ABSTRACT
The health benefits of purple sweetpotato, which is used as an edible food in its natural state and in processed foods and as a natural color pigment, have been recognized. In Japan, sweetpotato has been economically produced in regions below 36°4ʹN latitude; however, cultivation areas are beginning to expand further north. The anthocyanin and polyphenolics in three purple sweetpotatoes cultivated in different locations; I (42°92ʹN, 143°04ʹE) and II (31°72ʹN, 131°03ʹE), were compared over two years. Total anthocyanin and polyphenolic contents in purple sweetpotatoes tended to be high in location I. The anthocyanin and polyphenolic compositions differed between locations. The peonidin/cyanidin ratios were higher in location II compared with I in all varieties. The relative amount of 3,4- and 4,5-di-O-caffeoyolquinic acids were higher in location II. Their contents and composition also differed between years. These results suggest that the variability of the anthocyanin and polyphenolic content and composition was influenced by climate conditions, probably temperature during cultivation. This study indicates that northern areas in Japan are an alternative production area and may yield higher amounts of anthocyanin and polyphenolics.
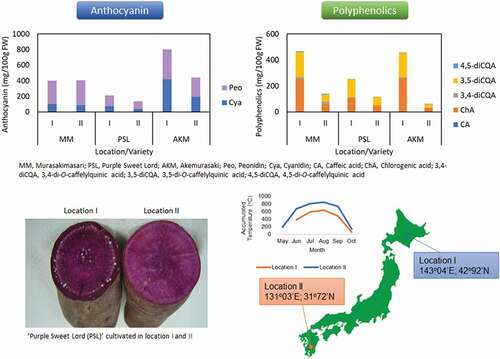
Graphical abstract
1. Introduction
Sweetpotato (Ipomoea batatas Lam.) is a tropical and subtropical root crop originally from central America or south America. It can be cultivated under a wide range of climatic conditions, between 30° and 40° latitude in both hemispheres, and in many countries, including Asia, Africa, America and Oceania (Paneque Ramirez, Citation2017). In Japan, sweetpotato is economically grown in regions below about 36°4ʹN latitude to produce starch, a distilled alcoholic drink, processed foods and natural color pigment as well as edible food. Sweetpotato has a number of health benefits due to its content of anthocyanins, carotenoids, polyphenolics, vitamins, minerals and dietary fibers (Tanaka et al., Citation2017).
Purple sweetpotatoes, which contain anthocyanins in their flesh, have been developed over the last two decades in Japan (Tanaka et al., Citation2017). Their production has gradually increased in the south of Japan and processed foods containing purple sweetpotato have been developed and become widely available since the first variety, Ayamusasaki, was released (Yoshinaga, Citation1995). The major coloring constituents in purple sweetpotato have been identified as peonidin- and cyanidin-based anthocyanins, named as YGM-1a, YGM-1b, YGM-2, YGM-3, YGM-4b, YGM-5a, YGM-5b and YGM-6 (Terahara et al., Citation1999) (). Considerable differences in anthocyanin compositions were recognized among purple sweetpotato varieties, and they can be classified into peonidin dominant type and cyanidin dominant type based on peonidin/cyanidin ratio (Montilla et al., Citation2011). Anthocyanins from purple sweetpotatoes show high heat and light stability due to their acylated structures (Montilla et al., Citation2011). Anthocyanin extracts, or the purified form, have been shown to have many physiological functions in vitro and in vivo: anti-oxidation (Kano et al., Citation2005), antimutagenicity (Yoshimoto et al., Citation2001), antihyperglycemic (Matsui et al., Citation2002), anti-atherosclerotic (Miyazaki et al., Citation2008), hepatoprotective and anti-hypertension (Suda et al., Citation2003) effects have been reported.
Figure 1. Structures of the sweetpotato anthocyanins (a) and polyphenolics (caffeoylquinic acids) (b). Glu, Glucopyranoside; HB, p-Hydroxybenzoic acid; Cf, Caffeic acid; Fr; Ferulic acid.
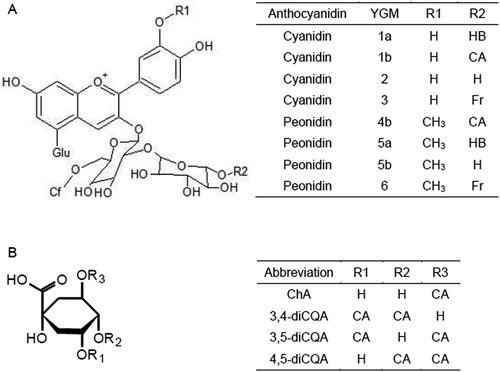
Sweetpotato storage roots contain polyphenolics, which are mainly caffeoylquinic acid derivatives, namely, caffeic acid (CA), chlorogenic acid (ChA), 3,4-di-O-caffeoylquinic acid (3,4-diCQA), 3,5-di-O-caffeoylquinic acid (3,5-diCQA) and 4,5-di-O-caffeoylquinic acid (4,5-diCQA) (Tanaka et al., Citation2017) (). Polyphenolics play an important role in in vivo defence systems against insects and other pathogens: the caffeoylquinic acids in sweetpotato roots have antifungal and antiviral activities (Harrison et al., Citation2001; Peterson et al., Citation2005). The concentration of these polyphenolics has been reported to increase as a function of stress, i.e. wounding, infection, drought, and storage at low temperature (Harrison et al., Citation2006; Ishiguro et al., Citation2007; Reyes et al., Citation2007; Uritani, Citation1999). Caffeoylquinic acids also have been shown to exhibit radical scavenging activity (Yoshimoto et al., Citation2006), antimutagenicity (Yoshimoto et al., Citation2002), anticancer activity (Kurata et al., Citation2007), anti-hyperglycaemic activity (Matsui et al., Citation2004), anti-inflammatory activity (Peluso et al., Citation1995), anti-HIV activity (Mahmood et al., Citation1993), and inhibit melanin production (Shimozono et al., Citation1996) in vitro or in vivo. In addition, there is a report that these caffeoylquinic acids suppress discoloration of anthocyanins (Nishiyma et al., Citation2000).
