
ABSTRACT
We present the longest tree-ring chronology to date in northeastern North America (2233 years; 227 BCE – 2005 CE), resulting from several research projects conducted at the subarctic treeline in northern Quebec. This raw chronology of tree-ring width includes 464 black spruce (Picea mariana (Mill.) B.S.P.) shrubs (krummholz) growing in wetlands and preserved within peatlands. An indexed series of 152 erect black spruce trees that have lived in wetlands is also presented, covering the period 216 BCE to 1619 CE. We compared these chronologies to a tree-ring series including 116 black spruce trees and krummholz having grown on well-drained lichen woodlands over the period 1304–2000 CE. These chronologies highlight the major climatic periods of the last two millennia. Floating chronologies dating from 2500 to 3500 years ago were also developed from trees preserved in frozen peat. Growth rings from this period are much wider than those of the last 2233 years, suggesting warm climatic conditions and permafrost-free peatlands during the transition from mid- to late Holocene. The three subarctic chronologies presented here underscore the relevance and usefulness of tree growth rings and growth forms as ecological tools to assess the influence of climate on subarctic ecosystems.
Résumé
Nous présentons une synthèse des recherches menées à la limite des arbres du Québec subarctique qui ont permis la confection d’une série dendrochronologique couvrant 2233 ans, entre l’an 227 AEC (AEC ‘avant l’ère courante’, soit avant l’an 0 de notre ère) et l’an 2005 EC (EC ‘ère courante’, soit après l’an 0 de notre ère). Il s’agit de la plus longue série produite à ce jour dans le nord-est de l’Amérique du Nord. Cette série de largeur de cernes a été confectionnée à partir de 464 épinettes noires (Picea mariana (Mill.) B.S.P.) arbustives (krummholz) établies dans des milieux humides et préservées dans des tourbières. Une série indicée de 152 épinettes noires arborescentes ayant vécu dans des milieux humides est également présentée. Elle couvre la période 216 AEC à 1619 EC. Ces séries sont comparées à une série confectionnée à partir de 116 épinettes noires (arbres et arbustes) de sols bien drainés de pessières à lichens couvrant la période 1304-2000 EC. Les séries dendrochronologiques mettent en relief les grandes périodes climatiques des deux derniers millénaires. Des chronologies flottantes datées entre 2500 et 3500 ans avant aujourd’hui ont été construites à partir d’arbres ayant poussé dans des tourbières maintenant sous l’influence du pergélisol. Les cernes de croissance des arbres de cette période sont beaucoup plus larges que ceux des 2233 dernières années, suggérant l’influence d’un climat plus chaud qu’aujourd’hui, au passage de l’Holocène moyen à l’Holocène supérieur. Les chronologies subarctiques présentées dans ce travail soulignent la pertinence et l’utilité des formes de croissance et des cernes de croissance des arbres comme outils écologiques pour évaluer l’influence du climat sur les écosystèmes subarctiques.
Introduction
Trees are natural ecological archives with annual resolution informing on ecological processes, abiotic and biotic disturbances, ecosystem development and climate change. Measurements of tree-ring width can be used to construct chronologies of variable length, from a few decades to several hundred or even thousand years. Millennial growth ring series are useful to reconstruct long-term climatic variations (Helama et al. Citation2005; Büntgen et al. Citation2011) and past ecosystem responses to climate change (Arseneault and Payette Citation1997a; Payette and Delwaide Citation2004). Long chronologies also allow dating of trees having died following natural disturbances such as wildfires, landslides, rockfalls (Brown et al. Citation1999; Corona et al. Citation2013), or wood structures in buildings and archeological sites (Delwaide and Filion Citation1999; Querrec et al. Citation2009; Martin-Benito et al. Citation2014; Rochner et al. Citation2017). A Scots pine (Pinus sylvestris L.) chronology of more than 7650 years, the first and longest conifer tree-ring chronology in northern Eurasia, was constructed with tree megafossils found in lakes of northeastern Finnish Lapland (Helama et al. Citation2008). The longest chronology in North America covers 8837 years and was constructed from trees with a longevity greater than 1000 years (Pinus aristata Engelm., Pinus longaeva Bailey) growing in the dry, mountainous areas of western United States (Ferguson Citation1969; LaMarche and Harlan Citation1973; Salzer et al. Citation2019). In northeastern North America, the longest chronologies to date cover the period 5 BCE-2016 CE (Arseneault et al. Citation2013; Büntgen et al. Citation2020) and 572 CE-2016 CE (Gennaretti et al. Citation2014) and were constructed with black spruce (Picea mariana (Mill.) B.S.P.) trees growing in the northern boreal forest. Two other long chronologies were developed from white cedar (Thuja occidentalis L.) in dry environments of the Niagara Escarpment in Ontario (798–2001 CE; Kelly et al. Citation1994) and on islands of Lac Duparquet in Quebec (1186–1987 CE; Archambault and Bergeron Citation1992).
In this study, we present the longest tree-ring chronology in northeastern North America, developed from black spruce sampled in the subarctic region of the Rivière Boniface–Lac Bush area (RBA) in northern Quebec (). The construction of this long chronology (WkC, ) began during the 1980s. Well-preserved dead stems and living trees in lichen woodlands of the Lac Bush area made it possible to develop a first black spruce chronology spanning more than 600 years (, ; Payette et al. Citation1985). Additional data from RBA (; Payette et al. Citation1989) were subsequently obtained from a site that escaped wildfire for more than 1000 years (; Vallée and Payette Citation2004). Additional chronologies were constructed from black spruce growing in or at the edge of peatlands (hereafter called wetlands, ). Lavoie and Payette (Citation1997) constructed seven floating chronologies using black spruce trunks preserved in permafrost peatlands (). Arseneault and Payette (Citation1997a) provided a chronology covering the period 690–1591 CE, as well as three older floating chronologies (). A reconstruction of lake level fluctuations made it possible to develop nine chronologies from samples preserved in two ponds and at the bottom of two lakes (; Payette and Delwaide Citation2004). Another chronology was prepared in 2010 using samples preserved in a palsa bog (; Cyr and Payette Citation2010). Finally, Asselin and Payette (Citation2006) constructed 10 chronologies from wetland sites () and successfully aligned them to the floating chronologies of Arseneault and Payette (Citation1997a).
Figure 1. Location of the study area in the Rivière Boniface and Lac Bush regions, subarctic Québec
Figure 2. Time periods covered by the growth ring chronologies developed in the Rivière Boniface region according to the type of environment (LWC: Lichen Woodland Chronology; WkC: Wetland krummholz Chronology; WtC: Wetland tree Chronology). (A) Payette et al. (Citation1985); (B) Payette et al. (Citation1989); (C) Vallée and Payette (Citation2004); (D) Lavoie and Payette (Citation1997); (E) Arseneault and Payette (Citation1997a); (F) Pond 1 (P1), Pond 2 (P2), Lac Bon (LB), Lac à l’eau (LE): Payette and Delwaide (Citation2004); (G) Cyr and Payette (Citation2010); (H) Asselin and Payette (Citation2006). Solid lines correspond to dated growth ring chronologies and dashed lines to floating chronologies. Juxtaposing the two lines (solid and dashed) shows that these chronologies were first published as floating, then dated for the present study
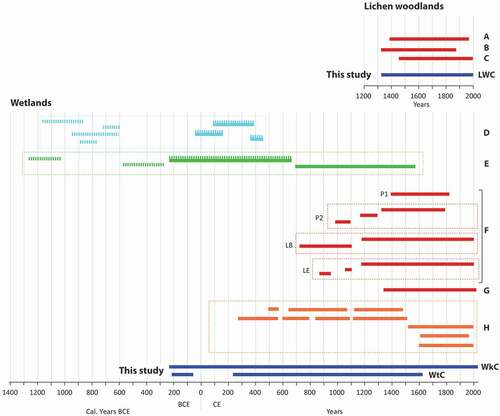
Figure 3. Photographs from the Rivière Boniface area: (a, b) black spruce-lichen woodland with living erect trees developed during the 20th century and dead sub-arborescent trees-krummholz (in the foreground) that lived during the Little Ice Age; (c) wetland spruce environment; (d) stunted spruce that lived during the Little Ice Age; (e) erect spruce trees that lived during the 16th century in contrast with living krummholz in a permafrost peatland; (f) stunted spruce clones with thufurs of Rhododendron subarcticum. Photo credit: S. Payette

The main objective of this research was thus to construct long black spruce chronologies highlighting the dominant climatic periods that had a major influence on ecosystem development in the Subarctic of eastern North America. Specifically, our study combined all datasets produced to date for RBA in order to generate three long growth-ring chronologies including two encompassing more than two millennia. Two chronologies were developed in wetland sites, one from krummholz (stunted) individuals (hereafter WkC) covering 2233 years, and another one from erect trees (hereafter WtC) covering 1662 years. The third chronology is from lichen woodland sites (hereafter LWC) and covers 697 years. Ring width patterns and growth forms of black spruce were used concurrently to highlight the influence of climate over the time period covered by the chronologies.
