-induced production of MMP1 by inhibiting JNK phosphorylation. The mixture may function as an anti-photoaging agent.[Citation135] Melanin content determines the skin color and synthesized by melanocytes and catalyzed by tyrosinase. Melanosomal pH is most important in the process of melanin synthesis and PMFs has a visible role in suppressing the melanogenesis. A study found the significant role of PMS in keeping skin young and slowing the aging process.[Citation47] The most important preclinical pharmacological studies of PMFs were shown in .
Table 2. The most important preclinical pharmacological studies of PMFs (this must be completed with all written studies – model below).
Discussion
Citrus flavonoids are an essential component of traditional medicine and are recommended as dietary supplements in addition to being a staple of the daily diet. Human organic anion transporter polypeptides are significant carriers that regulate drug interactions and nutrition. They are inhibited by the polymethoxyflavones nobiletin, sinensetin, and tangeretin, as well as by the flavonoids didymin, hesperidin, and narirutin.[Citation136] A study found that the inhibition was determined by quantifying the decrease in the absorption of the fluorescent substrate OATP1B1 and the substrate 8fluoresceincAMP OATP1B3 and the fluorescent substrate OATP1‘d. d2B3 examines the fluorescence from HEP2K29, in cells with excessive fluorescence from HEP2K29. Results showed that all tested flavonoids inhibit OATP2B1 (IC50 1.6 to 14.2 μM) in the lower micromolar range, but only polymethoxy flavonoids inhibited OATP1B1 and 1B3 (IC50 2.1 to 21 μM). Concluded that all the flavonoids studied can promote the intestinal interaction of OATP2B1 with drugs in juice or citrus fruits.[Citation137] Contrarily, the levels of polymethoxy flavonoids after drinking citrus or fruit juice may not be high enough to inhibit OATP1B1 and OATP1B3 in the liver, and when used as a medicine or dietary supplement, there may be side effects.[Citation137,Citation138] The polymethoxyflavones nobiletin, sinensetin, and tangeretin reduced the 8FcA fluorescence in OATP1B1 and OATP1B3. They overexpressed the cell concentration according to the cell concentration. This clearly shows that these OATP inhibits and excludes nonspecific effects, such as extinction inhibition is in the lowest micromolar range (OATP1B1 is 2–8 μM, OATP1B3 is 6–21 μM), Nob is OATP1B1 Tan is the strongest inhibitor. OATP1B3. Polymethoxyflavonoids and known powerful rifampicin inhibitors OATP1B1/1B3 indicate that these natural substances are effective inhibitors, except for for the line, which is approximately beneficial to the effectiveness of OATP1B3. Ten times that of rifampicin. However, rutin flavonoids are not related inhibitors of these OATPs. All tested flavonoids reduced the DBF fluorescence in the OATP 2B1 of overexpressing cells, but not in the parental cell line, indicating an inhibitory effect on OATP2B1, with IC50 values in the lower micro molar range.[Citation139]
Many fruit juices, especially grapefruit juice, are widely recognized for their interaction with clinically used capsules due to the inhibition of OATP. The drug capture transporters OATP1B1 and OATP1B3 are important regulators of the pharmacokinetics of several capsules, including statins, telmisartan, and irbesartan, in the liver absorption process in humans. When these OATPs are inhibited, the plasma concentration of the medication as a substrate may rise and its rate of clearance may drop. Which increased toxicity, but the scientific effectiveness of capsules containing statins decreased. For OATP1B1/1B3 that inhibits citrus fruits/juices or citrus flavonoids, there is simpler and more concise in vitro information so far. As far as we know, this is the first document about polymethoxy flavonoids, chuanxifen, chuannetin and hesperetin as inhibitors of OATP1B1 and OATP1B3. And inhibit OATP1B3 from clementine and citrus juice, we have previously determined.[Citation140] The inhibitory effect on OATP1B1 is the same as the well-known powerful inhibitor of rifampicin. OATP1B1/1B3 inhibitors must, however, reach adequate systemic concentrations in order to be clinically meaningful. The clinical relevance of the three polymethoxy flavonoids tested cannot be predicted because there is no human pharmacokinetic data. Nob and Tan’s bioavailability in rats is not anticipated to be very high.[Citation141] However, pure Nob and Tan or drugs made from PMFs have many health-promoting effects. These high levels of PMFs may lead to higher systemic concentrations, which may pose a risk of collateral interactions with the most commonly used substrate OATP1B1/1B3. OATP2B1 was considered recently the main intestinal subtype that can mediate intestinal drug absorption and drug-food interactions. OATP isoforms mediate the interaction between citrus juice and the drugs originally attributed to OATP1A2. We confirmed the inhibitory effects of Nob, Tan and hesperidin on OATP2B1.[Citation142]
Although there is no agreement between the IC50 value and the previously published value, it is generally accepted that the absolute value cannot be compared across different detection methods. Data also revealed that sinensetin, didymin, and narirutin block OATP2B1. The concentration of oral medications or food ingredients in the intestine is typically significantly higher than that in portal blood, despite the fact that OATP2B1 is expressed in both the liver and the gut. Orange juice lowered celiprolol’s bioavailability, grapefruit and orange juice decreased aliskiren’s area under the curve (AUC), and ronacaleret decreased celiprolol’s bioavailability. The results showed that all of the flavonoids tested are OATP2B1 inhibitors and may therefore contribute to the lowered exposure to rosuvastatin. Citrus species and different batches of juice and intestinal flavonoids have different concentrations. The potential is influenced by the citrus variety and batch in addition to the quantity of juice or fruit consumed. It has been reported that the concentration of flavonoids in fruits or juices is >100 μM (up to 20 mM) which is usually much higher than the effective concentration for inhibiting OATP2B1 (IC50 = 8.3 μM) when taken as dietary supplements or drugs, the intestinal concentration of these flavonoids may be higher, so the risk of interaction with drugs that are substrates of OATP2B1 must be higher.[Citation136]
The poor solubility of PMFs limits their bioaccessibility in the human gut system. In addition, a high metabolic rate influences the amount of PMFs entering the human circulatory system. This ultimately leads to poor bioavailability and reduces the therapeutic dosages they can achieve in the target organs. Researchers have attempted to use emulsions to improve the bioavailability of PMFs. The PMF formulations are intended for the oral delivery of bioactive-based medicines or nutraceuticals.
Conclusion and future perspectives
The citrus genus is rich in citrus PMFs, including nobiletin and tangeretin, which have a variety of biological characteristics. As shown in this review, research on citrus PMFs has increasingly acquired information from animal experiments down to the level of cell lines, proteins, and genes. The excellent bioactivities of PMFs in vitro and in vivo are attracting more and more nutritional and medical specialists for the benefit of humans in illness prevention and treatment. They regulate differentiation, proliferation, angiogenesis, and metabolism processes by controlling the signaling cascade, gene transcription, protein function, and enzyme activity.
Disclosure statement
The authors of the study declare that there were no business or financial connections that may be interpreted as having a potential conflict of interest.
References
- Sharifi-Rad, J.; Cristina Cirone Silva, N.; Jantwal, A.; Bhatt, D.; Sharopov, I.; Cho, F. ;. C.; Martins, W. N. Therapeutic Potential of Allicin-Rich Garlic Preparations: Emphasis on Clinical Evidence Toward Upcoming Drugs Formulation. Appl. Sci. 2019, 9(24), 5555. DOI: 10.3390/app9245555.
- Gao, Z.; Gao, W.; Zeng, S. L.; Li, P.; Liu, E. H. Chemical Structures, Bioactivities and Molecular Mechanisms of Citrus Polymethoxyflavones. J. Funct. Foods. 2018, 40, 498–509. DOI: 10.1016/j.jff.2017.11.036.
