
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
Supraliminal central cue words elicit spatial attention shifts to the cued side. To investigate, if this also holds true for subliminal central words, masked cue/prime words, left or right, were shown prior to a visual colour search display. Both response times to targets and event-related brain potentials (ERPs) reflecting spatial attention (EDAN, ADAN, LDAP, N2pc) were analyzed. In Experiment 1, we found no evidence for spatial attention shifts by subliminal central words; neither significant ERPs (EDAN, ADAN, LDAP) nor significant validity effects in the behavioural data. To control for the processing of the subliminal words, we included trials with a congruent or incongruent target word (left or right) in a classic target discrimination task in Experiment 2, and additionally analyzed an ERP reflecting semantic congruence, the N400. We mainly replicated the results from Experiment 1 (no spatial attention shifts by subliminal words) and found a difference depending on the congruence between subliminal word and target word (N400) as well as a negative congruence effect in the reaction times. These findings demonstrate that the subliminal word was processed. We therefore conclude that subliminally presented central cue words – even though they are processed – cannot elicit spatial attention shifts in visual colour search.
KEYWORDS:
Many studies have shown that humans can process subliminally presented stimuli of which they are unaware (Greenwald, Draine, & Abrams, Citation1996; Marcel, Citation1983). However, it is still debated if there are differences between processing of subliminal and supraliminal stimuli and what these differences might be (e.g., Ansorge, Kunde, & Kiefer, Citation2014; Dehaene, Changeux, Naccache, Sackur, & Sergent, Citation2006; Hassin, Citation2013).
Here, we tested if subliminal words can trigger an attention shift via their spatial meaning. Many masked-priming studies have shown that the meaning of subliminal words can be processed (Carr & Dagenbach, Citation1990; Kiefer, Citation2002). For example, a visually masked and therefore subliminally presented prime word can create a congruence effect: faster responses where meanings of prime word and target word are semantically related (or congruent) than unrelated (or incongruent; Kiefer, Citation2002). In addition, at least supraliminally presented words (i.e., words of which observers are aware) can elicit an attention shift corresponding to their meaning (Gibson & Kingstone, Citation2006; Hommel, Pratt, Colzato, & Godijn, Citation2001). For example, searching for and responding to a visual target on the same side as indicated by the task-irrelevant meaning of a preceding centrally presented cue word (e.g., a target on the left following the centrally presented cue word left) is faster than searches for a target on the opposite side (e.g., a target on the right following the same cue word left), even if across trials cue words are not predictive of the most likely target position (Hommel et al., Citation2001). Apart from behavioural cueing effects, Brignani, Guzzon, Marzi, and Miniussi (Citation2009) found evidence for spatial attention shifts by endogenous cues (patterns with arbitrarily assigned directions) in electrophysiological data. Our subliminal direction words resemble the pattern cues of Brignani et al. (Citation2009), as they both carry their direction information endogenously via their meaning.
Although a wide range of research has investigated on the one hand the processing of subliminal words and on the other hand attention shifts elicited by central word cues, the combination of both, namely the question if subliminal word cues can elicit spatial attention shifts, has not been answered. Yet, this is important to test in order to understand if subliminal semantic processing also elicits associated attentional processes, much as this would be the case for supraliminal processing. In other words, our study will help to understand if there are limits to consciousness-independent processing or if consciousness-independent processing follows the same laws as any other form of processing accompanied by consciousness does.
The suspicion that subliminal words might elicit an attention shift was nurtured by a recent study of Ansorge, Khalid, and Laback (Citation2016). These authors presented masked direction words (e.g., above or below) at screen centre prior to an auditory target from above or below and found that responses to the targets were faster where prime-word meaning and target position were congruent than where they were incongruent (Ansorge et al., Citation2016). However, it remained unclear if the spatial words elicited an attention shift, such that attention was already at target position at the time of target onset in congruent but not in incongruent conditions, or if the words affected tone processing via spatial semantics, for example, allowing a faster processing of target location information under congruent than incongruent conditions. Prior research has shown that masked stimuli, such as shapes, have an influence on spatial attention (e.g., Mattler, Citation2003; Schmidt & Schmidt, Citation2010). However, is this also the case for subliminal words?
In the current study, to shed a light on the underlying mechanisms in the processing of subliminally presented direction words, we took a maximally sensitive and exhaustive approach, and recorded a variety of potential electrophysiological indices of attention by the respective event-related potentials (ERPs). Across two experiments, we also took different approaches, such as presenting subliminal cues within or without the context of supraliminal words that were predictive of the target locations (Experiment 1 vs. Experiment 2), to see if at least in the former conditions, subliminal cues may trigger a spatial attention shift.
Event-related potentials reflecting spatial attention
To examine if masked (subliminal) words can elicit spatial attention shifts, we used four different lateralized event-related potentials (ERPs). Most straightforwardly, three components of the lateralized ERPs: The anterior directing-attention negativity (ADAN), a robust component concerning attentional shifts within the coordinates of external space (Eimer, Forster, Fieger, & Harbich, Citation2004), the early directing-attention negativity (EDAN; e.g., Feng & Zhang, Citation2014; Hopf & Mangun, Citation2000), and the late directing-attention positivity (LDAP; e.g., Kiss, van Velzen, & Eimer, Citation2008). Those ERPs are stronger contralateral to the direction of the induced attention shift and appear already in the interval between a centrally presented cue stimulus and a peripherally presented target (e.g., Harter, Miller, Price, LaLonde, & Keyes, Citation1989; Jongen, Smulders, & van der Heiden, Citation2007; Kelly, Gomez-Ramirez, & Foxe, Citation2009). These electrophysiological measures thus provide a window into word-elicited processing even prior to the overt response. They thus also give us a more exhaustive picture of the processing than the overt responses alone that sometimes are not optimally sensitive (e.g., where subliminal influences decay across time).
A fourth well-known lateralized ERP reflecting covert spatial attention shifts is the N2pcFootnote1 (posterior contralateral negativity) starting around 200 ms following the onset of a peripheral stimulus (see, e.g., Ansorge, Kiss, & Eimer, Citation2009; Livingstone, Christie, Wright, & McDonald, Citation2017; Luck & Hillyard, Citation1994). A centrally presented cue/prime word cannot directly elicit an N2pc. However, if prior to a visual search display with a target on the left or on the right the central cue/prime word left or right is presented, the N2pc to the target might differ depending on the congruence (or better: validity, see next sentence) between the meaning of the cue/prime word and the location of the target. For the sake of not confusing this attentional influence of the cue/prime word with its semantic processing in general, in the following, we use the labels valid instead of congruent and invalid instead of incongruent to refer to the orthogonal manipulation of cue/prime word meaning and target side in the visual search task.
To clarify if subliminally presented words indeed elicit an attention shift, we recorded electroencephalographic (EEG) data and looked for specific ERPs reflecting spatial attention shifts. We masked and, thus, presented subliminally, a central cue/prime word (left; right) between a forward and backward mask following the protocol of Kiefer and Brendel (Citation2006; see also Ansorge, Khalid, & König, Citation2013). If the cue/prime word elicits an attention shift, lateralized ERPs should occur depending on the meaning of the cue/prime word.
In addition, we used a visual search task for targets on one side presented among distractors on the other side. Here, cueing effects should occur in the reaction times to the target (Posner, Snyder, & Davidson, Citation1980): If the cue/prime words trigger an attention shift, it should be easier for the participants to search for and respond to the targets in valid conditions than in invalid conditions. In the visual search task, an attention shift elicited by the masked cue/prime words might also show up as a modulation of the target-elicited N2pc in valid compared to invalid trials.
Finally, to test if the masked cue/prime words were indeed presented subliminally, in a second block, the participants had to classify the cue/prime word as being the word left or right. In this task, we expected chance performance if the participants were unaware of the cue/prime words (see, e.g., Kiefer, Citation2002).
Experiment 1
Method
Participants
Fourteen right-handed psychology students from the University of Vienna participated in Experiment 1 (MAge = 22.64 years, SDAge = 2.87 years). The students received course credit for their participation. Prior to the testing, they signed an informed consent form that included questions regarding their aptitude for the EEG-testing. Their vision and colour vision was normal or corrected to normal. The treatment of the participants was in line with established ethical standards and an approval from the Ethics Committee of the University of Vienna was obtained.
Apparatus and stimuli
The visual stimuli were presented on a 19″ VGA-monitor with an aspect ratio of 4:3, a resolution of 1,024 × 786 pixels, and a refresh rate of 59 Hz. For the visual search task, coloured numbers were used (height: 1.5° of visual angle). The colours of the numbers were isoluminant: green (CIE L*a*b*, 56.0/−62.6/50.2), blue (56.1/26.7/56.5), and red (55.9/71.3/56.5). The cue words and letters of the forward and backward masks were presented centrally and written in bold Courier New 12 pt, in black font colour (30.2/−6.6/−13.0). The background was grey (70.1/−5.4/−26.4) throughout the whole experiment. Participants sat comfortably in front of the monitor, the distance between their eyes and the screen was kept constant at 58 cm by a chinrest. The room was dimly lit and soundproof. To ensure the possibility to contact the experimenter at all times, a live webcam was installed. No footage was recorded. All responses were given by keypress on a standard keyboard. The experiment was programmed and conducted using E-Prime 2.0 (Psychology Software Tools, Pittsburgh, PA, Citation2013).
Electrophysiological recording
The EEG was recorded using 64 active electrodes (Brain Products, actiCAP system) in a 10/10 system cap (EASYCAP GmbH, Herrsching, Germany) at a sampling rate of 1000 Hz (full-band DC-EEG system, neuroConn GmbH, Ilmenau, Germany). The online reference during recording was the ground electrode (AFZ), offline re-referencing was calculated by the average of both mastoids. We applied a 40 Hz low-pass finite impulse response filter: cut-off 45 Hz (−6 dB), transition bandwidth 10 Hz. The horizontal electrooculogram (HEOG) was calculated using the difference between the two electrodes positioned at the outer canthi. Trials with wrong or too slow responses were removed, as well as trials with very low activity (less than 0.5 µV difference between subsequent samples within a time period of 500 ms), very high signal changes (more than 50 µV/ms), values exceeding 80 µV, horizontal eye movements (HEOG exceeding ± 30 µV), and vertical eye movements or blinks (Fp1/Fp2 ± 60 µV). All EEG data were processed in MATLAB (The MathWorks, Inc., Citation2014) using the EEGLAB toolbox (Delorme & Makeig, Citation2004) with the ERPLAB extensions (Lopez-Calderon & Luck, Citation2014).
