
ABSTRACT
As positive-sense RNA viruses, the genomes of flaviviruses serve as the template for all stages of the viral life cycle, including translation, replication, and infectious particle production. Yet, they encode just 10 proteins, suggesting that the structure and dynamics of the viral RNA itself helps shepherd the viral genome through these stages. Herein, we highlight advances in our understanding of flavivirus RNA structural elements through the lens of their impact on the viral life cycle. We highlight how RNA structures impact translation, the switch from translation to replication, negative- and positive-strand RNA synthesis, and virion assembly. Consequently, we describe three major themes regarding the roles of RNA structure in flavivirus infections: 1) providing a layer of specificity; 2) increasing the functional capacity; and 3) providing a mechanism to support genome compaction. While the interactions described herein are specific to flaviviruses, these themes appear to extend more broadly across RNA viruses.
Introduction
The Flavivirus genus includes several emerging and re-emerging viral pathogens of public health concern. The most well-known of these are the mosquito-borne flaviviruses, which include Zika virus (ZIKV), dengue virus (DENV), West Nile virus (WNV), yellow fever virus (YFV) and Japanese encephalitis virus (JEV); however, this genus also includes tick-borne, insect-specific, and no-known vector flaviviruses [Citation1]. Billions of people around the globe remain at risk of flavivirus infections, with climate change increasing the geographic risk as the range of their insect vectors expands [Citation2,Citation3]. Despite their high-risk profile, we still lack effective vaccines and/or antiviral treatments for many flaviviruses, necessitating a better understanding of the intricacies of their life cycles.
Flaviviruses enter cells through receptor-mediated endocytosis and subsequent endosome acidification triggers viral uncoating, releasing the viral genomic RNA into the cytoplasm. The capped ~11 kb flavivirus genome contains a single open reading frame that encodes a ~ 3400 amino acid polyprotein (). Upon uncoating, the positive-sense genomic RNA is immediately able to recruit the ribosome to produce the viral polyprotein, which is processed by both cellular and viral proteases into the 10 mature viral proteins. This includes three structural proteins (capsid, envelope, and pre-membrane) that form the viral particle, and seven non-structural (NS) proteins (NS1, NS2A, NS2B, NS3, NS4A, NS4B and NS5), which coordinate viral replication, virion assembly, and evasion of cellular antiviral responses. Once sufficient translation of the viral proteins has occurred, the genome is condensed into virally induced invaginations of the endoplasmic reticulum (ER) membrane to form the replication organelle (RO), the site of viral genome replication [Citation4,Citation5]. Within the RO, genome replication occurs via synthesis of a negative-strand replicative intermediate, followed by subsequent positive-strand genomic RNA synthesis. Together, the NS3 protein, which has protease, helicase, and nucleotide triphosphatase activities, and the NS5 protein, which has RNA-dependent RNA polymerase (RdRp) and methyltransferase (MTase) activities, contain all the enzymatic activity required for viral RNA replication and capping of newly synthesized positive-sense RNAs. Following genome replication, newly synthesized positive-sense genomic RNAs are released from the RO, where they can either undergo translation and eventually seed a new RO, or can be transported to the assembly site to be packaged into progeny virions [Citation4,Citation5]. Viral particles are assembled on the ER membrane adjacent to ROs and bud into the ER before exiting the cell via the secretory pathway.
Figure 1. Flavivirus genome organization.
Notably, the viral positive-sense genomic RNA must serve as a template for all the intracellular steps of the viral life cycle, including translation, replication, and packaging. As such, the genomic RNA itself must be highly dynamic and readily able to accommodate unwinding, elongation, and exposure of different regions of the RNA to viral and cellular proteins in a temporally and spatially regulated manner. Recent innovations in RNA structure-probing techniques have provided novel insights into the RNA structures involved in the flavivirus life cycle and their dynamics (see Box 1). In this review, we first provide an overview of flavivirus cis-acting RNA elements and then highlight the roles of these RNA structural elements and their interplay in the various stages of the viral life cycle. In doing so, we emphasize recent advances that have reshaped our understanding of flavivirus translation, the switch from translation to replication, negative- and positive-strand RNA synthesis, and virion assembly.
Organization of cis-acting RNA elements in the flavivirus genome
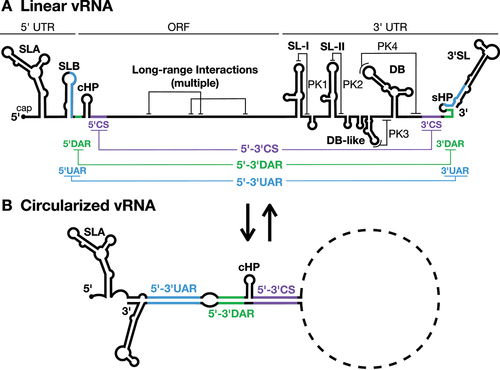
Flavivirus genomic RNAs are known to be highly structured, with the 5´ and 3´ ends of the genome being particularly well characterized (reviewed in [Citation17]) ( and ). Briefly, the 5´ untranslated region (UTR) contains stem-loop A (SLA) and SLB which are essential for viral RNA replication [Citation18,Citation19]. SLB also contains the 5´ upstream of AUG region (5´UAR), a sequence that is essential for a long-range RNA-RNA interaction that aids in genome cyclization and viral replication [Citation20]. Within the coding sequence, the stable capsid coding-region hairpin (cHP, ) is required for efficient translation and viral RNA replication [Citation21,Citation22]. Adjacent to cHP lies the 5´ downstream of AUG region (5´DAR) and 5´ cyclization sequence (5´CS), additional sequences that aid in genome cyclization for viral RNA replication () [Citation20,Citation23].
Table 1. RNA structures in the flavivirus genome and their roles in the viral life cycle.
Specific RNA structures in the 3´ UTR also play roles in viral translation, RNA replication, and virion assembly. The 3´ UTR of mosquito-borne flaviviruses can be divided into three distinct domains (). The first consists of the variable region, which typically contains one or more stem-loop (SL) structures; while the second consists of one to two dumbbell (DB) structures [Citation24]. Interestingly, both the SL and DB structures form local pseudoknots (PKs) that are 5´-3´ exoribonuclease 1 (XRN-1)-resistant, resulting in the generation of subgenomic flavivirus RNAs (sfRNAs). These sfRNAs have been implicated in inhibition of cellular antiviral responses and in host adaptation between mammals and their mosquito vectors [Citation25]. The final domain contains the 3´CS (which forms a long-range RNA-RNA interaction with the 5´CS), short hairpin (sHP), and the 3´SL [Citation20,Citation26]. The sHP and 3´SL are required for viral RNA replication and contain the 3´UAR and 3´DAR elements that help mediate genome cyclization with the 5´ end of the viral RNA [Citation24,Citation26,Citation27]. Notably, the 3´ UTR has also been implicated in viral genome packaging through interactions with the viral NS2A protein, a small hydrophobic protein implicated in both RO biogenesis and virion assembly [Citation28,Citation29].