- Md Idris, M. H.; Mohd Amin, S. N.; Mohd Amin, S. N.; Wibowo, A.; Zakaria, Z. A.; Shaameri, Z.; Hamzah, A. S.; Selvaraj, M.; Teh, L. K.; Salleh, M. Z. Discovery of Polymethoxyflavones as Potential Cyclooxygenase-2 (COX-2), 5-Lipoxygenase (5-LOX) and Phosphodiesterase 4B (PDE4B) Inhibitors. J. Recept. Signal Transduction. 2022, 42(4), 325–337. DOI: 10.1080/10799893.2021.1951756.
- Gao, Z.; Wang, Z. Y.; Guo, Y.; Chu, C.; Zheng, G. D.; Liu, E. H.; Li, F. Enrichment of Polymethoxyflavones from Citrus Reticulata ‘Chachi’peels and Their Hypolipidemic Effect. J. Chromatogr. B. 2019, 1124, 226–232. DOI: 10.1016/j.jchromb.2019.06.010.
- Wang, R. L.; Li, S.; Ho, C. T. Absorption of Polymethoxyflavones and Their Derivatives. J. Food Bioactives. 2018, 2, 82–90. DOI: 10.31665/JFB.2018.2142.
- Shajib, M. S.; Rashid, R. B.; Ming, L. C.; Islam, S.; Sarker, M. M. R.; Nahar, L.; Rashid, M. A.; Datta, B. K.; Rashid, M. A. Polymethoxyflavones from Nicotiana Plumbaginifolia (Solanaceae) Exert Antinociceptive and Neuropharmacological Effects in Mice. Front. Pharmacol. 2018, 9, 85. DOI: 10.3389/fphar.2018.00085.
- Faqueti, L. G.; Brieudes, V.; Halabalaki, M.; Skaltsounis, A. L.; Nascimento, L. F.; Barros, W. M.; Santos, A. R. S.; Biavatti, M. W. Antinociceptive and Anti-Inflammatory Activities of Standardized Extract of Polymethoxyflavones from Ageratum Conyzoides. J. Ethnopharmacol. 2016, 194, 369–377. DOI: 10.1016/j.jep.2016.09.025.
- Dong, Y.; Cao, A.; Shi, J.; Yin, P.; Wang, L.; Ji, G.; Xie, J.; Wu, D. Tangeretin, a Citrus Polymethoxyflavonoid, Induces Apoptosis of Human Gastric Cancer AGS Cells Through Extrinsic and Intrinsic Signaling Pathways. Oncol. Rep. 2014, 31(4), 1788–1794. DOI: 10.3892/or.2014.3034.
- Ting, Y.; Jiang, Y.; Ho, C. T.; Huang, Q. Common Delivery Systems for Enhancing in vivo Bioavailability and Biological Efficacy of Nutraceuticals. J. Funct. Foods. 2014, 7, 112–128. DOI: 10.1016/j.jff.2013.12.010.
- Rein, M. J.; Renouf, M.; Cruz‐hernandez, C.; Actis‐goretta, L.; Thakkar, S. K.; da Silva Pinto, M. Bioavailability of Bioactive Food Compounds: A Challenging Journey to Bioefficacy. Br. J. Clin. Pharmacol. 2013, 75(3), 588–602. DOI: 10.1111/j.1365-2125.2012.04425.x.
- Zhang, M.; Zhu, S.; Yang, W.; Huang, Q.; Ho, C. T. The Biological Fate and Bioefficacy of Citrus Flavonoids: Bioavailability, Biotransformation, and Delivery Systems. Food & Function. 2021, 12(8), 3307–3323. DOI: 10.1039/D0FO03403G.
- Alfei, S.; Schito, A. M.; Zuccari, G. Nanotechnological Manipulation of Nutraceuticals and Phytochemicals for Healthy Purposes: Established Advantages Vs. Still Undefined Risks. Poly. 2021, 13(14), 2262. DOI: 10.3390/polym13142262.
- Xu, Y.; Li, Y.; Xie, J.; Xie, L.; Mo, J.; Chen, W. Bioavailability, Absorption, and Metabolism of Pelargonidin-Based Anthocyanins Using Sprague–Dawley Rats and Caco-2 Cell Monolayers. J. Agric. Food Chem. 2021, 69(28), 7841–7850. DOI: 10.1021/acs.jafc.1c00257.
- Zhang, M.; Pan, Y.; Dong, Q.; Tang, X.; Xin, Y.; Yin, B.; Zhu, J.; Kou, X.; Ho, C. T.; Huang, Q. Development of Organogel-Based Emulsions to Enhance the Loading and Bioaccessibility of 5-Demethylnobiletin. Food Res. Int. 2021, 148, 110592. DOI: 10.1016/j.foodres.2021.110592.
- Singh, A. P.; Kandpal, J. B.; Sharma, R. K.; Chitme, H. Nobiletin a Biologically Active Phytoconstituent: Systematic Review. J. Biol. Act. Prod. Nat. 2021, 11(3), 204–211. DOI: 10.1080/22311866.2021.1920461.
- Wijaya, W.; Zheng, H.; Zheng, T.; Su, S.; Patel, A. R.; Van der Meeren, P.; Huang, Q. Improved Bioaccessibility of Polymethoxyflavones Loaded into High Internal Phase Emulsions Stabilized by Biopolymeric Complexes: A Dynamic Digestion Study via Tno’s Gastrointestinal Model. Curr. Res. Food Sci. 2020, 2, 11–19. DOI: 10.1016/j.crfs.2019.11.007.
- Ting, Y.; Jiang, Y.; Lan, Y.; Xia, C.; Lin, Z.; Rogers, M. A.; Huang, Q. Viscoelastic Emulsion Improved the Bioaccessibility and Oral Bioavailability of Crystalline Compound: A Mechanistic Study Using in vitro and in vivo Models. Mol. Pharmaceutics. 2015, 12(7), 2229–2236. DOI: 10.1021/mp5007322.
- Ting, Y.; Xia, Q.; Li, S.; Ho, C. T.; Huang, Q. Design of High-Loading and High-Stability Viscoelastic Emulsions for Polymethoxyflavones. Food Res. Int. 2013, 54(1), 633–640. DOI: 10.1016/j.foodres.2013.07.047.
- Bajraktari-Sylejmani, G.; Weiss, J. Potential Risk of Food-Drug Interactions: Citrus Polymethoxyflavones and Flavanones as Inhibitors of the Organic Anion Transporting Polypeptides (OATP) 1B1, 1B3, and 2B1. Eur. J. Drug Metab. Pharmacokinet. 2020, 45(6), 809–815. DOI: 10.1007/s13318-020-00634-4.
- Guo, J.; Tao, H.; Cao, Y.; Ho, C. T.; Jin, S.; Huang, Q. Prevention of Obesity and Type 2 Diabetes with Aged Citrus Peel (Chenpi) Extract. J. Agric. Food Chem. 2016, 64(10), 2053–2061. DOI: 10.1021/acs.jafc.5b06157.
- Cao, X.; Ma, C.; Gao, Z.; Zheng, J.; He, L.; McClements, D. J.; Xiao, H. Characterization of the Interactions Between Titanium Dioxide Nanoparticles and Polymethoxyflavones Using Surface-Enhanced Raman Spectroscopy. J. Agric. Food Chem. 2016, 64(49), 9436–9441. DOI: 10.1021/acs.jafc.6b03906.
- Faqueti, L. G.; Sandjo, L. P.; Biavatti, M. W. Simultaneous Identification and Quantification of Polymethoxyflavones, Coumarin and Phenolic Acids in Ageratum Conyzoides by UPLC-ESI-QToF-MS and UPLC-PDA. J. Pharm. Biomed. Anal. 2017, 145, 621–628. DOI: 10.1016/j.jpba.2017.07.034.
- Ververidis, F.; Trantas, E.; Douglas, C.; Vollmer, G.; Kretzschmar, G.; Panopoulos, N. Biotechnology of Flavonoids and Other Phenylpropanoid-Derived Natural Products. Part I: Chemical Diversity, Impacts on Plant Biology and Human Health. Biotechnol. J. 2007, 2(10), 1214–1234. DOI: 10.1002/biot.200700084.
