
Abstract
Drawing on object-oriented-ontology we aim for a hybrid language that not only processes formal symbols but interacts with the microbes in the speaker’s mouth. We argue that the metaphors historically used to frame the relationship between oral microbiota and speech cannot account for the co-creative material relationship between human speech and microbial, environmental and biotechnological needs. In our performances we harvest oral yeast microbes from the audience. Via a spectrogram, repetitively spoken phonemes drive pattern pumps, which add pheromones to the microbes, the latter which then are faded out. In the microbes, for some replication cycles an ecological adaptation to the individual phonemes persists, affirming – in our definition – some phonemes and deleting others, thereby improving the eco-sphere in the mouth by changing the alphabetical order of the input phonemes. The audience observes the adaptation by respelling words. We propose ‘microbial speech’ as a category between semantic and phonetic meaning, with a transcorporeal mattering between speech and the body as a new ecolinguistic quality. We aim at a language becoming a biological state in order to protect its own ecology. We consider these collective posthuman forms of relationality, subjectivity and biopolitical praxis as multiple ecologies of belonging and a more ethical mode of microbes existing in-common with language, affirming the transversal relations of all living and non-living matter.
In a participatory experiment entitled Microbial Keywording, our inter-disciplinary team explores links between spoken language, biotechnology and single-celled organisms that inhabit the human mouth. The experiment is based on theories of posthumanist performativityFootnote1 that call into question established dichotomies between matter and language, while building upon recent scientific studies on the cellular memory of microbes and their ability to anticipate future events.
Posthumanist views on language and matter, especially those elaborated by feminist scholar Karen Barad (2012) and political theorist Jane Bennett (Citation2010), enable us to consider the materiality of oral microbes as something fundamentally interwoven with, rather than separate from, technologies and our language- built scientific knowledge. Our team endeavoured to build an experimental phonetic set up where oral microbes are being tested and, at the same time, act as a force in itself to transform discourses about them. In the course of performative and experimental actions, keywords from conference lectures on the human microbiome are the starting point for the creation of a new language in which phonetic sound interacts with microbes via biotechnology.Footnote2 At the end of the process the keywords will make an impact twice: first, as meaningful phonemes of a word; second, as an oral medication, a linguistic-probiotic drug that may react on the speaker via well-known cross-talk mechanisms between the brain and the gut, and the altered mouth microbiota (Mayer et al. Citation2014; Gao et al. Citation2018). This last step is a speculative gesture based on our current knowledge of human microbiome– brain interactions. Recent scientific research suggests that single-celled organisms may be capable of memorizing past events (Caudron and Barral Citation2014) and of anticipating future events (Mitchell et al. Citation2009; Mitchell and Lim Citation2016). This research supports the theory that unicellular organisms can adapt to a changing environment such as the pH levels of speakers’ saliva. Based on an experiment on cell memory (Caudron and Barral Citation2013), we developed a set up in which phonemes – the smallest entities of a spoken word – would leave a memory trace on single- celled oral microbes enabling them to memorize and anticipate sound sequences of a word.
Our artistic experiment involves a live audience and aims at developing an understanding that language itself is material. It not only processes formal symbols but interacts directly with the actual agency of the microbes in the speaker’s mouth. Despite the direct oral contact we have with microbes as we speak, thus influencing their behaviour through our speech acts, we are unaware of these relations.Footnote3 In our performance series, we therefore encourage participants to shift their focus from the semantic content of spoken words to the biochemical milieu that speakers create for microbial organisms in their mouth cavity. This shift induces an increased awareness of the entanglement between biological matter and language, and more significantly, in the course of our participatory performances, reveals a better understanding of the general conditions that oral microbes live in and that their evolution and homeostasis may be related to the evolution of language in manifold ways:
Microbial Keywording, AIL Vienna, 2018. Rotating petri dish with injection of saliva harvest, designed by Joseph Knierzinger. © Spiess & Strecker

Oral microorganisms faced drastic fluctuations in their habitats in human mouths when people first started to utter phonemes. Although oral microbes have since then been specifically evolving to adapt to such changes, maximizing their likelihood of survival (Mitchell et al. Citation2009; Mitchell and Lim Citation2016) in the human mouth, recent findings show that microbial diversity has decreased (Moeller et al. Citation2014). This may prompt the assumption that the relationship between speech and oral microbial life is competitive. However, these current findings concerning the simultaneous loss of diversity in languages, phonemes and the microbiome may also be explained by microbial habitat destruction due to neoliberalism’s linguistic globalization and environmental damage (Fill and Penz Citation2017).