Methods
Study area
The study area is located in the subarctic zone of northern Quebec, along Rivière Boniface (57° 44ʹ N, 76° 10ʹ W), which is about 145 km south-east of the Inuit village of Inukjuak and 30 km from the Hudson Bay coast, and in the Lac Bush area (57° 47ʹ N, 75° 45ʹW), about 15 km east of Rivière Boniface (). This region corresponds to the arctic treeline located at the northern limit of the forest tundra zone (Payette Citation1983). The dominant tree species, black spruce, has been present in the region for at least 6450 years (Bhiry et al. Citation2007). Rare tamarack individuals (Larix laricina (Du Roi) K. Koch) are also present in the region. At their northern limit in northern Quebec, black spruce stands represent refugial forest enclaves in a shrub tundra-dominated landscape. Black spruce-moss stands are the most important forest type in protected valleys and shallow depressions, whereas lichen woodlands are distributed on slopes and exposed hilltops along with lichen-heath-dwarf birch (Betula glandulosa Michx.) communities. The fragmented RBA landscape resulted from fire-induced deforestation under deteriorating climatic conditions over the past millennia (Payette and Gagnon Citation1985; Payette and Morneau Citation1993; Asselin and Payette Citation2005a). The remaining black spruce stands have escaped fire and have persisted since initial post-glacial establishment (Payette and Morneau Citation1993; Vézeau and Payette Citation2016), shortly after the retreat of the Tyrrell Sea (Dyke Citation2004). The long persistence of these forests is attributed to the phenotypic plasticity of black spruce and its capacity to multiply vegetatively by layering, particularly during periods of climatic deterioration (Arseneault and Payette Citation1997b; Asselin and Payette Citation2005b).
Sample preparation and ring width measurement
All tree-ring studies in the RBA used the same methods for collecting and preparing samples. All relevant information pertaining to the growth form of black spruce individuals (e.g., erect trees vs stunted) were noted. Wood discs were collected as close as possible to the stem base or collar. Samples were dried and finely sanded until the wood cells were visible under a binocular microscope. Annual growth rings were measured with a Velmex micrometer (0.002 mm) or a Henson micrometer (0.01 mm) at a magnification of 40x. Only one radius per stem sample was measured because of frequent eccentric growth.
Stem dating
Samples were visually dated under a binocular microscope by identifying light ring patterns. Light rings are made of latewood tracheids delineated by very thin cell walls (Filion et al. Citation1986; Wang et al. Citation2000). They are often formed in years with unusually low summer temperatures (Liang et al. Citation1997; Wang et al. Citation2002), sometimes due to the cooling effect of major volcanic eruptions. At least 43 of the 65 light rings observed between 1500 CE and 1980 CE were indeed formed during the year of a major volcanic eruption or the following year (Filion et al. Citation1986). Light ring occurrence in black spruce increases with latitude in northern Quebec, with the highest frequencies recorded at the treeline (Delwaide et al. Citation1991). Accordingly, light rings have been shown to allow for robust crossdating of black spruce trees over the last 1500 years in the RBA (Payette et al. Citation1985 Payette et al. Citation1989; Payette and Delwaide Citation2004). Some specific and easily recognisable light ring patterns are highly reliable for crossdating (Arseneault and Payette Citation1998). Crossdating accuracy of all measured stems was also assessed using the program COFECHA (Holmes Citation1983).
Light rings are found in trees and stunted individuals, and their formation process is similar in dry sites (woodlands) and in wetlands (Arseneault and Payette Citation1998). While wood pieces buried in peat or in lake sediments have a darker color than living trees (Wilson et al. Citation2019) due to oxidation of dissolved ferrous iron (Wang et al. Citation2020), light rings can nevertheless be easily detected in buried trees and shrubs.
The initial development of the long light-ring chronology of RBA was based on living trees (1554–1978 CE: Filion et al. Citation1986, p. 1231–1978 CE: Delwaide et al. Citation1991). Arseneault and Payette (Citation1997a;Citation1998) extended the chronology to 690 CE using buried wood recovered from wetland sites. Several floating chronologies (Arseneault Citation1996; Lavoie and Payette Citation1997) were also constructed and radiocarbon-dated, including one series covering the period 178 BCE-785 CE. Asselin (Citation2005) fused the latter chronology to the calendar-dated one to produce a light ring chronology covering the 211 BCE-1992 CE period (Supplemental material S1). The floating chronologies (Lavoie and Payette Citation1997; Arseneault and Payette Citation1997a) were 14C-dated and calibrated using version 8.2 of the CALIB software (Stuiver et al. Citation2020).
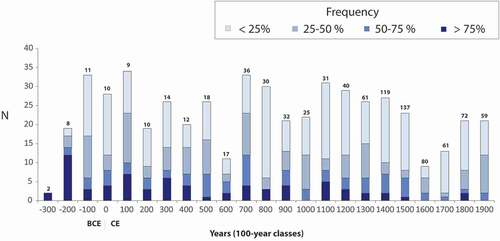
From 227 BCE to 2005 CE, 524 distinct light ring years were observed. Of these, 140, 129 and 255 light rings were found in more than 50%, 25–50% and less than 25% of the samples, respectively. These rings are distributed throughout the chronologies, with a mean of 23 light rings per century (). The lowest number of light rings was recorded during the 7th, 17th and 18th centuries. Light rings allowed dating of almost all treeline black spruce stems containing more than 100 growth rings between 227 BCE and today.
Figure 4. Frequency of light ring years per century in the Rivière Boniface area. The number of samples is indicated at the top of each bar. Different shades of blue indicate the percentage of samples for a given century on which a given number of light ring years were observed
Black spruce growth forms
Black spruce growing at their northern range limit are extremely sensitive to climatic variations and develop different growth forms depending on summer growth conditions and winter harshness (Bégin Citation1991; Lavoie and Payette Citation1992; Payette et al. Citation1994; Pereg and Payette Citation1998). The morphological development of black spruce follows Rauh’s architectural model (Bégin and Filion Citation1999). Vegetative multiplication by layering largely explains black spruce survival during cold years when sexual reproduction is impaired. Massive foliar loss, particularly at the snow–air interface, occurs because of mechanical damage and frost desiccation of leaves subjected to cold winter winds (Hadley and Smith Citation1983, Citation1986; Marchand Citation1987; Payette et al. Citation1996; Maher et al. Citation2020). Bud and leaf erosion above the snowpack during cold and dry winters often result in the transfer of apical dominance to infranival, undamaged dormant buds (Bégin and Filion Citation1999), causing the regression from erect (tree) to shrubby (krummholz) growth forms (). Eroded black spruce trees can survive for centuries as stunted individuals with a reduced leaf mass (Pereg and Payette Citation1998). Krummholz can also develop sub-arborescent growth forms (multi-stemmed or single-stemmed) when winter conditions become milder. Sub-arborescent krummholz are characterized by short (<2 m), damaged stems. In contrast, erect trees have taller (>2 m), undamaged supranival stems. Erect trees develop when sites are less exposed to snow-drift conditions, even at the northernmost range limit of black spruce (). Tree and shrub growth forms were differentiated based on the presence/absence of a supranival stem.
Climatic trends derived from changing black spruce growth forms have often been described in RBA (Payette et al. Citation1989, Citation1994; Lavoie and Payette Citation1992; Payette and Delwaide Citation1994), and they are summarized in .
Table 1. Characteristics of the WkC, WtC and LWC chronologies
Series standardization
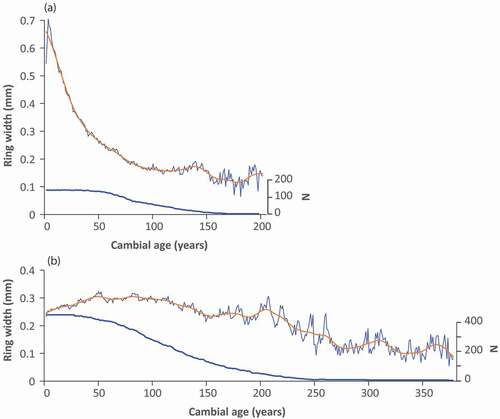
Black spruce trees produce gradually narrower growth rings as they age and become taller. This biological trend has to be removed to detect growth variations due to climate changes. To do so, we used the Regional Curve Standardization (RCS) method (Briffa et al. Citation1992; Esper et al. Citation2003). All dated erect trees from wetlands were used to develop a chronology (WtC chronology) of mean tree-ring width according to cambial age (). We applied a 32-year cubic spline to obtain the biological trend of decreasing ring width as a function of cambial age. We then divided each single series by this growth model to obtain a standardized chronology. In contrast, ring widths of stunted individuals do not show any significant change in radial growth with cambial age under 225 years of age ().
