Abstract
Cotton produces more biomass and economic yield when cluster planting pattern (three plants per hole) than in a traditional planting pattern (one plant per hole), even at similar plant densities, indicating that individual plant growth is promoted by cluster planting. The causal factors for this improved growth induced by cluster planting pattern, the light interception, canopy microclimate and photosynthetic rate of cotton were investigated in an arid region of China. The results indicated that the leaf area index and light interception were higher in cluster planting, and significantly different from those in traditional planting during the middle and late growth stages. Cotton canopy humidity at different growth stages was increased but canopy temperatures were reduced by cluster planting. In the later growth stage of cluster planting, the leaf chlorophyll content was higher and the leaf net photosynthetic rate and canopy photosynthetic rate were significantly increased in comparing with traditional planting pattern. We concluded that differences in canopy light interception and photosynthetic rate were the primary factors responsible for increased biomass production and economic yield in cluster planting compared with the traditional planting of cotton.
Comparisons of the growth and development of cotton grown in different planting patterns revealed that lint yield and water-use efficiency were higher with three plants per hole (cluster planting pattern) than with two or one plant per hole (traditional planting pattern) in the northwestern arid regions of China, although aboveground biomass did not differ significantly (Su et al., Citation2009). Through further research, we found that cluster planting with 50 cm between wide rows, 30 cm between narrow rows, and 28 cm between holes had the highest yield, and biomass production was also significantly increased (Xie et al., Citation2014).
Among the characteristics that contribute to increased economic yield and water-use efficiency, canopy microclimate is considered to be very important. Few studies have compared light interception and other microclimate characteristics in crops grown in different row-spacing systems, and none have examined cluster planting. Greater light interception has been reported with narrow row spacing in cotton (Gossypium hirsutum L.) (Heitholt et al., Citation1992) and other crops such as lucerne (Medicago sativa L.) (Mattera et al., Citation2013), maize (Zea mays L.) (Liu & Song, Citation2012) and potato (Solanum tuberosum L.) (Tarkalson et al., Citation2012). Gwathmey and Clement (Citation2010) found that early canopy development (leaf area) and, consequently, early light interception was higher in ultra-narrow rows than with conventional spacing. Yang et al. (Citation2008) reported that leaf temperature increased but humidity decreased with 15 cm row spacing than conventional row spacing, and this microclimate was better for photosynthesis. This increased light interception and optimized temperature and humidity in the growing season are suggested to be the primary reasons for increased yield in certain row-spacing systems.
Changes in yield and biomass production can also result from differences in photosynthetic capacity in later development, as observed in the cultivar Akenohoshi and Nipponbare of rice (Oryza sativa L.) (Jiang et al., Citation1988a), wheat (Triticum aestivum L.) (Nakagami et al., Citation2004) and maize (Zea mays L.) (Ma & Dwyer, Citation1998). The leaf photosynthesis of these plants with delayed leaf senescence was kept high in later growth stage. However, no studies have compared the photosynthetic capacity of cotton in cluster and traditional planting. San-oh et al. (Citation2006) found that one rice plant per hill produced more dry matter than three plants per hill, particularly during the ripening stage, and one reason could be that the former maintained higher rates of leaf net photosynthesis than the latter. Wang et al. (Citation2004) indicated the chlorophyll levels were higher for bed planting with furrow irrigation than flat planting with flood irrigation at all stages of wheat development and growth, especially during the later stages of the grain-fill period. Thus, the bed planting with furrow irrigation may lead to the longer ‘‘stay-green’’ and grain-fill period and therefore corresponding for higher grain yield.
To understand how the cluster planting pattern affects physiological characteristics contributing to higher yield, we tested the following hypotheses: (i) the cluster planting pattern increases light interception and improves microclimate and (ii) canopy development leads to greater photosynthesis in the late growth stage in cluster planting pattern.
2. Materials and methods
2.1. Experimental design
This study was conducted from late April to October (growing season) in 2012 and 2013 at the Linze Inland River Basin Research Station (39°21′N 100°02′E, 1,400 m a.s.l.), located in the oasis edge region of the northern part of Linze in the middle reach of Hexi Corridor in Gansu Province, northwest China. Traditional (one plant per hole) and cluster (three plants per hole) planting patterns were compared in both seasons. The traditional planting pattern had 30 cm between wide rows, 20 cm between narrow rows and 15 cm between holes (26.7 × 104 plants ha−1), while the cluster planting pattern had 50 cm between wide rows, 30 cm between narrow rows and 28 cm between holes (26.8 × 104 plants ha−1) (Figure ). A completely randomized plot design with six replications was used, and the area of each plot was 16 m2 (4 m × 4 m).