To take a critical stance on the neoliberal appropriation of all spheres of life, our performative experiment particularly underlines the connection between the fragility of phonetic diversity in language and oral biodiversity: during early childhood, 90 per cent of the originally present phonetic inventory is lost, and in today’s web language many phonetic utterances are replaced by emoticon-style signs and pictorial shortcuts, which are then reimported non- phonetically into language. Phonemes that are unnecessary for communication undergo evolutionary degeneration, resulting, for instance, in the phoneme H disappearing from numerous languages (Heller-Roazen Citation2005). It seems as if humans want to counteract this loss, for example when they imitate or invent phonemes while communicating with animals, or talking to a person in various states of pain, joy or sexual arousal. For such reasons, phonetic linguists also recommend specific pluri-lingual training of phonemes in order to prevent phoneme extinction (Fill and Penz Citation2017).
In the making of our experimental set up, we addressed the mouth in a synthesis of theory and practice as a performative area of complex ecological conditions and vulnerable psychosomatic balance. The French philosopher, theorist and writer Michel Serres (2016 [2008]) has emphasized the speaking and tasting mouth as a dual organ. Sharing what is in our mouths indicates a most intimate relationship. We experience pleasure and destruction when we speak and eat. Speech and food disappear at the height of pleasure, but on a biological level, the milieu in the mouth shares a unique bond with the outer environment: As a wet body cavity, although a small region, hundreds of endemic species populate its orifice in a particularly high concentration (Gao et al. Citation2018). This microbial density is the link to complex neuronal processes. As a small region, the mouth, the tongue and lips in particular are represented much larger in the human brain compared to other sensory body parts (Roux et al. Citation2018). As such, our mouth microbiota may connect both the brain and our environment at the microbial and neuronal level.
Microbial Keywording, Haus der Kulturen der Welt, Berlin, 2019. Guiding the harvest of sexually replicating Saccharomyces from the audience’s saliva. © Spiess & Strecker

The mouth is also one of the central organs of early literary concepts illustrating where language and materiality may connect: François Rabelais, the Renaissance-era physician and writer, made this relation the subject of his novels. He understood language as a composite, layered experience in a multi-faceted and hybrid reality. Attentive to the general transitions and dissolving orders taking place as feudalism transformed into capitalism, he questioned the relationship of the signifier to the signified: Rabelais’ literary figures used signs strategically, without an orientation towards communication (Ponziano Lavatori Citation1996). In his famous ‘frozen words’ episode, sailors, in spring, heard the screams of a battle that had been frozen during the winter. ‘He threw three or four handfuls of them on the deck; among which I perceived some very sharp words, and some bloody words. When they had been all melted together, we heard a strange noise’ (Rabelais 2008 [1532]: 126).
In the development of our performative experiment, we saw obvious parallels between the way Rabelais freezes signifiers and transforms them poetically into material and the way we ourselves transgressed material with data. Whereas Rabelais’ figures transformed linguistic information into ice, we transformed biological matter into linguistic information and vice versa. To do so, we either maintained the cellular processes of oral microbes running at the same rate as in the living body, or we slowed them down by freezing – a common practice in biotechnology. In this way, we were able to stop or accelerate the time of material processes in accordance with the needs of our dramaturgy. By slowing down biological time, we could even ‘biobank’ microbes and work with the same microbes at any time in future performances (Mitchell et al. Citation2008; Spiess and Strecker Citation2014).