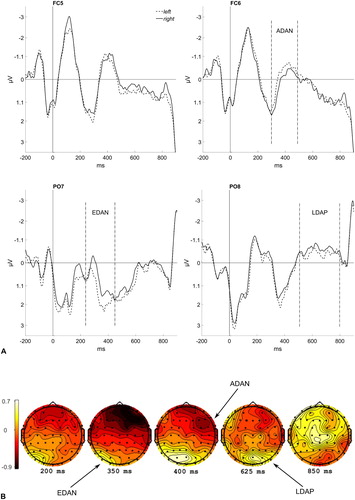
For ERP analyses, mean amplitudes of activities between two fixed latencies were extracted and analyzed in R (R Core Team, Citation2017) using the packages apa (Gromer, Citation2017) and ez (Lawrence, Citation2016). The respective electrode sites and time windows were broadly chosen after the literature and refined using visual inspection of scalp distributions and waveforms. As our study is sufficiently novel, no comparable prior studies can be used to define the time windows solely beforehand, as the timing of ERPs can change in relation to, for example, the luminance/shape/discriminability of stimuli. In this case, it is often the best approach to look at collapsed waveforms to refine the time windows, which are later used to analyze non-collapsed data (collapsed-localizers method; Luck & Gaspelin, Citation2017). The N2pc occurred between 200 and 300 ms after visual search target onset at the electrode sites PO7 and PO8 (e.g., Jolicœur, Brisson, & Robitaille, Citation2008). The EDAN occurred between 250 and 450 ms, the LDAP 500–800 ms after cue/prime word onset (both at the electrode sites PO7 and PO8), and the ADAN could be found between 300 and 500 ms after cue/prime onset, at the frontal electrode sites FC5 and FC6 (cf. Holmes, Mogg, Garcia, & Bradley, Citation2010; Jongen et al., Citation2007; Kelly et al., Citation2009; Talsma, Slagter, Nieuwenhuis, Hage, & Kok, Citation2005).
Task and design
The experiment consisted of two different tasks: visual search (experimental task of main interest; Blocks 1 and 2) and cue discrimination (control task for cue visibility; Blocks 3 and 4). In the visual search task, one number was presented to the left and one to the right of the fixation cross. The task was to react to the number presented in a to-be-searched for predefined target colour (e.g., in green) and report its identity (1, 2, 3, 4) by keypress (keys #F, #G, #H, and #J). As masked central cues produce more reliable cueing effects when combined with 100% valid unmasked cues (Al-Janabi & Finkbeiner, Citation2012; Reuss, Kiesel, Kunde, & Wühr, Citation2012), we also included trials in which the distractor had the same colour as the target. In these trials, the relevance of the cue word was increased, as colour search was not enough to find the target, but the cue word presented before the target denoted the location of the target with 100% certainty and had to be used to find the target. Participants knew this and were encouraged to use the cue word for this purpose. In the cue discrimination task, participants tried to discriminate the identity of the supraliminally or subliminally presented cue word (left or right) by keypress without time constraints. To ensure that the cue word was not consciously perceived in the subliminal condition, it was sandwiched between a forward- and backward-mask, both consisting of ten randomly drawn nonrecurring uppercase letters (see Kiefer & Brendel, Citation2006). This task was implemented as an additional task to the visual search task in the second half of the experiment; hence, in the second half of the experiment, every trial consisted of two consecutive tasks: visual search plus cue discrimination. Two different inter-stimulus-intervals (ISIs) between cue and target were implemented blockwise for both the first half of the experiment (visual search task only, Blocks 1 and 2) and the second half of the experiment (visual search plus cue discrimination, Blocks 3 and 4). The block order of the first two blocks and of the second two blocks as well as the target colour for the visual search task were counterbalanced across participants. The target (e.g., green number) appeared randomly left or right from the fixation cross, and the distractor was coloured randomly in one of two further colours (e.g., blue or red) in colour target trials, and had the same colour as the target in location targets. Trials with masked or unmasked cue words appeared equally often and in random order. The masked cue word was non-predictive (50% valid), and always followed by a single colour target. The unmasked cue word was 100% valid, and the two different target types (a single colour target, or two numbers of the same colour [location target]) followed equally often and in random order.
Procedure
At first, participants performed 20 practice trials, randomly drawn from Blocks 1 and 2, until they achieved a response accuracy of at least 80%. During practice, feedback for correct, incorrect, and slow responses (longer than 2 s) was given. Then, participants proceeded with Blocks 1 and 2, consisting of 240 trials each. Before Blocks 3 and 4 (240 trials each), an instruction for the additional cue discrimination task was given. The experiment consisted of 960 trials in total. After every 96 trials, participants were advised (by a corresponding display and a momentary halt of the experiment that only the participant could end by appropriate button press) to take a short break. Including time for preparation, testing, breaks, removing of electrodes, and debriefing, every participant stayed in the laboratory between 2 and 3 h.
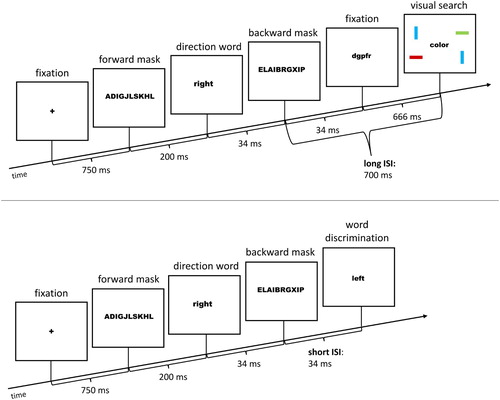
shows the structure of an exemplary trial of Block 1 or 2. Each trial started with the presentation of a fixation cross for 750 ms. Then the cue word was shown for 34 ms. In masked trials, it was rendered invisible between a forward mask (200 ms) and a backward mask (34 ms). The ISI between cue offset and target onset was either 34 ms or 700 ms. The shorter ISI might be necessary to demonstrate the validity effect in the visual search task (e.g., Berger, Citation1999; Castel, Chasteen, Scialfa, & Pratt, Citation2003; Reuss, Pohl, Kiesel, & Kunde, Citation2011; Schoeberl, Fuchs, Theeuwes, & Ansorge, Citation2015; Tassinari, Aglioti, Chelazzi, Peru, & Berlucchi, Citation1994) but the longer ISI was necessary for the registration of the cue-elicited ERP components free of visual searches’ target-elicited activity. The target display was shown for 150 ms. Until reaction, the screen was blank.
Figure 1. Exemplary trial (from left to right): The fixation cross, optional forward mask (in subliminal trials only), cue (direction word: right), optional backward mask (in subliminal trials only), and the target display are shown. The target is a colour target (green number at the right position). The preceding word right validly cued its location. Stimuli are not drawn to scale. The arrow depicts the flow of time. ISI: Inter-Stimulus Interval.
If subliminally presented central cue words can elicit spatial attention shifts, we would expect to find respective ERP components by masked cues: ADAN (more negative waveforms for the word left than right over the right hemisphere, FC6; more negative waveforms for the word right than left over the left hemisphere, FC5), EDAN (more negative waveforms for the word left than right over the right hemisphere, PO8; more negative waveforms for the word right than left over the left hemisphere, PO7), and LDAP (more positive waveforms for the word left than right over the right hemisphere, PO8; more positive waveforms for the word right than left over the left hemisphere, PO7). In addition, the size of the N2pc component might differ between validly and invalidly cued colour targets. If subliminally presented central cue words cannot elicit spatial attention shifts, we would expect to find N2pc components for colour targets independent from the validity of the preceding cue, and no ERPs reflecting spatial attention shifts (ADAN, EDAN, LDAP) for masked cues.
Results
Electroencephalographic data
For the cue-logged analyses (LDAP, ADAN, and EDAN), only trials with the long ISI were used, as the short ISI of 34 ms is not long enough to see any of those ERPs (arising between 250 and 800 ms post cue onset). In addition, the variable Validity does not play a role in the cue-elicited ERPs, as its steps are only relevant and become clear to the participants after target onset (734 ms post cue onset).
Late directing-attention positivity
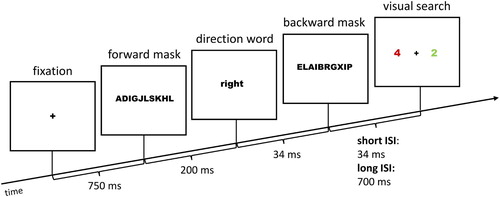
The late directing-attention positivity (LDAP) is an ERP component reflecting spatial attention shifts. It is a greater positivity at about 500–800 ms post-cue, contralateral to the side, the attention is shifted to. It is usually located at the posterior electrodes PO7 and PO8. If the subliminal cue word can elicit a spatial attention shift to the respective side, the amplitude at PO7 (left hemisphere) should be more positive after the word right than left, and respectively for PO8 (right hemisphere) more positive for the word left than right. We conducted two analyses of variance (ANOVAs; 2 × 2) of the long ISI conditions, with the variables Hemisphere (contralateral; ipsilateral) and Cue Word (left; right), separately for masked (subliminal) and unmasked (supraliminal) cues (see ). For masked cues, we found a significant main effect for Hemisphere (PO7: 0.41 µV, PO8: −0.64 µV), F(1, 13) = 14.64, p = .002, = .53. For unmasked cues, both the main effect for Hemisphere (PO7: 1.29 µV, PO8: −0.32 µV), F(1, 13) = 26.64, p < .001,
= .67, and the main effect for Cue Word (left: 0.78 µV, right: 0.19 µV), F(1, 13) = 6.04, p = .029,
= .32, were significant. The main effect for Cue Word (word right more negative than word left) corresponds to a significant LDAP over the right hemisphere (PO8).