Figure 5. Mean ring width of black spruce individuals from the Rivière Boniface area according to cambial age, along with the corresponding 32-year cubic spline and number of measured stems. (a) Erect trees (WtC) and (b) krummholz individuals (WkC)
Response function analysis
We quantified the relationship between tree growth and climate using response function analysis, a variant of multiple regression analysis which uses monthly climate predictor variables, orthogonalized as principal components, to identify the main climate drivers of growth (Zang and Biondi Citation2013). For this analysis, we used mean monthly temperature and total monthly precipitation (1940–2000 period) from the nearest meteorological station, located in Inukjuak, 145 km northwest of RBA. A weather station has been active at RBA since 1988 (CEN Citation2020). Despite the distance between the Inukjuak and RBA stations, a highly significant correlation (r = 0.996) was found between the annual temperature measured at the two stations over the 1988–2000 period. Although no precipitation data are available for RBA, a correlation between precipitation data at the Inukjuak station and RBA ring widths was calculated. Caution must be exercised given the fact that the Inukjuak station is under maritime conditions while RBA is under drier, continental conditions. We performed the response function analysis with the ‘bootRes’ package (Zang and Biondi Citation2013) in R (R Core Team Citation2019). We used 1000 bootstrap replicates to calculate significance at the 95% level.
Results
Wetlands
Establishment and lifespan of wetland trees and shrubs (krummholz)
We successfully crossdated 616 black spruce stems in wetlands using the light ring chronology and validating with the COFECHA program (mean correlation of all samples with the master chronology: 0.342). In total, 152 stems (25%) were classified as trees and 464 stems (75%) were shrubs (). All wetland individuals were preserved in peat and lake sediments over the last two millennia. Because some decayed stems were sampled above the root collar, the date of establishment could have been earlier than observed. Also, most measured stems had no bark, which indicates that the last growth ring was not necessarily present.
Figure 6. (a) Lifespan of sampled black spruce from the wetland environment of the Rivière Boniface area (WkC and WtC); (b) proportion of erect stems (in red) and krummholz (in black) based on the corresponding century of the first year of the lifespan. The number of samples per century is indicated above each bar
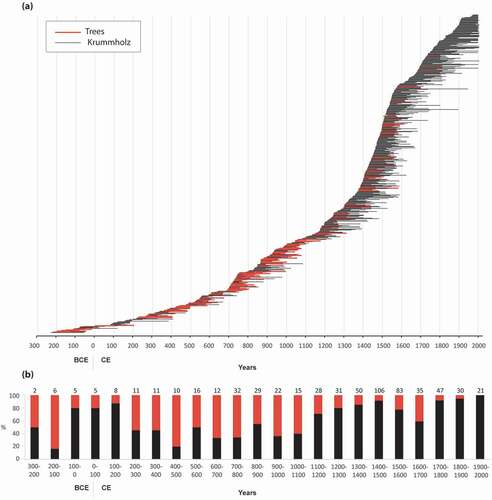
Black spruce individuals that established between 100 BCE and 100 CE were mostly krummholz (over 80% of the sampled stems; ). Krummholz also dominated between the 12th and 20th centuries. In contrast, most of the stems having lived prior to 100 CE or between 200 and 1000 CE displayed an erect growth form. It is noteworthy that a relatively high proportion (20–40%) of erect trees developed during the 16th and 17th centuries ().
Longevity of wetland trees and shrubs (krummholz)
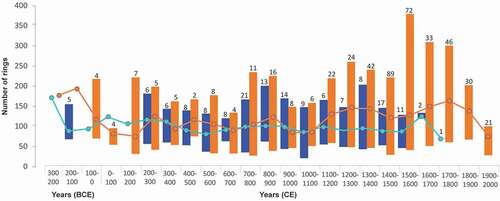
The mean number of growth rings in trees (WtC) and shrubs (WkC) was 93 (± 35) and 123 (± 55), respectively, suggesting greater longevity of stunted stems compared to erect stems. None of the erect stems contained more than 201 growth rings (). The mean number of growth rings was consistently greater and statistically different in krummholz than in trees (t-test: 7.125; p < 0.005). The number of rings varied little in trees and krummholz before 1100 CE, except between 227 and 100 BCE, a poorly replicated time interval with a small number of sampled stems. A marked increase in the number of growth rings in krummholz stems occurred between 1100 and 1900 CE.
Figure 7. Mean, minimum and maximum number of growth rings per century, for all spruce individuals sampled in wetlands of the Rivière Boniface area, either trees (WtC; blue) or krummholz (WkC; orange). Values are recorded according to the corresponding century of establishment. The number of samples established during each century is indicated above each bar. Dots indicate that only one stem established during the corresponding century
Wetland krummholz chronology (WkC)
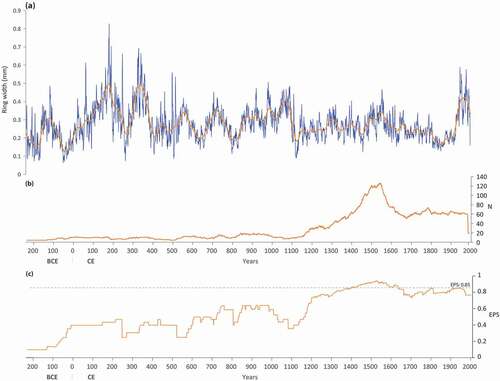
The mean ring width chronology of wetland krummholz covers 2233 years and clearly shows large variations in radial growth over time (). The period from 227 BCE to 100 CE is marked by slow growth, with the exception of a short episode around 120 BCE. Large growth rings occurred in 100–400 CE, 850–1110 CE and 1920–2005 CE, whereas the longest period of slow growth took place between 1110 and 1920 CE, except during the 16th century. The mean correlation (r) among all individuals within this chronology is 0.24 (p < 0.001). Highest EPS values are observed for the period 1200–2000 CE, particularly between 1400 and 1600 CE with a sample depth over 100 samples ().
Figure 8. (a) Mean ring width of krummholz from wetland sites (WkC) of the Rivière Boniface area, along with a superimposed 32-year cubic spline; (b) number of stems used in the chronology; (c) EPS (Expressed Population Signal) calculated according to period
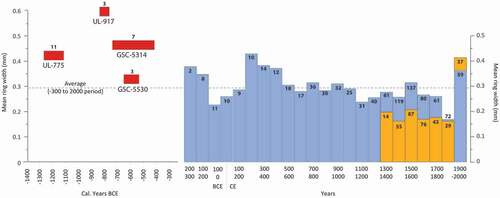
Four floating chronologies of Lavoie and Payette (Citation1997) cover the period between 1247 BCE and 539 BCE (calibrated years) with a mean ring width varying between 0.32 and 0.60 mm (). Most individuals from these chronologies had supranival stems (Lavoie Citation1994). Comparison of the ring width between these floating chronologies and the dated woodland and wetland chronologies shows greater mean growth of black spruce before 539 BCE than during the last 2200 years ().
Table 2. Statistics of the floating chronologies
Figure 9. Mean ring width of spruce trees and krummholz that have lived during each century in wetlands (WkC and WtC: blue) and lichen woodlands (LWC: yellow) of the Rivière Boniface area. The corresponding number of samples is indicated at the top of each bar. Because floating chronologies (red) were 14C dated, mean width and number of samples refer to the entire length of each chronology
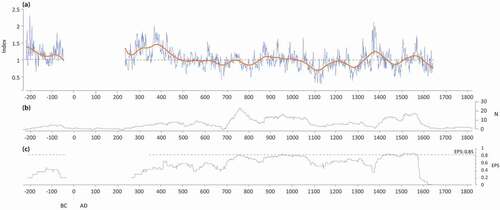
Wetland tree chronology (WtC)
The RCS chronology of wetland trees covers the period 216 BCE-1619 CE. The 50 BCE-225 CE interval is not presented because less than five trees were sampled (). Larger growth rings (above the mean) were formed in 225–400 CE, 1345–1425 CE and 1510–1570 CE.
Figure 10. (a) Mean standardized chronology of erect stems from wetland sites of the Rivière Boniface area (WtC), covering the 216 BCE-1619 CE period (except between 50 BCE and 225 CE for which less than five spruce individuals were available); (b) number of individuals included in the chronology; (c) EPS (Expressed Population Signal) calculated according to period
Lichen woodlands
Lichen woodland chronology (LWC)
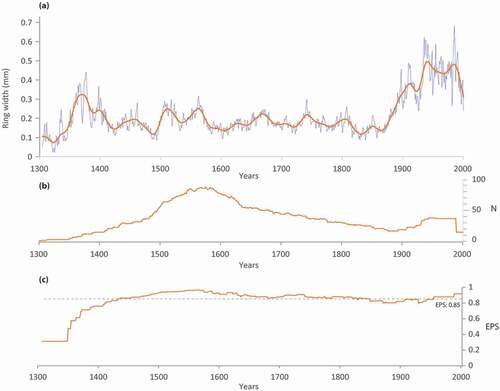
The lichen woodland chronology covers the years 1304 to 2000 CE (), and shows three periods of high radial growth at 1350–1403 CE, 1495–1575 CE, and 1880 CE to present. The length of the chronology was limited by the maximum conservation time of dry wood on the soil surface, which is about 400–600 years in RBA (Payette et al. Citation1989). We pooled together 116 stems from three datasets constructed in lichen woodlands of RBA (Payette et al. Citation1985, p. 1398–1982 CE, Citation1989, p. 1304–1988 CE; Vallée and Payette Citation2004, p. 1476–2000 CE). The mean ring-width chronologies from the three datasets are highly correlated (r = 0.52; p < 0.001) while all individual black spruce series are also significantly correlated (r = 0.293; p < 0.001). The mean number of growth rings per individual was 163, with a maximum of 504 (Payette et al. Citation1985).