Sweetpotato cultivation has spread to the north region of Japan where they have been shown to have a higher content of sugars (Nakamura et al., Citation2014); however, a comparison of the physiological active components, such as anthocyanins and polyphenolics, in sweetpotatoes grown in different locations has not yet been performed. In this study, we evaluated the differences in anthocyanin and polyphenolics in purple sweetpotatoes cultivated in different locations at north and south in Japan over two years. The contents and compositions of anthocyanin and polyphenolics were compared in three varieties and two locations over the two years. In addition, the relationship between climate conditions to anthocyanin and polyphenolic contents and components, and the expected advantages of cultivation in the north region are discussed.
2. Materials and methods
2.1. Reagents and chemicals
ChA was obtained from Sigma-Aldrich Japan Co., LLC. (Tokyo, Japan). 3,4-diCQA, 4,5-diCQA and 3,5-diCQA were purchased from ChemFaces (Wuhan, China). Purified YGM-6 (96% purity) was used as a standard for anthocyanin analysis by HPLC. Other reagents were purchased from Wako Pure Chemicals Ind. (Osaka, Japan).
2.2. Sample preparation
Three purple sweetpotato varieties were grown in two different locations in Japan in 2015 and 2016. Murasakimasari (MM) is a variety used in food pigment production and a material for ‘shochu’, a liquor distilled from sweetpotato, and has a medium content of anthocyanin. Akemurasaki (AKM) is also used in food pigment production and has a high content of anthocyanin. Purple Sweet Lord (PSL) is used as an edible food and has a rather low content of anthocyanin. The experimental locations were as follows: location I, Memuro, Hokkaido (longitude, 143°04E; latitude, 42°92ʹN); location II, Miyakonojo, Miyazaki (longitude, 131°03ʹE; latitude, 31°72ʹN). The sweetpotatoes were cultivated using the standard local methods, including planting and harvesting dates; transplanted and harvested days in location I were early June and early October, while in location II were middle May and early October, respectively. Chemical fertilizer was applied to the fields at the rates of 1.8, 15.0 and 7.8 kg 10a−1 for N, P2O5 and K2O, respectively, in location I, and 3.6, 4.8 and 7.2 kg 10a−1 for N, P2O5 and K2O, respectively, in location II in both years. Climate conditions of both locations, provided by the Automated Meteorological Data Acquisition System (AMeDAS), in 2015 and 2016, including precipitation, mean temperature, mean maximum temperature, mean minimum temperature and sunshine duration, and accumulated temperature are listed in . Harvested sweetpotatoes were freeze-dried and held at −35°C until analysis. Freeze-dried samples were used for anthocyanin and polyphenolic analyses.
Table 1. Weather data during cultivation in 2015 and 2016
2.3. Anthocyanin analyses
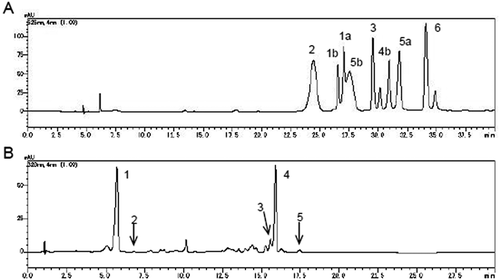
Anthocyanins were extracted according to the method by Oki et al. (Citation2011). One g of the freeze-dried samples was vigorously mixed in 9 mL of the extraction solution (methanol/water/trifluoroacetic acid = 40/60/0.5), mixed using a vortex mixer and sonicated in a water bath at 37°C for 5 min followed by continuous warming for 10 min. The extract was then centrifuged at 1,900 × g for 15 min and the supernatant was recovered. Extraction solution (8 mL) was added to the residue, and the extraction was repeated twice more. The extracts were combined and made up to 25 mL. A portion of the extract was filtered through a 0.45 μm membrane, and the anthocyanins were determined quantitatively by high-performance liquid chromatography (HPLC), as described (Terahara et al., Citation2007). The HPLC system consisted of two pumps (model LC-20AD; Shimadzu, Kyoto, Japan), an auto-injector (model SIL-20AC; Shimadzu, Kyoto, Japan), a column oven (model CTO-20AC; Shimadzu, Kyoto, Japan) and a photodiode array detector (model SPD-M20A; Shimadzu, Kyoto, Japan). A reversed phase column was used (250 × 4.6 mm i.d., Cadenza CD-C18, 3 μm: Imtakt Corp., Kyoto, Japan). The mobile phase consisted of water containing 0.4% formic acid (A) and 50% acetonitrile containing 0.4% formic acid (B). The elution profile was as follows: a linear gradient of 25% to 53% B from 0 to 40 min, and 25% B from 41 to 60 min at a flow rate of 0.6 mL min−1. The column oven was set at 30°C. Anthocyanins, YGM-1a, YGM-1b, YGM-2, YGM-3, YGM-4b, YGM-5a, YGM-5b and YGM-6, were identified by their retention time and the UV-vis spectra. Quantification of anthocyanins was performed using a YGM-6 external standard method based on detection at 520 nm. Each anthocyanin was quantified using each specific conversion factor (Terahara et al., Citation2007) and expressed as mg 100 g−1 fresh weight (FW).