Figure 1. A schematic diagram showing traditional planting pattern and cluster planting pattern.
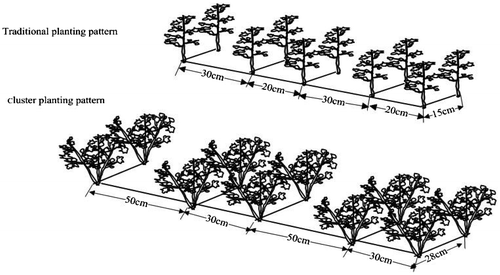
The cotton cultivar Xinluzao8 was used for this study and certain amount of cotton seeds was planted in sandy soil in late April and harvested in late September and early October. Five healthy seeds were sowed in one hole for cluster planting and three healthy seeds were sowed in one hole for traditional planting. Cotton seedlings were thinned when the height reached about 6–10 cm. 112 kg N ha−1 as urea and 56 kg P2O5 ha−1 as diammonium phosphate were applied just before sowing and at the squaring stage, and 56 kg N ha−1 as urea were applied at the bolling stage. During the growing season, precipitation was 92 mm in 2012 and 105 mm in 2013, and total water input was 6,000 m3 ha−1 by flood irrigation.
2.2. Leaf area index
Leaf area index (LAI) was measured by destructive sampling every 10 days beginning from the 60th day after sowing in six plots. Leaf area was determined by measuring sub-samples (6 plants per plot) with a Li-Cor planimeter (Model Li-3100, Li-COR Inc., Lincoln, NE, USA). These samples were dried and weighed, and specific leaf area was determined (m2 g−1). LAI was calculated as the product of specific leaf area and the amount of leaf dry matter (g m−2), and the plants in 1-meter-square were taken for leaf dry matter.
2.3. Light interception
The incoming photosynthetically active radiation (PAR) was measured at different heights in narrow and wide rows at three growth stages (full-flowering, full-bolling, and boll-opening). The heights were set as above the canopy (50 cm), and in the upper (3/4 of plant height), middle (1/2 of plant height) and lower (5 cm of above ground) of canopy layers using a sun scan canopy analysis system (Delta T Devices Ltd., Cambridge, UK). Six points (three points in the narrow rows and three points in the wide rows) per layer were measured within each plot between 11:00 and 13:00 h on a clear day, and light interception was calculated as the average value of narrow and wide rows. The proportion of PAR intercepted by the crop was calculated as: Li = (I0 − I)/I0 where I0 is the measured incident solar radiation at the top of the canopy and I is the transmitted radiation within the canopy.
2.4. CO2 concentration, relative humidity and temperature
The concentration of CO2 was measured by a portable infrared CO2 analyzer (GXH.3051, Beijing square physical and Chemical Technology Research Institute, Beijing, China) at the three different heights: the upper (3/4 of plant height), the middle (1/2 of plant height) and the lower (5 cm of above ground) canopy layers. Six points (three points in the narrow rows and three points in the wide rows) per layer were measured within each plot from 11:00 to 12:00 h on a clear day, and CO2 concentration was calculated as the average value of narrow and wide rows.
The humidity and temperature were measured by aspirated psychrometer (DHM-2A, Longtuo Instrument Equipment Ltd. Co., Shanghai, China). The measured location and time was same as for the measurement of CO2 concentration.
2.5. Leaf net photosynthesis
Leaf net photosynthetic rate (PN) was measured at the three growth stages. Single-leaf PN was recorded on the second or third fully expanded leaf from the top of the main axis using a Li-6400 portable photosynthetic system (Li-COR) between 12:00 and 14:00 local time. At measuring conditions, the photosynthetic photon flux density was 1900 μmol m−2 s−1, the ambient CO2 concentration (Ca) was 366 μmol mol−1, and the vapor pressure deficit was 3.5 kPa. Each measurement (3 readings per leaf) was replicated on three plants in each plot.
2.6. Leaf chlorophyll content
The content of leaf chlorophyll was measured in fully expanded leaves at the three growth stages using a portable chlorophyll meter (Model SPAD-502 Plus, Konica Minolta, Sakai, Japan). Within each plant, six leaves were measured to obtain an average SPAD value. Each measurement was replicated on three plants in each plot.