MICROBIAL KEYWORDING: A PARTICIPATORY EXPERIMENT
Imagine attending a three-day symposium in a large gallery. Glass vessels containing fluids sit on an outsize table, connected to cables, tubes and microphones. In the centre, cell cultures multiply in an incubator. The lids of Petri dishes filled with growth media show captions, phonetic characters and numerical values. Rotating devices in front of several seats hold similar Petri dishes, which circulate like vinyls, attached to a tube and a saliva ejector.
Several large screens are mounted on the walls around the audience. Over the course of four days, fifteen guest lecturers contribute their thoughts about artistic methods and scientific theories, looking at the microdynamics between living matter, technology, culture and economics. Symposium topics involve and reflect upon the agency of genes or cell fragments, proteins or enzymes, bacteria and fungi and how they relate to the macroscopic dynamics of power in contemporary biopolitics. The presentation format of the symposium alternates between lectures, performances, and the ongoing experimentation of this performance designated ‘Microbial Keywording’. To begin the experiment, each lecturer proposes a key term, for instance, extimacy.Footnote4
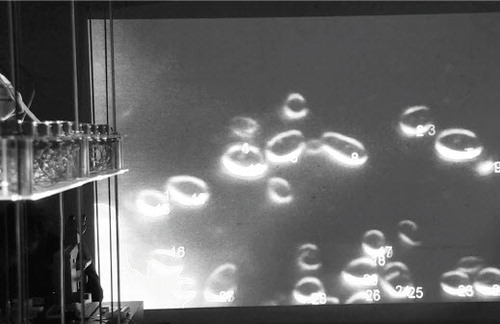
(right) Microbial Keywording, Ars Electronica, Linz, 2019. Blob Detection determines Replication– Apoptosis Ratio after Exposition. © Spiess & Strecker
Step 1: Changing the milieu
A small group of symposium guests encounters the material relationship between the selected keyword, its phonetic units and oral microbial cultures. Experimenters and technical assistants first facilitate identifying the keyword’s phonemes. The word extimacy is articulated as ekst m si. The experimenters perform a short phonological awareness exercise on the production of sounds, involving the tongue, palate, alveoli, teeth and breath, encouraging participants to focus on frequencies, vibrations and rhythms, rather than on meanings.
Chosen phonemes of the word extimacy, such as 'e k s t m s i, each affect the flow of saliva differently. Measuring the acidity level in the mouth with pH sensors before and after the repetition exercise, technical assistants evaluate changes in the milieu and their influence on specific microbes. Speakers’ saliva is thereby considered both a prerequisite of their speech and of the oral environment that their oral microbes are living in. Repetitive articulation may cause the pH value of donor saliva to become more acidic or more alkaline, triggering, as a consequence, particular types of microbes to adapt, with their fertility mode switching from asexual to sexual replication.Footnote5 Due to their adaptive drive to replicate sexually, these microbes become much more sensitive to pheromones. This characteristic will be relevant in later stages of the experiment. Not every repeated phoneme produces a change in the pH level of the saliva – inducing an alteration of the microbes’ mating behaviour. Thus, the team chooses to harvest the saliva of participants whose uttered phonemes have the desired effect. With a sterile saliva ejector, saliva is drawn into a rotating Petri dish by a vacuum pump.
Step 2: Adapting microbes
In the next step, the team staggers the biological process, alternating between freezing and unfreezing microbial cultures harvested prior from participants or by utilizing previously prepared vessels and bioreactors in which particular microbial alterations already occur. Since the growth and replication time of the microbes is slower than that of the staged performance, the team had also created a digital programme with moving data, charts and microscopic images for the audience to better visualize the biological cycles and compensate for the time gap.Footnote6
Microbial Keywording, AIL Vienna, 2018. Oleg Soulimenko performing with saliva ejector. © Spiess & Strecker

The team then transfers sensitized strains from the Petri dish to the first customized bioreactors. Participants and attendees gather around the conference table, asked to recall words from the previous lecture. Someone volunteers to pronounce the words ‘black box’, for example, another one ‘species’, and the last, ‘membrane’. Again, the keywords are broken down into their smallest sound units: b l æ k 'b a k s, 's p i : i : z, 'm ԑ m'b r e I n.