Figure 2. (A) Event-related potentials (ERPs) elicited by masked cues at electrode sites PO7 (left panel) and PO8 (right panel), depending on the identity of the cue word (dashed line: left, solid line: right). (B) ERPs elicited by unmasked cues at electrode sites PO7 (left panel) and PO8 (right panel), depending on the identity of the cue word (dashed line: left, solid line: right). For unmasked cues, the early directing-attention negativity (EDAN; not significant) can be seen between 250 and 450 ms (PO7) and the significant late directing-attention positivity (LDAP) between 500 ms and 800 ms (PO8).
Early directing-attention negativity
The early directing-attention negativity (EDAN) appears at the electrodes PO7 and PO8, but early following cue word onset in a time window between 250 and 450 ms. The amplitude at PO7 (left hemisphere) is expected to be more negative for the word right than left, respectively at PO8 (right hemisphere) more negative for the word left than right (see ). Two ANOVAs (2 × 2) of the long ISI conditions, with the variables Hemisphere (contralateral; ipsilateral) and Cue Word (left; right) were conducted for masked (subliminal) and unmasked (supraliminal) cues. For masked cues, the main effect for Hemisphere was significant (PO7: 0.68 µV, PO8: −0.71 µV), F(1, 13) = 11.09, p = .005, = .46. For unmasked cues, no effects were significant, all Fs(1, 13) < 1.48, all ps > .246, all
s < .10.
Anterior directing-attention negativity
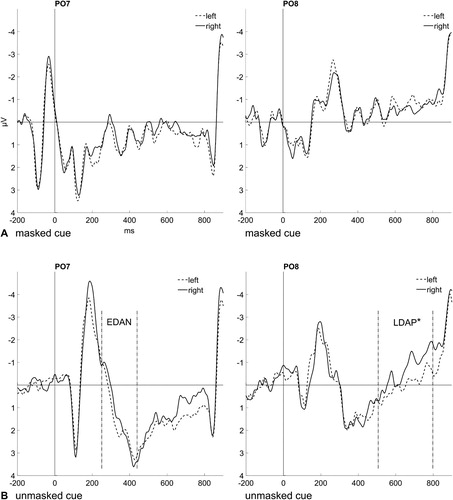
The anterior directing-attention negativity (ADAN) is a greater negativity contralateral to the side the attention is shifted to. It is usually strongest at the fronto-central electrodes FC5 and FC6 in the time window between 300 and 500 ms. Provided that the subliminal cue word can elicit an attention shift, the amplitude at FC5 (left hemisphere) is expected to be more negative for the word right than left, respectively at FC6 (right hemisphere) more negative for the word left than right (see ). Two repeated-measures ANOVAs (2 × 2), with mean amplitude values in the respective time window (300 ms to 500 ms) of the long ISI condition for the variables Hemisphere (contralateral; ipsilateral) and Cue Word (left; right) were conducted, one for masked and one for unmasked cues. There were no significant effects, neither for masked cues, all Fs(1, 13) < 1.12, all ps > .310, all s < .08, nor for unmasked cues, all Fs(1, 13) < 2.82, all ps > .117, all
s < .18.
Figure 3. (A) Event-related potentials (ERPs) elicited by masked cues at electrode sites FC5 (left panel) and FC6 (right panel), depending on the identity of the cue word (dashed line: left, solid line: right). (B) ERPs elicited by unmasked cues at electrode sites FC5 (left panel) and FC6 (right panel), depending on the identity of the cue word (dashed line: left, solid line: right). The anterior directing-attention negativity (ADAN; not significant) can be seen between 300 and 500 ms for unmasked cues only (FC5).
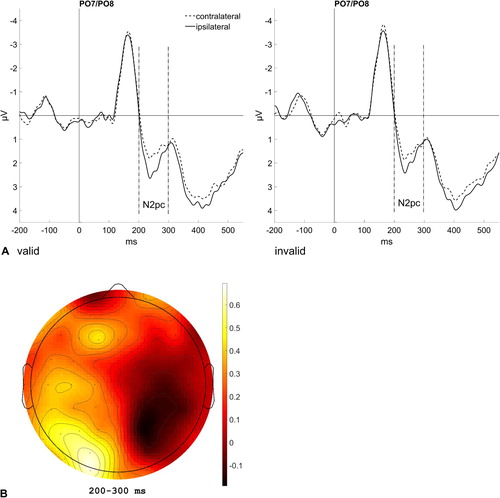
N2 posterior contralateral
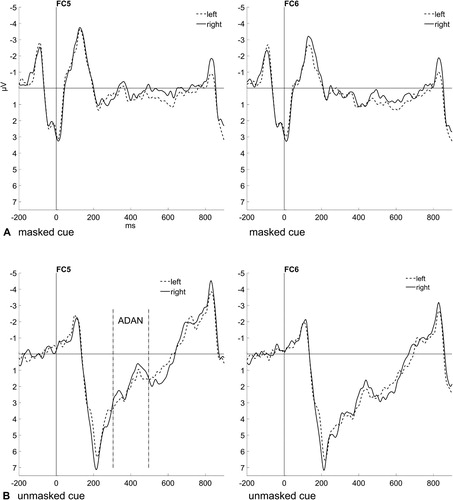
The N2pc is an ERP component reflecting spatial attention shifts. Its maximum is located at posterior electrode sites (PO7/PO8), and it shows as a greater negativity contralateral to the side of an attended stimulus between 200 and 300 ms post target (e.g., Jolicœur et al., Citation2008). The amplitude at PO7 (left hemisphere) should be more negative for a target presented at the right side of the screen than at the left side, and respectively for PO8 (right hemisphere) more negative for a target on the left side of the screen than on the right side. Additionally, if the subliminally presented cue word can elicit a spatial attention shift, the strength of the N2pc might differ depending on the validity or position relation between cue word and the side of the screen where the target is shown (see ). For targets following masked cue words, a repeated-measures ANOVA (2 × 2), with the variables Validity (cue word validly cued the target side; cue word invalidly cued the target side) and Hemisphere (contralateral; ipsilateral) was conducted. It yielded a significant main effect for Hemisphere (contralateral: 0.23 µV; ipsilateral: 1.06 µV), F(1, 13) = 45.65.23, p < .001, = .78, which is evidence for a significant N2pc, independent of the validity of the preceding masked cue. For targets following unmasked cue words, we computed the same repeated-measures ANOVA (2 × 2). It showed a significant interaction between Target Type and Hemisphere, F(1, 13) = 11.25, p = .005,
= .46. Post-hoc t tests (one-sided; α = .03, Bonferroni-corrected) comparing contralateral versus ipsilateral waveforms for both colour and location targets revealed a significant N2pc for colour targets (contralateral: 0.83; µV; ipsilateral: 1.97 µV), t(13) = −5.41, p < .001, d = −1.40, and no N2pc for location targets, t(13) = −1.61, p = .131, d = −0.42.
Figure 4. (A) Event-related potentials (ERPs) elicited by colour targets after valid masked cues (top left panel) and invalid masked cues (top right panel). (B) ERPs elicited by colour targets after valid unmasked cues (bottom left panel) and ERPs elicited by location targets after valid unmasked cues (bottom right panel). The ERPs are shown at the electrode sites PO7 and PO8 contralateral (dashed lines) and ipsilateral (solid lines) to the target position. The N2pc can be seen between 200 und 300 ms after target onset and is significant, except for location targets (bottom right panel).
Behavioural data
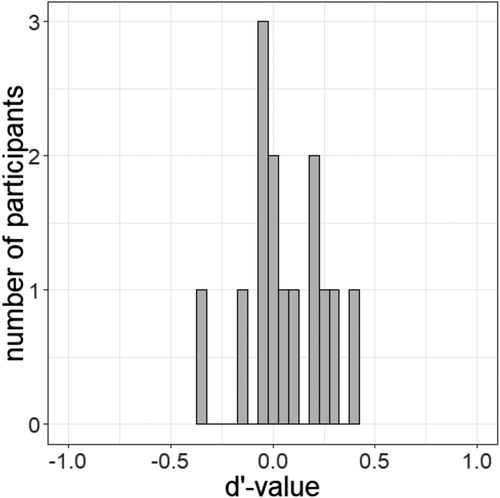
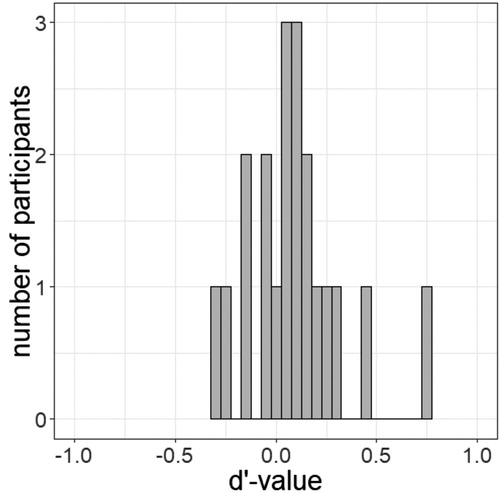
We asked participants to discriminate the cue word in Blocks 3 and 4 with a forced choice task to check if the masked presentation ensured subliminal presentation of the cue. Their performance was at chance level for masked cues (51.16%) but not for unmasked cues (95.27% correct judgments). d’ (cf. Green & Swets, Citation1966) for the masked cue was 0.06 (SDd’ = 0.19), which confirmed its invisibility, and 3.41 for the unmasked cue, which shows that it was consciously perceived in this condition. We used a t tests to compare the average of the d’-values per person to zero (see for the distribution of individual d’-values). The result confirmed the invisibility of the masked cue words, t(13) = 1.20, p = .252, d = 0.32 (no significant deviation from zero). Additionally, we calculated t tests comparing the accuracy of participants’ judgements to the guessing probability of 50%. For masked stimuli, the accuracy was equivalent to the guessing probability, t(13) = 1.17, p = .265, d = 0.31, whereas the accuracy to unmasked cues was significantly different from the guessing probability, t(13) = 36.79, p < .001, d = 9.83.