In addition to these well-characterized structures in the 5´ and 3´ UTRs, whole genome chemical mapping and cross-linking approaches have demonstrated a high degree of local and long-range secondary and tertiary structure across the flavivirus genome and a dynamic interplay between these structures [Citation30–33]. While there is still much left to learn regarding these RNA structures and their dynamics, the body of knowledge regarding the myriad roles of RNA structure in the viral life cycle continues to grow and inform our understanding of the complex and tightly regulated life cycles of flaviviruses. In the following sections, we provide a summary of recent advances that have shaped our understanding of how the viral RNA itself contributes to the coordination of viral translation, negative- and positive-strand RNA synthesis, RNA capping, and genome packaging. In light of these advances, we describe three major themes regarding the roles of RNA structure in the flavivirus life cycle: 1) RNA structure often provides specificity to the tightly regulated viral life cycle; 2) the ability of the RNA to fold into complex three-dimensional structures increases the functional capacity of the RNA beyond its sequence alone; and 3) the ability to form long-range RNA-RNA interactions likely facilitates viral genome compaction.
How does the flavivirus genomic RNA direct and regulate viral polyprotein production?
Flavivirus genomes contain a canonical type I (m7GpppN) cap structure and are thus primarily translated via cap-dependent translation, utilizing the same mechanism as cellular mRNAs. However, in most mosquito-borne flaviviruses, the start codon is in a canonically poor translation initiation context, often 40–50 nucleotides (nt) upstream of another start codon in a more favourable context [Citation22]. Despite this, initiation from the first start codon is heavily favoured in both mammalian and mosquito cells [Citation22,Citation34,Citation35].
This start codon preference is driven by the cHP, a small stem-loop structure that occurs approximately 12–16 nt downstream of the start codon ( and ) [Citation22]. Selection of the correct start codon occurs in a structure-dependent but sequence-independent manner, in which the positioning of the structure relative to the start codon appears to be the critical factor [Citation22,Citation33,Citation36]. Ribosomal pausing at the cHP structure thus appears to help select for the proper start codon, providing specificity to the translation initiation process [Citation22]. Despite this, flavivirus translation efficiency is not substantially affected by defects in cHP structure that result in improper start codon selection, raising the question of why proper start codon selection is maintained across flaviviruses [Citation21,Citation22]. One possible explanation is that proper start codon selection helps ensure the production of the full-length capsid protein. Without the cHP, initiation of translation in DENV occurs at a slightly downstream start codon resulting in loss of the first 15 amino acids of the capsid protein [Citation22,Citation37]. Interestingly, this region contains one of the two basic amino acid clusters at the N-terminus of the capsid protein that are critical for infectious particle production in DENV, and presumably interact with the viral RNA genome during the encapsidation process [Citation37]. However, it is still unclear whether this region is strictly required for packaging across all flaviviruses, as YFV can tolerate N-terminal capsid protein deletions of up to 40 amino acids while maintaining infectious particle production [Citation38]. Nonetheless, it is possible that YFV is the outlier in this regard, considering the strict conservation of the cHP structure across the Flavivirus genus, and the need for a sufficient level of positive-charge in the disordered N-terminus of the viral capsid protein for infectious particle production [Citation22,Citation37,Citation38].
In addition to the cHP, the viral 3´ UTR has also been shown to act as a translational enhancer, although the precise mechanism remains unclear [Citation39–42]. Notably, while capping of viral genomic RNAs is critical for successful infection, uncapped genomic RNAs are also capable of establishing productive infections when introduced into susceptible and permissive cells (see Box 2) [Citation39–43]. Thus, in addition to the cap structure, cis-acting RNA elements in the genomic RNA, including the 5´ UTR, cHP, and 3´ UTR, all contribute to translational regulation, thereby increasing the overall functional capacity of the flavivirus genome.
What mediates the switch from translation to replication in the flavivirus life cycle?
The switch from translation to replication is thought to be driven by ribosome exclusion and RO biogenesis [Citation45]. As positive-sense RNA viruses, flaviviral genomic RNAs are ribosome ready and can be translated immediately upon genome uncoating into the cytoplasm. However, once enough viral proteins have accumulated, flaviviruses are met with a dilemma – ribosomes travel in the 5´ to 3´ direction, while the viral NS5 RdRp travels in the 3´ to 5´ direction. Thus, translation and viral RNA replication cannot occur simultaneously on a single viral genomic RNA molecule. As such, there must be a mechanism to switch off translation for viral RNA replication to occur [Citation45,Citation46]. Presumably, the viral genomic RNA must also be efficiently condensed into a RO to establish viral RNA replication [Citation17,Citation45]. RO biogenesis likely occurs somewhat concurrently with ribosome exclusion from the viral genomic RNA and may contribute to establishing and/or maintaining translational inhibition. In this section, we discuss how viral genome cyclization and SLA interactions with the viral NS5 protein facilitate translational shutoff. We also discuss recent data that indicate that viral protein accumulation and genome compaction may be sufficient to establish ROs, albeit at low efficiency, even in the absence of an active mechanism(s) for ribosome exclusion.
Genome cyclization and the recruitment of NS5 to SLA contribute to translational shut-off
The importance of flaviviral genome cyclization as a precursor to viral RNA replication is well-established and has been the topic of several recent reviews [Citation17,Citation47]. In brief, genome cyclization involves long-range interactions between RNA elements at the 5´ and 3´ ends of the viral genome, including the 5´-3´UAR, DAR and CS elements, which are mutually exclusive with local RNA structures in the linear genome conformation ( and ). For example, in DENV, the highly stable DB pseudoknots in the 3´ UTR are only formed in the linear RNA conformation, implying that there must be a large energetic barrier for RNA rearrangement into the replication-competent circularized form of the genome [Citation48]. However, a precise equilibrium between linear and circular viral RNA seems to be essential for infection as stabilization of either RNA conformation is deleterious to viral RNA accumulation [Citation26,Citation49].