The volunteer repeats these phonemes, amplifying them in the gallery space over the microphone as other participants join in. Speed, volume and frequencies of the audience chanting the respective phonemes that are now emerging are interconnected with a microfluidic pump via voice spectrograms. In this way the collective voicescapes control the concentration and pressure applied to the flow of industrial pheromones into the bioreactor.Footnote7 These signalling molecules, however, come without what they purport to signify – in our case: mates. Because earlier in the experiment the cells’ drive to replicate sexually was increased, these microbes are now easily stimulated by the added pheromones. Multiple responses to this stimulation can be triggered: arrest of cell cycle, decrease in budding index, formation of cellular bulges, or apoptosis (Severin 1997). A visual map of pheromone/spectogram input and cell reactions shows participants how varying combined values can result in either wide microbial cell suicide or famine due to overpopulation, while other combinations are suitable to create a state of homeostasis – the condition for optimal performance of this particular group of oral microbes.Footnote8 Apoptosis, or programmed cell death, in single-celled organisms is a normal ecological response in a wide variety of physiological settings, where its role is to respond to environmental stimuli, to balance homeostatically between cell replication and cell death and to remove damaged cells (Gourlay et al. Citation2006). The resulting ratio of replication versus death indicates an ecological adaptation to each of the individual phonemes. In search of microbial colonies and phonemes that maintain an interdependent system of steady internal, physical and chemical relations, the team then collects the data of every processed phoneme, archiving the respective microbial colonies.
After repeating this procedure several times and over the course of multiple days, the cell cultures learn to anticipate the fluctuation of pheromones in close connection with the respective phonetic frequencies (Mitchell et al. Citation2009; Mitchell and Lim Citation2016). On the screen it can be seen that after the pheromones have been blocked the anticipation remains and microbes adapt to certain phonemes. Such anticipatory behaviour may be compared to Pavlovian conditioning in mammals. Footnote9 But we also know that such behaviour may cause epigenetic adaptation: cells that acquire certain behaviour form new protein clusters and even pass them on to next generations, independently from the input stimulus (Caudron and Barral Citation2014).
Step 3: Back to the oral cavity
Now it can be extrapolated that a development similar to that in the bioreactor in vitro is also conceivable in vivo, in the oral cavity itself. We conclude that the changing chemical composition of saliva, its concentration and flow – in short, the changes in the microclimate in the mouth – may also cause a conditioning between phonemes and microbes. We also identify the possibility that microbes from our experiment would inherit their memory of interactions with phoneme patterns. Because microbes are great communicators and pass on cellular memory through generations of reproduction cycles (Caudron and Barral Citation2014), we were challenged to reverse the process and reintroduce memories from the performance to the speaker’s mouth. The adapted oral microbes were actually given back to the participators as a ‘linguistic probiotic’, contributing to an improved milieu in the oral cavity.
(left) Microbial Keywording, Ars Electronica, Linz, 2019. Probiotic mouth spray labelled with the respective affirmed phoneme. © Spiess & Strecker

We approached the connection between speech acts, microbes and the ‘phonetic probiotic’ produced from various viewpoints.
Probiotics are living microorganisms promoted with claims that their consumption provides health benefits by improving or restoring the microbial flora (Abbasi Citation2019; Dassi et al. Citation2018). The concluding act of our experiment aims to create a drug that may influence the recipient not only via their direct oral ecology but also via the human microbiome–brain axis, by promoting a state of homeostasis in the speaker’s mouth.Footnote10 An increasing number of scientific articles propose intriguing data on the topic, employing terms such as ‘psychobiotic’ or ‘melancholic’ microbes (Cryan and Dinan 2012), suggesting humans are merely a vehicle for the 100 trillion microorganisms living inside them. Another intriguing hypothesis investigates microbes that manipulate their hosts, giving an evolutionary advantage to those microorganisms by encouraging human hosts to perform certain rituals that favour microbial transmission (Panchin et al. Citation2014).
