
ABSTRACT
DIRAS family GTPase 1 (DIRAS1) has been reported as a potential tumor suppressor in other human cancer. However, its expression pattern and role in cervical cancer remain unknown. Knockdown of DIRAS1 significantly promoted the proliferation, growth, migration, and invasion of C33A and SiHa cells cultured in vitro. Overexpression of DIRAS1 significantly inhibited the viability and motility of C33A and SiHa cells. Compared with normal cervical tissues, DIRAS1 mRNA levels were significantly lower in cervical cancer tissues. DIRAS1 protein expression was also significantly reduced in cervical cancer tissues compared with para-cancerous tissues. In addition, DIRAS1 expression level in tumor tissues was significantly negatively correlated with the pathological grades of cervical cancer patients. DNA methylation inhibitor (5-Azacytidine) and histone deacetylation inhibitor (SAHA) resulted in a significant increase in DIRAS1 mRNA levels in C33A and SiHa cells, but did not affect DIRAS1 protein levels. FTO inhibitor (FB23–2) significantly down-regulated intracellular DIRAS1 mRNA levels, but significantly up-regulated DIRAS1 protein levels. Moreover, the down-regulation of METTL3 and METTL14 expression significantly inhibited DIRAS1 protein expression, whereas the down-regulation of FTO and ALKBH5 expression significantly increased DIRAS1 protein expression. In conclusion, DIRAS1 exerts a significant anti-oncogenic function and its expression is significantly downregulated in cervical cancer cells. The m6A modification may be a key mechanism to regulate DIRAS1 mRNA stability and protein translation efficiency in cervical cancer.
Introduction
Cervical cancer is considered almost entirely preventable because of highly effective primary (HPV vaccine) and secondary (screening) prevention measures.Citation1 In 2018, the Director-General of the World Health Organization (WHO) called for global action to eliminate cervical cancer (≤4 per 100,000 women globally) through a triple-intervention strategy, that is projected to be achieved in the next century.Citation2,Citation3 However, until this goal is achieved, there will still be many women with advanced cervical cancer who have limited treatment options.Citation4 Exploring potential targeted therapies holds promise for the treatment of persistent, recurrent and metastatic cervical cancer.Citation4
DIRAS family GTPase 1 (DIRAS1), also known as Rig (Ras-related inhibitor of cell growth), is a member of the small GTPase Ras superfamily.Citation5 The DIRAS1 gene, located on chromosome 19p13.3, consists of 2 exons and encodes a protein of 198 amino acids. Unlike common oncogenic small GTPases (e.g., Ras or Rho family members), DIRAS1 has been reported as a potential tumor suppressor in human renal cell carcinoma,Citation6 ovarian cancer,Citation7 colorectal cancer,Citation8 gliomas,Citation9 and esophageal squamous cell carcinoma.Citation10 However, its expression pattern and role in cervical cancer remain unknown.
Epigenetic regulation mainly includes DNA methylation, histone modification and RNA modification, which are responsible for regulating gene expression and involved in normal biological functions and disease progression.Citation11 Among them, DNA methylation is closely related to cancer progression, and mainly refers to the hypermethylation of CpG island in the promoter of key anti-oncogenes in order to interfere with the reading of DNA information, thus silencing the expression of key anti-oncogenes.Citation12 Different histone marks determine the state of chromatin as well as the transcriptional activity of genes, both oncogenes and anti-oncogenes.Citation13 N6-methyladenosine (m6A) RNA modification is the most abundant internal modification in eukaryotic mRNAs, regulated by methyltransferases (METTL3/METTL14) and demethylases (FTO and ALKBH5), and recognized by a set of “readers”, which decode the m6A and further regulate the degradation, stability, translation initiation, and translation efficiency of the modified mRNAs.Citation14
In summary, this study will examine the anti-cancer function of DIRAS1 and its unique expression pattern in cervical cancer, and analyze the epigenetic regulatory mechanisms underlying the down-regulation of its expression.
Materials and methods
Cell lines and culture
Human cervical cancer cell lines (C33A and SiHa) were purchased from ATCC, and cultured in DMEM (KGM12800N–500, KeyGEN BioTECH) supplemented with 10% FBS (A31608, Gibco), 100 U/mL penicillin and 100 µg/mL streptomycin (KGY0023, KeyGEN BioTECH) in an incubator at 37°C with 5% CO2. 5-Azacytidine (5-aza-dC) (HY-10586)、Synonyms (SAHA) (HY-10221) and FB23–2 (HY-127103) were purchased from MCE.