- Xing, T. T.; Zhao, X. J.; Zhang, Y. D.; Li, Y. F. Fast Separation and Sensitive Quantitation of Polymethoxylated Flavonoids in the Peels of Citrus Using UPLC-Q-TOF-MS. J. Agric. Food Chem. 2017, 65(12), 2615–2627. DOI: 10.1021/acs.jafc.6b05821.
- Xu, J. J.; Wu, X.; Li, M. M.; Li, G. Q.; Yang, Y. T.; Luo, H. J.; Huang, W. H.; Chung, H. Y.; Ye, W. C.; Wang, G. C. L., et al. Antiviral Activity of Polymethoxylated Flavones from “Guangchenpi”, the Edible and Medicinal Pericarps of Citrus Reticulata ‘Chachi’. J. Agric. Food Chem. 2014, 62(10), 2182–2189. DOI: 10.1021/jf404310y.
- Salehi, B.; Jornet, P. L.; López, E. P. F.; Calina, D.; Sharifi-Rad, M.; Ramírez-Alarcón, K.; Forman, K.; Fernández, M.; Martorell, M.; Setzer, W. N., et al. Plant-Derived Bioactives in Oral Mucosal Lesions: A Key Emphasis to Curcumin, Lycopene, Chamomile, Aloe Vera, Green Tea and Coffee Properties. Biomol. 2019, 9(3), 106. DOI: 10.3390/biom9030106.
- Docea, A. O.; Mitrut, P.; Grigore, D.; Pirici, D.; Calina, D. C.; Gofita, E. Immunohistochemical Expression of TGF Beta (TGF-Beta), TGF Beta Receptor 1 (TGFBR1), and Ki67 in Intestinal Variant of Gastric Adenocarcinomas. Rom. J. Morphol. Embryol. 2012, 53(3), 683–692.
- Tung, Y. C.; Chou, Y. C.; Hung, W. L.; Cheng, A. C.; Yu, R. C.; Ho, C. T.; Pan, M. H. Polymethoxyflavones: Chemistry and Molecular Mechanisms for Cancer Prevention and Treatment. Curr. Pharmacol. Rep. 2019, 5(2), 98–113. DOI: 10.1007/s40495-019-00170-z.
- Surichan, S.; Arroo, R. R.; Ruparelia, K.; Tsatsakis, A. M.; Androutsopoulos, V. P. Nobiletin Bioactivation in MDA-MB-468 Breast Cancer Cells by Cytochrome P450 CYP1 Enzymes. Food Chem. Toxicol. 2018, 113, 228–235. DOI: 10.1016/j.fct.2018.01.047.
- Zhu, W. B.; Xiao, N.; Liu, X. J. Dietary Flavonoid Tangeretin Induces Reprogramming of Epithelial to Mesenchymal Transition in Prostate Cancer Cells by Targeting the PI3K/Akt/mTor Signaling Pathway. Oncol. Letters. 2018, 15(1), 433–440. DOI: 10.3892/ol.2017.7307.
- Li, J.; Li, Z.; Zheng, F. Nobiletin Inhibits Proliferation, Invasion, Migration and Angiogenesis in Colorectal Cancer Cells. J. Biomater. Tissue Eng. 2019, 9(5), 662–667. DOI: 10.1166/jbt.2019.2022.
- Goh, J. X. H.; Tan, L. T. H.; Goh, J. K.; Chan, K. G.; Pusparajah, P.; Lee, L. H.; Goh, B. H. Nobiletin and Derivatives: Functional Compounds from Citrus Fruit Peel for Colon Cancer Chemoprevention. Cancers. 2019, 11(6), 867. DOI: 10.3390/cancers11060867.
- Li, G. J.; Wang, J.; Cheng, Y. J.; Tan, X.; Zhai, Y. L.; Wang, Q.; Gao, F. J.; Liu, G. L.; Zhao, X.; Wang, H. Prophylactic Effects of Polymethoxyflavone-Rich Orange Peel Oil on Nω-Nitro-L-Arginine-Induced Hypertensive Rats. Appl. Sci. 2018, 8(5), 752. DOI: 10.3390/app8050752.
- Wu, J. C.; Tsai, M. L.; Lai, C. S.; Lo, C. Y.; Ho, C. T.; Wang, Y. J.; Pan, M. H. Polymethoxyflavones Prevent Benzo [A] Pyrene/Dextran Sodium Sulfate‐induced Colorectal Carcinogenesis Through Modulating Xenobiotic Metabolism and Ameliorate Autophagic Defect in ICR Mice. Int. J. Cancer. 2018, 142(8), 1689–1701. DOI: 10.1002/ijc.31190.
- Pereira, C. V.; Duarte, M.; Silva, P.; Bento da Silva, A.; Duarte, C. M.; Cifuentes, A.; García-Cañas, V.; Bronze, M. R.; Albuquerque, C.; Serra, A. T. Polymethoxylated Flavones Target Cancer Stemness and Improve the Antiproliferative Effect of 5-Fluorouracil in a 3D Cell Model of Colorectal Cancer. Nutrients. 2019, 11(2), 326. DOI: 10.3390/nu11020326.
- Li, Y. R.; Li, S.; Ho, C. T.; Chang, Y. H.; Tan, K. T.; Chung, T. W.; Wang, B. Y.; Chen, Y. K.; Lin, C. C. Tangeretin Derivative, 5-Acetyloxy-6, 7, 8, 4′-Tetramethoxyflavone Induces G2/M Arrest, Apoptosis and Autophagy in Human Non-Small Cell Lung Cancer Cells in vitro and in vivo. Cancer. Biol & Therapy. 2016, 17(1), 48–64. DOI: 10.1080/15384047.2015.1108491.
- Zhang, H.; Tian, G.; Zhao, C.; Han, Y.; DiMarco-Crook, C.; Lu, C.; Bao, Y.; Li, C.; Xiao, H.; Zheng, J. Characterization of Polymethoxyflavone Demethylation During Drying Processes of Citrus Peels. Food & Function. 2019, 10(9), 5707–5717. DOI: 10.1039/C9FO01053J.
- Tan, K. T.; Li, S.; Li, Y. R.; Cheng, S. L.; Lin, S. H.; Tung, Y. T. Synergistic Anticancer Effect of a Combination of Paclitaxel and 5-Demethylnobiletin Against Lung Cancer Cell Line in vitro and in vivo. Appl. Biochem. Biotechnol. 2019, 187, 1328–1343. DOI: 10.1007/s12010-018-2869-1.
- Abe, S.; Hirose, S.; Nishitani, M.; Yoshida, I.; Tsukayama, M.; Tsuji, A.; Yuasa, K. Citrus Peel Polymethoxyflavones, Sudachitin and Nobiletin, Induce Distinct Cellular Responses in Human Keratinocyte HaCat Cells. Biosci. Biotechnol. Biochem. 2018, 82(12), 2064–2071. DOI: 10.1080/09168451.2018.1514246.
- Yoshizaki, N.; Hashizume, R.; Masaki, H. A Polymethoxyflavone Mixture Extracted from Orange Peels, Mainly Containing Nobiletin3, 3’, 4’, 5, 6, 7, 8-Heptamethoxyflavone and Tangeretin, Suppresses Melanogenesis Through the Acidification of Cell Organelles, Including Melanosomes. J. Dermatol. Sci. 2017, 88(1), 78–84. DOI: 10.1016/j.jdermsci.2017.06.008.
- Sousa, D. P.; Pojo, M.; Pinto, A. T.; Leite, V.; Serra, A. T.; Cavaco, B. M. Nobiletin Alone or in Combination with Cisplatin Decreases the Viability of Anaplastic Thyroid Cancer Cell Lines. Nutr. Cancer. 2020, 72(2), 352–363. DOI: 10.1080/01635581.2019.1634745.
- Wei, G. J.; Chao, Y. H.; Tung, Y. C.; Wu, T. Y.; Su, Z. Y. A Tangeretin Derivative Inhibits the Growth of Human Prostate Cancer LNCaP Cells by Epigenetically Restoring P21 Gene Expression and Inhibiting Cancer Stem-Like Cell Proliferation. Aaps. J. 2019, 21(5), 1–12. DOI: 10.1208/s12248-019-0345-7.