Figure 5. Distribution of the individual d’-values of all 14 participants in the masked condition of Experiment 1.
Only correct responses as well as response times within ±2 SD from the median per person per condition were included in the analysis (10.52% removed in total). For trials with masked cues, a repeated-measures ANOVA (2 × 2), with the variables Validity (valid/invalid) and ISI (34 ms or 700 ms) was computed (see and in Appendix 1 for mean reaction times). There were no significant effects, all Fs(1, 13) < 0.26, all ps > .616, all s < .02. For trials with unmasked cues, we conducted a repeated-measures ANOVA (2 × 2), with the variables Target Type (location/colour) and ISI (34 ms or 700 ms); see and for mean reaction times. The interaction between Target Type and ISI was significant, F(1, 13) = 15.23, p = .002,
= .54. Post-hoc t tests (α = .025; Bonferroni-corrected for two comparisons) showed that reaction times for colour targets were significantly faster than for location targets, both in short ISI trials (colour: 736 ms, location: 924 ms), t(13) = 6.13, p < .001, d = 1.58, and long ISI trials (colour: 656 ms, location 739 ms), t(13) = 3.64, p = .003, d = 0.94.
Figure 6. Mean reaction times (RTs) in trials with masked cue words (left panel) and unmasked cue words (right panel). Left panel: Dashed lines represent valid trials, solid lines invalid trials. Right panel: Dashed lines represent trials with location targets, solid lines trials with colour targets. The blue lines show slow reaction times after median split, the red lines fast reaction times after median split and the green lines the average of the complete data. Error bars represent average SEs.
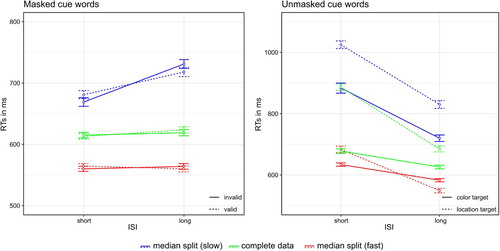
As the low error rate of the participants (Merror = 4.87%, SDerror = 3.57%) suggested the possibility of a speed-accuracy tradeoff, we repeated the ANOVAs after a median split (see ). For the trials with masked cue words, no effects were significant, all Fs(1, 13) < 1.35, all ps > .266, all s < .09. For trials with unmasked cues, we found significant main effects for Target Type (colour: 828 ms, location: 948 ms), F(1, 13) = 29.19, p < .001,
= .69, and ISI (short: 819 ms, long: 984 ms), F(1, 13) = 18.85, p < .001,
= .59, in the slow responses. In the fast responses, the interaction between Target Type and ISI was significant, F(1, 13) = 6.79, p = .002,
= .34. Post-hoc t tests (α = .025; Bonferroni-corrected for two comparisons) showed no significant effects: short ISI (colour: 663 ms, location 718 ms), t(13) = 2.84, p = .027, d = 0.64, long ISI (colour: 593 ms, location: 577 ms), t(13) = 0.16, p = .876, d = 0.04.
Discussion
We found no evidence for attention shifts elicited by masked cues: no validity effects in the reaction times, as well as no ERPs reflecting spatial attention shifts (EDAN, ADAN, and LDAP). In addition, the N2pc component to colour targets was independent of the validity of the preceding masked cue. For unmasked cues, we found a significant LDAP at the right hemisphere (PO8), which shows at least marginal influences of unmasked central cue words on spatial attention. (Given the current procedure, with 100% valid unmasked location word cues, we were not able to calculate a cueing effect of the unmasked word cues.) As expected, the N2pc was significant in the visual search task (colour targets). (For location targets, the contra-to-ipsilateral ERP difference might have started a little later, see .) Reaction times to colour targets were significantly faster than to location targets, which reflects the difference in difficulty.
Even though we ensured that the cue word was relevant for the task by using location targets (where only the identity of the cue word pointed out the target) and 100% valid visible cues as a context for encouraging the usage of cue information even in masked cue conditions, we found no validity effects by masked cue words in the reaction times. In addition, no ERPs showing spatial attention shifts to the left or right (EDAN, ADAN, and LDAP) were elicited by the masked cue words. To account for the possibility that the centrally presented masked cue words did not elicit an attention shift because they were not processed at all, we designed a second experiment.
Experiment 2
In Experiment 2, we included a target word discrimination task. By means of this additional task, we can control if the subliminal words were processed at all, even if they produce no attentional effects. In each trial of the target-word discrimination task, following the cue/prime word, the clearly visible word left or right was shown centrally as a target, and participants had to identify this target word as left or right by button press. Based on the existing literature, we expected a priming effect reflecting the processing of subliminal words: If subliminal cue/prime word and target word meaning are congruent, the reaction should be faster than if they are incongruent (for a review, see, e.g., Ansorge et al., Citation2013). Additionally, the ERP component N400 to the target word, which reflects semantic dissimilarities between sequentially presented words, should show as a more negative amplitude of activity in incongruent than in congruent trials (e.g., Kiyonaga, Grainger, Midgley, & Holcomb, Citation2007; Martens, Ansorge, & Kiefer, Citation2011; Ortells, Kiefer, Castillo, Megías, & Morillas, Citation2016).
Additionally, we wanted to replicate our findings from Experiment 1 with a slightly altered visual search task (search for a coloured bar presented amongst three distractors). This colour search task was used, as past research has shown that a cueing effect can be measured in this task (cf. Folk, Remington, & Johnston, Citation1992; Palmer & Mattler, Citation2013), even with subliminal (but peripheral) cues (Ansorge et al., Citation2009). This task provided an independent, second measure of any attentional effects of the masked words.
Method
Participants
We calculated the minimum required sample size to detect a small congruency effect (15 ms) for congruent trials in the short ISI condition while obtaining either no effect or an inversed congruency effect for incongruent trials in the long ISI condition. For this, we simulated data based on our design and the actual distribution of our data (cf. Brysbaert & Stevens, Citation2018): The design of Experiment 2 consisted of four cells (ISI short & congruent, ISI short & incongruent, ISI long & congruent, and ISI long & incongruent) with 48 trials per cell. We used the parameters of the actual distribution of our data to simulate data of 100 random studies following our design (4 cells, 48 trials each), with a desired power of at least 80%. The results showed that a minimum of 18 participants was required to detect the expected congruency effects in the word discrimination task (α = 0.05, power ≥ 0.80). Twenty-three right-handed psychology students from the University of Vienna participated in Experiment 2 (MAge = 22.00 years, SDAge = 3.37 years). Three students had to be excluded due to poor quality of the EEG data. The students received either course credit or a monetary expense allowance for their participation. Treatment of participants was the same as in Experiment 1.
Apparatus, stimuli, and electrophysiological recording
The setup remained equivalent to Experiment 1. The size of the coloured bars (visual search task) was 1.5° × 0.3° of visual angle. The four bars were arranged equidistant to the central fixation (4.4° visual angle), in the positions upper left, upper right, lower left, and lower right. The longer ISI of 700 ms was filled with a dummy word consisting of five randomly drawn lowercase letters, to keep the content of the screen preceding the targets constant for EEG-data analyses (both dummy word and backward mask consist of centrally presented letters).
In addition to the ERPs analyzed in Experiment 1, we examined the N400 reflecting semantic relatedness at 12 fronto-central electrodes (F1, Fz, F2, C1, Cz, C2, CP1, CPz, CP2, P1, Pz, P2) within a time window between 285 and 385 ms after word target onset (e.g., Martens et al., Citation2011; Ortells et al., Citation2016).
Task, design, and procedure
The experiment consisted of three different tasks: visual search (experimental task with the same purpose as in Experiment 1), target word discrimination (control task for processing of subliminal cue/prime word), and cue/prime discrimination (control task for cue/prime visibility). In the visual search task, following the masked cue/prime word, the word color in the centre of the screen indicated the task and helped keeping the gaze in the centre of the screen, while a peripherally presented bar in a predefined colour had to be searched for and its orientation (horizontal or vertical) had to be reported by keypress as fast as possible. The visual search task was built after existing protocols that have successfully been used to measure both conscious/supraliminal and unconscious/subliminal cueing effects many times by an N2pc before (cf. Ansorge et al., Citation2009; Eimer & Kiss, Citation2008). The target colour for the visual search task as well as the mapping of the response keys to horizontal versus vertical target bars were counterbalanced across participants. The cue/prime word was non-predictive (50% valid/congruent), the target appeared randomly at one out of four positions (upper left, upper right, lower left, lower right), and the three distractors were coloured randomly in two further colours (e.g., two blue distractors and one red distractor). The latter was done to ensure that the target does not pop out by colour and therefore−prevent singleton search for the target colour (Bacon & Egeth, Citation1994). This task still allows search for a single feature of colour but that does not preclude (subliminal) cueing effects in other cases (e.g., Ansorge et al., Citation2009; Ansorge, Horstmann, & Worschech, Citation2010; Folk et al., Citation1992; Schoeberl et al., Citation2015).
In the target-word discrimination task, following the masked cue/prime word, participants had to discriminate the central target word left or right by pressing the corresponding key as fast as possible. Trials of the visual search and word discrimination task were presented equally frequent and in random order. The third task, cue/prime discrimination (see Experiment 1), only occurred in the second half (Block 2) of Experiment 2 as an additional task in each trial, following the response to the first task (word discrimination or visual search). Experiment 2 consisted of 544 trials (384 in Block 1; 160 in Block 2). As we were only interested in the processing of the subliminal cue/prime words, the condition with supraliminally presented words from Experiment 1 was removed. See for exemplary trials of Experiment 2.
Figure 7. Exemplary trials of Experiment 2, Block 1 (from left to right): The fixation cross, forward mask, cue/prime word (right), backward mask, the optional longer ISI condition filled with a dummy word (upper trial), and two different versions of the target display are shown. The upper trial shows a target display of the visual search task. The target is the green bar at the upper right position (its location was validly cued by the preceding word right). The lower trial shows a target of the target-word discrimination task (target word left, incongruent to the preceding cue/prime word right). Stimuli are not drawn to scale. The arrow depicts the flow of time.