Recent advances indicate that genome cyclization may also be important for switching off translation. Interestingly, genome cyclization was shown to impair 48S ribosomal complex formation on the correct start codon of ZIKV and DENV genomes, inhibiting translation initiation at this site, and resulting in clearance of ribosomes from the viral genomic RNA [Citation35]. This finding suggests that the process of genome cyclization itself promotes translational inhibition and the switch between translation and replication in the flaviviral life cycle. Notably, this interplay between local and long-range structures to switch between these two stages of the viral life cycle provides another clear example of how RNA structures increase the functional capacity of the flaviviral genome.
In addition to the genome cyclization elements, interactions between the viral NS5 protein and the SLA structure at the 5´ terminus of the viral genomic RNA have also been implicated in translational shut-off (). Similar to genome cyclization, the NS5 protein is able to inhibit translation initiation of ZIKV and DENV genomic RNAs both in vitro and in cell culture, in a SLA-dependent manner [Citation34]. Interestingly, the interaction between NS5 and SLA appears to be further stabilized by the proximal SLB structure, suggesting that NS5-SLA interactions may precede genome cyclization in the flavivirus life cycle [Citation19]. While the precise mechanism of NS5-mediated inhibition of translation initiation remains to be deciphered, it is conceivable that NS5 binding may preclude recruitment of translation initiation factors to the 5´ cap. Additionally, NS5-SLA interactions likely induce RNA rearrangements that either promote or involve genome cyclization, perhaps via dimerization of NS5 proteins bound concurrently to SLA at the 5´ terminus and the 3´SL at the 3´ terminus of the viral genome [Citation35,Citation50,Citation51]. Nevertheless, whether binding of NS5 to SLA precedes or is concurrent with genome cyclization, these processes likely occur during pauses in translation of the viral genomic RNA following translational bursts, and together allow the viral RNA to be cleared of ribosomes and subsequently condensed into a RO [Citation52].
Viral protein accumulation and genome compaction may be sufficient for RO biogenesis even in the absence of active translational inhibition
While translational inhibition is an important element in the switch from translation to replication, RO biogenesis is also likely a central part of this process. Intriguingly, the recent development of a replication-independent plasmid-induced replication organelle biogenesis (pIRO) system revealed that it is possible for both DENV and ZIKV genomic RNAs to form ROs, even in the absence of the 5´ UTR and 5´ cyclization elements, albeit very inefficiently [Citation53]. These results suggest that viral protein accumulation, viral protein-RNA, and RNA-RNA interactions may be sufficient for facilitating RO biogenesis even in the absence of an active translational inhibition mechanism. Viral protein-RNA and RNA-RNA interactions may also aid in the necessary compaction of the flavivirus genome into the RO, perhaps through promoting phase separation or biomolecular condensate formation [Citation45]. This latter process has been implicated in ribosome exclusion, and phase separation is also hypothesized to underlie viral RO biogenesis in positive-sense, negative-sense, and double-stranded RNA viruses [Citation45]. Nevertheless, while the pIRO system indicates that the flaviviral RNA is capable of being condensed into a RO in the absence of the 5´ UTR and genome cyclization, this appears to be a rare event in a system where viral RNAs are produced constitutively [Citation53]. As such, during an infection where a single genome needs to be capable of establishing an infection, NS5-SLA interactions and genome cyclization may help facilitate the switch from translation to replication for efficient RO biogenesis.
Long-range RNA-RNA interactions and cellular RNA-binding proteins may facilitate genome compaction for RO biogenesis
While it is largely unclear at present what specific RNA-RNA and protein-RNA interactions underlie RO biogenesis in flaviviruses, it is likely that some of the same long-range RNA-RNA interactions that are necessary for nucleocapsid formation also promote genome compaction into ROs [Citation30–33]. Furthermore, cellular RNA-binding proteins (RBPs) are also likely to contribute to viral RNA compaction during RO biogenesis. For example, insulin-like growth factor 2 mRNA-binding protein 2 (IGF2BP2) can interact with ZIKV NS5 and both the viral 5´ and 3´ UTRs – an interaction that has been demonstrated to be critical for efficient viral RO biogenesis [Citation54]. However, DENV and WNV do not appear to have an IGF2BP2 dependency, implying that the specific cellular RBPs used to promote RO biogenesis may vary between flaviviruses [Citation54]. Additionally, as the viral genomic RNA must be accessible to NS5 to undergo RNA synthesis, it is likely that the degree of genome compaction in the RO is also important for efficient viral RNA replication. In line with this, cellular Ras GTPase-activating protein-binding protein 1 (G3BP1)-promoted genome compaction of non-translating ZIKV and WNV RNAs has an antiviral effect, impairing proper RO biogenesis [Citation55]. However, the eukaryotic initiation factor 4A (eIF4A) helicase activity counteracts G3BP1-mediated compaction, implying that unwinding of the viral RNA could help prevent over-compaction of the viral RNA during RO biogenesis [Citation55].
Taken together, recent studies have made it clear that a myriad of RNA sequences and structures are important in directing the switch from translation to replication in the flavivirus life cycle, by regulating both translational shutoff and facilitating RO biogenesis.
How do RNA structures direct viral RNA replication?
Flavivirus RNA replication is a two-step process that involves: 1) negative-strand RNA synthesis, which uses the single-stranded positive-sense genomic RNA as a template and results in a double-stranded (ds) RNA replicative intermediate; and subsequently, 2) positive-strand RNA synthesis, using the dsRNA replicative intermediate as a template. Notably, viral RNA replication is a tightly regulated process, with positive-strand genomic RNAs outnumbering negative-strands on the order of approximately 50:1 during infection [Citation56]. Genome cyclization is implicated in loading of the NS5 RdRp on the 3´ terminus of the positive-strand genomic RNA for negative-strand RNA synthesis. Yet, it remains unclear how the negative-strand is chosen as the template for subsequent positive-strand RNA synthesis, given the starting point is the dsRNA replicative intermediate. For example, the NS5 RdRp has specificity for a 3´-UC dinucleotide, which is present at both the 3´ terminus of the positive- and negative-strands [Citation57]. Moreover, the NS3 helicase requires a 3´ single-stranded overhang to unwind dsRNA; meaning that both NS3 and NS5 would presumably have similar affinity for either end of the dsRNA replicative intermediate. This lack of specificity from the viral proteins indicates that the viral RNAs themselves likely provide the direction necessary for the coupled processes of negative- and positive-strand RNA synthesis, and subsequent capping of newly synthesized positive-sense genomic RNAs. Notably, while SLA is already known to play a crucial role in RNA replication, recent advances regarding the tertiary structure of SLA provide new insights into how the NS5-SLA interaction could provide specificity to the genome replication process. In this section, we highlight new observations in NS5-SLA binding models, discuss how NS5-SLA interactions promote negative-strand RNA synthesis, and provide several models for the initiation and progression of negative-strand RNA synthesis. We then discuss positive-strand RNA synthesis from the dsRNA replicative intermediate, a putative positive-strand promoter, and how NS5-SLA interactions facilitate capping of newly-synthesized genomic positive-sense RNAs.