Figure 11. (a) Lichen woodland chronology (LWC) from the Rivière Boniface area, along with a superimposed 32-year cubic spline; (b) annual number of stems used in the mean chronology; (c) EPS (Expressed Population Signal) calculated according to period
Comparison between the lichen woodland and wetland chronologies
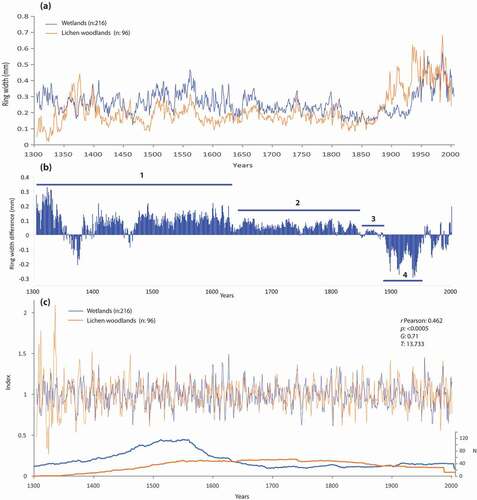
Wetland black spruce showed larger raw ring width than lichen woodland black spruce between 1304 and 1850 CE, except during a short interval in the 14th century (). Differences between the two environments were larger before 1630 CE (mean difference of 0.09 mm; b1) compared to the 1630–1880 CE period (mean difference: 0.06 mm; b2). Both chronologies showed similar, narrow growth rings between 1840 and 1880 CE (mean difference of 0.01 mm; b3). A sharp increase in radial growth occurred around 1880 CE in lichen woodlands, and 60 years later in wetlands (b4). The two chronologies display similar growth after 1940. We applied a 32-year cubic spline to the mean ring width chronologies to compare year-to-year variations. Despite long-term growth differences between the two environments, standardized values are significantly correlated (r = 0.462, p < 0.001; ; ).
Table 3. Statistics of the LWC and WkC chronologies. Chronologies were indexed using a 32-year cubic spline
Figure 12. (a) Mean ring width chronologies in lichen woodland (LWC) and wetland environments (WkC) of the Rivière Boniface area over the 1304–2000 CE period; (b) annual ring width difference between the two chronologies according to four periods (b1: 1304–1630; b2: 1630–1840; b3: 1840–1880; b4: 1880–1940); (c) mean standardized (32-year cubic spline) chronologies and annual number of samples
Response functions
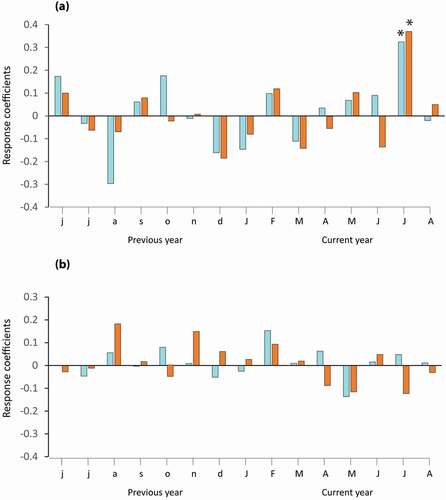
Monthly mean, maximum and minimum temperatures and monthly total precipitation of the Inukjuak station were compared to the mean LWC and WkC chronologies standardized with a 32-year cubic spline (period 1940–2000 CE from standardized values as presented in ; ). In both environments, response functions show a significant correlation with mean July temperature of the current year, which has a positive influence on radial growth (response correlations of 0.41 and 0.35 for lichen woodlands and wetlands, respectively), whereas mean maximum and minimum July temperatures showed high but nonsignificant correlation coefficients with the chronology (0.32 and 0.29, respectively, for lichen woodland and 0.28 and 0.24, respectively, for wetlands). No significant influence of precipitation on radial growth was found.
Figure 13. (a) Response coefficients between mean monthly temperature and LWC indexed (32-year cubic spline) chronology (in orange) and the indexed (32-year cubic spline) WkC chronology (in blue); (b) response coefficients between total monthly precipitation data and both chronologies. Stars represent significant correlations at the p < 0.05 confidence level (response coefficient: 0.41 and 0.35, respectively)
Discussion
The progressive reduction of solar radiation associated with orbital forcing over the northern hemisphere (Berger Citation1978), including northeastern North America, is among the main drivers of continental-wide cooling from the mid-Holocene to the present (Bradley Citation2014). The RBA floating and calendar-dated ring width chronologies spanning the last 3500 years highlight the direct response of black spruce to climate changes that occurred during this long period, at the turning point between the mid- and late Holocene (Walker et al. Citation2012). Due to their phenotypic plasticity, treeline black spruce in subarctic Quebec successfully adapted to extreme conditions and were able to persist for millennia despite the long-term cooling trend.
Together, the ring-width chronology of krummholz individuals (WkC) and the growth-form chronology seem to contain more robust climatic information than the RCS chronology of black spruce trees at RBA. Although the RCS method is widely used to remove the age-related biological trend, it has limitations that may have distorted the climatic signal in this chronology. For example, this method requires homogeneous edaphic conditions, very high replication, as well as stable mean cambial age and sampling height over time (Esper et al. Citation2008; Linderholm et al. Citation2010; Autin et al. Citation2015). In our study, living trees were sampled at the root collar in order to get the first growth ring, while dead and decayed stems were frequently sampled at an unknown position above the root collar. In addition, sample depth in the chronologies varied over time and was low for some periods. For all these considerations, and since the Expressed Population Signal (EPS) is below the well-established threshold value of 0.85 (Wigley et al. Citation1984) for almost all of the WtC chronology (), conditions for RCS standardization were not entirely met and this chronology may have diverged from climatic trends, especially during periods of low replication. Conversely, because ring width measurements from krummholz individuals do not contain an age-related growth trend, ring width variations over time are more directly linked to climate conditions.
Differential growth of black spruce in lichen woodlands and wetlands
Black spruce growth differed between the wetlands and the well-drained soils of the lichen woodlands, mostly during the 1304–1630 CE and 1880–1940 CE periods (b1 and 12b4). Before 1650 CE, spruce growth was greater in wetlands than in lichen woodlands. This period, particularly before the 17th century, was marked by low lake levels in RBA, most likely caused by reduced snow precipitation (Payette and Delwaide Citation2004). Because ring width in peatland black spruce is positively correlated to water table depth due to greater development of fine root biomass (Lieffers and Rothwell Citation1987), larger rings were probably associated with drier soil conditions during this period. In contrast, dryness may have been conducive to hydric stress and reduced growth in lichen woodlands during the same period. A close relationship between drought and reduced growth has been documented for black spruce in northern Québec and Alaska (Girard et al. Citation2011; Wolken et al. Citation2016; Sullivan et al. Citation2017) and white spruce (Picea glauca (Moench) Voss) in Alaska (Barber et al. Citation2000) during the 20th century. Smaller growth differences were found between lichen woodland and wetland black spruces after 1630 CE (b2). Higher precipitation may have contributed to reduce water stress in lichen woodland black spruce. The smallest growth difference between the two chronologies (b3) occurred between 1840 and 1880 CE. Additional factors may have contributed to growth differences between the two environments. Wetlands are less exposed to wind and accumulate more snow in winter compared to hillsides and hilltops. Krummholz growth rings in wetlands are generally larger than those in well-drained sites, most likely because of greater foliar mass below the snow cover and water availability.
The period 1880–1940 CE was marked by better growth in lichen woodlands than in wetlands (average ring-width difference of 0.16 mm). This large difference may be attributed to the increased snow cover recorded in the region since the end of the 19th century. Several studies carried out in northern Quebec have concluded that precipitation in the 20th century was greater than during the Little Ice Age, particularly in winter (Bégin and Payette Citation1988; Payette and Delwaide Citation2004). The basal level of tree stem erosion from windblown snow increased considerably between the end of the 19th century and the beginning of the 20th century (Bégin Citation1991; Lavoie and Payette Citation1994). Mechanical damage to black spruce around snow patches in the Lac Guillaume–Delisle area (Morin and Payette Citation1986) and increased frequency of ice scars on trees on the shore of Lac à l’Eau-Claire (Bégin and Payette Citation1988) are additional evidence of a trend towards snowier conditions from the late 19th century to early 20th century in subarctic Quebec. Increased thickness of the snow cover most likely allowed for the development of taller trees (from 0.8 to 1.6 m; Lavoie and Payette Citation1994) and reduced water stress in lichen woodlands.