2.7. Canopy apparent photosynthesis
Canopy photosynthetic rate (CAP) was measured at the three growth stages in an assimilation chamber (Beijing Ecotek Ltd. Co., Beijing, China) with a Li-8100 (Li-COR). The Li-8100 is a fully automated system for measuring soil CO2 flux. It uses IRGA to measure changes in CO2 and H2O concentrations in the chamber. The assimilation chamber (50 cm × 50 cm × 100 cm) was made of an acrylic material that allowed more than 95% light transmittance. Four fans ensured that the gas in the chamber was thoroughly mixed. The temperature was monitored with two temperature sensors, one inside and one outside the chamber. Temperature was controlled by electronic cooling components. A small cylinder attached to the roof and wall and driven by a small compressor opened and closed the roof, and the opened roof make the inner of assimilation chamber be in field conditions at interval between measurements, and the upper and lower edges of the chamber were sealed to ensure that it was gas tight. The assimilation chamber was connected with the Li-8100. When the measurement parameters were set up, the instrument operated automatically.
The pedestal which was used to place the assimilation chamber was a square frame of stainless steel, with an internal side length of 49 cm, an external side length of 51, and 3 cm height. The pedestal was embedded in the plots one day before the canopy photosynthetic measurement, and the periphery of the pedestal and the gaps of ground was filled with soil and compacted, thus to ensure the air tightness of the measurement system with soil. The assimilation chamber was placed on the pedestal and constituted a closed system with the Li-8100 for measurements. The measurements were taken once an hour between 12:00 and 14:00 local time. The measurement duration was 4 min, repeated three times to obtain average values. Six cotton plants were measured in the assimilation chamber in each plot in traditional planting and twelve cotton plants were measured in each plot in cluster planting.
2.8. Yield and component
Six plots in cluster or traditional planting pattern were used for yield measurement; it was measured by hand picking all open bolls in each plot. The number of bolls was recorded and seed cotton samples were ginned in a 10-saw gin. Seed cotton yield, lint yield (kg ha−1), lint percentage (%), lint per boll (g boll−1) and average boll size (g seed cotton boll−1) were calculated.
2.9. Statistical analyses
Statistical analyses were conducted using SPSS (IBM, Chicago, IL, USA). The t-test was performed at different significance levels to determine whether differences existed between cluster and traditional planting treatments. All graphical constructions were completed using the Origin 8.0 software package.
3. Results
3.1. LAI and light interception
Early LAI (<110 d after sowing) was not significantly different between cluster and traditional planting, but it tended to be significantly higher in cluster planting than traditional planting after 110 d from sowing (Figure ). Light interception of cotton in different layers in cluster planting was significantly higher than in traditional planting (Table ). At the boll-opening stage, light interception in cluster planting was also significantly higher than traditional planting in all layers (Table ).
Figure 2. Leaf area index of cotton at different growth stages in traditional and cluster planting patterns in 2012. Values represent means ± SD (n = 6).
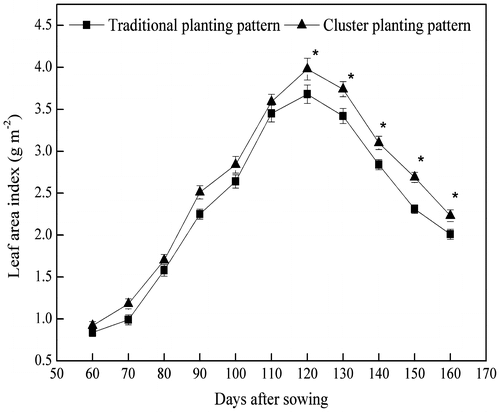
Table 1. Canopy light interception (Li) of cotton in different layers in three growths stages between traditional and cluster planting pattern in 2012.
3.2. CO2 concentration, temperature and relative humidity
Cluster planting affected the CO2 concentration within the canopy of cotton (Table ); CO2 concentrations in the same layer and growth stage were higher in cluster planting than in the traditional planting pattern; however, the differences were not significant (p > .05).
Table 2. Canopy CO2 concentration, temperature and relative humidity of cotton in three growth stages between traditional and cluster planting pattern in 2012.
Temperatures in the upper and lower layers were lower than those in the middle layer in any planting patterns. Temperatures in the cluster planting pattern were lower than those in the traditional pattern, but the differences were not significant (p > .05). The RH in cluster planting was significantly higher than that in traditional planting in all three growth stages (p < .05), indicating that RH in the canopy was significantly increased under cluster planting.