Plasmids and transfection
In loss-of-function experiments, the pLKO.1-puro empty vector (SHC001, Sigma-Aldrich) was used as a negative control (NC), and the shRNAs targeting sequences as shown in were designed and synthesized by Sigma-Aldrich. In the gain-of-function experiments, pcDNA3.1 empty vector (VT1001, YouBio) was used as the NC, and human DIRAS1 (NM_145173) cDNA sequence was cloned into pcDNA3.1 vector to constitute a recombinant expression plasmid. All plasmids were transiently transfected into cells using Lipofectamine 2000 (11668019, Invitrogen) according to the manufacturer’s instructions.
Table 1. shRnas targeting sequences.
Protein preparation and western blot
Cells were fully lysed with ice-cold strong RIPA lysate (CW2333S, CWBIO) on ice for 10 min. Cell lysates were centrifuged at 12,000 g for 10 min at 4°C to remove impurities. The supernatants after centrifugation were the protein solutions, and protein concentrations were quantified by BCA protein assay kit (CW0014S, CWBIO). Equal amounts of proteins from each group were electroblotted and separated by SDS-PAGE and electrotransferred onto PVDF membranes (0.45 μm, IPVH00010; 0.2 μm, ISEQ00010; Millipore). After blocking with 5% skimmed milk, the membranes were incubated overnight at 4°C with primary antibodies including DIRAS1 (ab65139, Abcam), METTL3 (ab195352), METTL14 (ab220030), ALKBH5 (ab195377), FTO (ab126605) and GAPDH (ab8245). Subsequently, the membranes were incubated with the corresponding secondary antibody at room temperature for 2 h. The immunoreactive bands were visualized by enhanced chemiluminescence (RPN2105, Amersham), and the gray values of the bands were read by Image J software.
Cell counting kit-8 (CCK-8) assay
Cells were implanted into a 96-well plate at a density of 5 × 103 cells/well 24 h after transfection, and cultured overnight. Cell viability was determined at 0 h, 24 h, 48 h, and 72 h of culture using CCK-8 (PF00004, PTG). The specific operations were as follows. Aspirate off the old culture medium and rinse the cells twice with PBS. 10% CCK-8 reagent (10 μL CCK-8 + 90 μL medium) was added back into the cell culture wells and incubation was continued for 2 h in the incubator. At the end of the incubation, the absorbance (OD) value of each well solution at 450 nm was measured using a microplate reader. The curve was plotted with time as the X axis and OD value as the Y axis.
Colony formation assay
After 24 h of transfection, cells were seeded in a 6-well plate at 500 cells per well and cultured for 2 weeks. At the end of the culture, the cells were fixed with 75% ethanol for 30 min and stained with 0.2% crystal violet. Cell clones were then observed and counted.
Transwell assay
5 × 105 cells after 24 h of transfection were inoculated in the upper chamber of the Transwell. The Transwell was placed in a 24-well plate, and 100 µL of FBS-free medium was added to the upper chamber, and 600 µL of complete medium with 10% FBS was added to the lower chamber to induce cell migration or invasion. In addition, the Transwell membrane needs to be coated with Matrigel in the invasion assay. After 24 h of culture, cells migrating or invading under the membrane of the chamber were fixed with 4% paraformaldehyde for 15 min, and stained with crystal violet. Finally, the cells were counted and photographed under a microscope.
RT-qCR
Cells were lysed using Trizol reagent, and total cellular RNA was extracted using RNA Extraction Kit (CW0581S, CWBIO). RNA was reverse transcribed to cDNA using the PrimeScript RT Master Mix (CW2569M, CWBIO). RT-PCR was performed with an SYBR Green master mix reagent (CW0957M, CWBIO). Β-actin was used as an internal reference, and relative gene expression was calculated using the 2−ΔΔCT method. The primer sequences used were designed and synthesized by ORIGENE as follows: DIRAS1 F 5’-CCTTCATCCTGGTGTTCTCCGT-3’, R 5’-CTGCGTCTCATCGCACTTGTTG-3’; β-actin F 5’-CACCATTGGCAATGAGCGGTTC-3’, R 5’-AGGTCTTTGCGGATGTCCACGT-3’.
Tissue microarray and immunohistochemical (IHC) analysis
Human cervical cancer tissue microarrays (ZL-Utr961) were obtained from Shanghai Biotechnology (Shanghai, China). Tissue sections were dewaxed and rehydrated by Van-Clear eco-friendly transparency (Shanghai Hongz Industrial Co., Ltd.) and gradient ethanol, then immersed in citrate buffer and microwaved for antigen repair. After sealing with 5% goat serum solution for 1 h at room temperature, the sections were incubated with anti-DIRAS1 solution (1:200; 12634–1-AP, Proteintech) overnight at 4°C, followed by incubation with goat anti-mouse/rabbit IgG polymer. Immunopositive staining on the sections was visualized with DAB chromogenic solution (DAB-2031, Fuzhou Maixin Biotechnology Development Co., Ltd.). Then, the sections were restained with hematoxylin solution and 1% hydrochloric acid solution. After dehydration with gradient ethanol and Van-Clear transparency, the sections were sealed with neutral gum and coverslips. The results were visualized with a microscopy and photographed (magnification 100× and 400×, respectively). All image processing was done using Image Pro Plus 6.0 software.