Relationships to climate
The climatic signatures of the Roman Climatic Optimum, Dark Ages Cold Period, Medieval Warm Period, Little Ice Age and the current warming trend are well expressed in the two long chronologies (). Whereas ring-width chronologies describe tree growth at an annual resolution, growth forms provide insights on the morphological adaptation of black spruce at a longer time scale, because of the delay between climate changes and the ecological response of the species.
Figure 14. (a) Lichen woodland chronology (LWC), along with a superimposed 32-year cubic spline. Horizontal bar indicates number of stems included in the chronology (yellow: 1–5; orange: 6–10; light red: 11–20; dark red: >20); (b) well-documented climatic periods; (c) wetland krummholz chronology (WkC), superimposed with a 128-year cubic spline. Horizontal bar indicates number of stems (same classes as LWC); (d) proportion of erect stems (grey bars) and krummholz (black bars) by 100-y periods. Periods of increased growth are highlighted as yellow zones, and periods of reduced growth as blue zones
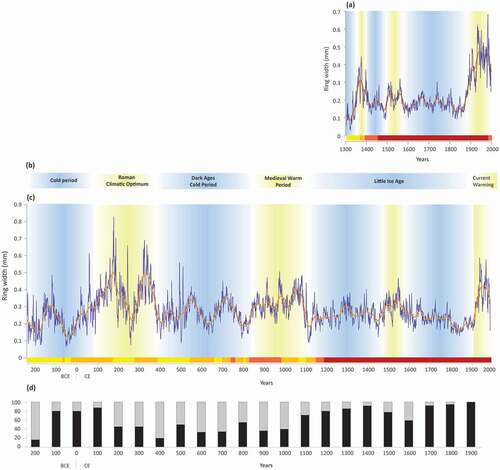
Period prior to 150 CE
Despite the low number of samples analyzed, the largest growth rings ever recorded in RBA were those of wetland black spruce having grown 2500 to 3500 years ago (). The majority of the sampled black spruce had erect growth forms (Lavoie Citation1994). Because these trees were mostly sampled in frozen peat (Lavoie and Payette Citation1997), they were buried before permafrost development, indicating subsequent climate cooling. At the global scale, this period was probably characterized by the most favorable climatic conditions for tree growth since the end of the mid-Holocene (Masson-Delmotte et al. Citation2018). In contrast, tree growth has been greatly reduced between 227 BCE and 100 CE, despite the smaller dataset covering this period. Cold conditions during this period were also deduced elsewhere based on ice core data in Greenland (Dahl-Jensen et al. Citation1998) and larch (Larix gmelinii Pilger) tree rings in Siberia (Naurzbaev and Vaganov Citation2000).
Roman Climatic Optimum
Despite the relatively low number of measured samples, black spruce krummholz which lived between 100 and 400 CE (n = 19) produced large growth rings suggesting favorable climatic conditions in RBA, except for a short episode between 250 and 280 CE (). High ring width values are also observed in the indexed series of erect trees (period 225 to 400 CE: ). The Roman Climatic Optimum coincided in Europe with the expansion of the Roman Empire (Wang et al. Citation2012), and was characterized by warm and dry summers in the British Isles and central Europe (Lamb Citation1995) along with relatively high sea surface temperatures north of Iceland (Eiriksson et al. Citation2006). In RBA, growth-ring width during this period showed the highest values across the entire chronology, including the Medieval Warm Period and present. A multi-proxy temperature reconstruction also suggested that the Roman Period (1–300 CE) was as warm as the twentieth century (Ljungqvist Citation2010). Reconstructed summer temperatures from tree-ring datasets of the northern hemisphere (Büntgen et al. Citation2020) are also in phase with this trend.
Dark Ages Cold Period
Relatively low black spruce growth occurred between 400 and 830 CE, except between 530 and 560 CE and between 690 and 740 CE (). This period is one of the six cold periods that occurred during the Holocene (Wanner et al. Citation2011). This agrees with a literature review of 114 paleoclimate studies indicating cold climatic conditions in the northern hemisphere between 400 and 765 CE (Helama et al. Citation2017).
Medieval Warm Period
Previous studies (Lamb Citation1965; Wilson et al. Citation2016; Büntgen et al. Citation2020) have suggested that the climate of the Medieval Warm Period (ca. 900 to 1300 CE) was warmer than the preceding centuries. At the treeline in subarctic Quebec, this period covered the years 850 to 1110 CE and was marked by large growth rings, particularly between 980 and 1100 CE. Furthermore, most black spruce individuals (65%) that established between 900 and 1000 CE developed an erect growth form (), indicating that warm summers were complemented by mild winters. Peak summer temperatures during the 990s and 1020s were also observed elsewhere across the northern hemisphere (Büntgen et al. Citation2020).
Little Ice Age
The Little Ice Age spanned the time interval between 1450 CE and 1850 CE (Lamb Citation1977; Grove Citation2004; Harland et al. Citation2013; Anchukaitis et al. Citation2017). During this period, krummholz formed very narrow growth rings in RBA, particularly between 1816 CE and 1900 CE (). July–August temperatures reconstructed from black spruce in the boreal forest of northeastern North America identified the 1816–1857 CE period as the coldest 40-year period of the last 1100 years (Gennaretti et al. Citation2014), likely in response to the Tambora volcanic eruption in 1815 (Filion et al. Citation1986; Gennaretti et al. Citation2014). An abrupt decrease in ring width also occurred in black spruce (Filion et al. Citation1985; Payette et al. Citation1985; Jacoby et al. Citation1988) and white spruce trees along the Hudson Bay coast (Caccianiga et al. Citation2008) and Labrador highlands (Payette Citation2007).
Predominance of krummholz with narrow growth rings suggests that unfavorable growth conditions in RBA already prevailed during the 12th century (). Porter (Citation1981) pointed out that the Little Ice Age may have begun as early as 1300 CE in the North Atlantic region. Field evidence for an early beginning of the Little Ice Age in North America was also provided by glacier advances in western Canada (Luckman Citation1995). Tree-ring dates of in-situ stumps indicated that the Robson Glacier began over-riding lowland forests between ca. 1142 and 1150 CE and continued until at least 1350 CE. Our data suggest that climatic conditions at the subarctic treeline deteriorated abruptly at the beginning of the 12th century, in agreement with glacial advances in western Canada (Luckman Citation1995). This period also coincides with an increase of krummholz lifespan (). While the number of growth rings is similar for trees and krummholz before 1100 CE, the mean longevity of black spruce krummholz increased to reach 378 years between 1100 and 1900 CE. In addition, during this period, more than 84% of the sampled black spruce individuals grew as krummholz (), with the exception of the 16th century (Payette et al. Citation1989; Arseneault and Payette Citation1992), a situation also observed in the Lac Bush area (Filion et al. Citation1985). The latter period was characterized by larger rings () and a greater proportion of the tree growth form in both lichen woodland and wetland environments ().
Current warming trend
Since the end of the 19th century, global warming has triggered a noticeable increase of black spruce radial growth (). While only krummholz established during the 20th century (), they experienced prominent growth increase, a generalized feature associated with recent climate warming across the northern hemisphere (Wilson et al. Citation2016). Indeed, the krummholz that established in the 18thth, 19th and 20th centuries shifted from stunted to sub-arborescent and even erect growth forms (), causing a slight treeline shift toward the Hudson Bay coast (Lescop-Sinclair and Payette Citation1995) as well as northward (Lavoie and Payette Citation1994). Simultaneously, tree height within the lichen woodlands increased by at least 1.6 m (see ; Vallée and Payette Citation2004).
Conclusion
Our tree-ring chronologies, complemented by the analysis of black spruce growth forms, show the responsiveness of subarctic ecosystems to climate change over the last 3500 years. The WkC 2233-year chronology is climate-sensitive, as shown by yearly radial growth variations correlated with July temperatures. Black spruce individuals of RBA adopt different growth forms in response to climatic conditions. Because growth rate is a function of the photosynthetic capacity of trees, the raw growth-ring values of RBA chronologies are a direct ecological proxy of long-term climatic conditions. The RBA growth-ring data fill an important gap, both spatially and temporally, for a better knowledge of the mid- to late Holocene climate in boreal-subarctic areas. Growth forms and ring widths of black spruce at its northern range limit were linked to climatic conditions over the last two millennia. We found evidence in our data of the previously documented Roman Climatic Optimum, Dark Ages Cold Period, Medieval Warm Period, Little Ice Age as well as the current warming trend. Floating chronologies dating from 2500 to 3500 years ago also suggest that climatic conditions during this period were favorable to black spruce growth in northern Québec, and likely even milder than anytime during the last two millennia. Overall, the RBA chronologies clearly show a trend of decreasing growth from the Roman Climatic Optimum to the end of the Little Ice Age suggesting progressive cooling, with growth resumption during the 20th century probably associated with continental-scale warming. Although the number of trees sampled is relatively small for the oldest parts of the chronologies, the warmest period of the last two millennia occurred at the beginning of the Common Era, as based on the wetland krummholz chronology. Sensitive subarctic trees in northern Québec, as described in this study, are indeed one of the best proxies for the reconstruction of climatic conditions over the late Holocene.
