
A key step in pre-mRNA processing is represented by splicing, the multilayered process operated by the spliceosome that removes the intervening non-coding introns and ligates adjacent exons. Splicing is necessary to yield a mature, translatable mRNA and its dysregulation underlies many human pathologies.Citation1 Notably, weak conservation of the sequences defining the exon-intron boundaries allows flexibility in the recognition of many exons by the spliceosome. As a consequence, alternative splicing (AS) of such variable exons generates multiple mRNAs, with potentially different coding properties and patterns of expression, from most mammalian genes.Citation1 Retention of select introns into mature mRNAs represents a peculiar pattern of AS that is emerging as a regulatory mechanism for developmentally-modulated gene expression patterns.Citation2 Granulocyte differentiation provided one of the first examples of intron retention (IR) program set in motion to regulate gene expression. Transcripts encoding for proteins no longer required for granulopoiesis, and potentially interfering with it, are eliminated by the nonsense-mediated (NMD) pathway through IR-mediated introduction of premature termination codons (PTCs).Citation3 Similar coordinated and widespread dampening of specific set of genes through IR has been described for several differentiation programs or cellular responses to external stimuli.Citation2 Spermatogenesis, however, represents a remarkable exception. Spermatogenesis involves profound genetic and morphological changes that are necessary for the differentiation of the male germ cell into a motile, fertile spermatozoon. Although proper progression of spermatogenesis requires the timely regulated expression of specific factors for each phase, transcription is not always active during this process. Indeed, nuclear condensation in post-meiotic male germ cells leads to a progressive decline of their transcriptional activity, which ultimately halts in spermatozoa.Citation4 We have recently shown that an orchestrated IR program activated during meiosis contributes to temporally regulate the expression of genes during spermatogenesis.Citation5 IR generates stable transcripts which persist in the nucleus of meiotic spermatocytes for several days after their synthesis, whose splicing and translation is delayed until the post-meiotic phases of spermatogenesis.Citation5 In this way, meiotic IR acts as a compensatory mechanism for the transcriptional inactivity of the terminal phases of germ cell differentiation. Of note, IR-regulated genes encode for proteins that are crucial for proper development and functionality of the spermatozoon, such as those involved in the maturation of the flagellum or in sperm-egg recognition. Interestingly, robust accumulation in the nucleus of stable intron-retaining transcripts was also observed during the cellular response to heat shock.Citation6 This observation suggests that IR stabilizes precursor transcripts before the global inhibition of RNA transcription caused by heat, and that their delayed splicing may promote efficient recovery of gene expression at the end of the stress. Furthermore, a “positive” role for IR was described in neurons. Post-transcriptional splicing of intron-retaining transcripts during neuronal activation allowed rapid expression of proteins encoded by genes that are too long to be rapidly transcribed, processed and translated in response to transient external stimuli.Citation7 Thus, regulation of IR is emerging as a mechanism that can compensate both deficiencies and inefficiencies of the transcriptional process in eukaryotic cells.Citation5,7 Notably, common traits of spermatogenic and neuronal IR programs are the nuclear preservation of intron-retaining transcripts and their protection from nuclear mechanisms of RNA surveillance.Citation5,7 Therefore, it might be of interest to understand whether common mechanisms underlying these features exist in germ cells and neurons, possibly relying on the activity of splicing factors that are selectively expressed in these cells, such as PTBP2 or the STAR protein SLM2Citation4.
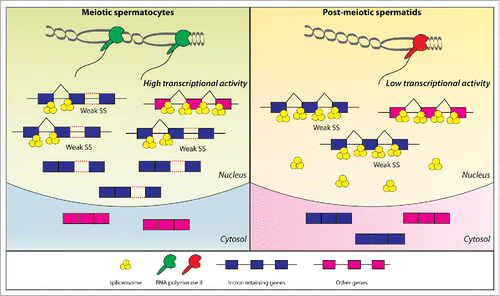
Intron-retaining genes are expressed at higher levels than properly spliced genes in meiotic cells, and splicing of their weak introns is improved by reducing the transcriptional load through inhibition of the RNA polymerase II activityCitation5 (). This finding suggests that an RNA synthetic activity exceeding the splicing capability of the cell represents the driver of the male meiotic IR program. Higher expression levels were also observed for heat shock-regulated intron-retaining genes and neuronal post-transcriptionally spliced pre-mRNAs.Citation6,7 Thus, competition of introns for limiting splicing factors could represent a conserved mechanism controlling eukaryotic gene expression through developmentally and physiologically regulated IR. A crucial point in this regulatory mechanism is the combination of high transcriptional levels with intronic sequence features predisposing to poor splicing efficiency. It would be interesting to investigate whether these features have been evolutionary conserved in genes that play key roles in cellular processes characterized by transcriptional insufficiency.