Recent advances in NS5-SLA interaction models
While the interaction between SLA and NS5 remains one of the best characterized interactions between the flavivirus RNA genome and a viral protein, recent advances have challenged the canonical view of the SLA structure. Traditionally, SLA was thought to take on a ‘Y’ shaped conformation, characterized by a base stem, side stem-loop (SSL) and a top stem-loop (TSL) () [Citation14,Citation58,Citation59]. However, recent crystallographic and cryo-EM-based structural studies suggest that SLA may at least sample an alternative ‘L’ or ‘V’ shaped structure upon NS5 binding () [Citation14,Citation15,Citation60]. In these models, the base stem and TSL are present, but the SSL bases remain unpaired in the central bulge. When interacting with NS5, the base stem and TSL of the ‘L’ or ‘V’ shaped SLA bridge the NS5 MTase and RdRp domains, respectively [Citation14,Citation15]. Kinetic analyses indicate that binding of NS5 to SLA is a 3-step process in which the RdRp first binds to the TSL of SLA, followed by binding of the MTase to the base stem, and finally a rearrangement that stabilizes these interactions [Citation61]. At present, it is unclear whether flavivirus SLAs exist only in the ‘L’ or ‘V’ conformation, or if SLA samples both the ‘Y’ and ‘L/V’ (or other) conformations at various points in the viral life cycle.
Figure 2. The ‘L/V’ conformations of DENV and ZIKV SLA.
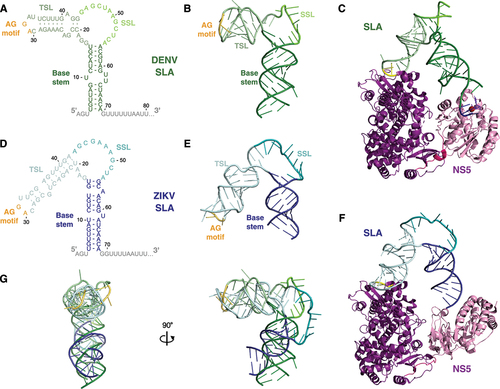
These new models for SLA and NS5 binding are largely consistent with known requirements for this interaction. Beyond the overall structural architecture, foot-printing and mutational studies have consistently demonstrated that NS5 binding to SLA also depends on a conserved ‘AG’ motif in the unpaired loop of the TSL that is maintained in all conformations [Citation14,Citation27,Citation51,Citation58,Citation62]. Despite the different angle of the TSL relative to the base of SLA in the ‘L’ and ‘V’ conformations, the AG motif in the unpaired loop of the TSL sits on the same face of the structure, implying that they both interact with their respective NS5 proteins along the same surface () [Citation14]. Furthermore, when the structures of ZIKV and DENV SLA are aligned, the TSL and base stems both largely occupy the same physical space due to different lengths of the helices in ZIKV relative to DENV [Citation14]. Thus, ZIKV and DENV SLAs likely bridge the NS5 RdRp and MTase domains in a similar manner . This spatial conservation of NS5 binding interfaces across species provides an explanation for the observation that the NS5s of different flaviviruses are generally interchangeable with respect to SLA binding and that SLA chimeric flaviviruses are replication competent [Citation63,Citation64]. In addition to the TSL ‘AG’ motif, the SSL sequence has also been demonstrated to be important for NS5 binding and viral RNA replication, but it remains unclear whether the sequence, structure, or both, mediate this effect [Citation14,Citation27]. In the ‘L/V’ conformations, potential SLA dimerization via kissing loop interactions of the SSL have been suggested in both DENV and ZIKV SLAs [Citation14]. However, the significance of this potential interaction in flaviviruses remains unclear (see Box 3) [Citation60]. Interestingly, while the predicted SSL is relatively short (2–3 bp) in the mosquito-borne flaviviruses, it is predicted to be significantly longer (~20 bp) in the tick-borne flaviviruses [Citation58]. It is not yet clear what implication this would have on the ability of SLA to arrange into the putative ‘L/V’ conformation and interact with NS5. Nonetheless, these advances in our understanding of the tertiary structure of SLA and its interactions with NS5 have implications for both negative- and positive-strand RNA synthesis, as well as capping, which will be discussed in detail in the following subsections.
NS5 interactions with SLA and the 3´SL promote negative-strand RNA synthesis
Recruitment of NS5 by SLA has repeatedly been demonstrated to be critical for replication of the viral genome, but precisely how NS5 localization to SLA at the 5´ end of the viral genome facilitates the initiation of negative-strand RNA synthesis at the 3´ terminus of the positive-sense genomic RNA remains unclear (). Based on recent data, we envision several models for the initiation of negative-strand RNA synthesis, two of which are SLA-centric, while the latter two additionally rely on interactions with the 3´SL ().
Figure 3. Models for the initiation of negative-strand RNA synthesis.
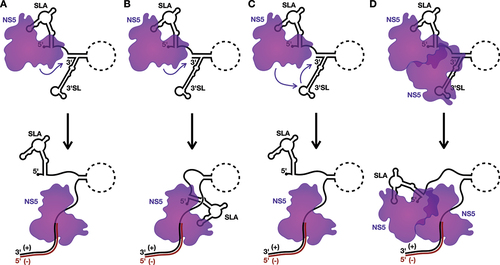
As a consequence of the ‘L/V’ conformation of SLA, the bound NS5 appears to sit below the SLA structure, where it would be proximal to the 3´ terminus as a consequence of genome cyclization [Citation14,Citation15,Citation60]. As such, in the first model, NS5 could initiate negative-strand RNA synthesis directly off the partially unwound 3´ terminus of the positive-strand (). However, in this first model, transfer of the NS5 from SLA to the 3´ terminus of the positive-strand RNA could potentially create an opening for reinitiation of translation at the now ‘free’ 5´ terminus of the viral genome [Citation34,Citation35]. Thus, in the second model, NS5 remains bound to SLA as it synthesizes the negative-strand, only releasing SLA when it needs to be unwound for completion of the dsRNA replicative intermediate (). Importantly, this continued engagement with the 5´ terminus during RNA synthesis from the 3´ terminus is not unprecedented, as several viral RdRps, including those of coronaviruses and influenza viruses, have been proposed to either directly or indirectly interact with the 5´ terminus of the template RNA as they synthesize the complementary strand [Citation66–69]. Since SLA remains bound to NS5 during the elongation phase of viral RNA synthesis in this second model, sequestration of the 5´ terminus could also help prevent reinitation of translation, which may be advantageous if compaction of the viral RNA into a RO is occurring concurrently with the initiation of negative-strand RNA synthesis.