Supplemental Material
Download Zip (10.2 KB)Acknowledgments
We thank François Cadrin, Éric Marcoux, Éric Poirier, Roger Saint-Arnault, Mylène Lévesque, Marie-Claude Dansereau, Annie St-Louis, Marie-Josée Laberge, Michel Bergeron, Marie-Josée Fortin, Kateri Lescop-Sinclair, Pierre Levasseur, Min Peng, Daria Péreg, Régis Pilote, Nadia Pitre, Martin Poitras, Michel Beauchemin, Marie-Claude Martel, Sébastien Champagne, Tamsin Laing, Reinhard Pienitz and Sheila Vallée for field and laboratory assistance. Thoughtful comments from two anonymous referees greatly improved the quality of the original manuscript.
Disclosure statement
No potential conflict of interest was reported by the authors.
Supplementary material
Supplemental data for this article can be accessed here
Additional information
Funding
Related Research Data
References
- Anchukaitis KJ, Wilson R, Briffa KR, Büntgen U, Cook ER, D’Arrigo R, Davi N, Esper J, Frank D, Gunnarson BE, et al. 2017. Last millennium Northern Hemisphere summer temperatures from tree rings: part II, spatially resolved reconstructions. Quaternary Sci Rev. 163:1–22. doi:https://doi.org/10.1016/j.quascirev.2017.02.020.
- Archambault S, Bergeron Y. 1992. An 802-year tree-ring chronology from the Quebec boreal forest. Can J Forest Res. 22(5):674–682. doi:https://doi.org/10.1139/x92-090.
- Arseneault D. 1996. Analyse dendroécologique des macrorestes ligneux des tourbières subarctiques. [PhD dissertation]. Québec: Université Laval.
- Arseneault D, Dy B, Gennaretti F, Autin J, Bégin Y. 2013. Developing millennial tree ring chronologies in the fire-prone North American boreal forest. J Quaternary Sci. 28(3):283–292. doi:https://doi.org/10.1002/jqs.2612.
- Arseneault D, Payette S. 1992. A postfire shift from lichen-spruce to lichen-tundra vegetation at tree line. Ecology. 73(3):1067–1081. doi:https://doi.org/10.2307/1940181.
- Arseneault D, Payette S. 1997a. Reconstruction of millennial forest dynamics from tree remains in a subarctic tree line peatland. Ecology. 78(6):1873–1883. doi:https://doi.org/10.1890/0012-9658(1997)078[1873:ROMFDF]2.0.CO;2.
- Arseneault D, Payette S. 1997b. Landscape change following deforestation at the arctic tree line in Québec, Canada. Ecology. 78:693–706.
- Arseneault D, Payette S. 1998. Chronologie des cernes pâles de l’épinette noire (Picea mariana [Mill.] B.S.P.) au Québec subarctique: de 706 à 1675 ap. J.-C. Géographie physique et Quaternaire. 52:1–8.
- Asselin H. 2005. Dynamique végétale d’une toposéquence subarctique. Interaction entre le climat et les incendies forestiers [PhD dissertation]. Québec: Université Laval.
- Asselin H, Payette S. 2005a. Late Holocene opening of the forest tundra landscape in northern Québec, Canada. Global Ecol Biogeogr. 14(4):307–313. doi:https://doi.org/10.1111/j.1466-822X.2005.00157.x.
- Asselin H, Payette S. 2005b. Late Holocene deforestation of a tree line site: estimation of pre-fire vegetation composition and black spruce cover using soil charcoal. Ecography. 28:801–805.
- Asselin H, Payette S. 2006. Origin and long-term dynamics of a subarctic tree line. Écoscience. 13(2):135–142. doi:https://doi.org/10.2980/i1195-6860-13-2-135.1.
- Autin J, Gennaretti F, Arseneault D, Bégin Y. 2015. Biases in RCS tree ring chronologies due to sampling heights of trees. Dendrochronologia. 36:13–22. doi:https://doi.org/10.1016/j.dendro.2015.08.002.
- Barber VA, Juday GP, Finney BP. 2000. Reduced growth of Alaskan white spruce in the twentieth century from temperature-induced drought stress. Nature. 405(6787):668–673. doi:https://doi.org/10.1038/35015049.
- Bégin C. 1991. Analyse architecturale et dendroécologique d’une pessière à lichens à la limite des forêts. [PhD dissertation]. Québec: Université Laval.
- Bégin C, Filion L. 1999. Black spruce (Picea mariana) architecture. Can J Bot. 77:664–672.
- Bégin Y, Payette S. 1988. Dendroecological evidence of lake-level changes during the last three centuries in subarctic Québec. Quaternary Res. 30(2):210–220. doi:https://doi.org/10.1016/0033-5894(88)90025-7.
- Berger A. 1978. Long-term variations of daily insolations and Quaternary climatic changes. J Atmos Sci. 35(12):2362–2367. doi:https://doi.org/10.1175/1520-0469(1978)035<2362:LTVODI>2.0.CO;2.
- Bhiry N, Payette S, Robert E. 2007. Peatland development at the arctic tree line (Québec, Canada) influenced by flooding and permafrost. Quaternary Res. 67(3):426–437. doi:https://doi.org/10.1016/j.yqres.2006.11.009.
- Bradley R. 2014. Paleoclimatology. Reconstructing climates of the Quaternary. 3rd ed. New York (NY): Academic Press.
- Briffa KR, Jones PD, Bartholin TS, Eckstein D, Schweingruber FH, Karlen W, Zetterberg P, Eronen M. 1992. Fennoscandian summers from AD 500: temperature changes on short and long timescales. Clim Dynam. 7(3):111–119. doi:https://doi.org/10.1007/BF00211153.
- Brown PM, Kaufmann MR, Shepperd WD. 1999. Long-term, landscape patterns of past fire events in a montane ponderosa pine forest of central Colorado. Landscape Ecol. 14(6):513–532. doi:https://doi.org/10.1023/A:1008137005355.
- Büntgen U, Arseneault D, É B, Churakova (Sidorova) OV, Gennaretti F, Crivellaro A, Mk H, Av K, Klippel L, PJ K, et al. 2020. Prominent role of volcanism in Common Era climate variability and human history. Dendrochronologia. 64:125757. doi:https://doi.org/10.1016/j.dendro.2020.125757.
- Büntgen U, Brazdil R, Heussner KU, Hofmann J, Kontic R, Kyncl T, Pfister C, Chroma K, Tegel W. 2011. Combined dendro-documentary evidence of Central European hydroclimatic springtime extremes over the last millennium. Quaternary Sci Rev. 30(27–28):3947–3959. doi:https://doi.org/10.1016/j.quascirev.2011.10.010.
- Caccianiga M, Payette S, Filion L. 2008. Biotic disturbance in expanding subarctic forests along the eastern coast of Hudson Bay. New Phytol. 178(4):823–834. doi:https://doi.org/10.1111/j.1469-8137.2008.02408.x.
- CEN. 2020. Données des stations climatiques de la région de la rivière Boniface au Nunavik, Québec, Canada, v. 1.3 (1988-2019). Nordicana D7, doi: https://doi.org/10.5885/45129SL-DBDA2A77C0094963.
- Corona C, Lopez Saez J, Stoffel M, Rovéra G, J-l E, Berger F. 2013. Seven centuries of avalanche activity at Echalp (Queyras massif, southern French Alps) as inferred from tree rings. Holocene. 23(2):292–304. doi:https://doi.org/10.1177/0959683612460784.
- Cyr S, Payette S. 2010. The origin and structure of wooded permafrost mounds at the arctic treeline in eastern Canada. Plant Ecol Divers. 3(1):35–46. doi:https://doi.org/10.1080/17550871003777176.
- Dahl-Jensen D, Mosegaard K, Gundestrup N, Clow GD, Johnsen SJ, Hansen AW, Balling N. 1998. Past temperatures directly from the Greenland Ice Sheet. Science. 282(5387):268–271. doi:https://doi.org/10.1126/science.282.5387.268.
- Delwaide A, Filion L. 1999. Dendroséries du pin blanc (Pinus strobus L.) et de la pruche de l’est (Tsuga canadensis L. Carr.) dans la région de Québec. Géographie physique et Quaternaire. 53(2):265–275. doi:https://doi.org/10.7202/004853ar.