Results were scored at 400× field of view with 3–5 fields of view selected for each sample. Scoring was done independently by 3 experienced pathologists and the final results were combined by the 3 experimenters. The scale was based on staining intensity x staining area (0–9 points). The intensity of staining was categorized into 0–3 points: no staining (0 points), mild staining (1 point), moderate staining (2 points), and strong staining (3 points). The area occupied by positively stained cells was categorized into 0–3: unstained (0 points), <33% positively stained cells (1 point), 33–66% positively stained cells (2 points), >66% positively stained cells (3 points).
Statistical analysis
The results of three independent replicate experiments were analyzed using SPSS 22.0 software, and presented as mean ± SD. Differences between the two groups were compared using the Student’s t-test, and a difference of p < .05 was considered statistically significant.
Results
Knockdown of DIRAS1 significantly promotes proliferation, growth and motility of C33A and SiHa cells in vitro
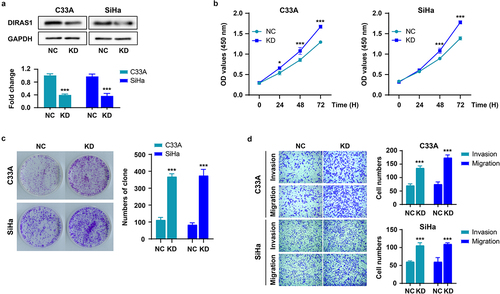
Firstly, DIRAS1 expression was significantly knocked down (KD) in C33A and SiHa cells cultured in vitro using targeted shRNA plasmid transfection (). Next, the effects of DIRAS1 knockdown on cell proliferation, growth, and motility were detected using CCK-8, clone formation and Transwell assays, respectively. As shown in , after 48–72 h of culture, the OD values at 450 nm of the KD group were significantly higher than that of the NC group, indicating that the cell number in the KD group was significantly greater than that in the NC group. In addition, the number and size of clones formed in the KD group were significantly higher than those in the NC group (). These data suggest that knockdown of DIRAS1 significantly promoted the proliferation and growth of C33A and SiHa cells in vitro. Moreover, the number of cells crossing the membrane (and Matrigel) and accomplishing migration or invasion was significantly greater in the KD group than in the NC group, indicating that DIRAS1 knockdown significantly promoted the motility of C33A and SiHa cells in vitro ().
Figure 1. Knockdown of DIRAS1 significantly promotes proliferation, growth and motility of C33A and SiHa cells.
DIRAS1 overexpression significantly inhibits the proliferation, growth and motility of C33A and SiHa cells in vitro
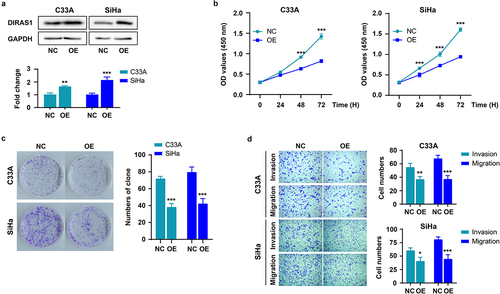
Along the lines of , the expression of DIRAS1 was significantly up-regulated in C33A and SiHa cells using expression plasmid transfection (), and the effects of its overexpression (OE) on cell proliferation, growth, and motility were further examined. As shown in , overexpression of DIRAS1 significantly inhibited the proliferation, growth and motility of C33A and SiHa cells.
Figure 2. DIRAS1 overexpression significantly inhibits the proliferation, growth and motility of C33A and SiHa cells.
Altered expression pattern of DIRAS1 in human cervical cancer tissues
We analyzed RNA-Seq data of cervical squamous cell carcinoma and endocervical adenocarcinoma (CESC)-related tissues from the TNMplot database (tnmplot.com), and found that DIRAS1 mRNA levels were significantly reduced in CESC tissues (N = 304), compared to normal cervical tissues (). This was despite the fact that there were only 3 cases of normal cervical tissues and only 2 cases of CESC metastatic tissues (). Therefore, we analyzed DIRAS1 mRNA levels in CESC tissues again using the GEPIA database (gepia.cancer-pku.cn), which contains 306 cases of CESC tissues and 13 cases of normal cervical tissues from TCGA and GTEx. Consistent with the results of the data analysis of TNMplot, the results of data analysis from GEPIA also showed significantly lower DIRAS1 mRNA levels in CESC tissues (N = 306) compared to normal cervical tissues (N = 13) ().
Figure 3. Altered expression pattern of DIRAS1 in cervical cancer tissues.