2.4. Polyphenolic analyses
Polyphenolics were extracted as previously reported (Ishiguro et al., Citation2007). Freeze-dried samples (100 mg) were vigorously mixed in 10 mL of 80% ethanol, and boiled at 100°C for 5 min. A portion of the extract was filtered through a 0.45 μm membrane, and the caffeoylquinic acids were determined quantitatively by HPLC, as described, with slight modification (Ishiguro et al., Citation2007). The HPLC system was the same as for anthocyanin analysis. A reversed phase column was used (75 × 4.6 mm i.d., 3 μm, Cadenza CD-C18: Imtakt, Kyoto, Japan). The mobile phase consisted of water containing 0.2% formic acid (A) and acetonitrile (B). The elution profile was as follows: a linear gradient of 8% to 30% B from 0 to 23 min, and 8% B from 23.01 to 30 min at a flow rate of 1 mL min−1. The column oven was set at 40°C. Caffeoylquinic acids, ChA, CA, 3,4-diCQA, 3,5-diCQA, 4,5-diCQA, were identified by their retention time and the UV-vis spectra of standards. Quantification of caffeoylquinic acids was performed using external standards based on detection at 326 nm. Total polyphenolics content was quantified as total contents of caffeoylquinic acids, ChA, CA, 3,4-diCQA, 3,5-diCQA and 4,5-diCQA, and were expressed as mg 100 g FW−1.
2.5. Statistical analysis
Four replications were performed for all analyses. Three-way analysis of variance (ANOVA) was performed to evaluate the differences in the contents of anthocyanin and polyphenolics among variety, location and year by Excel Statistics (Bellcurve, Tokyo, Japan). The proportions of each caffeoylquinic acid in each variety among locations and years were arcsine transformed, followed by three-way ANOVA. When their interactions were significant, the simple main effects tests are performed. The statistical differences between groups were evaluated by Tukey’s multiple comparison test.
3. Results
3.1. Anthocyanin content and composition of purple sweetpotatoes cultivated in different locations
The anthocyanin content and composition of three purple sweetpotato varieties cultivated in two different locations were analyzed in 2015 and 2016. Eight known anthocyanins, YGM-1a, YGM-1b, YGM-2, YGM-3, YGM-4b, YGM-5a, YGM-5b and YGM-6 were detected in all three varieties cultivated in different locations (). Three-way ANOVA indicated that there were significant differences in total anthocyanin content among locations, varieties, and years as well as significant interactions among location and variety, location and year, variety and year, and location, variety and year (). Total anthocyanin contents in PSL and AKM with an average of two years were 1.6 and 1.8 times higher in location I (210 mg 100 g FW−1 and 802 mg 100 g FW−1, respectively) than II (135 mg 100 g FW−1 and 440 mg 100 g FW−1, respectively). Meanwhile, the content in MM with an average of two years was not significantly different between locations (, suppl Fig. S1). The total anthocyanin content with an average of two locations was significantly higher in 2015 than 2016 in all varieties (, suppl Fig. S1).
Table 2. Total anthocyanin, cyanidin, peonidin content and peonidin/cyanidin ratio in individual varieties cultivated in two locations in 2015 and 2016
Figure 2. Representative HPLC chromatogram of sweetpotato YGM-anthocyanins (a) and polyphenolics (b). A: Indicated numbers means YGM-anthocyanins. B: 1, ChA; 2, CA; 3, 3,4-diCQA; 4, 3,5-diCQA; 5, 4,5-diCQA.
Three-way ANOVA indicated that there were significant differences in cyanidin and peonidin content, and peonidin/cyanidin ratio among locations, varieties and years as well as significant interactions between location and variety, location and year, variety and year, and location and variety and year (). The cyanidin content with an average of two years were 1.1, 2.2 and 2.2 times higher in location I (MM: 98 mg 100 g FW−1, PSL: 75 mg 100 g FW−1 and AKM: 418 mg 100 g FW−1, respectively) than II (MM: 86 mg 100 g FW−1, PSL: 39 mg 100 g FW−1 and AKM: 191 mg 100 g FW−1, respectively), and the content with an average of two locations was significantly higher in 2015 than 2016 in all varieties (). The peonidin content with an average of two years in PSL and AKM were 1.5 times higher in location I (142 mg 100 g FW−1 and 384 mg 100 g FW−1, respectively) than II (97 mg 100 g FW−1 and 250 mg 100 g FW−1, respectively). In MM, the content with an average of two years was 1.1 times higher in location II (320 mg 100 g FW−1) than I (302 mg 100 g FW−1) (). The peonidin content with an average of two locations was higher in 2015 than 2016 in all varieties (). The peonidin/cyanidin ratios with an average of two years were higher in location II than I in all varieties (). The ratio with an average of both locations in MM was not significantly different between 2015 and 2016, in PSL was higher in 2015 than in 2016, and in AKM was higher in 2016 than in 2015 ().
These results indicated that the total anthocyanin content of purple sweetpotato was variable depending on variety, location and year, and was obviously higher in location I than II, especially in the variety with lower ratio of peonidin/cyanidin (AKM and PSL), and that the ratio of peonidin/cyanidin became higher in location II than I. While, the total anthocyanin content of the variety with higher ratio of peonidin/cyanidin (MM) was not significantly different between locations as a result of higher cyanidin content in location I and higher peonidin content in location II.