We consider data that prove microbe–brain interactions as key drivers in the evolution of the social brain. Experimental changes to the microbiome show effects on emotional behaviour and related brain systems (Zaura and Mira Citation2015; Montiel-Castro et al. Citation2013; Sarkar et al. Citation2016). Thus, our participants’ microbiome is part of their intestine–brain axis, and as such mutually connects the affective and cognitive level of the participants’ speech with their oral microbes. Studies of faecal transplantation even show that bacteria may transfer certain behavioural characteristics, such as depression or addiction, from one person to another (de Groot et al. Citation2017; Evrensel and Ceylan Citation2016), giving a good reason for the application of microbial diversity by probiotics.
Another compound that provides a rational for our probiotic is that the oral microbial habitat is vulnerable to destructive impacts, such as high calorie intake, antibiotics, hygiene, fertilizer and exsiccation, and as such represents the general dependency of oral microbial biodiversity on cultural behaviour (Zaura and Mira Citation2015; Gao et al. Citation2018; Dassi et al. Citation2018). Today such cultural impacts have decreased the diversity of the human microbiome to 20 per cent in urban areas compared to native rural communities (Clemente et al. Citation2015). From that perspective, it is considered relevant to restore the microbial flora in diversifying the microbes by administering relevant strains.
An additional argument may be that speech acts are often linked to rituals or other cultural human interactions that favour microbial transmission and evolution (Panchin et al. Citation2014). While reciting psalms, prayers or mantras, words and phonemes are repeated by multiple people before kissing a sacred relic such as a cross, holy stone, book or painting. We assume that a reduction of such speech-act-related rituals in turn results in a loss of vertical microbial transmission, as each successive generation has fewer types of organism to pass on to the next.
Thinking of diversity against such a wide background has finally led to a paradigm shift in psychopharmacology, seeing the therapeutic potential in microbial-based behaviour modification (Mayer et al. Citation2014). Thus, the microbial drug we produced as the result of artistic practice and its related research aims to explore alternative values and forms of therapeutic designer microbiomes and to look at them from a broader bio-political and eco- linguistic perspective. In the manner of critical and speculative design the ‘linguistic probiotic’ we introduce in our performance gives way to playful research into how speech acts may be entangled with the human body and its non- human inhabitants.
Again, concluding from the perspective of performance studies, we understand our artistic work as turning J. L. Austin’s concept of the speech act (Austin Citation1975 [1962]) and his interest in the performative aspects and effects of language towards its ultimate materialization.Footnote11 Because the dynamic and agential potential of materiality of arbitrary systems have been much ignored, research has lately been presented that foregrounds the material aspects of language (De Freitas and Sinclair Citation2014; Cavanaugh and Shankar Citation2017). With our experiment, we also interpret Karen Barad, who opposed Austin by developing a theory of an agency of matter, which moves away from granting language only productive effects and radically decentres human language as a legislator of truth. Likewise, we recognize that non-human factors are involved in the creation of our discursive belief systems; in conditioning microbes by speech we aim for a more direct relationship between language and oral symbiosis and dysbiosis. As performance artists and biologists, we challenge a simplistic semiotic view that defines language as merely symbolic, as an arbitrary relationship the signifier (sound pattern of a word) has with the signified (the concept behind the word). Focusing instead on the indexical, material and interventional relation of phonemes and microbes with less respect to language’s arbitrariness, by Microbial KeywordingFootnote12 we rather set out to rethink and enact materiality and signification.
ACKNOWLEDGEMENTS
Funded by the Medical University of Vienna, the Austrian Science Fund (FWF) V501-G24 and the University of Applied Arts Vienna.
Supported by Ars Electronica, Linz, Angewandte Innovation Lab Vienna, brut Vienna, Haus der Kulturen der Welt, Berlin and the University of Salzburg, Department of Biosciences.
Notes
1 In such a theoretical approach the operational power of performativity is not simply based on language as doing, but on doing as materialization (Barad Citation2003). This includes the assumption that matter and language are neither exclusively given nor solely an effect of pure human action.