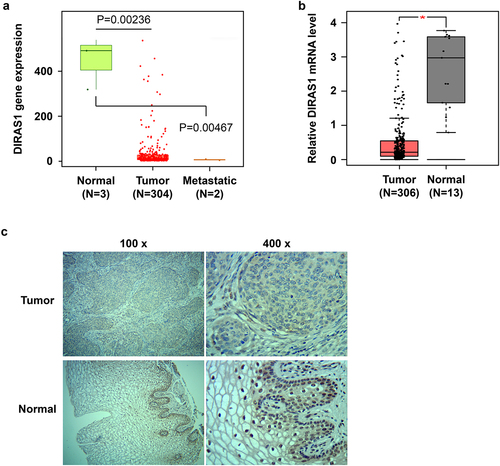
The results of IHC analysis performed on tumor tissues and para-cancerous tissues from 48 clinically collected patients with cervical cancer showed that, consistent with the mRNA levels, DIRAS1 protein expression was also significantly reduced in cervical cancer tissues compared to para-carcinoma tissues ( and ). In addition, in para-carcinoma tissues, DIRAS1 protein showed significant nucleus accumulation (). In contrast, in cervical cancer tissues, DIRAS1 protein expression was not only significantly reduced, but the nucleus accumulation also disappeared significantly (). Furthermore, the expression level of DIRAS1 in tumor tissues was significantly negatively correlated with the pathological grade of cervical cancer patients ().
Table 2. DIRAS1 expression in cervical cancer tissues compared with para-carcinoma tissues.
Table 3. DIRAS1 expression associated with the clinicopathological parameters in CCA.
M6A modification promotes DIRAS1 translation
Given the anti-oncogenic role of DIRAS1 and its specific expression downregulation in cervical cancer cells, we further explored the major epigenetic mechanisms that regulate the downregulation of DIRAS1 expression in cancer cells. The specific DNA methylation inhibitor 5-aza-dC (2 µM for 48 h) as well as the histone deacetylase inhibitor SAHA (1 µM for 48 h) were used to treat the cells. Their applications both resulted in a significant increase in DIRAS1 mRNA levels in C33A and SiHa cells (). Based on this result, we hypothesized that both DNA methylation and histone acetylation might be involved in the transcriptional regulation of DIRAS1 in cancer cells, with DNA methylation hindering DIRAS1 transcription and histone acetylation promoting DIRAS1 transcription. However, the results of western blot showed that the application of 5-aza-dC and SAHA did not significantly affect the intracellular DIRAS1 protein level (). This result suggests that there may be more critical post-transcriptional regulatory mechanisms to significantly reduce DIRAS1 protein levels in cervical cancer cells.
Figure 4. The mechanisms of epigenetic regulation to regulate DIRAS1.
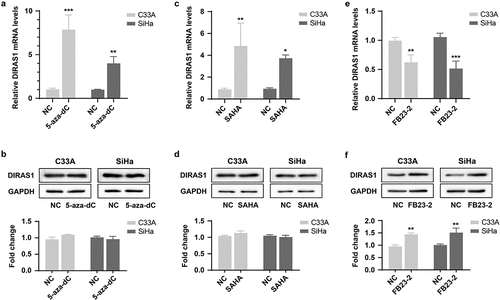
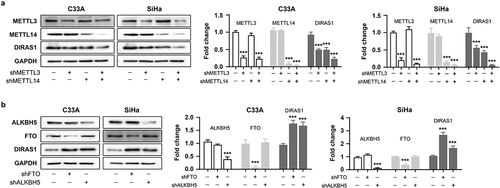
Subsequently, the treatment of cells with the FTO inhibitor FB23–2 (1 µM for 48 h) resulted in significant down-regulation of intracellular DIRAS1 mRNA levels, but significant up-regulation of DIRAS1 protein levels (). This result implies that m6A modification may be the key post-transcriptional regulatory mechanism to regulate DIRAS1 mRNA degradation and translation. Furthermore, the expression of METTL3, METTL14, METTL3/METTL14, FTO and ALKBH5 were significantly downregulated using specific shRNA plasmid transfection, respectively (). Downregulation of METTL3 and METTL14 expression significantly suppressed DIRAS1 protein expression, and the simultaneous downregulation of METTL3 and METTL14 expression had a synergistic effect (). Downregulation of FTO and ALKBH5 expression significantly increased DIRAS1 protein expression ().
Figure 5. M6A modification promotes DIRAS1 translation.
Discussion
In this study, we investigated the role of DIRAS1 expression in the proliferation, growth and motility of cervical cancer cells cultured in vitro using knockdown and exogenous overexpression of DIRAS1 in C33A and SiHa cells. The results of indicate that DIRAS1 has a significant inhibitory effect on the proliferation, growth, and motility of cervical cancer cells cultured in vitro, which is consistent with its role in other types of solid tumors. Unfortunately, this study was not in a position to validate its role on the in vivo experiments. The results of this study, in combination with those of other studies, suggest that DIRAS1 has a potent cancer inhibitory effect. Focusing on DIRAS1 may be beneficial for the advancement of cancer treatment.