3.2. Polyphenolic content and composition of purple sweetpotatoes cultivated in different locations
The polyphenolic content and composition of three purple sweetpotato varieties cultivated in two different locations were analyzed in 2015 and 2016. Five known caffeoylquinic acids, ChA, CA, 3,4-diCQA, 3,5-diCQA and 4,5-diCQA, were detected by HPLC in all the varieties cultivated in different locations (). Three-way ANOVA indicated that there were significant differences in total polyphenolic content among locations, varieties and years as well as significant interactions between location and variety, variety and year, and location and variety and year (). The contents in MM, PSL and AKM with an average of two years were 3.3, 2.2 and 7.5 times higher in location I (466 mg 100 g FW−1, 254 mg 100 g FW−1 and 458 mg 100 g FW−1, respectively) than II (140 mg 100 g FW−1, 114 mg 100 g FW−1 and 61 mg 100 g FW−1, respectively) and the content with an average of two locations was much higher in 2015 than 2016 in all varieties (, supple Fig. S2).
Table 3. The contents and proportions of total polyphenolics and caffeoylquinic acids in individual varieties cultivated in two locations in 2015 and 2016
Three-way ANOVA indicated that there were significant differences in each caffeoylquinic acid content among locations, varieties and years as well as strong significant interactions (>1% significant level) among location and variety, variety and year, and location and variety and year, except the differences among locations, and the interaction among location and year, and location and variety and year were not significant in 3,4-diCQA (). ChA and 3,5-diCQA were the major polyphenolics in all three varieties cultivated in both locations (). All the contents of caffeoylquinic acids with an average of two years were higher in location I than II except for 3,4-diCQA and 4,5-diCQA content in MM. And their contents with an average for two locations were significantly higher in 2015 than in 2016 except for 3,4-diCQA content in AKM.
Three-way ANOVA indicated that there were significant differences in relative amount of individual caffeoylquinic acid among locations, varieties and years as well as significant interactions between location and variety, variety and year, and location and variety and year (). The proportion of ChA to total polyphenolics in MM and AKM with an average of two years were higher in location I than II, while in PSL were slightly higher in location II than I (). The proportion of 3,5-diCQA to total polyphenolics in MM was not significantly different between locations, in PSL was higher in location I than II, and in AKM was higher in location II than I (). Meanwhile, the proportions of other minor polyphenolics, 3,4-diCQA and 4,5-diCQA, in all varieties with an average of two years were higher in location II than I (). The proportion of ChA to total polyphenolics in MM with an average of both locations were higher in 2015 than 2016, in PSL was higher in 2016 than 2015 and in AKM was not significantly different between years (). The proportion of 3,5-diCQA to total polyphenolics in MM with an average of two locations was not significantly different between years, in PSL and AKM were higher in 2015 than 2016 (). The proportions of other minor polyphenolics, 3,4-diCQA and 4,5-diCQA, in all varieties with an average of two locations were higher in 2016 than 2015, with the single exception that the proportion of 4,5-diCQA to total polyphenolics in PSL was not significant between years ().
These results indicated that the total polyphenolic content of purple sweetpotato was variable depending on variety, location and year. The content was clearly higher in location I than II, and in 2015 than 2016. The proportions of major polyphenolics, ChA and 3,5-diCQA to total polyphenolics, were also variable depending on variety, location and year, and the tendency was somewhat different in varieties. While the proportions of other minor polyphenolics, 3,4-diCQA and 4,5-diCQA, showed higher content in location II than I, and in 2016 than 2015 in all varieties except a single exception.
4. Discussion
The anthocyanin and polyphenolic contents in plants are affected by environmental conditions, such as temperature, light, plant growth regulator, and sugars (Matsushita et al., Citation2016; Schwartz et al., Citation2009; Takos et al., Citation2006). The content of anthocyanins in the periderm of sweetpotatoes was higher when grown in lower temperature growth chambers (Villavicencio et al., Citation2007). A highly negative correlation was recognized between soil temperature and anthocyanin content in the flesh of purple sweetpotato (Kobayashi et al., Citation1998). In this study, the total anthocyanin contents in purple sweetpotatoes, AKM and PSL which showed lower peonidin/cyanidin rate, were significantly higher in location I than location II (, suppl Fig. S1). The total polyphenolics in all purple sweetpotato varieties also showed higher content in location I than in location II during the two years cultivation (, suppl Fig. S2). Location I is in the north of Japan and had the lower temperatures, while location II is in the south of Japan and had the higher temperatures during cultivation (). These results suggest that the north region of Japan is a good location for sweetpotato production to maximize anthocyanins and polyphenolics. Their contents in all varieties cultivated in both locations were significantly higher in 2015 than in 2016 ( and ). The average mean temperature, mean maximum temperature, mean minimum temperature and accumulated temperature were lower in 2015 compared with 2016 in the latter stages of cultivation in location I and throughout cultivation in location II (). The lower temperatures in cultivation years were, the higher anthocyanin and polyphenolic contents in sweetpotato tubers raised ( and ). It is suggested that the temperature during cultivation has a large influence on the polyphenolic content as well as anthocyanin content in purple sweetpotatoes.
The anthocyanin composition differed between cultivation locations. The peonidin/cyanidin ratio was higher in location II compared with I with an exception of PSL in 2016 (). Peonidin is produced by the methylation of the precursor cyanidin (Holton & Cornish, Citation1995). In sweetpotato cell culture, temperature significantly affected the peonidin/cyanidin ratio. A low temperature (15°C) suppressed accumulation of peonidin-based pigments and a higher temperature (25–30°C) favored methylation (Konczak et al., Citation2005). It is possible that temperature effects methyltransferase activity, which would change peonidin production.