2 Klaus Spiess and Lucie Strecker’s Microbial Keywording performance first took place within the context of the festival and symposium ‘Applied Microperformativity: Live arts for a radical socio-economic turn’ held at the University of Applied Arts Innovation Lab, Vienna 2018, https://bit.ly/2CP51ck
3 We aim to not foreground linguistic models of speech acts, as developed by the British philosopher J. L. Austin, but models of posthumanist performativity. Therefore, we consider speech acts here as flow of saliva, its changes in acidity, tongue friction on teeth, palate and mucous membrane or air pressure.
4 Extimacy describes a very intimate connection between subject and object, which nevertheless remains external. Jacques Lacan (Citation1986 [1959/60]), who coined the term, considered the voice as an element with which the subject is connected by means of extimacy.
5 We decided to work with eucaryotic cells of the Saccharomyces boulardii subtype of yeast (Sen and Mansell Citation2020), mouth fungi with probiotic characteristics that represent less than 1 per cent of the oral mycobiome, yet is characterized by its extraordinary ability to both link environment and brain and to memorize stress.
6 We used the Python Programming Language, Pure Data visual programming language and speech-to-text- recognition software for programming. This programmed predictive network was developed based on data from our previous performances (saliva acidity, replication and apoptosis rates, spectrograms, phonemes). A logistic equation was adopted to model how oral microbes and linguistic fields interacted. Despite trivial denominations of data, the programmed network aimed at non-trivial modelling with hysteresis and memory as an internal part of programming itself.
7 Pheromones are secreted or excreted chemical factors that trigger a social response. They can also be purchased as engineered products from biotech companies. We ordered them from Genscript: https://bit.ly/3aHtDjs
8 We adapted Lindeberg’s (1993) software to measure the death-related shrinkage and replication- related bulging of the cells.
9 We hypothesize that the pathway that was triggered in the later phase of our experiment is based on weak cross-modal correspondences (Suslick Citation2012) between pheromones, phonemes and the anticipated homeostasis based on cellular pattern recognition (Trdá et al. Citation2015). However, one has to consider that what we performed was less a controlled experiment to test a hypothesis than a weighted manipulation of conditions in order to generate a desired artistic outcome.
10 In an early version of the performance with a slightly different concept, a drug was administered as a rectal suppository (see also K. T. Zakravsky’s article in this issue that foregrounds a non- material humanist critics of the microbial process involved). In this version, we filled small bottles with microbial cultures, labelling them with the corresponding letter of the phonetic alphabet. Due to an unknown medical risk–benefit ratio and safety of the application, we did not invite the audience to consume. The performance can be considered as a phase 0 study, which would have to go through phases I to IV before becoming an approved medical treatment.
11 The speech act theory of J. L. Austin was later further developed by American philosopher John Rogers Searle (1969), devising a system of speech act categorization. Utterances are considered not only to convey information but also to perform actions at different levels.
12 Watch videos of the project on the PR YouTube channel and on Vimeo: Angewandte Innovation Lab December 2018 https://bit.ly/2Ri2A5K  Microbial Keywording. Animation https://bit.ly/2QTqILK
Microbial Keywording. Animation https://bit.ly/2QTqILK  Microbial Keywording at Ars Electronica https://bit.ly/3iAdRdH
Microbial Keywording at Ars Electronica https://bit.ly/3iAdRdH
REFERENCES
- Abbasi, Jennifer (2019) ‘Are probiotics money down the toilet? Or worse?’ Journal of the American Medical Association JAMA 321(7): 633–5. doi: 10.1001/jama.2018.20798
- Austin, J. L. (1975 [1962]) in James Opie Urmson and Maria Sbisà (eds) How to do Things with Words. The William James Lectures delivered in Harvard University in 1955, Oxford: Clarendon Press.
- Barad, Karen (2003) ‘Posthumanist performativity: Toward an understanding of how matter comes to matter’, Signs 28(3): 801–31. doi: 10.1086/345321
- Bennett, Jane (2010) Vibrant Matter. A political ecology of things, London: Duke University Press.
- Caudron, Fabrice and Barral, Yves (2013) ‘A super-assembly of Whi3 encodes memory of deceptive encounters by single cells during yeast courtship’, Cell 5, 155 (6): 1,244–57.