- Lin, C. C.; Chen, K. B.; Tsai, C. H.; Tsai, F. J.; Huang, C. Y.; Tang, C. H.; Yang, J. S.; Hsu, Y. M.; Peng, S. F.; Chung, J. G. Casticin Inhibits Human Prostate Cancer DU 145 Cell Migration and Invasion via Ras/Akt/Nf‐κb Signaling Pathways. J. Food Biochem. 2019, 43(7), e12902. DOI: 10.1111/jfbc.12902.
- Docea, A. O.; Calina, D.; Buga, A. M.; Zlatian, O.; Paoliello, M. M. B.; Mogosanu, G. D.; Streba, C. T.; Popescu, E. L.; Stoica, A. E.; Bîrcă, A. C., et al. The Effect of Silver Nanoparticles on Antioxidant/pro-Oxidant Balance in a Murine Model. Int. J. Mol. Sci. 2020, 21(4), 1233. DOI: 10.3390/ijms21041233.
- Padureanu, R.; Albu, C. V.; Mititelu, R. R.; Bacanoiu, M. V.; Docea, A. O.; Calina, D.; Padureanu, V.; Olaru, G.; Sandu, R. E.; Malin, R. D., et al. Oxidative Stress and Inflammation Interdependence in Multiple Sclerosis. J. Clin. Med. 2019, 8(11), 1815. DOI: 10.3390/jcm8111815.
- Mititelu, R. R.; Pădureanu, R.; Băcănoiu, M.; Pădureanu, V.; Docea, A. O.; Calina, D.; Barbulescu, A. L.; Buga, A. M. Inflammatory and Oxidative Stress Markers—mirror Tools in Rheumatoid Arthritis. Biomed. 2020, 8(5), 125. DOI: 10.3390/biomedicines8050125.
- Salehi, B.; Sestito, S.; Rapposelli, S.; Peron, G.; Calina, D.; Sharifi-Rad, M.; Sharopov, F.; Martins, N.; Sharifi-Rad, J. Epibatidine: A Promising Natural Alkaloid in Health. Biomol. 2018, 9(1), 6. DOI: 10.3390/biom9010006.
- Lakshmi, A.; Subramanian, S. P. Tangeretin Ameliorates Oxidative Stress in the Renal Tissues of Rats with Experimental Breast Cancer Induced by 7, 12-Dimethylbenz [A] Anthracene. Toxicol. Lett. 2014, 229(2), 333–348. DOI: 10.1016/j.toxlet.2014.06.845.
- Daniyal, A.; Sue, E. S.; Ashok Kumar, G. How Effective are Antioxidant Supplements in Obesity and Diabetes? Med. Prin and Practice. 2015, 24(3), 201–215. DOI: 10.1159/000375305.
- Chen, X. Y.; Zhang, J.; Zhu, J. S.; Chen, X.; Han, X.; Yu, J.; Wang, W.; Liang, L.; Liu, Z.; Zheng, Y., et al. Epigenetically Upregulated Oncoprotein PLCE1 Drives Esophageal Carcinoma Angiogenesis and Proliferation via Activating the PI-PLCε-NF-Κb Signaling Pathway and VEGF-C/ Bcl-2 Expression. Mol. Cancer. 2019, 18(1), 1–9. DOI: 10.1186/s12943-018-0930-x.
- Wang, Y.; Chen, Y.; Zhang, H.; Chen, J.; Cao, J.; Chen, Q.; Li, X.; Sun, C. Polymethoxyflavones from Citrus Inhibited Gastric Cancer Cell Proliferation Through Inducing Apoptosis by Upregulating RARβ, Both in vitro and in vivo. Food Chem. Toxicol. 2020, 146, 111811. DOI: 10.1016/j.fct.2020.111811.
- Liang, F.; Fang, Y.; Cao, W.; Zhang, Z.; Pan, S.; Xu, X. Attenuation of Tert-Butyl Hydroperoxide (T-BHP)-Induced Oxidative Damage in HepG2 Cells by Tangeretin: Relevance of the Nrf2–ARE and MAPK Signaling Pathways. J. Agric. Food Chem. 2018, 66(25), 6317–6325. DOI: 10.1021/acs.jafc.8b01875.
- Hagenlocher, Y.; Feilhauer, K.; Schäffer, M.; Bischoff, S. C.; Lorentz, A. Citrus Peel Polymethoxyflavones Nobiletin and Tangeretin Suppress LPS-And IgE-Mediated Activation of Human Intestinal Mast Cells. Eur. J. Nutr. 2017, 56(4), 1609–1620. DOI: 10.1007/s00394-016-1207-z.
- Lai, C. S.; Tsai, M. L.; Cheng, A. C.; Li, S.; Lo, C. Y.; Wang, Y.; Xiao, H.; Ho, C. T.; Wang, Y. J.; Pan, M. H. Chemoprevention of Colonic Tumorigenesis by Dietary Hydroxylated Polymethoxyflavones in Azoxymethane‐treated Mice. Mole. Nutri & Food Res. 2011, 55(2), 278–290. DOI: 10.1002/mnfr.201000224.
- Wu, X.; Song, M.; Rakariyatham, K.; Zheng, J.; Guo, S.; Tang, Z.; Zhou, S.; Xiao, H. Anti-Inflammatory Effects of 4′-Demethylnobiletin, a Major Metabolite of Nobiletin. J. Funct. Foods. 2015, 19, 278–287. DOI: 10.1016/j.jff.2015.09.035.
- Kim, T. W.; Lee, D. R.; Choi, B. K.; Kang, H. K.; Jung, J. Y.; Lim, S. W.; Yang, S. H.; Suh, J. W. Hepatoprotective Effects of Polymethoxyflavones Against Acute and Chronic Carbon Tetrachloride Intoxication. Food Chem. Toxicol. 2016, 91, 91–99. DOI: 10.1016/j.fct.2016.03.004.
- Devi, A. D.; Devi, O. I.; Singh, T. C.; Singh, E. J. A Study of Aromatic Plant Species Especially in Thoubal District, Manipur, North East India. Int. J. Sci. Res. Publ. 2014, 4, 2250–3153.
- Lee, Y. Y.; Lee, E. J.; Park, J. S.; Jang, S. E.; Kim, D. H.; Kim, H. S. Anti-Inflammatory and Antioxidant Mechanism of Tangeretin in Activated Microglia. J. Neuroimmune Pharmacol. 2016, 11(2), 294–305. DOI: 10.1007/s11481-016-9657-x.
- Shu, Z.; Yang, B.; Zhao, H.; Xu, B.; Jiao, W.; Wang, Q.; Wang, Z.; Kuang, H. Tangeretin Exerts Anti-Neuroinflammatory Effects via NF-Κb Modulation in Lipopolysaccharide-Stimulated Microglial Cells. Int. Immunopharmacol. 2014, 19(2), 275–282. DOI: 10.1016/j.intimp.2014.01.011.
- Gosslau, A.; Chen, K. Y.; Ho, C. T.; Li, S. Anti-Inflammatory Effects of Characterized Orange Peel Extracts Enriched with Bioactive Polymethoxyflavones. Food Sci. Hum. Wellness. 2014, 3(1), 26–35. DOI: 10.1016/j.fshw.2014.02.002.
- Zhang, D.; Hou, L.; Peng, W. Tangeritin Attenuates Oxidative Stress, Apoptosis and Inflammation in Cadmium-Induced Cardiotoxicity in Rats by Activating Nrf2 Signaling Pathway. Trop. J. Pharm. Res. 2018, 17(12), 2421–2426. DOI: 10.4314/tjpr.v17i12.16.