Caffeoylquinic acids and anthocyanins are synthesized via the phenylpropanoid pathway (Vogt, Citation2010). Accumulation of anthocyanins in sweetpotato storage roots is regulated by the MYB-domain-containing transcription factor, IbMYB1 (Mano et al., Citation2007). Overexpression of IbMYB1 led to higher anthocyanin and polyphenolic acid levels in storage roots. In the IbMYB1 transgenic sweetpotato, expression of the genes involved in the phenylpropanoid pathway such as phenyl alanine ammonia lyase (PAL) and cinnamate 4-hydroxylase (C4H), and following anthocyanin pathway including chalcone synthase (CHS), chalcone isomerase (CHI), flavonone-3-hydroxylase (F3H), UDP glucose: flavonoid-3-O-glucosyltransferase (UF3GT), dihydroflavonol reductase (DFR) and anthocyanidin synthase (ANS) were elevated. In addition, levels of proanthocyanidins and total phenolics were also elevated in the IbMYB1 transgenic sweetpotato (Park et al., Citation2015). In strawberry, the expression of MYB and anthocyanin biosynthetic genes, DFR, ANS and UF3GT, decreased significantly at a high temperature (Matsushita et al., Citation2016). Low-temperature stress enhanced the transcripts on general phenylpropanoid (PAL, 4CL) and anthocyanin pathways (CHS, CHI) in maize seedlings (Christie et al., Citation1994). We found higher accumulation of anthocyanins and caffeoylquinic acids in purple sweetpotatoes cultivated in the northern location and in the year with lower temperatures, probably due to elevation of gene expression involved phenylpropanoid pathway followed by anthocyanin and caffeoylquinic acid pathway at low temperatures.
The general phenylpropanoid pathway leads to several branches including caffeoylquinic acids and anthocyanins. In MM, the biochemical pathway of caffeoylquinic acids might be dominant over that of anthocyanins, because the total polyphenolic content was highest among varieties, while the total anthocyanin content was middle level among varieties. ChA is synthesized by hydroxycinnamoyl-CoA:quinate hydroxycinnamoyl transferase (HQT) or hydroxycinnamoyl-CoA:shikimate/quinate hydroxycinnamoyl transferase (HCT) and p-coumarate-3ʹ-hydroxylase (C3ʹH) from p-coumaroyl-CoA (Valiñas et al., Citation2017). The biosynthetic pathway of di-caffeoylquinic acids has not been well studied. In this study, 3,4-diCQA and 4,5-diCQA were more abundant in location II and in 2016 which temperature were high compared with in location I and in 2015. Higher enzyme activity might explain the higher amounts of 3,4- and 4,5-diCQA at location II and in 2016 with higher temperature. Our results will help in the understanding of the relationship between environmental conditions and polyphenolic composition in sweetpotato.
Sugars enhance anthocyanin and chlorogenic acids accumulation and phenylpropanoid-related genes in potato (Payayavula et al., Citation2013). Wang et al. (Citation2016) demonstrated by proteomic approach that starch degradation, which provides abundant substrates for anthocyanin biosynthesis, may contribute to anthocyanin accumulation in purple sweetpotato. The effect of their factors on accumulation of anthocyanin and polyphenolics in sweetpotato at low temperature are future study issue.
In conclusion, the accumulation of anthocyanin and polyphenolics was higher in purple sweetpotatoes cultivated in a northern location (location I), suggesting that lower temperatures enhanced their contents. The varieties used for natural pigment production, such as MM and AKM, are cultivated in southern location (location II). Our study showed that a northern location could be used as an alternative area for purple sweetpotato growth for natural pigment production. Our results also suggest that the amounts of anthocyanin and polyphenolics easily fluctuates and are influenced by climate conditions, probably temperature during cultivation. The cultivation of purple sweetpotato in a northern region could compensate for decrease in pigment production by high temperatures in southern location. In addition, the anthocyanin in sweetpotatoes cultivated in northern region might be more stable in high heat and light because caffeoylquinic acids contribute to anthocyanin stability (Nishiyma et al., Citation2000). The purple sweetpotato varieties used for edible food, such as PSL, are usually cultivated at lower latitudes like location II: a northern location may also be a suitable area for such varieties, because the sugar content is higher (Nakamura et al., Citation2014) and health benefits due to the higher contents of anthocyanin and polyphenolics are expected. For the economical cultivation of purple sweetpotatoes in northern locations, the development of a variety with higher yield in low temperature is desirable, because existing varieties showed relatively lower tuber yield. The breeding for a low-temperature-tolerant sweetopotato is in progress (Kuranouchi et al., Citation2020). Dehydration-responsive element-biding/C-repeat-binding factor (DREB1/CBF) proteins play important roles in cold stress tolerance. Transgenic sweetpotato plants overexpressing IbCBF3 gene had significantly improved tolerance to both cold and drought stress compared with the wild type (Jin et al., Citation2017). The molecular breeding might also be useful for developing of a variety with low temperature tolerance.