- Delwaide A, Filion L, Payette S. 1991. Spatiotemporal distribution of light rings in subarctic black spruce, Québec. Can J Forest Res. 21(12):1828–1832. doi:https://doi.org/10.1139/x91-252.
- Dyke AS. 2004. Deglaciation with emphasis on central and northern Canada. In: Ehlers J, Gibbard PL, editors. Quaternary glaciations: extent and chronology. part II: North America. developments in quaternary science. Vol. 2, New York (NY): Elsevier; p. 373–424.
- Eiriksson J, Bartels-Jonsdottir HB, Cage AG, Gudmundsdottir ER, Klitgaard- Kistensen D, Marret F, Rodrigues T, Abrantes F, Austin WEN, Jiang H, et al. 2006. Variability of the North Atlantic current during the last 2000 years based on shelf bottom water and sea surface temperatures along an open ocean/shallow marine transect in western Europe. Holocene. 16(7):1017–1019. doi:https://doi.org/10.1177/0959683606hl991rp.
- Esper J, Cook ER, Krusic PJ, Peters K, Schweingruber FH. 2003. Tests of the RCS method for preserving low-frequency variability in long tree-ring chronologies. Tree-Ring Res. 59:81–98.
- Esper J, Niederer R, Bebi P, Frank D. 2008. Climate signal age effects evidence from young and old trees in the Swiss Engadin. Forest Ecol Manag. 255(11):3783–3789. doi:https://doi.org/10.1016/j.foreco.2008.03.015.
- Ferguson CW. 1969. A 7104-year annual tree-ring chronology for bristlecone pine, Pinus aristata, from the White Mountains, California. Tree-Ring Bull. 29:3–29.
- Filion L, Payette S, Gauthier L. 1985. Analyse dendroclimatique d’un krummholz à la limite des arbres, lac Bush, Québec nordique. Géographie physique et Quaternaire. 39(2):221–226. doi:https://doi.org/10.7202/032604ar.
- Filion L, Payette S, Gauthier L, Boutin Y. 1986. Light rings in subarctic conifers as a dendrochronological tool. Quaternary Res. 26(2):272–279. doi:https://doi.org/10.1016/0033-5894(86)90111-0.
- Gennaretti F, Arseneault D, Nicault A, Perreault L, Bégin Y. 2014. Volcano-induced regime shifts in millennial tree-ring chronologies from Northeastern North America. P Natl Acad Sci USA. 111(28):10077–10082. doi:https://doi.org/10.1073/pnas.1324220111.
- Girard F, Payette S, Gagnon R. 2011. Dendroecological analysis of black spruce in lichen-spruce woodlands of the closed-crown forest zone in eastern Canada. Écoscience. 18(3):279–294. doi:https://doi.org/10.2980/18-3-3438.
- Grove JM. 2004. Little ice ages: ancient and modern. Boca Raton (FL): Routledge.
- Hadley JL, Smith WK. 1983. Influence of wind exposure on needle desiccation and mortality for timberline conifers in Wyoming, USA. Arct Alp Res. 15(1):127–135. doi:https://doi.org/10.2307/1550988.
- Hadley JL, Smith WK. 1986. Wind erosion of leaf surface wax in alpine timberline conifers. Arct Alp Res. 67:12–19.
- Harland R, Asteman IP, Nordberg K. 2013. A two-millennium dinoflagellate cyst record from Gullmar Fjord, a Swedish skagerrak sill fjord. Palaeogeogr Palaeoclimatol Palaeoecol. 392:247–260. doi:https://doi.org/10.1016/j.palaeo.2013.09.006.
- Helama S, Jones PD, Briffa KR. 2017. Dark ages cold period: a literature review and directions for future research. Holocene. 27(10):1600–1606. doi:https://doi.org/10.1177/0959683617693898.
- Helama S, Timonen M, Lindholm M, Merilainen J, Eronen M. 2005. Extracting long-period climate fluctuations from tree-ring chronologies over timescales of centuries to millennia. Int J Climatol. 25(13):1767–1779. doi:https://doi.org/10.1002/joc.1215.
- Helama SK, Mielikäinen K, Timonen M, Eronen M. 2008. Finnish supra-long tree-ring chronology extended to 5634 BC. Norsk Geogr Tidsskrift. 62(4):271–277. doi:https://doi.org/10.1080/00291950802517593.
- Holmes RL. 1983. Computer-assisted quality control in tree ring dating and measurement. Tree-ring Bull. 43:69–78.
- Jacoby GC, Ivanciu IS, Ulan LD. 1988. A 263-year record of summer temperature for northern Quebec reconstructed from tree-ring data and evidence of a major climatic shift in the early 1800s. Palaeogeogr Palaeoclimatol Palaeoecol. 64(1–2):69–78. doi:https://doi.org/10.1016/0031-0182(88)90143-5.
- Kelly PE, Cook ER, Larson DW. 1994. A 1397-year tree-ring chronology of Thuja occidentalis from cliff faces of the Niagara escarpment, southern Ontario, Canada. Can J Forest Res. 24(5):1049–1057. doi:https://doi.org/10.1139/x94-137.
- LaMarche VC Jr, Harlan TP. 1973. Accuracy of tree-ring dating of bristlecone pine for calibration of the radiocarbon time scale. J Geophys Res. 78:8849–8858.
- Lamb HH. 1965. The early medieval warm epoch and its sequel. Palaeogeogr Palaeoclimatol Palaeoecol. 1:13–37. doi:https://doi.org/10.1016/0031-0182(65)90004-0.
- Lamb HH. 1977. Climate, past, present and future. Vol. 2, London: Methuen: Climatic history and the future.
- Lamb HH. 1995. Climate history and the modern world. New York: Routledge.
- Lavoie C. 1994. Dynamique holocène de la limite des forêts au Québec subarctique. [PhD dissertation]. Québec: Université Laval.
- Lavoie C, Payette S. 1992. Black spruce growth forms as a record of a changing winter environment at treeline, Québec, Canada. Arct Alp Res. 24(1):40–49. doi:https://doi.org/10.2307/1551318.
- Lavoie C, Payette S. 1994. Recent fluctuations of the recent spruce forest limit in subarctic Québec. J Ecol. 82(4):725–734. doi:https://doi.org/10.2307/2261438.
- Lavoie C, Payette S. 1997. Late-Holocene light-ring chronologies from subfossil black spruce stems in mires of subarctic Québec. Holocene. 7(2):129–137. doi:https://doi.org/10.1177/095968369700700201.
- Lescop-Sinclair K, Payette S. 1995. Recent advance of the arctic treeline along the eastern coast of Hudson Bay. J Ecol. 83(6):929–936. doi:https://doi.org/10.2307/2261175.
- Liang C, Filion L, Cournoyer L. 1997. Wood structure of biotically and climatically induced light rings in eastern larch (Larix laricina). Can J Forest Res. 27(10):1538–1547. doi:https://doi.org/10.1139/x97-117.
- Lieffers VJ, Rothwell RL. 1987. Rooting of peatland black spruce and tamarack in relation to depth of water table. Can J Bot. 65(5):817–821. doi:https://doi.org/10.1139/b87-111.
- Linderholm HW, Gunnarson BE, Liu Y. 2010. Comparing scots pine tree-ring proxies and detrending methods among sites in Jamtland, west-central, Scandinavia. Dendrochronologia. 28(4):239–249. doi:https://doi.org/10.1016/j.dendro.2010.01.001.
- Ljungqvist FC. 2010. A new reconstruction of temperature variability in the extra-tropical Northern Hemisphere during the last two millennia. Geogr Ann. 92A(3):339–351. doi:https://doi.org/10.1111/j.1468-0459.2010.00399.x.
- Luckman BH 1995. Calendar-dated, early little ice age glacier advance at Robson Glacier, British Columbia, Canada. Holocene. 5(2):149–159. doi:https://doi.org/10.1177/095968369500500203.
- Maher CT, Nelson CR, Larson AJ. 2020. Winter damage is more important than summer temperature for maintaining the krummholz growth form above alpine treeline. J Ecol. 108(3):1074–1087. doi:https://doi.org/10.1111/1365-2745.13315.
- Marchand PJ. 1987. Life in the cold:An introduction to winter ecology. Hanover (New Hampshire): University Press of New-England.
- Martin-Benito D, Pederson N, McDonald M, Krusic P, Fernandez JM, Buckley B, Anchukaitis KJ, D’Arrigo R, Andreu-Hayles L, Cook E. 2014. Dendrochronological dating of the world trade center ship, lower Manhattan, New York City. Tree-Ring Res. 70(2):65–77. doi:https://doi.org/10.3959/1536-1098-70.2.65.