- Sharifi-Rad, J.; Rodrigues, C. F.; Sharopov, F.; Docea, A. O.; Can Karaca, A.; Sharifi-Rad, M.; Kahveci Karıncaoglu, D.; Gülseren, G.; Şenol, E.; Demircan, E., et al. Diet, Lifestyle and Cardiovascular Diseases: Linking Pathophysiology to Cardioprotective Effects of Natural Bioactive Compounds. Int. J. Environ. Res. Public Health. 2020, 17(7), 2326. DOI: 10.3390/ijerph17072326.
- Sharifi-Rad, M.; Anil Kumar, N. V.; Zucca, P.; Varoni, E. M.; Dini, L.; Panzarini, E.; Rajkovic, J.; Tsouh Fokou, P. V.; Azzini, E.; Peluso, I., et al. Lifestyle, Oxidative Stress, and Antioxidants: Back and Forth in the Pathophysiology of Chronic Diseases. Front. Physiol. 2020, 11, 694. DOI: 10.3389/fphys.2020.00694.
- Shammugasamy, B.; Valtchev, P.; Dong, Q.; Dehghani, F. Effect of Citrus Peel Extracts on the Cellular Quiescence of Prostate Cancer Cells. Food & Function. 2019, 10(6), 3727–3737. DOI: 10.1039/C9FO00455F.
- Zhou, M. X.; Li, G. H.; Wu, X. Y.; Sun, L.; Li, Y. R.; Yang, W. J.; Ren, D. M.; Wang, X. N.; Xiang, L.; Lou, H. X., et al. (2S)-5,6,7,3′,4′-Pentamethoxyflavanone, a Citrus Polymethoxyflavone Ameliorates Arsenic- and Cigarette Smoke Extract-Induced Cytotoxicity via Activating Nrf2-Mediated Defense System. J. Funct. Foods. 2019, 54(54), 337–347. DOI: 10.1016/j.jff.2019.01.019.
- Lakshmi, A.; Subramanian, S. Chemotherapeutic Effect of Tangeretin, a Polymethoxylated Flavone Studied in 7, 12-Dimethylbenz (A) Anthracene Induced Mammary Carcinoma in Experimental Rats. Biochimie. 2014, 99, 96–109. DOI: 10.1016/j.biochi.2013.11.017.
- Salehi, B.; Prakash Mishra, A.; Nigam, M.; Karazhan, N.; Shukla, I.; Kiełtyka‐dadasiewicz, A.; Sawicka, B.; Głowacka, A.; Abu‐darwish, M. S.; Hussein Tarawneh, A., et al. Ficus Plants: State of the Art from a Phytochemical, Pharmacological, and Toxicological Perspective. Phytotherapy. Res. 2021, 35(3), 1187–1217. DOI: 10.1002/ptr.6884.
- Ochiai, M.; Takeuchi, T.; Nozaki, T.; Ishihara, K. O.; Matsuo, T. Kaempferia Parviflora Ethanol Extract, a Peroxisome Proliferator‐activated Receptor γ Ligand‐binding Agonist, Improves Glucose Tolerance and Suppresses Fat Accumulation in Diabetic NSY Mice. J. Food Sci. 2019, 84(2), 339–348. DOI: 10.1111/1750-3841.14437.
- Wang, L.; Wang, J.; Fang, L.; Zheng, Z.; Zhi, D.; Wang, S.; Li, S.; Ho, C. T.; Zhao, H. Anticancer Activities of Citrus Peel Polymethoxyflavones Related to Angiogenesis and Others. Biomed Res. Int. 2014, 2014, 1–10. DOI: 10.1155/2014/453972.
- Guo, J. Application of 5-oh polymethoxyflavones and safe mitochondrial uncouplers in preventing and treating type 2 diabetes and fatty liver diseases. (Doctoral dissertation, Rutgers University-School of Graduate Studies). 2019.
- Fayek, N. M.; El-Shazly, A. H.; Abdel-Monem, A. R.; Moussa, M. Y.; Abd-Elwahab, S. M.; El-Tanbouly, N. D. Comparative Study of the Hypocholesterolemic, Antidiabetic Effects of Four Agro-Waste Citrus Peels Cultivars and Their HPLC Standardization. Rev. Bras. Farmacogn. 2017, 27(4), 488–494. DOI: 10.1016/j.bjp.2017.01.010.
- Keshtkar, S.; Kaviani, M.; Jabbarpour, Z.; Geramizadeh, B.; Motevaseli, E.; Nikeghbalian, S.; Shamsaeefar, A.; Motazedian, N.; Al-Abdullah, I. H.; Ghahremani, M. H., et al. Protective Effect of Nobiletin on Isolated Human Islets Survival and Function Against Hypoxia and Oxidative Stress-Induced Apoptosis. Sci. Rep. 2019, 9(1), 1–13. DOI: 10.1038/s41598-019-48262-6.
- Yuk, T.; Kim, Y.; Yang, J.; Sung, J.; Jeong, H. S.; Lee, J. Nobiletin Inhibits Hepatic Lipogenesis via Activation of AMP-Activated Protein Kinase. Evid. Based Complement. Altern. Med. 2018, 2018, 1–8. DOI: 10.1155/2018/7420265.
- Feng, K.; Zhu, X.; Chen, T.; Peng, B.; Lu, M.; Zheng, H.; Huang, Q.; Ho, C. T.; Chen, Y.; Cao, Y. Prevention of Obesity and Hyperlipidemia by Heptamethoxyflavone in High-Fat Diet-Induced Rats. J. Agric. Food Chem. 2019, 67(9), 2476–2489. DOI: 10.1021/acs.jafc.8b05632.
- Pan, M. H.; Yang, G.; Li, S.; Li, M. Y.; Tsai, M. L.; Wu, J. C.; Badmaev, V.; Ho, C. T.; Lai, C. S. Combination of Citrus Polymethoxyflavones, Green Tea Polyphenols, and Lychee Extracts Suppresses Obesity and Hepatic Steatosis in High‐fat Diet Induced Obese Mice. Mole.Nutri & Food Res. 2017, 61(11), 1601104. DOI: 10.1002/mnfr.201601104.
- Guo, J.; Cao, Y.; Ho, C. T.; Jin, S.; Huang, Q. Aged Citrus Peel (Chenpi) Extract Reduces Lipogenesis in Differentiating 3T3-L1 Adipocytes. J. Funct. Foods. 2017, 34, 297–303. DOI: 10.1016/j.jff.2017.04.042.
- Yun, Y. R.; Lee, J. E. Alliin, Capsaicin, and Gingerol Attenuate Endoplasmic Reticulum Stress-Induced Hepatic Steatosis in HepG2 Cells and C57BL/6N Mice. J. Funct. Foods. 2022, 95, 105186. DOI: 10.1016/j.jff.2022.105186.
- Tung, Y. C.; Li, S.; Huang, Q.; Hung, W. L.; Ho, C. T.; Wei, G. J.; Pan, M. H. 5-Demethylnobiletin and 5-Acetoxy-6, 7, 8, 3′, 4′-Pentamethoxyflavone Suppress Lipid Accumulation by Activating the LKB1-AMPK Pathway in 3T3-L1 Preadipocytes and High Fat Diet-Fed C57BL/6 Mice. J. Agric. Food Chem. 2016, 64(16), 3196–3205. DOI: 10.1021/acs.jafc.6b00706.
- Tsatsakis, A.; Docea, A. O.; Calina, D.; Tsarouhas, K.; Zamfira, L. M.; Mitrut, R.; Sharifi-Rad, J.; Kovatsi, L.; Siokas, V.; Dardiotis, E., et al. A Mechanistic and Pathophysiological Approach for Stroke Associated with Drugs of Abuse. J. Clin. Med. 2019, 8(9), 1295. DOI: 10.3390/jcm8091295.
- Salehi, B.; Sharifi-Rad, J.; Cappellini, F.; Reiner, Ž.; Zorzan, D.; Imran, M.; Sener, B.; Kilic, M.; El-Shazly, M.; Fahmy, N. M., et al. The Therapeutic Potential of Anthocyanins: Current Approaches Based on Their Molecular Mechanism of Action. Front. Pharmacol. 2020, 11, 1300. DOI: 10.3389/fphar.2020.01300.