Results
EEG data
N400
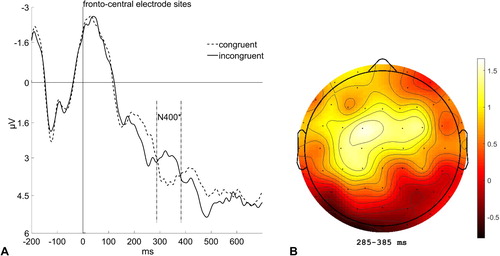
The N400 is an ERP component reflecting semantic relatedness. It is usually located with a maximum at fronto-central electrodes and shows as a greater negativity at about 400 ms post-target, if the semantic relatedness is weak or low. We calculated the mean amplitude values of 12 fronto-central electrodes, in the time window between 285 and 385 ms (see ). Only trials with the short ISI (34 ms) were analyzed, as the N400 in trials with the long ISI (700 ms) would only reflect the constant incongruence between the target word and the preceding supraliminally presented dummy word, which is of no interest to our research question. A repeated-measures ANOVA (2 × 3 × 4), with the variables Congruence (cue/prime word and target word congruent; cue/prime word and target word incongruent), Laterality (left: F1, C1, CP1, P1; middle: Fz, Cz, CPz, Pz; right: F2, C2, CP2, P2), and Caudality (frontal: F1, Fz, F2; central: C1, Cz, C2; centro-parietal: CP1, CPz, CP2; parietal: P1, Pz, P2) was computed. The degrees of freedom were adapted using a Greenhouse-Geisser correction. There was a marginally significant main effect for Congruence, F(1, 19) = 4.39, p = .050, = .19, which reflects a less positive (i.e., more negative) mean amplitude for incongruent than for congruent cue/prime-target word relations (2.89 and 3.86 µV), as well as a main effect for Caudality, F(1.89, 35.92) = 7.51, p = .002,
= .28, which shows generally increasing mean amplitudes from rostral to caudal electrode sites (frontal: 2.45 µV; central: 3.50 µV; centro-parietal: 3.86 µV; parietal: 3.99 µV). No further main effects or interaction effects were significant, all ps > .126, all
s < .10. For an omnibus ANOVA including the variables Word Identity and Block, the reader should refer to Appendix 2.
Figure 8. (A) Grand average of event-related potentials (ERPs) elicited at fronto-central electrode sites (F1, C1, CP1, P1, Fz, Cz, CPz, Pz, F2, C2, CP2, P2) in the target-word discrimination task depending on whether a trial is congruent (dashed line) or incongruent (solid line). Data is averaged across the steps of the nonsignificant variables Identity of the cue/prime word and Block. (B) Scalp distribution map showing the difference in brain activity between congruent and incongruent trials (congruent minus incongruent).
ADAN, EDAN, and LDAP
The ADAN, EDAN, and LDAP were analyzed as in Experiment 1 (see and Results from Experiment 1 for more information). The repeated-measures analyses of variance (ANOVAs; 2 × 2) of the long ISI conditions, with the variables Hemisphere (contralateral; ipsilateral) and Cue/Prime Word (left; right), yielded the following results (refer to Appendix 2 for an omnibus ANOVA including the variable Block): For the ADAN, a significant interaction effect of Hemisphere and Cue/Prime Word was found, F(1, 19) = 12.06, p = .003, = .39. Post-hoc t tests (one-sided; α-level Bonferroni-corrected) comparing the words left and right over each hemisphere show a significant difference between mean amplitudes over the right hemisphere: more negative voltages for the word left (−0.72 µV) than right (−0.18 µV), t(19) = −3.01, p = .004, d = −0.66. The difference was not significant over the left hemisphere, more negative for the word right (−0.24 µV) than left (−0.23 µV), t(19) = 0.02, p = .493, d < 0.01. As it is a common finding that the ADAN is only found over the right hemisphere (Hietanen, Leppänen, Nummenmaa, & Astikainen, Citation2008; Talsma et al., Citation2005), this effect demonstrates that spatial attention shifts might be elicited by subliminally presented spatial words.
Figure 9. (A) This figure shows event-related potentials (ERPs) elicited by the centrally presented masked cue word, depending on its identity: cue word left (dashed line), cue word right (solid line). Electrodes of the left hemisphere are depicted on the left (upper panel: FC5; lower panel: PO7). Electrodes of the right hemisphere are depicted on the right (upper panel: FC6; lower panel: PO8). Data are averaged across steps of the nonsignificant variable Block. The anterior directing-attention negativity (ADAN) can be seen between 300 and 500 ms (FC6), the early directing-attention negativity (EDAN) between 250 and 450 ms (PO7) and the late attention-directing positivity (LDAP) between 500 and 800 ms (PO8). (B) Scalp distribution maps showing the difference between brain activity for the cue/prime word left and right at selected time points.
Analysis of the EDAN showed a significant main effect for Cue/Prime Word, F(1, 19) = 5.43, p = .031, = .22 (left: 1.27 µV, right: 0.91 µV). This difference might reflect the different features of the words (cf. van Velzen & Eimer, Citation2003) and proves that the subliminal words have been processed. There were no further significant effects, all Fs(1, 19) < 2.61, all ps > .123, all
s < .12. These results show no proof for attention shifts elicited by the cue/prime word.
For the LDAP, only the main effect of Hemisphere was significant, F(1, 19) = 7.09, p = .015, = .27 (PO7: 0.61 µV, PO8: −0.66 µV). No further effects were significant, Fs(1, 19) < 3.11, all ps > .094, all
s < .14. As no interaction between Hemisphere and Cue/Prime Word was significant, all Fs(1, 19) < 0.55, all ps > .469, all
s < .03, there is no proof for spatial attention shifts elicited by the subliminal cue/prime word in the LDAP.
N2 posterior contralateral
The N2pc was defined as in Experiment 1 (see Results of Experiment 1 for more details). A repeated-measures ANOVA (2 × 2), with the variables Validity (cue/prime word validly cued the target side; cue/prime word invalidly cued the target side) and Hemisphere (contralateral; ipsilateral) was computed (see for graphs and scalp maps as well as , Appendix 1, for the mean amplitude differences). It yielded a significant interaction between Validity and Hemisphere, F(1, 19) = 7.23, p = .015, = .28. Post-hoc t tests (one-sided; α = .03, Bonferroni-corrected for two comparisons) examining the difference between contralateral and ipsilateral electrodes (variable Hemisphere) for both valid and invalid trials (variable Validity) showed a marginally significant difference in valid trials, t(19) = −2.44, p = .025, d = −0.53 (contralateral: 1.75 µV, ipsilateral: 2.25 µV), and no significant difference in invalid trials, t(19) = 0.11, p = .911, d = 0.02 (contralateral: 1.83 µV, ipsilateral: 1.81 µV). For an omnibus ANOVA including the variables ISI and Block, see Appendix 2.
Figure 10. (A) Event-related potentials (ERPs) elicited in the visual search task in valid trials (left panel) and invalid trials (right panel) at the electrode sites PO7 and PO8 contralateral (dashed lines) and ipsilateral (solid lines) to the target bar. Data are averaged across the steps of the nonsignificant variables (ISI and Block). The N2pc can be seen between 200 und 300 ms after target onset. (B) Scalp distribution map showing the difference in brain activity for a target at the left versus right side.
Behavioural data
With a forced-choice task, asking participants to discriminate the masked cue/prime word in Block 2 of Experiment 2, we checked if the masked presentation ensured subliminal presentation of the cue/prime. Participants’ performance was at chance level (51.63%). We computed the mean d’-value for all participants (cf. Green & Swets, Citation1966) for the masked prime word, mean d’ = 0.08 (SDd’ = 0.24; see for the distribution). A t test comparing the average of the d’-values per person against zero showed no significant effect, t(19) = 1.59, p = .127, d = 0.36, neither did a t test comparing the discrimination performance with the guessing probability of 50%, t(19) = 1.63, p = .120, d = 0.36, which both confirmed the invisibility of the cue/prime word.
Figure 11. Distribution of the individual d’-values of all 20 participants of Experiment 2.
Only data from Block 1 (without additional cue/prime discrimination task) was analyzed for congruency/validity effects. Trials with false responses were removed, as well as responses faster or slower than two SDs from the median per person and condition (total removal: 8.79%). Separately for each task (visual search and target-word discrimination), a repeated-measures ANOVA (2 × 2), with the variables Validity (valid/invalid; for visual search trials) or Congruence (congruent/incongruent; for target word discrimination trials) and ISI (34 ms or 700 ms), was computed (see and for mean reaction times). There was a significant main effect for ISI in the target word discrimination task, F(1, 19) = 26.46, p < .001, = .58. Participants reacted significantly faster when the ISI was long (683 ms) than when it was short (709 ms). In the visual search task, there were no significant effects, all Fs(1, 19) < 3.69, all ps > .070, all
s < .16.
Figure 12. Mean reaction times (RTs) to visual search targets (left panel) and word discrimination targets (right panel). Dashed lines represent valid/congruent trials, solid lines invalid/incongruent trials. The blue lines show slow reaction times after median split, the red lines fast reaction times after median split and the green lines the average of the complete data. Error bars represent average SEs.
We did the same ANOVAs after a median split (see ), as the low error rate of the participants (Merror = 3.80%, SDerror = 1.91%) suggested the possibility of a speed-accuracy tradeoff. The median split was a post-hoc decision. The mean reaction times after the median split can be found in . In the word discrimination task, we have a significant main effect for ISI in both the slow, F(1, 19) = 22.02, p < .001, = .54 (long ISI: 764 ms; short ISI: 791 ms), and the fast reaction times, F(1, 19) = 18.93, p < .001,
= .50 (long ISI: 613 ms; short ISI: 637 ms). Also, a significant main effect for congruence (congruent: 784 ms; incongruent: 771 ms) was found in the slow reaction times, F(1, 19) = 5.82, p = .026,
= .23. This effect reflects a negative congruence effect (NCE; Eimer & Schlaghecken, Citation1998), an inversion of the congruence effect showing as faster responses in incongruent than congruent trials if additional time has passed since the masked prime (e.g., Boy & Sumner, Citation2010). The NCE often occurs only in the slow RTs (Ocampo & Finkbeiner, Citation2013). In the visual search task, there was also a significant main effect for ISI in the fast reaction times, F(1, 19) = 4.90, p < .039,
= .21 (long ISI: 745 ms; short ISI: 732 ms), but no significant effect in the slow reaction times, F(1, 19) = 2.71, p = .116,
= .12. No further effects were significant, all Fs(1, 19) < 3.21, all ps > .089, all
s < .14.