Regarding the mechanism of cancer inhibition of DIRAS1, DIRAS1 appears to inhibit cancer by competitively binding SmgGDS with other proteins with strong affinity.Citation15 SmgGDS is a key promoter of tumorigenesis and cancer cell proliferation with cytoplasmic-nuclear shuttling. In the cytoplasm, SmgGDS binds to small GTPases and facilitates their transport to the plasma membrane.Citation15 These small GTPases normally exert pro-oncogenic activity. In the nucleus, SmgGDS can regulate the expression of more than 600 gene products, including mainly proteins controlling the cell cycle, by interacting with transcription factor complexes.Citation16 DIRAS1 is expressed in normal cells and binds to SmgGDS, blocking the action of SmgGDS with small GTPases or transcription factor complexes.Citation15,Citation16 In cancer cells, deletion of DIRAS1 expression eliminates this brake and allows SmgGDS to exert oncogenic activity.Citation17 However, other mechanisms of cancer inhibition by DIRAS1 need to be further explored.
Furthermore, using RNA-Seq data as well as IHC, this study determined that DIRAS1 expression was significantly downregulated in cervical cancer tissues. The same trend was also found in other types of solid tumor tissues. Similar downregulation of DIRAS1 expression was found in renal cell carcinoma,Citation6 ovarian cancer,Citation7 colorectal cancer,Citation8 gliomaCitation9 and esophageal squamous cell carcinomaCitation10 tissues. The low-level expression of the DIRAS1 protein in tumor tissues may be the main reason for its lack of nuclear localization. Generally, the protein first accumulates in the cytoplasm before being translocated to the nucleus. For example, nuclear localization is also found to occur with high expression of DIRAS1 in IHC analysis of ovarian cancer tissues.Citation7 The mechanisms mediating the nucleoplasmic shuttling of DIRAS1 protein remain to be explored. In addition, statistical analysis of the data showed that low expression of DIRAS1 was associated with high pathological stage in cervical cancer patients. A study indicates that low level of DIRAS1 is significantly associated with advanced clinical stage and lymph node metastasis in patients with esophageal squamous cell carcinoma.Citation10 Unfortunately, this study failed to better complete patient follow-up and tracking to analyze the correlation between DIRAS1 expression and patient prognosis. However, one study has reported that downregulation of DIRAS1 expression is associated with decreased disease-free survival and overall survival in ovarian cancer patients.Citation7 In addition, it has been suggested that DIRAS1 is a good predictor of overall survival in HER2+ breast cancer patients receiving neoadjuvant chemotherapy.Citation18
The focus of this study is on the regulatory mechanisms underlying the downregulation of DIRAS1 expression in cancer cells. Current mutation analysis did not identify inactivating mutations on the DIRAS1 coding region.Citation9 Heterozygous deletion of the DIRAS1 gene is present in esophageal squamous cell carcinoma tissues.Citation10 The present study was not equipped to detect mutations in DIRAS1 occurring in cervical cancer tissues and mainly focused on epigenetic regulation. Most current studies have also focused on the epigenetic regulation mechanisms of DIRAS1 expression, mainly DNA methylation and histone modification. Aberrant hypermethylation of CpG islands in the promoter region of DIRAS1 is identified by bisulfite sequencing in renal cell carcinoma cell lines (ACHN, 786-O and Caki-1).Citation6 Hypermethylation of the DIRAS1 promoter region and the resulting deletion or reduction of DIRAS1 expression have also been identified in colorectal cancer samples, esophageal squamous cell carcinoma tissues, and corresponding cell lines.Citation8,Citation10
In addition, experimental data have shown that 5-aza-dC treatment of renal cell carcinoma cell lines (ACHN, 786-O, and Caki-1) (10 µM for 96 h),Citation6 colorectal cancer cell lines (2 µM for 96 h, with fluid changes every 24 h),Citation8 glioblastoma cell lines (U251MG and Hs683) (1 µM for 72 h)Citation9 and esophageal squamous cell carcinoma cell lines (KYSE30 and KYSE510)Citation10 result in a significant elevation of DIRAS1 mRNA levels. Treatment of both glioblastoma cell linesCitation9 and renal cell carcinoma cell line (UOK146)Citation19 with histone deacetylase inhibitors also result in a significant increase in DIRAS1 mRNA levels. These findings seem to suggest that DIRAS1 expression is regulated by DNA methylation and histone modifications. However, none of these studies detected changes in DIRAS1 protein levels after inhibitor treatment.