3.3. Leaf chlorophyll content
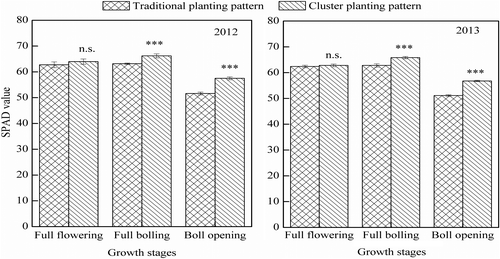
The SPAD values in two years in the cluster planting pattern were significantly higher than those in traditional planting at the full-bolling and boll-opening stages (p < .001) (Figure ). At the full-opening stage, the SPAD values in the cluster planting pattern were 11.2% higher than those in the traditional pattern. The years had significant effect on SPAD values in the full-bolling and full-opening stages (p < .05), but the interaction of planting pattern treatments × years was not significant in the three growth stages (p > .05).
Figure 3. Leaf chlorophyll content (SPAD) of cotton at different growth stages in traditional and cluster planting patterns in 2012 and 2013. Values represent means ± SD (n = 6). n.s.: not significant. ***p < .001.
3.4. Leaf and canopy net photosynthetic rates
Table showed that the patterns of variation in PN and CAP were similar under traditional and cluster planting patterns, with a rise from the full-flowering to the full-bolling stage but a subsequently decline in the boll-opening stage. PN and CAP were higher in cluster planting than those in traditional planting at full bolling and boll opening stages. In the full-flowering stage, PN and CAP were significantly effected by years (p < .01), planting pattern treatments and planting pattern treatments × years interaction had no significant effect on PN and CAP (p > .05). In the full bolling and boll opening stages, planting pattern treatments had extremely significant effect on PN and CAP (p < .001); the years had significant effect on PN (p < .01) and CAP (p < .05) in the full-bolling stage, and it had extremely significant effect on PN (p < .001) and CAP (p < .01) in the boll-opening stage. The interaction of planting pattern treatments × years had no significant effect on PN and CAP (p > .05) except of the PN on boll-opening stage.
Table 3. Leaf and canopy photosynthetic rate of cotton in three growth stages between traditional and cluster planting pattern in 2012 and 2013 (μmolCO2 m−2 s−1).
3.5. Cotton yield
A combined analysis across 2 years showed that seed cotton yield and lint yield, and final boll numbers in the cluster planting pattern were significantly higher than those in the traditional planting pattern. Lint percentage was higher and mean boll size was larger in the cluster planting pattern than those in the traditional pattern (Table ). Effects of planting pattern treatments and years had extremely significant effect on yield and yield components (p < .001), but planting pattern treatments only had significant effect on final boll numbers (p < .05). The interaction of planting pattern treatments and years had extremely significant effect on yield (p < .001), but it had significant effect on lint percentage and mean boll size (p < .05) except of final boll numbers.
Table 4. Effect of planting pattern treatments on cotton yield and yield components in 2012 and 2013.
4. Discussion
Canopy development and consequent light interception in the middle and late growth stages were higher in cluster planting than those in the traditional planting, and canopy development and light interception were significantly different from those in traditional planting in the two growth stages. These two parameters followed a similar pattern to biomass accumulation and crop growth rate. This pattern of light interception and LAI in cluster planting contradicted the results of Brodrick et al. (Citation2013) in which the canopy development and light interception in the early growth stage were higher in ultra-narrow row crops, but the canopy development and light interception were lower in ultra-narrow row crops in later growth stage. Existing research shows that, under the field conditions, light interception is the most important factor influencing photosynthesis (Agele et al., Citation2007; Kim et al., Citation2006, Citation2007; Maddonni et al., Citation2006), thereby, we inferred that the greater light interception in the cluster planting pattern benefitted cotton photosynthesis.