Abbreviations
CA: caffeic acid; ChA: chlorogenic acid; 3,4-diCQA: 3,4-di-O-caffeoylquinic acid; 3,5-diCQA: 3,5-di-O-caffeoylquinic acid; 4,5-diCQA: 4,5-di-O-caffeoylquinic acid; MM: Murasakimasari; AKM: Akemurasaki; PSL: Purple Sweet Lord; HPLC: high-performance liquid chromatography; PAL: phenyl alanine ammonia lyase; C4H: cinnamate 4-hydroxylase; CHS: chalcone synthase; CHI: chalcone isomerase; F3H: flavonone-3-hydroxylase; UF3GT: UDP glucose: flavonoid-3-O-glucosyltransferase; DFR: dihydroflavonol reductase; ANS: anthocyanidin synthase; HQT: hydroxycinnamoyl-CoA:quinate hydroxycinnamoyl transferase; HCT: hydroxycinnamoyl-CoA:shikimate/quinate hydroxycinnamoyl transferase; C3ʹH: p-coumarate-3ʹ-hydroxylase (C3ʹH); ANOVA: analysis of variance Comparison of Anthocyanin and Polyphenolics in Purple Sweetpotato (Ipomoea batatas Lam.) Grown in Different Locations in Japan.
Authors Contribution
Koji Ishiguro: Conceived and designed the experiments; performed the experiments; Analyzed and interpreted the data; Contributed reagents, materials, analysis tools or data; Wrote the paper.
Toshikazu Kuranouchi: Conceived and designed the experiments; Contributed materials.
Yumi Kai; Performed the experiments; Contributed materials.
Kenji Katayama; Conceived and designed the experiments; Contributed materials.
PPS2021_003RP-File008.tif
Download TIFF Image (48.2 KB)PPS2021_003RP-File007.tif
Download TIFF Image (42.6 KB)Acknowledgments
This research was supported by grants from the Project of the NARO Bio-oriented Technology Research Advancement Institution (Research program on development of innovative technology). The authors are grateful to Dr. Satoshi Yokota, Dr. Shigenobu Takigawa, Dr. Chie Endo, Dr. Takahiro Noda and Ms. Minako Saito for their assistance.
Disclosure statement
No potential conflict of interest was reported by the authors.
Supplementary material
Supplemental data for this article can be accessed here.
Additional information
Funding
References
- Christie, P. J., Alfenito, M. R., & Walbot, V. (1994). Impact of low-temperature stress on general phenylpropanoid and anthocyanin pathways: Enhancement of transcript abundance and anthocyanin pigmentation in maize seedlings. Planta, 194(4), 541–549. https://doi.org/https://doi.org/10.1007/BF00714468
- Harrison, H., Peterson, J., Clark, C., & Snook, M. (2001). Sweetpotato periderm components inhibit in vitro growth of root rotting fungi. Hortscience, 36(5), 927–930. https://doi.org/https://doi.org/10.21273/HORTSCI.36.5.927
- Harrison, H., Peterson, J., & Snook, M. E. (2006). Simulated drought induces high caffeic acid contents in storage root periderm of greenhouse grown sweetpotatoes. Hortscience, 41(1), 277–278. https://doi.org/https://doi.org/10.21273/HORTSCI.41.1.277
- Holton, T., & Cornish, E. (1995). Genetics and biochemistry of anthocyanin biosyntesis. The Plant Cell, 7(7), 1071–1083. https://doi.org/https://doi.org/10.1105/tpc.7.7.1071
- Ishiguro, K., Yahara, S., & Yoshimoto, M. (2007). Changes in polyphenolic content and radical-scavenging activity of sweetpotato (Ipomoea batatas L.) during storage at optimal and low temperatures. Journal of Agricultural and Food Chemistry, 55(26), 10773–10778. https://doi.org/https://doi.org/10.1021/jf072256v
- Jin, R., Kim, B. H., Ji, C. Y., Kim, H. S., Li, H. M., Ma, D. F., & Kwak, S. S. (2017). Overexpressing IbCBF3 increases low temperature and drought stress tolerance in transgenic sweetpotato. Plant Physiology and Biochemistry, 118, 45–54. https://doi.org/https://doi.org/10.1016/j.plaphy.2017.06.002
- Kano, N., Takayanagi, T., Harada, K., Makino, K., & Ishikawa, F. (2005). Antioxidative activity of anthocyanins from purple sweet potato, Ipomoea batatas cultivar Ayamurasaki. Bioscience, Biotechnology, and Biochemistry, 69(5), 979–988. https://doi.org/https://doi.org/10.1271/bbb.69.979
- Kobayashi, T., Ikoma, H., & Mochida, H. (1998). Effect of cultural conditions on anthocyanin content of purple-colored sweetpotato. Sweetpotato Research Front, 6, 2. https://www.naro.go.jp/publicity_report/publication/archive/files/SPORF06.pdf
- Konczak, I., Terahara, N., Yoshimoto, M., Nakatani, M., Yoshinaga, M., & Yamakawa, O. (2005). Regulating the composition of anthocyanins and phenolic acids in a sweetpotato cell culture towards production of polyphenolic complex with enhanced physiological activity. Trends in Food Science & Technology, 16(9), 377–388. https://doi.org/https://doi.org/10.1016/j.tifs.2005.02.007
- Kuranouchi, T., Nishinaka, M., Takada, A., Kumagai, T., Nakatani, M., & Katayama, K. (2020). Recent progress in breeding low-temperature-tolerant sweetpotato lines in Japan. Japan Agricultural Research Quarterly: JARQ, 54(1), 1–6. https://doi.org/https://doi.org/10.6090/jarq.54.1