In the present study, we also treated C33A and SiHa cells with 5-aza-dC and histone deacetylase inhibitors and found that DIRAS1 mRNA levels were significantly increased after inhibitor treatment. However, western blot results showed that DIRAS1 protein levels were not significantly altered after inhibitor treatment, suggesting the existence of a more critical post-transcriptional regulatory mechanism for DIRAS1 that further controls the low-level expression of DIRAS1 protein in cancer cells. The application of the FTO inhibitor FB23–2 as well as the knockdown of FTO and ALKBH5 significantly reduced intracellular protein expression of DIRAS1, while knockdown of METTL3/14 significantly increased intracellular DIRAS1 protein expression. This result suggests that the m6A modification mechanism may be involved in the low-level expression of DIRAS1 protein in cancer cells. To the best of our knowledge, studies related to DIRAS1 mRNA m6A modification are lacking. However, the results of this study are still preliminary and fail to clarify the specific m6A modification sites of DIRAS1 mRNA and how m6A modification affects the altered mRNA and protein levels of DIRAS1.
One study evaluates the total mRNA m6A levels in 286 pairs of cervical cancer samples and their adjacent normal tissues, and find that the total mRNA m6A levels in cervical cancer tissues are significantly lower compared with adjacent normal tissues, m6A level reduction is also significantly correlated with FIGO stage, tumor size, degree of differentiation, lymphatic invasion, cancer recurrence and patient survival.Citation20 In addition, some studies have reported that the expression of FTO and ALKBH5 is significantly up-regulated in cervical cancer and significantly enhances the malignant phenotype of cervical cancer cells.Citation21–23 The pro-cancer mechanism of FTO and ALKBH5 may also include the removal of m6A modification of DIRAS1 mRNA, which reduces the DIRAS1 protein levels.
What cannot be ignored is the recognition and interpretation of RNA m6A modifications by m6A “readers”, which really affects the fate of RNA. An m6A “reader” recognizes and binds an m6A site and may mediate different biological processes and changes. For example, YTHDC1 affects pre-mRNA splicingCitation24 and nuclear export.Citation25 YTHDC2 accelerates the translation and decay of mRNAs.Citation26 YTHDF1 enhances the translation efficiency of mRNAs, whereas YTHDF2 promotes the degradation of mRNAs.Citation27,Citation28 YTHDF3 promotes the production of proteins through its interactions with YTHDF1, and influences mRNA decay through YTHDF2.Citation29 IGF2BP1/2/3 improves mRNA stability and translation efficiency.Citation30 Combined with the result that treatment with FTO inhibitors decreased DIRAS1 mRNA levels but increased DIRAS1 protein expression, we hypothesized that m6A modification of DIRAS1 mRNA at least mediated the degradation or decay of its mRNAs while promoting its translation. However, as mentioned earlier, this study was unable to identify the specific m6A modification sites of DIRAS1 mRNA, as well as the specific “readers” that recognize the m6A and the specific roles they mediate.
In summary, DIRAS1 exerts a significant anti-oncogenic function and its expression is significantly downregulated in cervical cancer cells. m6A modification is a key mechanism to regulate DIRAS1 translation and protein levels.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Data availability statement
The datasets generated during and/or analyzed during the current study are not publicly available, but are available from the corresponding author on reasonable request.
Additional information
Funding
Notes on contributors
Yu-Yan Wang
Yu-yan Wang is a Master’s degree, graduated from Jinzhou Medical University, is currently working in the Department of Obstetrics and Gynecology of the First Affiliated Hospital of Jinzhou Medical University, as the deputy chief physician and deputy director of the department, main academic research direction is gynecological tumors.
Lian-Hua Ye
Lian-Hua Ye is studying for a master’s degree at Jinzhou Medical University, specializing in oncology.
An-Qi Zhao
An-Qi Zhao is studying for a master’s degree at Capital Medical University, specializing in obstetrics and gynecology.
Wei-Ran Gao
Wei-Ran Gao is a Master’s degree, graduated from Jinzhou Medical University, is currently working in the Department of Oncology, the First Affiliated Hospital of Jinzhou Medical University, as the deputy chief physician, and main academic research direction is gynecological tumors.
Ning Dai
Ning Dai is a Master’s degree, graduated from Dalian University, is currently working in the Department of Oncology of the First Affiliated Hospital of Jinzhou Medical University, and main academic research direction is gynecological oncology.
Yu Yin
Yu Yin is working in the First Affiliated Hospital of Jinzhou Medical University, and main academic research direction is gynecological tumors.
Xin Zhang
Xin Zhang is a Ph.D., graduated from Shandong Medical University with a major in oncology, and is currently working in the Department of Oncology, the First Affiliated Hospital of Jinzhou Medical University, Chief Physician and Director of the Department of Oncology, with the main academic research direction of gynecological tumors.