However, these SLA-centric models do not directly account for the role of NS5–3´SL interactions in viral RNA replication [Citation50,Citation51,Citation70,Citation71]. Notably, the 3´SL also contains a conserved ‘AG’ dinucleotide in its TSL that has been shown to be critical for viral RNA replication and NS5 recruitment, and could facilitate NS5–3´SL binding similarly to NS5-SLA interactions [Citation14,Citation15,Citation27,Citation51,Citation58,Citation60,Citation62,Citation70,Citation71]. To account for this interaction, in the third model for the initiation of negative-strand RNA synthesis, the NS5 bound to SLA is transferred to the 3´SL due to the higher affinity of NS5 for the 3´SL over SLA following the cyclization-induced unwinding of SLB [Citation19]. Subsequently, the transferred NS5 could then initiate at the 3´ terminus to synthesize the negative-strand replicative intermediate () [Citation72]. However, this model has the same potential translation reinitiation drawback as the first model, which leads us to the final model, whereby the NS5 bound to SLA does not directly synthesize the negative-strand, but rather interacts with a second NS5 bound to the 3´SL, and facilitates rearrangement to allow this latter NS5 molecule to initiate negative-strand RNA synthesis in the context of an NS5 dimer (). Similarly to the second model, the 5´ terminus would thus remain associated with the SLA-bound NS5 during negative-strand RNA synthesis until this region requires unwinding for completion of the dsRNA replicative intermediate. While it is well accepted that flaviviral NS5 can dimerize (or even form higher-ordered oligomers), the orientation of a putative NS5 dimer in the initiation of negative-strand RNA synthesis remains unclear, as several NS5 dimer/oligomer orientations have been observed in crystal structures [Citation73–75]. While more experimentation will be needed to provide definitive support for any one of these four models, it seems clear that cis-acting RNA elements, including SLA and perhaps also 3´SL, as well as the genome cyclization sequences, play key roles in directing the initiation of negative-strand RNA synthesis in the flavivirus life cycle.
SLA may also have an active role in positive-strand RNA synthesis
Compared with negative-strand RNA synthesis, the requirements for positive-strand RNA synthesis are more poorly defined, although it seems increasingly likely that SLA also plays a central role in this process ( and ). The major difference between negative- and positive-strand RNA synthesis relates to the nature of the template RNA, which is single-stranded positive-sense genomic RNA and the dsRNA replicative intermediate, respectively. Although each end of the dsRNA replicative intermediate is biochemically similar and possesses all the features needed for the core replicase to act, there is a clear preference for the initiation of positive-strand RNA synthesis relative to negative-strand RNA synthesis [Citation56,Citation57,Citation76]. As discussed previously, since the specificity needed to maintain this preference does not come from the viral proteins involved, it is likely that features of the viral RNAs themselves are responsible for guiding this process. A recent study that used ribozymes to explore internal RNA folding in negative-strand replicative intermediates suggests that regions of the negative-strand in YFV, the related hepatitis C virus (HCV), and other positive-sense RNA viruses can be locally structured during viral replication, albeit at low frequency [Citation77]. Thus, perhaps in combination with terminal ‘breathing’ or a yet undiscovered active unwinding mechanism, SLA may be able to spontaneously reform at the 5´ terminus of the positive-strand in the dsRNA replicative intermediate [Citation78].
Figure 4. Models for the initiation of positive-strand RNA synthesis.
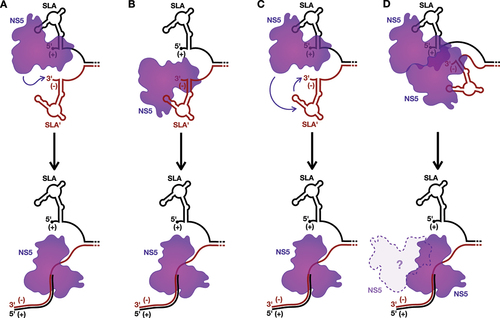
As such, SLA could also play a role in directing the initiation of positive-strand RNA synthesis, in parallel to the models discussed above for the initiation of negative-strand RNA synthesis (). For example, binding of NS5 to the re-folded SLA could stabilize the 5´ terminus of the positive-strand, resulting in a single-stranded 3´ overhang on the negative-strand that is necessary for NS3 to initiate unwinding of the dsRNA intermediate [Citation76]. Similarly to the first model for negative-strand RNA synthesis (), it is possible that the NS5 recruited to the reformed SLA could then directly interact with the 3´ terminus of the negative-strand to initiate positive-strand RNA synthesis from the dsRNA template (). Alternatively, akin to the role of 3´SL in the models for negative-strand RNA synthesis (), it is possible that the terminal structure at the 3´ end of the negative-strand, termed SLA´ herein, serves as the promoter for positive-strand RNA synthesis (). Interestingly, despite the presence of a couple alternative G:U wobble base pairs, SLA´ is predicted to essentially form a mirror image of SLA, with a base stem, SSL, and a conserved unpaired ‘AG motif’ in its TSL [Citation79]. This mirroring is somewhat unexpected, as in the related HCV, the structure of the 5´ terminus of the positive-strand and the 3´ terminus of the negative-strand possess completely independent architectures due to G-U wobble base pairing, and the latter has been demonstrated to serve as the promoter for positive-strand RNA synthesis [Citation80–82]. Furthermore, in support of a role for flaviviral SLA´ in positive-strand RNA synthesis, in vitro binding studies with JEV show that NS5 has a greater affinity for the 3´ terminus of the negative-strand than the 3´ terminus of the positive-strand [Citation83]. Moreover, in ZIKV, the terminal 105 nucleotides at the 3´ terminus of the negative-strand is important for efficient regulation of positive-strand RNA synthesis in vitro, a region which includes SLA´ [Citation84]. As such, it is possible that NS5 is directly recruited to SLA´ (), or transferred from SLA to SLA´ (), to initiate positive-strand RNA synthesis. Alternatively, the NS5 bound to SLA could interact with, or even aid in the recruitment of, a second NS5 bound to SLA´, to promote initiation of positive-strand RNA synthesis (). However, in contrast to the fourth model for negative-strand RNA synthesis (), it seems less clear during the elongation stage whether the NS5 bound to SLA would be displaced or remain bound to the NS5 carrying out positive-strand RNA synthesis, since the displaced positive-strand needs to exit the RO to maintain the largely uniform size of the flaviviral RO () [Citation4,Citation5]. Beyond these models, it is also possible that the SLA´ structure may simply serve to promote positive-strand RNA synthesis by reinforcing strand separation at the terminus of the dsRNA replicative intermediate. Regardless, both SLA and/or SLA´ would presumably need to reform for each subsequent round of positive-strand RNA synthesis, as this process results in regeneration of the dsRNA intermediate. Nevertheless, while the mechanism of initiating positive-strand RNA synthesis remains largely a black box, it is likely that SLA, and possibly also SLA´, help direct the process.