- Masson-Delmotte V, Zhai P, Pörtner HO, Roberts D, Skea J, Shukla PR, Pirani A, Moufouma-Okia W, Péan C, Pidcock R, et al., editors. 2018. IPCC. 2018: global warming of 1.5°C. An IPCC special report on the impacts of global warming of 1.5°C above pre-industrial levels and related global greenhouse gas emission pathways, in the context of strengthening the global response to the threat of climate change, sustainable development, and efforts to eradicate poverty. Geneva (Switzerland): IPCC.
- Morin H, Payette S. 1986. La dynamique récente des combes à neige du golfe de Richmond (Québec nordique): une analyse dendrochronologique. Can J Bot. 64(9):2113–2119. doi:https://doi.org/10.1139/b86-278.
- Naurzbaev MM, Vaganov EA. 2000. Variation of early summer and annual temperature in east Taymir and Putoran (Siberia) over the last two millennia inferred from tree rings. J Geophys Res. 105(D6):7317–7326. doi:https://doi.org/10.1029/1999JD901059.
- Payette S. 1983. The forest tundra and the present tree-lines of the Northern Québec-Labrador Peninsula. Nordicana. 47:141–151.
- Payette S. 2007. Contrasted dynamics of Northern Labrador tree lines caused by climate change and migration lag. Ecology. 88(3):770–780. doi:https://doi.org/10.1890/06-0265.
- Payette S, Delwaide A. 1994. Growth of black spruce at its northern range limit in Arctic Quebec, Canada. Arct Alp Res. 26(2):174–179. doi:https://doi.org/10.2307/1551781.
- Payette S, Delwaide A. 2004. Dynamics of subarctic wetland forests over the past 1500 years. Ecol Monogr. 74(3):373–391. doi:https://doi.org/10.1890/03-4033.
- Payette S, Delwaide A, Morneau C, Lavoie C. 1994. Stem analysis of a long-lived black spruce clone at treeline. Arct Alp Res. 26(1):56–59. doi:https://doi.org/10.2307/1551877.
- Payette S, Delwaide A, Morneau C, Lavoie C. 1996. Patterns of tree stem decline along a snow-drift gradient at treeline: a case study using stem analysis. Can J Bot. 74(11):1671–1683. doi:https://doi.org/10.1139/b96-203.
- Payette S, Filion L, Delwaide A, Bégin C. 1989. Reconstruction of tree-line vegetation responses to long-term climate change. Nature. 341(6241):429–432. doi:https://doi.org/10.1038/341429a0.
- Payette S, Filion L, Gauthier L, Boutin Y. 1985. Secular climate change in old-growth tree-line vegetation of northern Quebec. Nature. 315(6015):135–138. doi:https://doi.org/10.1038/315135a0.
- Payette S, Gagnon R. 1985. Late Holocene deforestation and tree regeneration in the forest-tundra of Québec. Nature. 313(6003):570–572. doi:https://doi.org/10.1038/313570a0.
- Payette S, Morneau C. 1993. Holocene relict woodlands at the eastern Canadian treeline. Quaternary Res. 39(1):84–89. doi:https://doi.org/10.1006/qres.1993.1010.
- Pereg D, Payette S. 1998. Development of black spruce growth forms at treeline. Plant Ecol. 138(2):137–147. doi:https://doi.org/10.1023/A:1009756707596.
- Porter SC. 1981. Glaciological evidence of Holocene climatic change. In: Wigley TML, Ingram MJ, Farmer G, editors. Climate and history studies on past climates and their impact on man. Cambridge: Cambridge University Press; p. 82–110.
- Querrec L, Filion L, Auger R, Arseneault D. 2009. Tree-ring analysis of white cedar (Thuja occidentalis L.) archaeological and historical wood in Québec City (Québec, Canada). Dendrochronologia. 27(3):199–212. doi:https://doi.org/10.1016/j.dendro.2009.05.004.
- R Core Team. 2019. R: a language and environment for statistical computing. Vienna (Austria): R Foundation for Statistical Computing.
- Rochner ML, Kelley HW, Wilson CS, Bennett T, Grissino-Mayer HD. 2017. Dendrochronological dating of the warner house and barn (20LV334), Livingston County, Michigan, USA. Dendrochronologia. 43:4–11. doi:https://doi.org/10.1016/j.dendro.2016.11.002.
- Salzer MW, Pearson CL, Baisan CH 2019. Dating the methuselah walk bristlecone pine floating chronologies. Tree-Ring Res. 75(1):61–66. doi:https://doi.org/10.3959/1536-1098-75.1.61.
- Stuiver M, Reimer PJ, Reimer RW. 2020. CALIB 8.2. http://calib.org; [ accessed 2020 Sept 13].
- Sullivan PF, Pattison RR, Brownlee AH, Cahoon SMP, Hollingsworth TN. 2017. Limited evidence of declining growth among moisture‐limited black and white spruce in interior Alaska. Sci Rep. 7(1):1–14. doi:https://doi.org/10.1038/s41598-017-15644-7.
- Vallée S, Payette S. 2004. Contrasted growth of black spruce (Picea mariana) forest trees at treeline associated with climate change over the last 400 years. Arct Antarct Alp Res. 36(4):400–406. doi:https://doi.org/10.1657/1523-0430(2004)036[0400:CGOBSP]2.0.CO;2.
- Vézeau C, Payette S. 2016. Gap expansion in old-growth subarctic forests: the climate-pathogen connection. New Phytol. 212(4):1044–1056. doi:https://doi.org/10.1111/nph.14081.
- Walker MJC, Berkelhammer M, Björck S, Cwynar LC, Fisher DA, Long AJ, Lowe JJ, Newnham RM, Rasmussen SO, Weiss H. 2012. Formal subdivision of the Holocene series/epoch: a discussion paper by a working group of INTIMATE (integration of ice-core, marine and terrestrial records) and the subcommission on quaternary stratigraphy (international commission on stratigraphy). J Quat Sci. 27:649–659.
- Wang F, Arseneault D, É B, Yu S, Ouellet S, Chaillou G, Delwaide A, Wang L. 2020. Chemical destaining and the delta correction for blue intensity measurements of stained lake subfossil trees. Biogeosciences. 17(D6):4559–4570. doi:https://doi.org/10.5194/bg-17-4559-2020.
- Wang L, Payette S, Bégin Y. 2000. A quantitative definition of light rings in black spruce (Picea mariana) at the Arctic treeline in Northern Québec, Canada. Arct Antarct Alp Res. 32(3):324–330. doi:https://doi.org/10.1080/15230430.2000.12003370.
- Wang L, Payette S, Bégin Y. 2002. Relationships between anatomical and densitometric characteristics of black spruce and summer temperature at tree line in northern Quebec. Can J Forest Res. 32(3):477–486. doi:https://doi.org/10.1139/x01-208.
- Wang T, Surge D, Mithen S. 2012. Seasonal temperature variability of the Neoglacial (3300–2500 BP) and roman warm period (2500–1600 BP) reconstructed from oxygen isotope ratios of limpet shells (Patella vulgata), Northwest Scotland. Palaeogeogr Palaeoclimatol Palaeoecol. 317-318:104–113. doi:https://doi.org/10.1016/j.palaeo.2011.12.016.
- Wanner H, Solomina O, Grosjean M, Ritz SP, Jetel M. 2011. Structure and origin of Holocene cold events. Quaternary Sci Rev. 30(21–22):3109–3123. doi:https://doi.org/10.1016/j.quascirev.2011.07.010.
- Wigley TML, Briffa KR, Jones PD. 1984. On the average of correlated time series, with applications in dendroclimatology and hydrometeorology. J Clim Appl Meteorol. 23(2):201–213. doi:https://doi.org/10.1175/1520-0450(1984)023<0201:OTAVOC>2.0.CO;2.
- Wilson R, Anchukaitis K, Andreu-Hayles L, Cook E, D’Arrigo R, Davi N, Haberbauer L, Krusic P, Luckman B, Morimoto D, et al. 2019. Improved dendroclimatic calibration using blue intensity in the southern Yukon. Holocene. 29(11):1817–1830. doi:https://doi.org/10.1177/0959683619862037.
- Wilson R, Anchukaitis K, Briffa KR, Buntgen U, Cook E, D’Arrigo R, Davi N, Esper J, Frank D, Gunnarson B, et al. 2016. Last millennium northern hemisphere summer temperatures from tree rings: part I: the long term context. Quaternary Sci Rev. 134:1–18. doi:https://doi.org/10.1016/j.quascirev.2015.12.005.
- Wolken JM, Mann DH, Grant III TA, Lloyd AH, Scott Rupp T, Hollingsworth TN. 2016. Climate-growth relationships along a black spruce toposequence in interior Alaska. Arct Antarct Alp Res. 48(4):637–652. doi:https://doi.org/10.1657/AAAR0015-056.
- Zang C, Biondi F. 2013. Dendroclimatic calibration in R: the boot res package for response and correlation function analysis. Dendrochronologia. 31(1):68–74. doi:https://doi.org/10.1016/j.dendro.2012.08.001.