- Kurata, R., Adachi, M., Yamakawa, O., & Yoshimoto, M. (2007). Growth suppression of human cancer cells by polyphenolics from sweetpotato (Ipomoea batatas L.) leaves. Journal of Agricultural and Food Chemistry, 55(1), 185–190. https://doi.org/https://doi.org/10.1021/jf0620259
- Mahmood, N., Moore, P., Detommasi, N., Desimone, F., Colman, S., Hay, A., & Pizza, C. (1993). Inhibition of HIV-infection by caffeoylquinic acid-derivatives. Antiviral Chemistry & Chemotherapy, 4(4), 235–240. https://doi.org/https://doi.org/10.1177/095632029300400406
- Mano, H., Ogasawara, F., Sato, K., Higo, H., & Minobe, Y. (2007). Isolation of a regulatory gene of anthocyanin biosynthesis in tuberous roots of purple-fleshed sweet potato. Plant Physiology, 143(3), 1252–1268. https://doi.org/https://doi.org/10.1104/pp.106.094425
- Matsui, T., Ebuchi, S., Fujise, T., Abesundara, K., Doi, S., Yamada, H., & Matsumoto, K. (2004). Strong antihyperglycemic effects of water-soluble fraction of Brazilian propolis and its bioactive constituent, 3,4,5-tri-o-caffeoylquinic acid. Biological & Pharmaceutical Bulletin, 27(11), 1797–1803. https://doi.org/https://doi.org/10.1248/bpb.27.1797
- Matsui, T., Ebuchi, S., Kobayashi, M., Fukui, K., Sugita, K., Terahara, N., & Matsumoto, K. (2002). Anti-hyperglycemic effect of diacylated anthocyanin derived from Ipomoea batatas cultivar Ayamurasaki can be achieved through the alpha-glucosidase inhibitory action. Journal of Agricultural and Food Chemistry, 50(25), 7244–7248. https://doi.org/https://doi.org/10.1021/jf025913m
- Matsushita, K., Sakayori, T., & Ikeda, T. (2016). The effect of high air temperature on anthocyanin concentration and the expressions of its biosynthetic genes in strawberry‚ ‘Sachinoka’. Environmental Control in Biology, 54(2), 101–107. https://doi.org/https://doi.org/10.2525/ecb.54.101
- Miyazaki, K., Makino, K., Iwadate, E., Deguchi, Y., & Ishikawa, F. (2008). Anthocyanins from purple sweet potato Ipomoea batatas cultivar ayamurasaki suppress the development of atherosclerotic lesions and both enhancements of oxidative stress and soluble vascular cell adhesion molecule-1 in apolipoprotein E-deficient mice. Journal of Agricultural and Food Chemistry, 56(23), 11485–11492. https://doi.org/https://doi.org/10.1021/jf801876n
- Montilla, E. C., Hillebrand, S., & Winterhalter, P. (2011). Anthocyanins in purple sweet potato (Ipomoea batatas L.) varieties. Fruit, Vegetable and Cereal Science and Biotechnology, 5(2), 19–24. https://www.researchgate.net/publication/283429468_Anthocyanins_in_purple_sweet_potato_Ipomoea_batatas_L_Varieties
- Nakamura, Y., Kuranouchi, T., Ohara-Takada, A., & Katayama, K. (2014). The effects of beta-amylase activity and starch pasting temperature on maltose generation in steamed storage roots of sweet potato. Journal of Japan Society of Nutrition and Food Science, 61 (12), 577–585. (in Japanese with English summary). https://doi.org/https://doi.org/10.3136/nskkk.61.577
- Nishiyma, K., Ichi, T., & Kawamoto, M. (2000). Japanese patent (P2000-270807A). (in Japanese).
- Oki, T., Sawai, Y., Sato-Furukawa, M., & Suda, I. (2011). Validation of pH differential method for the determination of total anthocyanin content in black rice and black soybean with interlaboratory comparison. Bunseki Kagaku, 60 (10), 819–824. (in Japanese with English summary). https://doi.org/https://doi.org/10.2116/bunsekikagaku.60.819
- Paneque Ramirez, G. (2017). Cultivation harvesting and storage of sweet potato products. http://www.fao.org/docrep/003/T0554E/T0554E14.htm.
- Park, S. C., Kim, Y. H., Kim, S. H., Jeong, Y. J., Kim, C. Y., Lee, J. S., Bae, J. Y., Ahn, M. J., Jeong, J. C., Lee, H. S., & Kwak, S. S. (2015). Overexpression of the IbMYB1 gene in an orange-fleshed sweet potato cultivar produces a dual-pigmented transgenic sweet potato with improved antioxidant activity. Physiologia Plantarum, 153(4), 525–537. https://doi.org/https://doi.org/10.1111/ppl.12281
- Payayavula, R. S., Singh, R. K., & Navarre, D. A. (2013). Transcription factors, sucrose, and sucrose metabolic genes interact to regulate potato phenylpropanoid metabolism. Journal of Experimental Botany, 64(16), 5115–5131. https://doi.org/https://doi.org/10.1093/jxb/ert303
- Peluso, G., Defeo, V., Desimone, F., Bresciano, E., & Vuotto, M. (1995). Studies on the inhibitory effects of caffeoylquinic acids on monocyte migration and superoxide ion production. Journal of Natural Products, 58(5), 639–646. https://doi.org/https://doi.org/10.1021/np50119a001
- Peterson, J., Harrison, H., Snook, M., & Jackson, D. (2005). Chlorogenic acid content in sweetpotato germplasm: Possible role in disease and pest resistance. Allelopathy Journal, 16(2), 239–249. https://www.ars.usda.gov/research/publications/publication/?seqNo115=144515