NS5-SLA interactions provide specificity for genome capping
In addition to its roles in viral RNA replication, SLA has also been implicated in the capping of newly-synthesized viral genomes (). While the RTPase activity of NS3 is responsible for reducing the 5´ triphosphate to a monophosphate moiety, the latter steps of the capping reaction are catalysed by NS5’s MTase activity, which adds a guanyl cap and then methylates at the N-7 and 2´-O positions of the 5´ cap. In both DENV and WNV, 2´-O-methylation (2´-OMe) of the A1 position of the genome is RNA structure-independent, and only dependent on a G at position 2, which stacks with the A1 base and aids in coordinating the Mg2+ ion bound to NS5 [Citation85,Citation86]. Together with the requirement of the NS5 RdRp to initiate RNA synthesis on a 3´-UC dinucleotide, these requirements for 2´-OMe contribute to the strict conservation of an AG dinucleotide at the 5´ terminus of the RNA [Citation57]. In contrast to 2´-OMe, efficient N-7 methylation in WNV appears to be SLA-dependent, with the interaction between the base stem and NS5 being of particular importance [Citation85]. NS5 binding to the ‘L/V’ conformation of SLA in DENV and ZIKV is consistent with a SLA-mediated capping model, with the capped 5´-tetranucleotide (5´-GpppAGUU) well accommodated in the catalytic pocket of the NS5 MTase domain () [Citation14,Citation15]. While the addition of the G cap itself has not been studied in the context of SLA, the positioning of the 5´ terminus in the MTase active site implies that this step is also likely to be SLA-mediated. Thus, beyond the importance of SLA-NS5 interactions in providing the specificity for regulating negative- and positive-strand RNA synthesis, these new interaction models also reveals a role for SLA-NS5 interactions in viral RNA capping.
What features of the genomic RNA provide specificity for genome packaging?
To date, virion assembly remains structurally and biochemically one of the least defined stages of the flavivirus life cycle [Citation87,Citation88]. In brief, the nascent viral genomic RNA must be localized to the assembly site (often located in close proximity to the RO pore), condensed, and encapsidated as it buds into the ER lumen, gaining a viral envelope studded with the viral pre-membrane and envelope proteins [Citation88]. However, at least in vitro, the capsid protein does not appear to have any specificity for the viral genomic RNA, raising the question as to how viral genomes are specifically packaged into assembling viral particles [Citation89]. Recent data has indicated that the viral NS2A protein may provide specificity for genome packaging, through specific interactions with the viral 3´ UTR [Citation28,Citation29]. In this section, we will discuss the RNA structures and sequences of the putative viral packaging signal, as well as long-range RNA-RNA interactions that likely facilitate compaction of the viral genome into the assembling virion.
NS2A–3´ UTR interactions facilitate packaging of flavivirus genomic RNAs
Having no clearly defined ‘packaging signal’ within the flaviviral RNA, the mechanism(s) surrounding specificity in genome packaging have remained elusive [Citation17]. However, interactions between NS2A and the 3´ UTR are necessary for genome packaging in ZIKV, DENV, and Kunjin virus (KUNV), suggesting that NS2A–3´ UTR interactions provide specificity to the virion assembly process () [Citation28,Citation29]. However, the 3´ UTR structures bound by NS2A appear to differ across these flaviviruses. Based on competitive pull-down experiments, ZIKV NS2A binds to SL-II specifically, while the last 285 nucleotides of the 3´ UTR (corresponding to DBI, DBII, sHP and the 3´SL) are needed for successful interaction with DENV NS2A, implying that DENV NS2A interacts with the overall RNA structure in this region rather than an individual structure [Citation28,Citation29]. Taken together, these findings imply that there is no singular conserved RNA structure that acts as the flaviviral packaging signal, but instead flavivirus NS2A proteins may have each divergently evolved to recognize RNA sequences and/or structures in the 3´ UTR as a means of providing specificity to the virion assembly process.
Beyond interacting with the 3´ UTR of the viral genome, NS2A is hypothesized to serve as a central hub at the virion assembly site due to its interactions with the other viral proteins required for assembly (including the capsid, pre-membrane, and envelope complex, as well as NS2B and NS3). The ability of NS2A to facilitate the co-localization of the viral genome and capsid proteins would bring them into close proximity, possibly driving genome condensation and formation of the nucleocapsid [Citation28,Citation29,Citation90]. The NS2A–3´ UTR interaction could thereby circumvent the capsid protein’s apparent lack of specificity for the viral RNA genome, and thus serve as a noncanonical packaging signal for the flavivirus genomic RNA [Citation91].