References
- Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, Bray F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021;71(3):209–9. doi:10.3322/caac.21660.
- Canfell K, Kim JJ, Brisson M, Keane A, Simms KT, Caruana M, Burger EA, Martin D, Nguyen DTN, Bénard É, et al. Mortality impact of achieving WHO cervical cancer elimination targets: a comparative modelling analysis in 78 low-income and lower-middle-income countries. Lancet. 2020;395(10224):591–603. doi:10.1016/s0140-6736(20)30157-4.
- Brisson M, Kim JJ, Canfell K, Drolet M, Gingras G, Burger EA, Martin D, Simms KT, Bénard É, Boily M-C, et al. Impact of HPV vaccination and cervical screening on cervical cancer elimination: a comparative modelling analysis in 78 low-income and lower-middle-income countries. Lancet. 2020;395(10224):575–90. doi:10.1016/s0140-6736(20)30068-4.
- Mutlu L, Tymon-Rosario J, Harold J, Menderes G. Targeted treatment options for the management of metastatic/persistent and recurrent cervical cancer. Expert Rev Anticancer Ther. 2022;22(6):633–45. doi:10.1080/14737140.2022.2075348.
- Kontani K, Tada M, Ogawa T, Okai T, Saito K, Araki Y, Katada T. Di-ras, a distinct subgroup of ras family GTPases with unique biochemical properties. J Biol Chem. 2002;277(43):41070–41078. doi:10.1074/jbc.M202150200.
- Xu X, Li J, Wang S, Zheng X, Xie L. Rnaa and Vector-mediated overexpression of DIRAS1 suppresses tumor growth and migration in renal cell carcinoma. Mol Ther Nucleic Acids. 2018;12:845–53. doi:10.1016/j.omtn.2018.07.019.
- Sutton MN, Yang H, Huang GY, Fu C, Pontikos M, Wang Y, Mao W, Pang L, Yang M, Liu J, et al. RAS-related GTPases DIRAS1 and DIRAS2 induce autophagic cancer cell death and are required for autophagy in murine ovarian cancer cells. Autophagy. 2018;14(4):637–53. doi:10.1080/15548627.2018.1427022.
- Zheng R, Gao D, He T, Zhang M, Zhang X, Linghu E, Wei L, Guo M. Methylation of DIRAS1 promotes colorectal cancer progression and may serve as a marker for poor prognosis. Clin Epigenetics. 2017;9(1):50. doi:10.1186/s13148-017-0348-0.
- Rothhammer-Hampl T, Liesenberg F, Hansen N, Hoja S, Delic S, Reifenberger G, Riemenschneider MJ. Frequent epigenetic inactivation of DIRAS-1 and DIRAS-2 contributes to chemo-resistance in gliomas. Cancers Basel. 2021;13(20):5113. doi:10.3390/cancers13205113.
- Zhu YH, Fu L, Chen L, Qin YR, Liu H, Xie F, Zeng T, Dong S-S, Li J, Li Y, et al. Downregulation of the novel tumor suppressor DIRAS1 predicts poor prognosis in esophageal squamous cell carcinoma. Cancer Res. 2013;73(7):2298–309. doi:10.1158/0008-5472.Can-12-2663.
- Deans C, Maggert KA. What do you mean, “epigenetic”? Genetics. 2015;199(4):887–96. doi:10.1534/genetics.114.173492.
- Smith ZD, Meissner A. DNA methylation: roles in mammalian development. Nat Rev Genet. 2013;14(3):204–20. doi:10.1038/nrg3354.
- Xiao C, Fan T, Tian H, Zheng Y, Zhou Z, Li S, Li C, He J. H3K36 trimethylation-mediated biological functions in cancer. Clin Epigenetics. 2021;13(1):199. doi:10.1186/s13148-021-01187-2.
- Pan Y, Ma P, Liu Y, Li W, Shu Y. Multiple functions of m(6)A RNA methylation in cancer. J Hematol Oncol. 2018;11(1):48. doi:10.1186/s13045-018-0590-8.
- Brandt AC, Koehn OJ, Williams CL. SmgGDS: an emerging master regulator of prenylation and trafficking by small GTPases in the ras and rho families. Front Mol Biosci. 2021;8:685135. doi:10.3389/fmolb.2021.685135.
- Gonyo P, Bergom C, Brandt AC, Tsaih SW, Sun Y, Bigley TM, Lorimer EL, Terhune SS, Rui H, Flister MJ, et al. SmgGDS is a transient nucleolar protein that protects cells from nucleolar stress and promotes the cell cycle by regulating DREAM complex gene expression. Oncogene. 2017;36(50):6873–83. doi:10.1038/onc.2017.280.