- Reyes, L., Villarreal, J., & Cisneros-Zevallos, L. (2007). The increase in antioxidant capacity after wounding depends on the type of fruit or vegetable tissue. Food Chemistry, 101(3), 1254–1262. https://doi.org/https://doi.org/10.1016/j.foodchem.2006.03.032
- Schwartz, E., Tzulker, R., Glazer, I., Bar-Ya’akov, I., Wiesman, Z., Tripler, E., Bar-Ilan, I., Fromm, H., Borochov-Neori, H., Holland, D., & Amir, R. (2009). Environmental conditions affect the color, taste, and antioxidant capacity of 11 pomegranate accessions’ fruits. Journal of Agricultural and Food Chemistry, 57(19), 9197–9209. https://doi.org/https://doi.org/10.1021/jf901466c
- Shimozono, H., Kobori, M., Shinmoto, H., & Tsushida, T. (1996). Suppression of the melanogenesis of mouse melanoma B 16 cells by sweet potato extract. Nippon Shokuhin Kagaku Kogaku Kaishi, 43 (3), 313–317. (in Japanese with English summary). https://doi.org/https://doi.org/10.3136/nskkk.43.313
- Suda, I., Oki, T., Masuda, M., Kobayashi, M., Nishiba, Y., & Furuta, S. (2003). Physiological functionality of purple-fleshed sweet potatoes containing anthocyanins and their utilization in foods. Japan Agricultural Research Quarterly: JARQ, 37(3), 167–173. https://doi.org/https://doi.org/10.6090/jarq.37.167
- Takos, A. M., Jaffé, F. W., Jacob, S. R., Bogs, J., Robinson, S. P., & Walker, A. R. (2006). Light-induced expression of a MYB gene regulates anthocyanin biosynthesis in red apples. Plant Physiology, 142(3), 1216–1232. https://doi.org/https://doi.org/10.1104/pp.106.088104
- Tanaka, M., Ishiguro, K., Oki, T., & Okuno, S. (2017). Functional components in sweetpotato and their genetic improvement. Breeding Science, 67(1), 52–61. https://doi.org/https://doi.org/10.1270/jsbbs.16125
- Terahara, N., Oki, T., Matsui, T., Fukui, K., Sugita, K., Matsumoto, K., & Suda, I. (2007). Simultaneous determination of major anthocyanins in purple sweet potato. Nippon Shokuhin Kagaku Kogaku Kaishi, 54 (1), 33–38. (in Japanese with English summary). https://doi.org/https://doi.org/10.3136/nskkk.54.33
- Terahara, N., Shimizu, T., Kato, Y., Nakamura, M., Maitani, T., Yamaguchi, M., & Goda, Y. (1999). Six diacylated anthocyanins from the roots of purple sweet potato, Ipomoea batatas. Bioscience, Biotechnology, and Biochemistry, 63(8), 1420–1424. https://doi.org/https://doi.org/10.1271/bbb.63.1420
- Uritani, I. (1999). Biochemistry on postharvest metabolism and deterioration of some tropical tuberous crops. Botanical Bulletin of Academia Sinica, 40(3), 177–183. https://ejournal.sinica.edu.tw/bbas/content/1999/3/bot403-01.html
- Valiñas, M. A., Lanteri, M. L., Have, A. T., & Andreu, A. B. (2017). Chlorogenic acid, anthocyanin and flavan-3-ol biosynthesis in flesh and skin of Andean potato tubers (Solanum tuberosum subsp. Andigena). Food Chemistry, 229, 837–846. https://doi.org/https://doi.org/10.1016/j.foodchem.2017.02.150
- Villavicencio, L., Blankenship, S., Yencho, G., Thomas, J., & Raper, C. (2007). Temperature effect on skin adhesion, cell wall enzyme activity, lignin content, anthocyanins, growth parameters, and periderm histochemistry of sweetpotato. Journal of the American Society for Horticultural Science, 132(5), 729–738. https://doi.org/https://doi.org/10.21273/JASHS.132.5.729
- Vogt, T. (2010). Phenylpropanoid biosynthesis. Molecular Plant, 3(1), 2–20. https://doi.org/https://doi.org/10.1093/mp/ssp106
- Wang, S., Pan, D., Lv, X., Song, X., Qiu, Z., Huang, C., Huang, R., & Chen, W. (2016). Proteomic approach reveals that starch degradation contributes to anthocyanin accumulation in tuberous root of purple sweet potato. Journal of Proteomics, 143, 298–305. https://doi.org/https://doi.org/10.1016/j.jprot.2016.03.010
- Yoshimoto, M., Kurata, R., Okuno, S., Ishiguro, K., Yamakawa, O., Tsubata, M., Mori, S., & Takagaki, K. (2006). Nutritional value and physiological functions of sweetpotato leaves. Acta Horticulturae, 703(703), 107–116. https://doi.org/https://doi.org/10.17660/ActaHortic.2006.703.11
- Yoshimoto, M., Okuno, S., Yamaguchi, M., & Yamakawa, O. (2001). Antimutagenicity of deacylated anthocyanins in purple-fleshed sweetpotato. Bioscience, Biotechnology, and Biochemistry, 65(7), 1652–1655. https://doi.org/https://doi.org/10.1271/bbb.65.1652
- Yoshimoto, M., Yahara, S., Okuno, S., Islam, M., Ishiguro, K., & Yamakawa, O. (2002). Antimutagenicity of mono-, di-, and tricaffeoylquinic acid derivatives isolated from sweetpotato (Ipomoea batatas L.) leaf. Bioscience, Biotechnology, and Biochemistry, 66(11), 2336–2341. https://doi.org/https://doi.org/10.1271/bbb.66.2336
- Yoshinaga, M. (1995). New cultivar “Ayamurasaki” for colorant production. Sweetpotato Research Front, 1, 2. https://www.naro.go.jp/publicity_report/publication/archive/files/SPORF01_1.pdf