Long-range RNA-RNA interactions likely facilitate compaction of the viral RNA into the assembling virion
The structure of the packaged genome has not been elucidated in recent high-resolution cryo-electron microscopy studies of flaviviral virions, but the need for compaction of the viral genomic RNA to package it inside the virion is clear [Citation92]. For instance, the hydrodynamic radius of the DENV2 genome is ~45 nm in the absence of viral and host proteins, while its virion has a radius of ~25 nm, meaning that the viral genome needs to be reduced to <17% of its volume to be successfully packaged [Citation30]. In addition, the need to compact the viral genome is not limited to virion assembly, but is also a requirement for RO biogenesis, as discussed above [Citation45]. To solve this dilemma, the genomes of flaviviruses have evolved to have numerous long-range RNA-RNA interactions (generally greater than 0.5 kb of separation between the interacting elements) () [Citation30–33]. In infected cells, only 34% of RNA interactions are classified as long-range, while this number rises to 77% when looking solely at RNA within the virion [Citation33]. Interestingly, there appears to be high plasticity in these interactions across flaviviruses. Rather than utilizing the same long-range RNA-RNA interaction in a particular region of the genome, different flaviviruses have often evolved a comparable interaction close by that promotes genome compaction [Citation30,Citation33]. Even within a single strain, a given RNA element is more likely to have an alternative long-range RNA interaction partner within a packaged virion than within an infected cell, pointing towards an overall structural heterogeneity to compact the genome into the assembling virion [Citation32]. Despite this heterogeneity and plasticity in the RNA-RNA interactions used to condense the genome, the overall degree of these interactions appears to be tightly balanced, as disruption of several individual long-range RNA-RNA interactions in DENV was sufficient to impair viral fitness and increase the genome volume by ~30–50% [Citation30]. Interestingly, while the DENV capsid protein does not form specific interactions with the genomic RNA, it has been shown to preferentially bind to regions of the genome involved in these long-range RNA-RNA interactions within the virion [Citation31]. Thus, genome compaction could also be assisted during virion assembly by capsid’s RNA chaperone activity. This relationship between compaction and encapsidation would be mutually synergistic, with long-range RNA-RNA interactions in the genome facilitating capsid binding, while capsid in turn aids in their stabilization and thus promotes long-range RNA-RNA interactions [Citation89]. In addition to capsid, cellular RBPs also contribute to the stability and remodelling of these long-range RNA-RNA interactions (see Box 4). Overall, the formation of long-range RNA-RNA interactions is a highly plastic, but critical step in genome compaction and encapsidation.
Taken together, both specific interactions between the NS2A protein and the 3´ UTR of the viral genomic RNA (that likely serves as the viral packaging signal), as well as more loosely defined long-range RNA-RNA interactions across the viral genomic RNA (which facilitate genome compaction and encapsidation), help drive genome packaging in the flavivirus life cycle.
Concluding remarks
In summary, three major themes are starting to emerge regarding the general roles played by viral RNA structure in the flavivirus life cycle, with parallels in a diverse range of RNA viruses. These themes include providing a layer of specificity across the viral life cycle, increasing the functional capacity of the genome, and providing a mechanism to support genome compaction.
The first of these themes is that RNA structure provides specificity to a tightly regulated system in which the viral proteins generally act non-specifically. During translation, the viral genomic RNA provides specificity via the cHP, which ensures proper start codon selection [Citation22]. Interestingly, alphaviruses utilize a similar mechanism, whereby a stable downstream stem-loop on the subgenomic mRNA is hypothesized to form an RNA trap and stall ribosomal scanning at the start codon for production of the viral structural proteins [Citation20,Citation101,Citation102]. Since stable stem-loop structures shortly downstream of the start codon lower the requirement for optimal start codon context in eukaryotic translation, it is not surprising that this mechanism of start codon selection extends broadly across RNA viruses [Citation103]. Additionally, during RNA replication, SLA provides specificity to negative- and positive-strand RNA synthesis, and the capping reaction through its interactions with NS5, the viral RdRp and MTase. Similarly, the poliovirus 5´ cloverleaf structure is also known to recruit the viral RdRp and promote both negative- and positive-strand RNA synthesis [Citation104]. Furthermore, while the mechanism of positive-strand RNA synthesis initiation is largely unknown in flaviviruses, RNA structures at the 3´ terminus of the negative-strand of the closely related HCV also serve as the promoter for initiating positive-strand RNA synthesis [Citation80–82]. Beyond positive-sense RNA viruses, the 5´ and 3´ termini of the influenza virus genome segments (a multi-segmented negative-sense RNA virus) form a corkscrew-like structure that recruits the viral RdRp [Citation105]. Thus, the concept of 5´ and 3´ terminal RNA structures directing viral RNA synthesis is also likely to apply broadly across RNA viruses. Finally, interactions between NS2A and structures in the 3´ UTR of flaviviruses provide specificity for genome packaging. Analogous to differences in the packaging signals in the related ZIKV and DENV, distinct betacoronaviruses have vastly different structural requirements in their packaging signals, ranging from a 95-nt structure in mouse hepatitis virus to a 1.4 kb region in severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), further illustrating the diversity of packaging signals even among highly related viruses [Citation106–108]. Conversely, selective genome packaging of each of the eight influenza virus genome segments relies upon RNA-RNA interactions between the packaging signals of the different genome segments, providing an elegant solution to a complex packaging specificity problem [Citation109]. Taken together, these examples help to demonstrate how viral RNA structure provides specificity, not only in flaviviruses but across RNA viruses more broadly.
The second theme regarding the role of RNA structure in the flavivirus life cycle is to increase the functional capacity of their small RNA genomes, in which the ~11 kb RNA must encode the 10 viral proteins and all the functional sequence/structural information needed to carry out the complex infectious cycle. For example, encoding a cap-independent mechanism for translation initiation provides a layer of redundancy in times of stress (see Box 1). Possessing such a cap-independent mechanism is incredibly valuable to RNA viruses as infection generally leads to downregulation of cellular cap-dependent translation due to interferon-induced phosphorylation of the eukaryotic initiation factor 2α [Citation40,Citation41,Citation43,Citation44]. As such, it is not surprising that many RNA viruses, including poliovirus and HCV, encode IRES structures that allow them to recruit the ribosome in a cap-independent manner using a fraction of the eukaryotic translation initiation factors [Citation110]. Another example of how RNA structure increases the functional capacity of flavivirus genomes are their mechanisms for translational shutoff. In a parallel manner to NS5-SLA interactions in the flavivirus life cycle, the cloverleaf structure at the 5´ terminus of the poliovirus genome also participates in translational shutoff and the switch from translation to replication [Citation67]. Additionally, like the cyclization elements of flaviviruses, interactions between the poly-C binding protein 2 and the poliovirus cloverleaf structure at the 5´ terminus and poly-A binding protein at the 3´ terminus aid in genome circularization for viral RNA replication [Citation67]. Mechanisms for genome circularization are pervasive across diverse RNA viruses, representing a common approach for increasing the functional capacity of their genomes [Citation66–69]. Interestingly, these genome circularization mechanisms inhibit viral translation in both flaviviruses and picornaviruses [Citation26,Citation67]. This observation may also be true for coronaviruses, where nucleocapsid-mediated circularization promotes negative-strand RNA synthesis, although the mechanism is still unclear [Citation68]. As such, circularization of the genomes of positive-sense RNA viruses may represent a common mechanism for mediating translational shut-off. Beyond the structures covered in this review, there are several well-characterized examples of alternative RNA conformations that aid in increasing the functional capacity of viral RNAs. These include the flaviviral pseudoknots, which promote sfRNA production to evade antiviral responses; the subgenomic mRNA promoter region of togaviruses; and the transcriptional regulatory sequences in coronaviruses that mediate discontinuous transcription for the production of subgenomic mRNAs [Citation24–48–Citation101,Citation102–113]. Thus, it seems clear that RNA secondary and tertiary structures allow RNA viruses to increase the functional capacity of their genomes beyond that of their limited coding capacity.