- Bergom C, Hauser AD, Rymaszewski A, Gonyo P, Prokop JW, Jennings BC, Lawton AJ, Frei A, Lorimer EL, Aguilera-Barrantes I, et al. The tumor-suppressive small GTPase DiRas1 binds the noncanonical guanine nucleotide exchange factor SmgGDS and antagonizes SmgGDS interactions with oncogenic small GTPases. J Biol Chem. 2016;291(12):6534–45. doi:10.1074/jbc.M115.696831.
- Barrón-Gallardo CA, Garcia-Chagollán M, Morán-Mendoza AJ, Delgadillo-Cristerna R, Martínez-Silva MG, Villaseñor-García MM, Aguilar-Lemarroy A, Jave-Suárez LF. A gene expression signature in HER2+ breast cancer patients related to neoadjuvant chemotherapy resistance, overall survival, and disease-free survival. Front Genet. 2022;13:991706. doi:10.3389/fgene.2022.991706.
- Verma SP, Agarwal A, Das P. Sodium butyrate induces cell death by autophagy and reactivates a tumor suppressor gene DIRAS1 in renal cell carcinoma cell line UOK146. Vitro Cell Dev Biol Anim. 2018;54(4):295–303. doi:10.1007/s11626-018-0239-5.
- Wang X, Li Z, Kong B, Song C, Cong J, Hou J, Wang S. Reduced m(6)A mRNA methylation is correlated with the progression of human cervical cancer. Oncotarget. 2017;8(58):98918–98930. doi:10.18632/oncotarget.22041.
- Huo FC, Zhu ZM, Du WQ, Pan YJ, Jiang X, Kang MJ, Liu B-W, Mou J, Pei D-S. HPV E7-drived ALKBH5 promotes cervical cancer progression by modulating m6A modification of PAK5. Pharmacol Res. 2023;195:106863. doi:10.1016/j.phrs.2023.106863.
- Huang J, Yang J, Zhang Y, Lu D, Dai Y. FTO promotes cervical cancer cell proliferation, colony formation, migration and invasion via the regulation of the BMP4/Hippo/YAP1/TAZ pathway. Exp Cell Res. 2023;427(1):113585. doi:10.1016/j.yexcr.2023.113585.
- Zhou S, Bai ZL, Xia D, Zhao ZJ, Zhao R, Wang YY, Zhe H. FTO regulates the chemo-radiotherapy resistance of cervical squamous cell carcinoma (CSCC) by targeting β-catenin through mRNA demethylation. Mol Carcinog. 2018;57(5):590–597. doi:10.1002/mc.22782.
- Xiao W, Adhikari S, Dahal U, Chen YS, Hao YJ, Sun BF, Sun H-Y, Li A, Ping X-L, Lai W-Y, et al. Nuclear m(6)A Reader YTHDC1 Regulates mRNA Splicing. Mol Cell. 2016;61(4):507–519. doi:10.1016/j.molcel.2016.01.012.
- Roundtree IA, Luo GZ, Zhang Z, Wang X, Zhou T, Cui Y, Sha J, Huang X, Guerrero L, Xie P, et al., YTHDC1 mediates nuclear export of N(6)-methyladenosine methylated mRnas. Elife. 2017;6:e31311. doi:10.7554/eLife.31311.
- Hsu PJ, Zhu Y, Ma H, Guo Y, Shi X, Liu Y, Qi M, Lu Z, Shi H, Wang J, et al. Ythdc2 is an N(6)-methyladenosine binding protein that regulates mammalian spermatogenesis. Cell Res. 2017;27(9):1115–1127. doi:10.1038/cr.2017.99.
- Wang X, Zhao BS, Roundtree IA, Lu Z, Han D, Ma H, Weng X, Chen K, Shi H, He C, et al. N(6)-methyladenosine modulates messenger RNA translation efficiency. Cell. 2015;161(6):1388–1399. doi:10.1016/j.cell.2015.05.014.
- Wang X, Lu Z, Gomez A, Hon GC, Yue Y, Han D, Fu Y, Parisien M, Dai Q, Jia G, et al. N6-methyladenosine-dependent regulation of messenger RNA stability. Nature. 2014;505(7481):117–20. doi:10.1038/nature12730.
- Shi H, Wang X, Lu Z, Zhao BS, Ma H, Hsu PJ, Liu C, He C. YTHDF3 facilitates translation and decay of N(6)-methyladenosine-modified RNA. Cell Res. 2017;27(3):315–328. doi:10.1038/cr.2017.15.
- Huang H, Weng H, Sun W, Qin X, Shi H, Wu H, Zhao BS, Mesquita A, Liu C, Yuan CL, et al. Recognition of RNA N(6)-methyladenosine by IGF2BP proteins enhances mRNA stability and translation. Nat Cell Biol. 2018;20(3):285–295. doi:10.1038/s41556-018-0045-z.