Figure 1. Balance between transcriptional activity and splicing capability regulates intron-retention during germ cell differentiation. High transcriptional activity of meiotic spermatocytes (left panel) generates high levels of transcripts for intron-retaining genes (blue genes). Weak introns of these genes are not efficiently recognized by the spliceosome and their unspliced transcripts are consequently retained in the nucleus. The lower transcriptional activity of post-meiotic spermatids (right panel) then allows efficient splicing of such intron-retaining genes, whose transcripts are efficiently exported in the cytoplasm and translated into proteins.
IR as a consequence of a transcriptional burst highlights the importance of maintaining a tight balance between transcription and splicing efficiency in eukaryotic cells. In line with this, the general increase in RNA synthesis elicited by oncogenic MYC was shown to render cancer cells more susceptible to spliceosome inhibition, which caused pervasive IR.Citation8 Thus, while perturbing this balance augments vulnerability of proliferating cells, it appears to spare post-mitotic cells in which global IR regulation may have evolved as a fine-tuned differentiation/developmental program.
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
References
- Lee Y, Rio DC. Mechanisms and regulation of alternative Pre-mRNA splicing. Annu Rev Biochem 2015; 84:291-323; PMID:25784052; https://doi.org/10.1146/annurev-biochem-060614-034316
- Jacob AG, Smith CW. Intron retention as a component of regulated gene expression programs. Hum Genet 2017; Epub ahead of print; PMID:28391524; https://doi.org/10.1007/s00439-017-1791-x
- Wong JJ, Ritchie W, Ebner OA, Selbach M, Wong JW, Huang Y, Gao D, Pinello N, Gonzalez M, Baidya K, Thoeng A, Khoo TL, Bailey CG, Holst J, Rasko JE. Orchestrated intron retention regulates normal granulocyte differentiation. Cell 2013; 154(3):583-95; PMID:23911323; https://doi.org/10.1016/j.cell.2013.06.052
- Paronetto MP, Sette C. Role of RNA-binding proteins in mammalian spermatogenesis. Int J Androl 2010; 33:2-12; PMID:19281489; https://doi.org/10.1111/j.1365-2605.2009.00959.x
- Naro C, Jolly A, Di Persio S, Bielli P, Setterblad N, Alberdi AJ, Vicini E, Geremia R, De la Grange P, Sette C. An orchestrated intron retention program in meiosis controls timely usage of transcripts during germ cell differentiation. Dev Cell 2017; 41:82-93.e4; PMID:28366282; https://doi.org/10.1016/j.devcel.2017.03.003
- Shalgi R, Hurt JA, Lindquist S, Burge CB. Report widespread inhibition of posttranscriptional splicing shapes the cellular transcriptome following heat shock. Cell Rep 2014; 7:1362-70; PMID:24857664; https://doi.org/10.1016/j.celrep.2014.04.044
- Mauger O, Lemoine F, Scheiffele P. Targeted intron retention and excision for rapid gene regulation in response to neuronal activity. Neuron 2016; 92:1266-78; PMID:28009274; https://doi.org/10.1016/j.neuron.2016.11.032
- Hsu TY, Simon LM, Neill NJ, Marcotte R, Sayad A, Bland CS, Echeverria GV, Sun T, Kurley SJ, Tyagi S, Karlin KL, Dominguez-Vidaña R, Hartman JD, Renwick A, Scorsone K, Bernardi RJ, Skinner SO, Jain A, Orellana M, Lagisetti C, Golding I, Jung SY, Neilson JR, Zhang XH, Cooper TA, Webb TR, Neel BG, Shaw CA, Westbrook TF. The spliceosome is a therapeutic vulnerability in MYC-driven cancer. Nature [ Internet] 2015; 525:384-8; PMID:26331541; https://doi.org/10.1038/nature14985