- Kang, S. I.; Shin, H. S.; Kim, S. J. Sinensetin Enhances Adipogenesis and Lipolysis by Increasing Cyclic Adenosine Monophosphate Levels in 3T3-L1 Adipocytes. Biol. Pharm. Bull. 2015, 38(4), 552–558. DOI: 10.1248/bpb.b14-00700.
- Adebiyi, A. O.; Adebiyi, O. O.; Owira, P. M. Naringin Mitigates Cardiac Hypertrophy by Reducing Oxidative Stress and Inactivating C-Jun Nuclear Kinase-1 Protein in Type I Diabetes. J. Cardiovasc. Pharmacol. 2016, 67(2), 136–144. DOI: 10.1097/FJC.0000000000000325.
- Jenie, R. I.; Setiawan, I. M.; Jenie, I. M.; Muflikhasari, H. A. Cytoprotective Effect of Tangeretin in Hydrogen Peroxyde-Induced oxidative Stress on Human Umbilical Vein Endothelial Cells (HUVECs). J. Ilm. Farm. 2015, 11(1), 16–19. DOI: 10.20885/jif.vol11.iss1.art3.
- Kim, J. J.; Kim, K.; Jung, Y. R.; Bian, Y.; Ngo, T.; Bae, O. N.; Lim, K. M.; Chung, J. H. Co-Existence of Hypertensive and Anti-Hypertensive Constituents, Synephrine, and Nobiletin in Citrus Unshiu Peel. Molecules. 2019, 24(7), 1197. DOI: 10.3390/molecules24071197.
- Cummings, J. L.; Tong, G.; Ballard, C. Treatment Combinations for Alzheimer’s Disease: Current and Future Pharmacotherapy Options. J. Alzheimer’s dis. 2019, 67(3), 779–794. DOI: 10.3233/JAD-180766.
- Calina, D.; Buga, A. M.; Mitroi, M.; Buha, A.; Caruntu, C.; Scheau, C.; Bouyahya, A.; El Omari, N.; El Menyiy, N.; Docea, A. O. The Treatment of Cognitive, Behavioural and Motor Impairments from Brain Injury and Neurodegenerative Diseases Through Cannabinoid System Modulation—evidence from in vivo Studies. J. Clin. Med. 2020, 9(8), 2395. DOI: 10.3390/jcm9082395.
- Youn, K.; Yu, Y.; Lee, J.; Jeong, W. S.; Ho, C. T.; Jun, M. Polymethoxyflavones: Novel β-Secretase (BACE1) Inhibitors from Citrus Peels. Nutrients. 2017, 9(9), 973. DOI: 10.3390/nu9090973.
- Nakajima, A.; Aoyama, Y.; Shin, E. J.; Nam, Y.; Kim, H. C.; Nagai, T.; Yokosuka, A.; Mimaki, Y.; Yokoi, T.; Ohizumi, Y., et al. Nobiletin, a Citrus Flavonoid, Improves Cognitive Impairment and Reduces Soluble Aβ Levels in a Triple Transgenic Mouse Model of Alzheimer’s Disease (3xtg-AD). Behav. Brain Res. 2015, 289, 69–77. DOI: 10.1016/j.bbr.2015.04.028.
- Youn, K.; Lee, S.; Jun, M. Discovery of Nobiletin from Citrus Peel as a Potent Inhibitor of β-Amyloid Peptide Toxicity. Nutrients. 2019, 11(11), 2648. DOI: 10.3390/nu11112648.
- Kawahata, I.; Yoshida, M.; Sun, W.; Nakajima, A.; Lai, Y.; Osaka, N.; Matsuzaki, K.; Yokosuka, A.; Mimaki, Y.; Naganuma, A., et al. Potent Activity of Nobiletin-Rich Citrus Reticulata Peel Extract to Facilitate cAmp/PKA/ERK/CREB Signaling Associated with Learning and Memory in Cultured Hippocampal Neurons: Identification of the Substances Responsible for the Pharmacological Action. J. Neural Trans. 2013, 120(10), 1397–1409. DOI: 10.1007/s00702-013-1025-x.
- Nakajima, A.; Ohizumi, Y. Potential Benefits of Nobiletin, a Citrus Flavonoid, Against Alzheimer’s Disease and Parkinson’s Disease. Int. J. Mol. Sci. 2019, 20(14), 3380. DOI: 10.3390/ijms20143380.
- Cioboată, R.; Găman, A.; Traşcă, D.; Ungureanu, A.; Docea, A. O.; Tomescu, P.; Gherghina, F.; Arsene, A. L.; Badiu, C.; Tsatsakis, A. M., et al. Pharmacological Management of Non-Alcoholic Fatty Liver Disease: Atorvastatin versus Pentoxifylline. Exp. Ther. Med. 2017, 13(5), 2375–2381. DOI: 10.3892/etm.2017.4256.
- Han, H. Y.; Lee, S. K.; Choi, B. K.; Lee, D. R.; Lee, H. J.; Kim, T. W. Preventive Effect of Citrus Aurantium Peel Extract on High-Fat Diet-Induced Non-Alcoholic Fatty Liver in Mice. Biol. Pharm. Bull. 2019, 42(2), 255–260. DOI: 10.1248/bpb.b18-00702.
- Yen, J. H.; Weng, C. Y.; Li, S.; Lo, Y. H.; Pan, M. H.; Fu, S. H.; Ho, C. T.; Wu, M. J. Citrus Flavonoid 5‐demethylnobiletin Suppresses Scavenger Receptor Expression in THP‐1 Cells and Alters Lipid Homeostasis in HepG2 Liver Cells. Mole. Nutri & Food Res. 2011, 55(5), 733–748. DOI: 10.1002/mnfr.201000226.
- Cushnie, T. T.; Hamilton, V. E.; Lamb, A. J. Assessment of the Antibacterial Activity of Selected Flavonoids and Consideration of Discrepancies Between Previous Reports. Microbiol. Res. 2003, 158(4), 281–289. DOI: 10.1078/0944-5013-00206.
- Cushnie, T. T.; Lamb, A. J. Antimicrobial Activity of Flavonoids. Int. J. Antimicrob. Agents. 2005, 26(5), 343–356. DOI: 10.1016/j.ijantimicag.2005.09.002.
- Alvarez, M. A.; Debattista, N. B.; Pappano, N. B. Antimicrobial Activity and Synergism of Some Substituted Flavonoids. Folia. microbiologica. 2008, 53(1), 23. DOI: 10.1007/s12223-008-0003-4.
- Basile, A.; Giordano, S.; López-Sáez, J. A.; Cobianchi, R. C. Antibacterial Activity of Pure Flavonoids Isolated from Mosses. Phytochemistry. 1999, 52(8), 1479–1482. DOI: 10.1016/S0031-9422(99)00286-1.
- Basile, A.; Sorbo, S.; Lopez-Saez, J. A.; Cobianchi, R. C. Effects of Seven Pure Flavonoids from Mosses on Germination and Growth of Tortula Muralis Hedw.(bryophyta) and Raphanus Sativus L.(magnoliophyta). Phytochemistry. 2003, 62(7), 1145–1151. DOI: 10.1016/S0031-9422(02)00659-3.
- Mori, A.; Nishino, C.; Enoki, N.; Tawata, S. Antibacterial Activity and Mode of Action of Plant Flavonoids Against Proteus Vulgaris and Staphylococcus Aureus. Phytochemistry. 1987, 26(8), 2231–2234. DOI: 10.1016/S0031-9422(00)84689-0.
- Cushnie, T. T.; Lamb, A. J. Detection of Galangin-Induced Cytoplasmic Membrane Damage in Staphylococcus Aureus by Measuring Potassium Loss. J. Ethnopharmacol. 2005, 101(1–3), 243–248. DOI: 10.1016/j.jep.2005.04.014.
- Butler, E. E.; Hine, R. B. Use of Novobiocin for Isolation of Fungi from Soil. Soil. Sci. 1958, 85(5), 250–254. DOI: 10.1097/00010694-195805000-00003.