Discussion
To check for the processing of the subliminal cue/prime word, we analyzed the N400 and response times to the targets in the target word discrimination task. The significant difference between congruent and incongruent words both in the N400 (greater negativity for incongruent than congruent words) and the negative congruence effect (NCE) in the slow response times after median split showed that the subliminal cue/prime word had been processed and influenced both the neurophysiological and behavioural reaction depending on its congruence with the target word. In addition, the significant main effect of the identity of the cue/prime word in the EDAN analysis proves that the prime words had been discriminated. At first glance, the mere NCE in the slow RTs of the word discrimination task might be surprising, as the significant N400 proves the influence of the congruence between cue/prime word and target word (word discrimination task). However, the lacking reaction time effects might be due to the difficulty of working on two different task randomly mixed throughout the experiment or to the different ISIs presented randomly intermixed in Experiment 2: It has been shown in the past that temporal attention to the target is essential to give rise to congruence effects but with intermixed ISIs participants could not anticipate when the target was presented very well. Such temporal uncertainty abolishes (cf. Naccache, Blandin, & Dehaene, Citation2002) or at least considerably weakens the behavioural effects (Doherty, Rao, Mesulam, & Nobre, Citation2005; Rohenkohl, Gould, Pessoa, & Nobre, Citation2014). This would also explain why the N400 did not produce larger effects in our design (Kiefer & Brendel, Citation2006).
Regarding spatial attention shifts, our results are in line with the results from Experiment 1: The lack of reliable validity effects in the reaction times and the missing ERP effects (no significant EDAN or LDAP differences; significant ADAN only over one hemisphere) can be seen as evidence against the idea that subliminal cue words can alter spatial attention in visual colour search.
General Discussion
We examined the influence of subliminally and centrally presented cue/prime words on spatial attention shifts in two experiments. Specific ERP components reflecting spatial attention shifts following cue/prime word onset (EDAN, ADAN, and LDAP) were analyzed. Additionally, we inspected the N2pc and reaction times to a visual search target depending on the validity between the identity of the subliminal cue/prime word and the side of the screen where the target was shown. In Experiment 1, we included a second target type (location target), where the target was not recognizable by colour, but only by the preceding cue word. To boost the relevance of the cue word even more, we also included unmasked cue words, which validly cued the target location in 100% of the trials and had to be used to find the location targets. To control for the processing of the subliminal cue/prime word, we added an additional target discrimination task in Experiment 2 and analyzed the N400 reflecting semantic dissimilarities and reaction times depending on the congruence between subliminal cue/prime word and target word. Both the sensitivity index d’ as well as the discrimination ability on chance level verified that the cue/prime words in Experiments 1 and 2 were presented subliminally.
In Experiment 1, we found no evidence for spatial attention shifts elicited by subliminally presented words, neither in the reaction times nor in the ERPs. In Experiment 2, apart from replicating the results of Experiment 1 in most of our dependent variables, we found a significant ADAN over one hemisphere as well as a difference in the N2pc amplitude to the target, depending on the validity of the subliminal cue word. The N2pc has been analyzed depending on a preceding cue before. For example, Kiss et al. (Citation2008) and Praamstra (Citation2006) found a reduced N2pc amplitude for informative cues providing advance spatial information. In Experiment 2, we found a difference depending on the relation between cue and target (enhanced target-elicited N2pc amplitude in valid compared to invalid trials). This is expected, as Kiss et al. found a consistent posterior negativity contralateral to the cued side even in target-absent trials, starting at the same time window as the N2pc in target-present trials. This posterior negativity due to the cue might add up with the target induced N2pc in valid trials to produce a greater summed activity than in invalid trials, where the negativity elicited by the cue is on the other side than that elicited by the target.
Although in the ERPs of Experiment 2, we might have found some evidence for spatial attention shifts by subliminal cues words, in Experiment 1, this was not the case: The subliminal central cue words neither elicited ERPs reflecting spatial attention shifts (EDAN, ADAN, LDAP), nor did the target N2pc components differ depending on the validity of the preceding masked cue. In addition, a behavioural cueing effect of the subliminal cues was missing. As in Experiment 1, under conditions of certainty about the time of the stimulus presentation and in the context of a 100% valid supraliminal word cue, no subliminal word cueing effect could be found, we would conclude that subliminal words do not elicit attention shifts, as this was sensitive and exhaustive methodology.
In addition, we found a congruence effect of the masked prime words in the control conditions of Experiment 2. Though some authors believe that such effects of subliminal stimuli are stimulus-driven, this cannot be concluded from the present study. It is true that the masked cue/prime words were uninformative of target word identities (Exp. 2). However, task relevance of the word cues was ensured by asking the participants to discriminate the same words as visible targets (Exp. 2). Prior studies have shown that such instructions to process the visible targets are sufficient to reliably elicit top-down dependent processing of the same stimuli if masked (Klinger, Burton, & Pitts, Citation2000; Kunde, Kiesel, & Hoffmann, Citation2003). In addition, although we observed an N400 in response to the congruence of the relation between masked cue/prime word and target word, and this N400 would be evidence of a semantic processing of the cue/prime words, one cannot conclude that semantic processing also underlies the found inverse reaction-time congruence effect in the slow responses of Experiment 2. These effects could have likewise reflected the congruence versus incongruence of the responses that were activated by the cue/prime words and the target words, respectively.
Conclusion
We exhaustively tested whether subliminally presented central word cues can elicit spatial attention shifts, by looking at behavioural data (validity effects) as well as EEG-data (EDAN, ADAN, LDAP, N2pc, N400) in two experiments. We varied the relevance of the cue word, the predictability of the target onset, and the difficulty of the search. Yet, our findings show that although subliminally presented central word cues are being processed, they do not elicit spatial attention shifts in visual colour search.
Acknowledgements
We are grateful to Christoph Huber-Huber for his assistance in processing the EEG data, and to Sophie Hanke, Marie-Luise Augsten, Marlene Forstinger and Valeria Natalia La Notte for helping with the collection of data.
Disclosure statement
No potential conflict of interest was reported by the authors.
ORCID
Diane Baier http://orcid.org/0000-0002-2293-8572
Ulrich Ansorge http://orcid.org/0000-0002-2421-9942
Notes
* The data that support the findings of this study are available from the corresponding author, D.B., upon request.
1 The N2pc cannot be observed in direct response to a centrally presented stimulus alone. The N2pc can only be recorded in response to a peripheral stimulus, for instance, a target of a visual search task when the target is presented on either the left or the right side of the screen. In many cases, including the present study, the N2pc to a relevant target on one side is measured in the presence of irrelevant distractors on the other side. In these conditions, the recorded contra-ipsilateral difference probably reflects a mixture of directing of attention to the target side (an NT), with more negative activity contra- than ipsilateral to the target, and a distractor positivity (PD)–that is, more positive activity contralateral than ipsilateral of the distractor, indicative of suppression of the distractor (Hickey, Di Lollo, & McDonald, Citation2009; Li, Liu, & Hu, Citation2017). However, in keeping with prior studies, in the following we shall continue to label the contra-ipsilateral difference in response to the lateral targets of a visual search N2pc.
References
- Al-Janabi, S., & Finkbeiner, M. (2012). Effective processing of masked eye gaze requires volitional control. Experimental Brain Research, 216, 433–443. doi: 10.1007/s00221-011-2944-0
- Ansorge, U., Horstmann, G., & Worschech, F. (2010). Attentional capture by masked colour singletons. Vision Research, 50, 2015–2027. doi: 10.1016/j.visres.2010.07.015
- Ansorge, U., Khalid, S., & König, P. (2013). Space-valence priming with subliminal and supraliminal words. Frontiers in Psychology, 4, 81. doi: 10.3389/fpsyg.2013.00081
- Ansorge, U., Khalid, S., & Laback, B. (2016). Unconscious cross-modal priming of auditory sound localization by visual words. Journal of Experimental Psychology: Learning, Memory, and Cognition, 42, 925–937. doi: 10.1037/xlm0000217
- Ansorge, U., Kiss, M., & Eimer, M. (2009). Goal-driven attentional capture by invisible colours: Evidence from event-related potentials. Psychonomic Bulletin & Review, 16, 648–653. doi:10.3758/PBR.16.4.64 doi: 10.3758/PBR.16.4.648
- Ansorge, U., Kunde, W., & Kiefer, M. (2014). Unconscious vision and executive control: How unconscious processing and conscious action control interact. Consciousness and Cognition, 27, 268–287. doi: 10.1016/j.concog.2014.05.009
- Bacon, W. F., & Egeth, H. E. (1994). Overriding stimulus-driven attentional capture. Perception & Psychophysics, 55, 485–496. doi: 10.3758/BF03205306
- Berger, A. (1999). Peripheral non-informative cues do induce early facilitation of target detection. European Journal of Cognitive Psychology, 11, 119–137. doi: 10.1080/713752304
- Boy, F., & Sumner, P. (2010). Tight coupling between positive and reversed priming in the masked prime paradigm. Journal of Experimental Psychology: Human Perception and Performance, 36, 892–905. doi: 10.1037/a0017173
- Brignani, D., Guzzon, D., Marzi, C. A., & Miniussi, C. (2009). Attentional orienting induced by arrows and eye-gaze compared with an endogenous cue. Neuropsychologia, 47, 370–381. doi: 10.1016/j.neuropsychologia.2008.09.011
- Brysbaert, M., & Stevens, M. (2018). Power analysis and effect size in mixed effects models: A tutorial. Journal of Cognition, 1, 1–20. doi: 10.5334/joc.10
- Carr, T. H., & Dagenbach, D. (1990). Semantic priming and repetition priming from masked words: Evidence for a center-surround attentional mechanism in perceptual recognition. Journal of Experimental Psychology: Learning, Memory, and Cognition, 16, 341–350. doi:10.1037/0278-7393.16.2.341.