- Caudron, Fabrice and Barral, Yves (2014) ‘Mnemons: Encoding memory by protein super-assembly’, Microbial Cell 1(3): 100–2. doi: 10.15698/mic2014.01.134
- Cavanaugh, Jillian R. and Shalini Shankar, eds (2017) Language and Materiality, New York: Cambridge University Press.
- Clemente, Jose C., Pehrsson, Erica C., Blaser, Martin J., Sandhu, Kuldip, Gao, Zhan, Wang, Bin, Magris, Magda, Hidalgo, Gilda, Contreras, Monica, Noya-Alarcón, Óscar, Lander, Orlana, McDonald, Jeremy, Cox, Mike, Walter, Jens, Lyn Oh, Phaik, Ruiz, Jean F., Rodriguez, Selena, Shen, Nan, Song, Se Jin, Metcalf, Jessika, Knight, Robert, Dantas, Guatam and Dominguez-Bello, M. Gloria (2015) ‘The microbiome of uncontacted Amerindians’, Science Advances 3;1(3) e1500183. doi: 10.1126/sciadv.1500183
- Dassi, Erik, Ferretti, Pamela, Covello, Giuseppina, Bertorelli, Roberto, Denti, Michaela A., De Sanctis, Veronica, Tett, Adrian and De Segata, Veronica (2018) ‘The short-term impact of probiotic consumption on the oral cavity microbiome’, Scientific Reports 8: 10,476.
- De Freitas, Elizabeth and Sinclair, Natalie (2014) Mathematics and the Body. Material Entanglement in the Classroom, New York: Cambridge University Press.
- De Groot, Pieter F., Myrthe, N., Frissen, M. N., de Clercq, Nicolien C. and Nieuwdorp, Max (2017) ‘Fecal microbiota transplantation in metabolic syndrome: History, present and future’, Gut Microbes 4;8(3): 253–67. doi: 10.1080/19490976.2017.1293224
- Evrensel, Alper and Ceylan, Mehmet Emin (2016) ‘Fecal microbiota transplantation and its usage in neuropsychiatric disorders’, Clinical Psychopharmacolgy and Neuroscience 31;14(3): 231–7. doi: 10.9758/cpn.2016.14.3.231
- Fill, Alwin F. and Penz, Hermine (2017) The Routledge Handbook of Ecolinguistics, New York, NY: Routledge.
- Gao, Lu, Xu T., Tiansong, Huang, Gang, Jiang, Song, Gu, Yan and Chen, Feng (2018) ‘Oral microbiomes: More and more importance in oral cavity and whole body’, Protein Cell 9(5): 488–500. doi: 10.1007/s13238-018-0548-1
- Gourlay, Campbell W., Du, Wei and Ayscough, Kathryn R. (2006) ‘Apoptosis in yeast-mechanisms and benefits to a unicellular organism’, Molecular Microbiology 62(6): 1,515–21. doi: 10.1111/j.1365-2958.2006.05486.x
- Heller-Roazen, Daniel (2005) Echolalias – On the forgetting of language, Cambridge: MIT Press.
- Lacan, Jacques (1986 [1959/60]) in Jacques Alain-Miller (ed.) Le séminaire. Livre VII. L’éthique de la psychoanalyse, Paris: Seuil.
- Lindeberg, Tony (1993) ‘Detecting salient blob-like image structures and their scales with a scale-space primal sketch: A method for focus-of-attention’, International Journal of Computer Vision 11(3): 283–318. doi: 10.1007/BF01469346
- Mayer, Emeran A., Knight, Rob, Mazmanian, Sarkis K., Cryan, John F. and Tillisch, Kirsten (2014) ‘Gut microbes and the brain: Paradigm shift in neuroscience’, The Journal of Neuroscience 34(46): 15,490–6. doi: 10.1523/JNEUROSCI.3299-14.2014
- Mitchell, Amir and Lim, Wendell (2016) ‘Cellular perception and misperception: Internal models for decision-making shaped by evolutionary experience’, BioEssays 38(9): 845–9. doi: 10.1002/bies.201600090
- Mitchell, Robert, Burgess, Helen J. and Thurtle, Phillip (2008) Biofutures: Owning body parts and information, DVD-ROM: University of Pennsylvania Press.