Photosynthesis is affected by many interacting external conditions, such as light, CO2 concentration, temperature and RH. In this study, we found RH in the canopy had significant difference between the cluster planting and traditional planting, which probably results from the combined effects of temperature, intercepted PAR and ventilation. In our previous study, we found that cotton cultivated in cluster planting produced more biomass and lint yield (Xie et al., Citation2014). We assumed that increased economic yield was a consequence of higher net photosynthetic rate in later growth stage. In general, the net photosynthetic rates of plants have been examined in fully expanded leaves (Murata, Citation1961) and during leaf senescence (Jiang et al., Citation1988b; Makino et al., Citation1985). At the full-flowering stage, there were no significant differences in the leaf and CAPs between traditional and cluster planting (Table ). These results indicated that cotton’s capacity for photosynthesis was similar in both planting patterns in early growth. During senescence (boll-opening stage), cottons in cluster planting pattern maintained higher rates of leaf and canopy net photosynthesis than in traditional planting pattern (Table ). This higher photosynthesis in cluster planting pattern maybe associated with the delay in leaf senescence which related with the higher levels of chlorophyll content in the leaves in this stage. The delays in leaf senescence under different planting patterns have been reported in rice (Oryza sativa L.) (San-oh et al., Citation2006), wheat (Triticum aestivum L.) (Nakamura et al., Citation2003), and maize (Zea mays L.) (Fujita et al., Citation2002). In these plants, the rate of leaf photosynthesis was kept high during ripening. The rate of photosynthesis during senescence was also correlated with levels of Rubisco in the leaves (Makino et al., Citation1985) and a strong correlation has been observed between leaf nitrogen and Rubisco content during senescence (Makino et al., Citation1984; Ookawa et al., Citation2004), so these factors will be measured in the next study.
5. Conclusion
Cotton grown in the cluster planting pattern had higher economic yield because of higher levels of light interception, leaf chlorophyll content, and leaf and canopy photosynthesis in the middle and late growth stages due to improved microclimate compared with the traditional planting pattern.
| Abbreviations | ||
| Li | = | light interception; |
| PAR | = | photosynthetically active radiation; |
| LAI | = | leaf area index; |
| RH | = | relative humidity; |
| PN | = | net photosynthetic rate; |
| PPFD | = | photosynthetic photon flux density; |
| Ca | = | ambient CO2 concentration; |
| VPD | = | vapor pressure deficit; |
| CAP | = | canopy photosynthetic rate. |
Disclosure statement
No potential conflict of interest was reported by the authors.
Acknowledgements
The study was carried out at Linze Inland River Basin Research Station.
Funding
This work was supported by the National Natural Sciences Foundation of China [31300323], [91325104], [41361100]; China Postdoctoral Science Foundation Funded Project [2014M552515].
References
- Agele, S. O., Maraiyesa, I. O., Adeniji, I. A. (2007). Effects of variety and row spacing on radiation interception, partitioning of dry matter and seed set efficiency in late season sunflower (Helianthus annuus L.) in a humid zone of nigeria. African Journal of Agricultural Research, 2, 80–88.
- Brodrick, R., Bange, M. P., Milroy, S. P., & Hammer, G. L. (2013). Physiological determinants of high yielding ultra-narrow row cotton: Canopy development and radiation use efficiency. Field Crops Research, 148, 86–94.10.1016/j.fcr.2012.05.008
- Fujita, M., Ookawa, T., & Hirasawa, T., (2002). Effects of soil moisture conditions before flowering on photosynthesis rate, nitrogen accumulation and cytokinin activity in xylem exudates during ripening in maize plants. Japanese Journal of Crop Science, 71, 170–171.
- Gwathmey, C. O., & Clement, J. D. (2010). Alteration of cotton source-sink relations with plant population density and mepiquat chloride. Field Crops Research, 116, 101–107.10.1016/j.fcr.2009.11.019
- Heitholt, J. J., Pettigrew, W., & Meredith, W., (1992). Light interception and lint yield of narrow-row cotton. Crop Science, 32, 728–733.10.2135/cropsci1992.0011183X003200030030x
- Jiang, C. Z., Hirasawa, T., & Isihara, K. (1988a). Physiological and ecological characteristics of high yielding varieties in rice plants. I. Yield and dry matter production. Japanese journal of crop science, 57, 132–138.10.1626/jcs.57.132
- Jiang, C. Z., Hirasawa, T., & Isihara, K. (1988b). Physiological and ecological characteristics of high yielding varieties in rice plants. II. Leaf photosynthetic rates. Japanese Journal of Crop Science, 57, 139–145.10.1626/jcs.57.139
- Kim, S. H., Sicher, R. C., Bae, H., Gitz, D. C., Baker, J. T.Timlin, J. D., & Reddy, V. R. (2006). Canopy photosynthesis, evapotranspiration, leaf nitrogen, and transcription profiles of maize in response to CO2 enrichment. Global Change Biology, 12, 588–600.10.1111/gcb.2006.12.issue-3
- Kim, S. H., Gitz, D., Sicherb, R. C., Baker, J. T., Timlin, D. J., & Reddy, V. R. (2007). Temperature dependence of growth, development, and photosynthesis in maize under elevated CO2. Environmental and Experimental Botany, 61, 224–236.10.1016/j.envexpbot.2007.06.005
- Maddonni, G. A., Cirilo, A. G., & Otegul, M. E. (2006). Row width and maize grain yield. Agronomy Journal, 98, 1532–1543.10.2134/agronj2006.0038
- Liu, T. D., & Song, F. B. (2012). Maize photosynthesis and microclimate within the canopies at grain-filling stage in response to narrow-wide row planting patterns. Photosynthetica, 50, 215–222.10.1007/s11099-012-0011-0
- Ma, B. L., & Dwyer, L. (1998). Nitrogen uptake and use of two contrasting maize hybrids differing in leaf senescence. Plant and Soil, 199, 283–291.10.1023/A:1004397219723
- Makino, A., Mae, T., & Ohira, K., (1984). Relation between nitrogen and ribulose-1,5-bisphosphate carboxylase in rice leaves from emergence through senescence. Plant and Cell Physiology, 25, 429–437.