- Castel, A. D., Chasteen, A. L., Scialfa, C. T., & Pratt, J. (2003). Adult age differences in the time course of inhibition of return. The Journals of Gerontology Series B: Psychological Sciences and Social Sciences, 58, 256–259. doi: 10.1093/geronb/58.5.P256
- Dehaene, S., Changeux, J. P., Naccache, L., Sackur, J., & Sergent, C. (2006). Conscious, preconscious, and subliminal processing: A testable taxonomy. Trends in Cognitive Sciences, 10, 204–211. doi: 10.1016/j.tics.2006.03.007
- Delorme, A., & Makeig, S. (2004). EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. Journal of Neuroscience Methods, 134, 9–21. doi: 10.1016/j.jneumeth.2003.10.009
- Doherty, J. R., Rao, A., Mesulam, M. M., & Nobre, A. C. (2005). Synergistic effect of combined temporal and spatial expectations on visual attention. Journal of Neuroscience, 25, 8259–8266. doi: 10.1523/JNEUROSCI.1821-05.2005
- Eimer, M., Forster, B., Fieger, A., & Harbich, S. (2004). Effects of hand posture on preparatory control processes and sensory modulations in tactile-spatial attention. Clinical Neurophysiology, 115, 596–608. doi: 10.1016/j.clinph.2003.10.015
- Eimer, M., & Kiss, M. (2008). Involuntary attentional capture is determined by task set: Evidence from event-related brain potentials. Journal of Cognitive Neuroscience, 20, 1423–1433. doi: 10.1162/jocn.2008.20099
- Eimer, M., & Schlaghecken, F. (1998). Effects of masked stimuli on motor activation: Behavioral and electrophysiological evidence. Journal of Experimental Psychology: Human Perception and Performance, 24, 1737–1747. doi: 10.1037/0096-1523.24.6.1737
- Feng, Q., & Zhang, X. (2014). Eye gaze triggers reflexive attention shifts: Evidence from lateralised ERPs. Brain Research, 1589, 37–44. doi: 10.1016/j.brainres.2014.09.029
- Folk, C. L., Remington, R. W., & Johnston, J. C. (1992). Involuntary covert orienting is contingent on attentional control settings. Journal of Experimental Psychology: Human Perception and Performance, 18, 1030–1044. doi: 10.1037/0096-1523.18.4.1030
- Gibson, B. S., & Kingstone, A. (2006). Visual attention and the semantics of space beyond central and peripheral cues. Psychological Science, 17, 622–627. doi: 10.1111/j.1467-9280.2006.01754.x
- Green, D. M., & Swets, J. A. (1966). Signal detection theory and psychophysics. Oxford, England: John Wiley.
- Greenwald, A. G., Draine, S. C., & Abrams, R. L. (1996). Three cognitive markers of unconscious semantic activation. Science, 273, 1699–1702. doi: 10.1126/science.273.5282.1699
- Gromer, D. (2017). apa: Format Outputs of Statistical Tests According to APA Guidelines. R package version 0.2.0. https://CRAN.R-project.org/package=apa
- Harter, M. R., Miller, S. L., Price, N. J., LaLonde, M. E., & Keyes, A. L. (1989). Neural processes involved in directing attention. Journal of Cognitive Neuroscience, 1, 223–237. doi: 10.1162/jocn.1989.1.3.223
- Hassin, R. R. (2013). Yes it can: On the functional abilities of the human unconscious. Perspectives on Psychological Science, 8, 195–207. doi: 10.1177/1745691612460684
- Hickey, C., Di Lollo, V., & McDonald, J. J. (2009). Electrophysiological indices of target and distractor processing in visual search. Journal of Cognitive Neuroscience, 21, 760–775. doi: 10.1162/jocn.2009.21039
- Hietanen, J. K., Leppänen, J. M., Nummenmaa, L., & Astikainen, P. (2008). Visuospatial attention shifts by gaze and arrow cues: An ERP study. Brain Research, 1215, 123–136. doi: 10.1016/j.brainres.2008.03.091
- Holmes, A., Mogg, K., Garcia, L. M., & Bradley, B. P. (2010). Neural activity associated with attention orienting triggered by gaze cues: A study of lateralized ERPs. Social Neuroscience, 5, 285–295. doi: 10.1080/17470910903422819
- Hommel, B., Pratt, J., Colzato, L., & Godijn, R. (2001). Symbolic control of visual attention. Psychological Science, 12, 360–365. doi: 10.1111/1467-9280.00367
- Hopf, J. M., & Mangun, G. R. (2000). Shifting visual attention in space: An electrophysiological analysis using high spatial resolution mapping. Clinical Neurophysiology, 111, 1241–1257. doi: 10.1016/S1388-2457(00)00313-8
- Jolicœur, P., Brisson, B., & Robitaille, N. (2008). Dissociation of the N2pc and sustained posterior contralateral negativity in a choice response task. Brain Research, 1215, 160–172. doi: 10.1016/j.brainres.2008.03.059
- Jongen, E. M., Smulders, F. T., & van der Heiden, J. S. (2007). Lateralized ERP components related to spatial orienting: Discriminating the direction of attention from processing sensory aspects of the cue. Psychophysiology, 44, 968–986. doi: 10.1111/j.1469-8986.2007.00557.x
- Kelly, S. P., Gomez-Ramirez, M., & Foxe, J. J. (2009). The strength of anticipatory spatial biasing predicts target discrimination at attended locations: A high-density EEG study. European Journal of Neuroscience, 30, 2224–2234. doi: 10.1111/j.1460-9568.2009.06980.x
- Kiefer, M. (2002). The N400 is modulated by unconsciously perceived masked words: Further evidence for a spreading activation account of N400 priming effects. Cognitive Brain Research, 13, 27–39. doi: 10.1016/S0926-6410(01)00085-4
- Kiefer, M., & Brendel, D. (2006). Attentional modulation of unconscious ‘automatic’ processes: Evidence from event-related potentials in a masked priming paradigm. Journal of Cognitive Neuroscience, 18, 184–198. doi: 10.1162/jocn.2006.18.2.184
- Kiss, M., van Velzen, J., & Eimer, M. (2008). The N2pc component and its links to attention shifts and spatially selective visual processing. Psychophysiology, 45, 240–249. doi: 10.1111/j.1469-8986.2007.00611.x
- Kiyonaga, K., Grainger, J., Midgley, K., & Holcomb, P. J. (2007). Masked cross-modal repetition priming: An event-related potential investigation. Language and Cognitive Processes, 22, 337–376. doi: 10.1080/01690960600652471
- Klinger, M. R., Burton, P. C., & Pitts, G. S. (2000). Mechanisms of unconscious priming. I. Response competition, not spreading activation. Journal of Experimental Psychology: Learning, Memory, and Cognition, 26, 441–455. doi: 10.1037//0278-7393.26.2.441
- Kunde, W., Kiesel, A., & Hoffmann, J. (2003). Conscious control over the content of unconscious cognition. Cognition, 88, 223–242. doi: 10.1016/S0010-0277(03)00023-4
- Lawrence, M. A. (2016). ez: Easy Analysis and Visualization of Factorial Experiments. R package version 4.4-0. https://CRAN.R-project.org/package=ez
- Li, C., Liu, Q., & Hu, Z. (2017). Further evidence that N2pc reflects target enhancement rather than distracter suppression. Frontiers in Psychology, 8, 2275. doi: 10.3389/fpsyg.2017.02275
- Livingstone, A. C., Christie, G. J., Wright, R. D., & McDonald, J. J. (2017). Signal enhancement, not active suppression, follows the contingent capture of visual attention. Journal of Experimental Psychology: Human Perception and Performance, 43, 219–224. doi: 10.1037/xhp0000339
- Lopez-Calderon, J., & Luck, S. J. (2014). ERPLAB: An open-source toolbox for the analysis of event-related potentials. Frontiers in Human Neuroscience, 8, 213. doi: 10.3389/fnhum.2014.00213
- Luck, S. J., & Gaspelin, N. (2017). How to get statistically significant effects in any ERP experiment (and why you shouldn't). Psychophysiology, 54, 146–157. doi: 10.1111/psyp.12639
- Luck, S. J., & Hillyard, S. A. (1994). Spatial filtering during visual search: Evidence from human electrophysiology. Journal of Experimental Psychology: Human Perception and Performance, 20, 1000–1014. doi:10.1037/0096–1523.20.5.1000
- Marcel, A. (1983). Conscious and unconscious perception: Experiments on visual masking and word recognition. Cognitive Psychology, 15, 197–237. doi:10.1016/0010-0285(83)90010-5 doi: 10.1016/0010-0285(83)90009-9
- Martens, U., Ansorge, U., & Kiefer, M. (2011). Controlling the unconscious: Attentional task sets modulate subliminal semantic and visuomotor processes differentially. Psychological Science, 22, 282–291. doi: 10.1177/0956797610397056
- MATLAB Release. (2014). The MathWorks, Inc., Natick, MA, USA.