- Tanaka, H.; Sato, M.; Fujiwara, S.; Hirata, M.; Etoh, H.; Takeuchi, H. Antibacterial Activity of Isoflavonoids Isolated from Erythrina Variegata Against Methicillin‐resistant Staphylococcus Aureus. Lett Appl. Microbiol. 2002, 35(6), 494–498. DOI: 10.1046/j.1472-765X.2002.01222.x.
- Chew, Y. L.; Ling Chan, E. W.; Tan, P. L.; Lim, Y. Y.; Stanslas, J.; Goh, J. K. Assessment of Phytochemical Content, Polyphenolic Composition, Antioxidant and Antibacterial Activities of Leguminosae Medicinal Plants in Peninsular Malaysia. BMC Complementary Altern. Med. 2011, 11(1), 1–10. DOI: 10.1186/1472-6882-11-12.
- Tsuchiya, H.; Sato, M.; Miyazaki, T.; Fujiwara, S.; Tanigaki, S.; Ohyama, M.; Tanaka, T.; Iinuma, M. Comparative Study on the Antibacterial Activity of Phytochemical Flavanones Against Methicillin-Resistant Staphylococcus Aureus. J. Ethnopharmacol. 1996, 50(1), 27–34. DOI: 10.1016/0378-8741(96)85514-0.
- García-Mediavilla, V.; Crespo, I.; Collado, P. S.; Esteller, A.; Sánchez-Campos, S.; Tuñón, M. J.; González-Gallego, J. The Anti-Inflammatory Flavones Quercetin and Kaempferol Cause Inhibition of Inducible Nitric Oxide Synthase, Cyclooxygenase-2 and Reactive C-Protein, and Down-Regulation of the Nuclear Factor kappaB Pathway in Chang Liver Cells. Eur. J. Pharmacol. 2007, 557(2–3), 221–229. DOI: 10.1016/j.ejphar.2006.11.014.
- Gado, D. A.; Abdalla, M. A.; Ahmed, A. S.; Madikizela, B.; Nkadimeng, S. M.; Ehlers, M. M.; McGaw, L. J. In vitro Antibacterial Activity of Loxostylis Alata Extracts and Isolated Compounds Against Salmonella Species. BMC Comple. Med and Therap. 2021, 21(1), 1–16. DOI: 10.1186/s12906-021-03292-4.
- Li, S.; Lo, C. Y.; Ho, C. T. Hydroxylated Polymethoxyflavones and Methylated Flavonoids in Sweet Orange (Citrus Sinensis) Peel. J. Agric. Food Chem. 2006, 54(12), 4176–4185. DOI: 10.1021/jf060234n.
- Manvar, D.; Mishra, M.; Kumar, S.; Pandey, V. N. Identification and Evaluation of Anti Hepatitis C Virus Phytochemicals from Eclipta Alba. J. Ethnopharmacol. 2012, 144(3), 545–554. DOI: 10.1016/j.jep.2012.09.036.
- Farkas, O.; Palócz, O.; Pászti-Gere, E.; Gálfi, P. Polymethoxyflavone Apigenin-Trimethylether Suppresses LPS-Induced Inflammatory Response in Nontransformed Porcine Intestinal Cell Line IPEC-J2. Oxid. Med. Cell. Longev. 2015, 2015, 1–10. DOI: 10.1155/2015/673847.
- Bachmetov, L.; Gal‐tanamy, M.; Shapira, A.; Vorobeychik, M.; Giterman‐galam, T.; Sathiyamoorthy, P.; Golan‐goldhirsh, A.; Benhar, I.; Tur‐kaspa, R.; Zemel, R. Suppression of Hepatitis C Virus by the Flavonoid Quercetin is Mediated by Inhibition of NS3 Protease Activity. J. Viral. Hepatitis. 2012, 19(2), e81–88. DOI: 10.1111/j.1365-2893.2011.01507.x.
- Huang, H. C.; Tao, M. H.; Hung, T. M.; Chen, J. C.; Lin, Z. J.; Huang, C. (−)-Epigallocatechin-3-Gallate Inhibits Entry of Hepatitis B Virus into Hepatocytes. Antiviral. Res. 2014, 111, 100–111. DOI: 10.1016/j.antiviral.2014.09.009.
- Zhang, Y.; Wang, R.; Wu, J.; Shen, Q. Characterization and Evaluation of Self-Microemulsifying Sustained-Release Pellet Formulation of Puerarin for Oral Delivery. Int. J. Pharmaceutics. 2012, 427(2), 337–344. DOI: 10.1016/j.ijpharm.2012.02.013.
- Le Lee, J.; Loe, M. W. C.; Lee, R. C. H.; Chu, J. J. H. Antiviral Activity of Pinocembrin Against Zika Virus Replication. Antiviral. Res. 2019, 167, 13–24. DOI: 10.1016/j.antiviral.2019.04.003.
- Ghildiyal, R.; Prakash, V.; Chaudhary, V. K.; Gupta, V.; Gabrani, R. Phytochemicals as Antiviral Agents: Recent Updates. Plant-Derived Bioactives: Production, Properties and Therapeutic Applications; Springer: Singapore, 2020, 279–295. ISBN: 978-981-15-1761 DOI: 10.1007/978-981-15-1761-7_12.
- Fahmy, N. M.; Al‐sayed, E.; Moghannem, S.; Azam, F.; El‐shazly, M.; Singab, A. N. Breaking Down the Barriers to a Natural Antiviral Agent: Antiviral Activity and Molecular Docking of Erythrina Speciosa Extract, Fractions, and the Major Compound. Chem. Biodivers. 2020, 17(2), e1900511. DOI: 10.1002/cbdv.201900511.
- Moghaddam, E.; Teoh, B. T.; Sam, S. S.; Lani, R.; Hassandarvish, P.; Chik, Z.; Yueh, A.; Abubakar, S.; Zandi, K. Baicalin, a Metabolite of Baicalein with Antiviral Activity Against Dengue Virus. Sci. Rep. 2014, 4(1), 1–8. DOI: 10.1038/srep05452.
- Li, S.; Hattori, T.; Kodama, E. N. Epigallocatechin Gallate Inhibits the HIV Reverse Transcription Step. Antivir. Chem. Chemother. 2011, 21(6), 239–243. DOI: 10.3851/IMP1774.
- Ahmed-Belkacem, A.; Guichou, J. F.; Brillet, R.; Ahnou, N.; Hernandez, E.; Pallier, C.; Pawlotsky, J. M. Inhibition of RNA Binding to Hepatitis C Virus RNA-Dependent RNA Polymerase: A New Mechanism for Antiviral Intervention. Nucleic Acids. Res. 2014, 42(14), 9399–9409. DOI: 10.1093/nar/gku632.
- Zetterberg, C.; Öfverholm, T. Carpal Tunnel Syndrome and Other Wrist/Hand Symptoms and Signs in Male and Female Car Assembly Workers. Int. J. Ind. Ergon. 1999, 23(3), 193–204. DOI: 10.1016/S0169-8141(97)00054-1.
- Yasuda, H.; Shima, N.; Nakagawa, N.; Yamaguchi, K.; Kinosaki, M.; Mochizuki, S. I.; Tomoyasu, A.; Yano, K.; Goto, M.; Murakami, A., et al. Osteoclast Differentiation Factor is a Ligand for Osteoprotegerin/osteoclastogenesis-Inhibitory Factor and is Identical to TRANCE/RANKL. Proce. National Acade. Sci, 1998, 95(7), 3597–3602.
- Tanabe, N.; Maeno, M.; Suzuki, N.; Fujisaki, K.; Tanaka, H.; Ogiso, B.; Ito, K. IL-1α Stimulates the Formation of Osteoclast-Like Cells by Increasing M-CSF and PGE2 Production and Decreasing OPG Production by Osteoblasts. Life. Sci. 2005, 77(6), 615–626. DOI: 10.1016/j.lfs.2004.10.079.