- Mitchell, Amir, Romano, Gal H., Groisman, Bella, Yona, Avihu, Dekel, Erez, Kupiec, Martin, Dahan, Orna and Pilpel, Yitzhak (2009) ‘Adaptive prediction of environmental changes by microorganisms’, Nature 9;460(7,252): 220–4. doi: 10.1038/nature08112
- Moeller, Andrew H., Li, Yingying, Ngole, Eitel Mqoudi, Ahuka-Mundeke, Steven, Lonsdorf, Elizabeth V., Pusey, Anne E., Peeters, Martine, Hahn, Beatrice H. and Ochman, Howard (2014) ‘Rapid changes in the gut microbiome during human evolution’, PNAS 111(46): 16,431–5. doi: 10.1073/pnas.1419136111
- Montiel-Castro, Augusto J., González-Cervantes, Rina M., Bravo-Ruiseco, Gabriela and Pacheco-López, Gustavo (2013) ‘The microbiota–gut–brain axis: Neurobehavioral correlates, health and sociality’, Frontier in Integrative Neuroscience 7(7): 70.
- Panchin, Alexander Y., Tuzhikov, Alexander I. and Panchin, Yuri V. (2014) ‘Midichlorians – the biomeme hypothesis: Is there a microbial component to religious rituals?’, Biology Direct 9: 14. doi: 10.1186/1745-6150-9-14
- Ponziano Lavatori, Gerard (1996) Language and Money in Rabelais, New York, NY: Peter Lang Publishing.
- Rabelais, François (2008 [1532]) Gargantua and Pantagruel, Moscow: Dodo Press.
- Roux, Franck-Emmanuel, Djidjeli, Imène and Durand, Jean-Baptiste (2018) ‘Functional architecture of the somatosensory homunculus detected by electrostimulation’, The Journal of Physiology 596(5): 941–56. DOI: 10.1113/JP275243 . Epub 2018 Jan 19.
- Sarkar, Amar, Lehto, Soili M., Harty, Siobhán, Dinan, Timothy G., Cryan, John F. and Burnet, Philip W. J. (2016) ‘Psychobiotics and the manipulation of bacteria–gut–brain signals’, Trends in Neuroscience 39(11): 763–81. doi: 10.1016/j.tins.2016.09.002
- Searl, John Rogers (1969) Speech Acts: An essay in the philosophy of languae, Cambridge: Cambridge University Press
- Sen, Swastik and Mansell, Thomas J. (2020) ‘Yeasts as probiotics: Mechanisms, outcomes, and future potential’, Fungal Genetics Biology 137: 103333. DOI: 10.1016/j.fgb.2020.103333. [Epub ahead of print.]
- Serres, Michel (2016 [2008]) The Five Senses: A philosophy of mingled bodies, London: Bloomsbury Revelations.
- Spiess, Klaus and Strecker, Lucie (2014) ‘Spitparty: DNA imaging and aesthetic mimicry’, Performance Research 19(4): 38–44. doi: 10.1080/13528165.2014.947136
- Suslick, Kenneth S. (2012) ‘Synesthesia in science and technology: More than making the unseen visible’, Current Opinion in Chemical Biology 16(5–6): 557–63. doi: 10.1016/j.cbpa.2012.10.030
- Trdá, Lucie, Boutrot, Freddy, Claverie, Justine, Brulé, Daphnée, Dorey, Stephan and Poinssot, Benoit (2015) ‘Perception of pathogenic or beneficial bacteria and their evasion of host immunity: Pattern recognition receptors in the frontline’, Frontiers in Plant Science. DOI: 10.3389/fpls.2015.00219
- Zaura, Egija and Mira, Alex (2015) ‘The oral microbiome in an ecological perspective’, Frontiers in Cellular and Infection Microbiology, https://bit.ly/34jrUzY , 29 April, accessed 20 February 2020.