- Mattler, U. (2003). Priming of mental operations by masked stimuli. Perception & Psychophysics, 65, 167–187. doi: 10.3758/BF03194793
- Naccache, L., Blandin, E., & Dehaene, S. (2002). Unconscious masked priming depends on temporal attention. Psychological Science, 13, 416–424. doi: 10.1111/1467-9280.00474
- Ocampo, B., & Finkbeiner, M. (2013). The negative compatibility effect with relevant masks: A case for automatic motor inhibition. Frontiers in Psychology, 4, 822. doi: 10.3389/fpsyg.2013.00822
- Ortells, J. J., Kiefer, M., Castillo, A., Megías, M., & Morillas, A. (2016). The semantic origin of unconscious priming: Behavioral and event-related potential evidence during category congruence priming from strongly and weakly related masked words. Cognition, 146, 143–157. doi: 10.1016/j.cognition.2015.09.012
- Palmer, S., & Mattler, U. (2013). Masked stimuli modulate endogenous shifts of spatial attention. Consciousness and Cognition, 22, 486–503. doi: 10.1016/j.concog.2013.02.008
- Posner, M. I., Snyder, C. R. R., & Davidson, B. J. (1980). Attention and the detection of signals. Journal of Experimental Psychology: General, 109, 160–174. doi: 10.1037/0096-3445.109.2.160
- Praamstra, P. (2006). Prior information of stimulus location: Effects on ERP measures of visual selection and response selection. Brain Research, 1072, 153–160. doi: 10.1016/j.brainres.2005.11.098
- Psychology Software Tools, Inc. [E-Prime 2.0]. (2013). Retrieved from http://www.pstnet.com
- R Core Team. (2017). R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. URL https://www.R-project.org/
- Reuss, H., Kiesel, A., Kunde, W., & Wühr, P. (2012). A cue from the unconscious–masked symbols prompt spatial anticipation. Frontiers in Psychology, 3, 397. doi: 10.3389/fpsyg.2012.00397
- Reuss, H., Pohl, C., Kiesel, A., & Kunde, W. (2011). Follow the sign! Top-down contingent attentional capture of masked arrow cues. Advances in Cognitive Psychology, 7, 82–91. doi: 10.2478/v10053-008-0091-3
- Rohenkohl, G., Gould, I. C., Pessoa, J., & Nobre, A. C. (2014). Combining spatial and temporal expectations to improve visual perception. Journal of Vision, 14, 8. doi: 10.1167/14.4.8
- Schmidt, F., & Schmidt, T. (2010). Feature-based attention to unconscious shapes and colors. Attention, Perception, & Psychophysics, 72, 1480–1494. doi: 10.3758/APP.72.6.1480
- Schoeberl, T., Fuchs, I., Theeuwes, J., & Ansorge, U. (2015). Stimulus-driven attentional capture by subliminal onset cues. Attention, Perception, & Psychophysics, 77, 737–748. doi: 10.3758/s13414-014-0802-4
- Talsma, D., Slagter, H. A., Nieuwenhuis, S., Hage, J., & Kok, A. (2005). The orienting of visuospatial attention: An event-related brain potential study. Cognitive Brain Research, 25, 117–129. doi: 10.1016/j.cogbrainres.2005.04.013
- Tassinari, G., Aglioti, S., Chelazzi, L., Peru, A., & Berlucchi, G. (1994). Do peripheral non-informative cues induce early facilitation of target detection? Vision Research, 34, 179–189. doi: 10.1016/0042-6989(94)90330-1
- van Velzen, J., & Eimer, M. (2003). Early posterior ERP components do not reflect the control of attentional shifts toward expected peripheral events. Psychophysiology, 40, 827–831. doi: 10.1111/1469-8986.00083
Appendices
Appendix 1
Tables of reaction times (RT) and mean amplitude differences (MAD), for the different conditions of Experiments 1 and 2.
Table A1. Mean Reaction Times of Experiment 1: For Trials With Masked Cues, the Variables Validity and Inter-Stimulus Interval (ISI). For Trials With Unmasked Cues, the Variables Target Type and Inter-Stimulus Interval (ISI).
Table A2. Mean Amplitude Difference (MAD) of Experiment 2 in µV (Ipsilateral Minus Contralateral) in the Time Window Between 200 and 300 ms at the Electrode Sites PO7 and PO8 (N2pc) as a Function of the Variables Block, ISI and Validity.
Table A3. Mean Reaction Times of Experiment 2 in ms as a Function of the Variables Validity (Visual Search Trials)/Congruence (Word Discrimination Trials) and Inter-Stimulus Interval (ISI).
Table A4. Mean Reaction Times of Experiment 2 in ms After Median Split as a Function of the Variables Validity (Visual Search Trials)/Congruence (Word Discrimination Trials) and Inter-Stimulus Interval (ISI).
Appendix 2
Analyses of ERPs including all variables (not only the one we had a priori hypotheses on), can be found below.
A.1. LDAP
A repeated-measures analysis of variance (ANOVA; 2 × 2 × 2) of the long ISI conditions, with the variables Hemisphere (contralateral; ipsilateral), Cue/Prime Word (left; right) and Block (Block1; Block2 with additional cue/prime discrimination), yielded only a significant main effect of Hemisphere, F(1, 19) = 7.09, p = .015, = .27 (PO7: 0.61 µV, PO8: −0.66 µV). No further effects were significant, Fs(1, 19) < 3.11, all ps > .094, all
s < .14. As no interaction between Hemisphere and Cue/Prime Word was significant, all Fs(1, 19) < 0.55, all ps > .469, all
s < .03, there is no proof for spatial attention shifts elicited by the subliminal cue/prime word in the LDAP.
A.2. ADAN
A repeated-measures ANOVA (2 × 2 × 2) with mean amplitude values in the respective time window of the long ISI condition for the variables Hemisphere (contralateral; ipsilateral), Cue/Prime Word (left; right), and Block (Block1; Block2 with additional cue/prime discrimination) yielded a significant interaction effect of Hemisphere and Cue/Prime Word, F(1, 19) = 12.06, p = .003, = .39. Post-hoc t tests (one-sided; α-level Bonferroni-corrected) comparing the words left and right over each hemisphere show a significant difference between mean amplitudes over the right hemisphere: more negative Voltages for the word left (−0.72 µV) than right (−0.18 µV), t(19) = −3.01, p = .004, d = −0.66. This effect demonstrates that spatial attention shifts can be elicited by subliminally presented spatial words. The difference was not significant over the left hemisphere: more negative for the word right (−0.24 µV) than left (−0.23 µV), t(19) = 0.02, p = .493, d < 0.01. Irrelevant to the research question, the ANOVA held a significant interaction effect between Hemisphere and Block, F(1, 19) = 4.85, p = .040,
= .20. Mean amplitudes were more negatively lateralized in Block 1 (left hemisphere: −0.90 µV, right hemisphere: −0.49) than in Block 2, with the additional cue/prime discrimination task (left hemisphere: 0.43 µV, right hemisphere: −0.40 µV). However, post-hoc t tests (two-sided; α-level Bonferroni-corrected) looking into the difference between Blocks 1 and 2 were not significant: t(19) = −2.19, p = .042, d = −0.48 (left hemisphere) and t(19) = −0.19, p = .854, d = −0.04 (right hemisphere). No further effects were significant in the ANOVA, all Fs(1, 19) < 2.58, all ps > .125, all
s < .12.
A.3. EDAN
A repeated-measures ANOVA (2 × 2 × 2), with mean amplitude values in the respective time window of the long ISI condition for the variables Hemisphere (contralateral; ipsilateral), Cue/Prime Word (left; right), and Block (Block1; Block2 with additional cue/prime discrimination) held a significant main effect for Cue/Prime Word, F(1, 19) = 5.43, p = .031, = .22 (left: 1.27 µV, right: 0.91 µV). This difference proves that the subliminal word has been processed. However, there were no further significant effects, all Fs(1, 19) < 2.61, all ps > .123, all
s < .12. The results show no proof for attention shifts elicited by the cue/prime word.
A.4. N2pc
A repeated-measures ANOVA (2 × 2 × 2 × 2), with the variables Validity (cue/prime word validly cued the target side; cue/prime word invalidly cued the target side), ISI (34 ms; 700 ms), Hemisphere (contralateral; ipsilateral), and Block (Block1; Block2 with additional cue/prime discrimination) was computed. It yielded two significant interactions, Validity × Hemisphere, F(1, 19) = 7.23, p = .015, = .28, as well as ISI × Block, F(1, 19) = 4.99, p = .038,
= .21. Post-hoc t tests (one-sided; α = .03, Bonferroni-corrected for two comparisons) examining the difference between contralateral and ipsilateral electrodes (variable Hemisphere) for both valid and invalid trials (variable Validity) showed a marginally significant difference in valid trials, t(19) = −2.44, p = .025, d = −0.53 (contralateral: 1.75 µV, ipsilateral: 2.25 µV), and no significant difference in invalid trials, t(19) = 0.11, p = .911, d = 0.02 (contralateral: 1.83 µV, ipsilateral: 1.81 µV). Post-hoc t tests for the interaction of ISI and Block (two-sided; α = .03, Bonferroni-corrected) revealed a significant difference between long and short ISI in Block 1 trials (only visual search task), t(19) = −4.78, p < .001, d = 1.05 (short: 0.42 µV, long: 3.07 µV), and no difference in Block 2 trials (with additional cue/prime discrimination task), t(19) = 1.85, p = .081, d = 0.40 (short: 1.39 µV, long: 2.76 µV).
A.5. N400
A repeated-measures ANOVA (2 × 2 × 2 × 3 × 4), with the variables Congruence (cue/prime word and target word congruent; cue/prime word and target word incongruent), Block (Block1; Block2 with additional cue/prime discrimination), Cue/Prime Word (left; right), Laterality (left: F1, C1, CP1, P1; middle: Fz, Cz, CPz, Pz; right: F2, C2, CP2, P2), and Caudality (frontal: F1, Fz, F2; central: C1, Cz, C2; centro-partietal: CP1, CPz, CP2; parietal: P1, Pz, P2) was computed. The degrees of freedom were adapted using a Greenhouse-Geisser correction. There was a marginally significant main effect for Congruence, F(1, 19) = 4.39, p = .050, = .19, which reflects a less positive (i.e., more negative) mean amplitude for incongruent than for congruent cue/prime-target word relations (2.89 and 3.86 µV), as well as a main effect for Caudality, F(1.89, 35.92) = 7.51, p = .002,
= .28, which shows generally increasing mean amplitudes from rostral to caudal electrode sites (frontal: 2.45 µV; central: 3.50 µV; centro-parietal: 3.86 µV; parietal: 3.99 µV). No further main effects or interaction effects were significant, all ps > .050, all
s < .15.