- Suda, K.; Udagawa, N.; Sato, N.; Takami, M.; Itoh, K.; Woo, J. T.; Takahashi, N.; Nagai, K. Suppression of Osteoprotegerin Expression by Prostaglandin E2 is Crucially Involved in Lipopolysaccharide-Induced Osteoclast Formation. J. Immunol. 2004, 172(4), 2504–2510. DOI: 10.4049/jimmunol.172.4.2504.
- Hirata, M.; Kobayashi, M.; Takita, M.; Matsumoto, C.; Miyaura, C.; Inada, M. Hyaluronan Inhibits Bone Resorption by Suppressing Prostaglandin E Synthesis in Osteoblasts Treated with Interleukin-1. Biochem. Biophys. Res. Commun. 2009, 381(2), 139–143. DOI: 10.1016/j.bbrc.2009.01.146.
- Juluri, R.; Prashanth, E.; Gopalakrishnan, D.; Kathariya, R.; Devanoorkar, A.; Viswanathan, V.; Romanos, G. E. Association of Postmenopausal Osteoporosis and Periodontal Disease: A Double-Blind Case-Control Study. J. Int. Oral. Health. 2015, 7(9), 119.
- Miyaura, C.; Kusano, K.; Masuzawa, T.; Chaki, O.; Onoe, Y.; Aoyagi, M.; Sasaki, T.; Tamura, T.; Koishihara, Y.; Ohsugi, Y., et al. Endogenous Bone‐resorbing Factors in Estrogen Deficiency: Cooperative Effects of IL‐1 and IL‐6. J. Bone Miner. Res. 1995, 10(9), 1365–1373. DOI: 10.1002/jbmr.5650100914.
- Lai, C. S.; Li, S.; Chai, C. Y.; Lo, C. Y.; Dushenkov, S.; Ho, C. T.; Pan, M. H.; Wang, Y. J. Anti-Inflammatory and Antitumor Promotional Effects of a Novel Urinary Metabolite, 3′, 4′-Didemethylnobiletin, Derived from Nobiletin. Carcinogenesis. 2008, 29(12), 2415–2424. DOI: 10.1093/carcin/bgn222.
- Matsumoto, C.; Inoue, H.; Tominari, T.; Watanabe, K.; Hirata, M.; Miyaura, C.; Inada, M. Heptamethoxyflavone, a Citrus Flavonoid, Suppresses Inflammatory Osteoclastogenesis and Alveolar Bone Resorption. Biosci. Biotechnol. Biochem. 2015, 79(1), 155–158. DOI: 10.1080/09168451.2014.952616.
- Tominari, T.; Hirata, M.; Matsumoto, C.; Inada, M.; Miyaura, C. Polymethoxy Flavonoids, Nobiletin and Tangeretin, Prevent Lipopolysaccharide-Induced Inflammatory Bone Loss in an Experimental Model for Periodontitis. J. Pharmacol. Sci. 2012, 119(4), 390–394. DOI: 10.1254/jphs.11188SC.
- Yoshizaki, N.; Fujii, T.; Hashizume, R.; Masaki, H. A Polymethoxyflavone Mixture, Extracted from Orange Peels, Suppresses the UVB‐induced Expression of MMP‐1. Experi. dermatol. 2016, 25, 52–56. DOI: 10.1111/exd.13087.
- Murakami, A.; Song, M.; Katsumata, S. I.; Uehara, M.; Suzuki, K.; Ohigashi, H. Citrus Nobiletin Suppresses Bone Loss in Ovariectomized ddY Mice and Collagen-Induced Arthritis in DBA/1J Mice: Possible Involvement of Receptor Activator of NF-kappaB Ligand (RANKL)-Induced Osteoclastogenesis Regulation. Bio. Factors. 2007, 30(3), 179–192. DOI: 10.1002/biof.5520300305.
- Padalhin, A. R. Brief Retrospect on the Use of Photobiomodulation (PBM) Therapy for Augmented Bone Regeneration (ABR). Med. Las; Engine, Basic Res, and Clin. Appl. 2021, 10(1), 15–21. DOI: 10.25289/ML.2021.10.1.15.
- Kawaii, S.; Ikuina, T.; Hikima, T.; Tokiwano, T.; Yoshizawa, Y. Relationship Between Structure and Antiproliferative Activity of Polymethoxyflavones Towards HL60 Cells. Anticancer. Res. 2012, 32(12), 5239–5244.
- Ianoşi, S.; Ianoşi, G.; Neagoe, D.; Ionescu, O.; Zlatian, O.; Docea, A. O.; Badiu, C.; Sifaki, M.; Tsoukalas, D.; Tsatsakis, A. M., et al. Age-Dependent Endocrine Disorders Involved in the Pathogenesis of Refractory Acne in Women. Mol. Med. Rep. 2016, 14(6), 5501–5506. DOI: 10.3892/mmr.2016.5924.
- Phung, H. M.; Lee, S.; Hong, S.; Lee, S.; Jung, K.; Kang, K. S. Protective Effect of Polymethoxyflavones Isolated from Kaempferia Parviflora Against TNF-α-Induced Human Dermal Fibroblast Damage. Antioxidants. 2021, 10(10), 1609. DOI: 10.3390/antiox10101609.
- Vogiatzoglou, A.; Mulligan, A. A.; Lentjes, M. A.; Luben, R. N.; Spencer, J. P.; Schroeter, H.; Khaw, K. T.; Kuhnle, G. G.; Wang, D. Flavonoid Intake in European Adults (18 to 64 Years). PLoS One. 2015, 10(5), e0128132. DOI: 10.1371/journal.pone.0128132.
- Shitara, Y.; Maeda, K.; Ikejiri, K.; Yoshida, K.; Horie, T.; Sugiyama, Y. Clinical Significance of Organic Anion Transporting Polypeptides (OATPs) in Drug Disposition: Their Roles in Hepatic Clearance and Intestinal Absorption. Biopharm. Drug Dispos. 2013, 34(1), 45–78. DOI: 10.1002/bdd.1823.
- Tamai, I.; Nakanishi, T. OATP Transporter-Mediated Drug Absorption and Interaction. Curr. Opin. Pharmacol. 2013, 13(6), 859–863. DOI: 10.1016/j.coph.2013.09.001.
- Hung, W. L.; Chang, W. S.; Lu, W. C.; Wei, G. J.; Wang, Y.; Ho, C. T.; Hwang, L. S. Pharmacokinetics, Bioavailability, Tissue Distribution and Excretion of Tangeretin in Rat. J. Food Drug Anal. 2018, 26(2), 849–857. DOI: 10.1016/j.jfda.2017.08.003.
- Kumar, A.; Devaraj, V. C.; Giri, K. C.; Giri, S.; Rajagopal, S.; Mullangi, R. Development and Validation of a Highly Sensitive Lc‐ms/Ms‐esi Method for the Determination of Nobiletin in Rat Plasma: Application to a Pharmacokinetic Study. Biomed. Chromatogr. 2012, 26(12), 1464–1471. DOI: 10.1002/bmc.2717.
- Bailey, D. G. Fruit Juice Inhibition of Uptake Transport: A New Type of Food–Drug Interaction. Br. J. Clin. Pharmacol. 2010, 70(5), 645–655. DOI: 10.1111/j.1365-2125.2010.03722.x.
- Tapaninen, T.; Neuvonen, P. J.; Niemi, M. Orange and Apple Juice Greatly Reduce the Plasma Concentrations of the OATP2B1 Substrate Aliskiren. Br. J. Clin. Pharmacol. 2011, 71(5), 718–726. DOI: 10.1111/j.1365-2125.2010.03898.x.
- Lilja, J. J.; Juntti‐patinen, L.; Neuvonen, P. J. Orange Juice Substantially Reduces the Bioavailability of the Β‐adrenergic–Blocking Agent Celiprolol. Clin. Pharmacol & Therapeutics. 2004, 75(3), 184–190. DOI: 10.1016/j.clpt.2003.11.002.