- Makino, A., Mae, T., & Ohira, K., (1985). Photosynthesis and ribulose-1,5-bisphosphate carboxylase/oxygenase in rice leaves from emergence through senescence. Quantitative analysis by carboxylation/oxygenation and regeneration of ribulose-1,5-bisphosphate. Planta, 166, 414–420.10.1007/BF00401181
- Mattera, J., Romero, L. A., Cuatrin, A. L., Cornaglia, P. S., & Grimoldi, A. A (2013). Yield components, light interception and radiation use efficiency of lucerne (Medicago sativa L.) in response to row spacing. European Journal of Agronomy, 45, 87–95.10.1016/j.eja.2012.10.008
- Murata, Y. (1961). Studies on the photosynthesis of rice plants and its culture significance. Bulletin of the National Institute of Agricultural Sciences (Japan), D9, 1–169.
- Nakagami, K., Ookawa, T., & Hirasawa, T. (2004). Effects of a reduction in soil moisture from one month before flowering through ripening on dry matter production and ecophysiological characteristics of wheat plants. Plant Production Science, 7, 143–154.10.1626/pps.7.143
- Nakamura, E., Ookawa, T., Ishihara, K. & Hirasawa, T. (2003). Effects of soil moisture depletion for one month before flowering on dry matter production and ecophysiological characteristics of wheat plants in wet soil during grain filling. Plant Production Science, 6, 195–205.10.1626/pps.6.195
- Ookawa, T., Naruoka, Y., Sayama, A. & Hirasawa, T. (2004). Cytokinin effects on ribulose-1,5-bisphosphate carboxylase/oxygenase and nitrogen partitioning in rice during ripening. Crop Science, 44, 2107–2115.10.2135/cropsci2004.2107
- San-oh, Y., Sugiyama, T., Yoshita, D., Ookawa, T. & Hirasawa, T. (2006). The effect of planting pattern on the rate of photosynthesis and related processes during ripening in rice plants. Field Crops Research, 96, 113–124.10.1016/j.fcr.2005.06.002
- Su, P. X., Xie, T. T., & Ding, S. S. (2009). Experimental studies on high-yield cluster cultivation of cotton in the Hexi Corridor oases of northwestern China. Agricultural Research in the Arid Areas, 27, 108–113.
- Tarkalson, D. D., King, B. A., Biorneberg, D. L., & Taberna, J. P. (2012). Effects of planting configuration and in-row plant spacing on photosynthetically active radiation interception for three irrigated potato cultivars. Potato Research, 55, 41–58.10.1007/s11540-011-9205-2
- Wang, F. H., Wang, X. Q., & Ken, S. (2004). Comparison of conventional, flood irrigated, flat planting with furrow irrigated, raised bed planting for winter wheat in China. Field Crops Research, 87, 35–42.
- Xie, T. T., Su, P. X., Zhou, Z. J., Zhang, H. N., & Li, S. J. (2014). Effect of cluster planting on field microclimate and yield of cotton. Acta Agriculture Boreali-Occident Sin, 23, 55–61.
- Yang, W. P., Guo, T. C., Liu, S. B., Wang, C. Y., Wang, Y. H., & Ma, D. Y. (2008). Effects of row spacing in winter wheat on canopy structure and microclimate in later growth stage. Journal of Plant Ecology, 32, 485–490.