Finally, the third major theme regarding the role of RNA structure in the viral life cycle is compaction of the viral genome. In all RNA viruses, there are two major stages of the viral life cycle that require viral genome compaction: 1) RO biogenesis, in which the viral RNA is condensed together with viral replication proteins into a replication compartment; and 2) virion assembly, in which the viral genome is condensed into the nucleocapsid and infectious viral particle [Citation30,Citation45]. In flaviviruses, it appears that genome compaction is driven by long-range RNA-RNA interactions across the viral genomic RNA [Citation30–33]. While similar whole-genome analyses have been performed in other RNA viruses, these studies have largely not included information on long-range RNA-RNA interactions [Citation114]. However, a recent study of SARS-CoV-2 genome structure revealed numerous long-range RNA-RNA interactions within the virion, resulting in an unentangled globule conformation of the genome [Citation115]. It is unclear at present whether these long-range RNA-RNA interactions promote compaction of the viral genome, as they do in flaviviruses, or are a consequence of compaction of the viral genome into the assembling virion. Nonetheless, RNA secondary and tertiary structures are likely to play a central role in the process of genome compaction for both RO biogenesis and virion assembly.
Despite recent advances in our understanding of the roles of RNA structure in the flavivirus life cycle, there remain a number of outstanding questions and challenges in elucidating the precise roles of RNA structures during infection. For example, to further interrogate which of the aforementioned models for negative- and positive-strand RNA synthesis best reflects the role of SLA-NS5 interactions, it will be necessary to develop experimental systems that uncouple the potential roles of SLA in the switch from translation to replication, negative- and positive-strand RNA synthesis, and genome capping (). The complexity needed for such a system has thus far limited efforts to further interrogate each individual role of SLA, and the overlaps in sequence and structural requirements for SLA across these roles renders the application of traditional stem disruption and rescue approaches for assessing function very difficult in the context of the viral life cycle. Moving forward, creative approaches that allow the isolation of one or more of these roles, analogous to the pIRO system for analysing requirements for RO biogenesis independently of viral RNA replication, will be needed [Citation4]. While such systems may not provide direct equivalents to infection, they can provide insight into the players involved and will help to refine models for viral translation, replication, and packaging in the flavivirus life cycle.
Lastly, that a single piece of viral genomic RNA could play such myriad roles across the viral life cycle speaks to the highly sophisticated nature of flaviviruses. Moreover, the themes discussed herein, including specificity, functional capacity, and genome compaction extend beyond flaviviruses, with parallel roles described for RNA structures in the life cycles of a diverse array of RNA viruses.
Author contributions
Q.H.A. and S.M.S. conceived of the article. Q.H.A., B.N.L., A.B.W., R.F.K., and H.C.H.H. researched data for the article. All authors contributed to the discussion of content, as well as drafting and editing of the manuscript.
Highlights
RNA structures provide a layer of specificity across the flavivirus life cycle.
RNA sequence and structural elements, and their dynamics, increase the functional capacity of flaviviral genomes.
SLA is a critical element in the switch from translation to replication, negative- and positive-strand RNA synthesis, and capping of newly-synthesized genomic RNAs.
New models for interactions between SLA and NS5 shed light on potential mechanisms for the initiation of negative- and positive-strand RNA synthesis.
Long-range RNA-RNA interactions in flavivirus genomes are highly plastic and critical for compaction of the viral genome into replication organelles and infectious viral particles.
Acknowledgments
The authors apologize to all colleagues whose work they could not cite due to space limitations. Q.H.A. would like to thank the Frederick Banting and Charles Best Canada Graduate Scholarships – Doctoral Award (CGS-D) and Tomlinson Doctoral Scholarship for graduate support. B.N.L. and A.B.W. were supported by Canada Graduate Scholarship – Master’s Awards (CGS-M) from the Natural Sciences and Engineering Research Council (NSERC).
Disclosure statement
No potential conflict of interest was reported by the author(s).
Data availability statement
The authors confirm that the data supporting the findings of this study are available within the article.
Additional information
Funding
References
- Gould EA, Solomon T. Pathogenic flaviviruses. Lancet. 2008;371(9611):500–509. doi: 10.1016/S0140-6736(08)60238-X PubMed PMID: 18262042.
- Iwamura T, Guzman-Holst A, Murray KA. Accelerating invasion potential of disease vector Aedes aegypti under climate change. Nat Commun. 2020;11(1):2130. doi: 10.1038/s41467-020-16010-4 Epub 20200501. PubMed PMID: 32358588; PubMed Central PMCID: PMC7195482.
- Pierson TC, Diamond MS. The continued threat of emerging flaviviruses. Nat Microbiol. 2020;5(6):796–812. doi: 10.1038/s41564-020-0714-0 Epub 20200504. PubMed PMID: 32367055; PubMed Central PMCID: PMC7696730.
- Cortese M, Goellner S, Acosta EG, et al. Ultrastructural characterization of Zika Virus replication factories. Cell Rep. 2017;18(9):2113–2123. doi: 10.1016/j.celrep.2017.02.014 PubMed PMID: 28249158; PubMed Central PMCID: PMC5340982.
- Welsch S, Miller S, Romero-Brey I, Merz A, Bleck CK, Walther P, et al. Composition and three-dimensional architecture of the dengue virus replication and assembly sites. Cell Host Microbe. 2009;5(4):365–375. doi: 10.1016/j.chom.2009.03.007 PubMed PMID: 19380115; PubMed Central PMCID: PMC7103389.
- Spitale RC, Crisalli P, Flynn RA, et al. RNA SHAPE analysis in living cells. Nat Chem Biol. 2013;9(1):18–20. doi: 10.1038/nchembio.1131 Epub 2012 Nov 28. PubMed PMID: 23178934; PubMed Central PMCID: PMC3706714.
- Weeks KM. Advances in RNA structure analysis by chemical probing. Curr Opin Struct Biol. 2010;20(3):295–304. doi: 10.1016/j.sbi.2010.04.001 Epub 2010 Jun 08. PubMed PMID: 20447823; PubMed Central PMCID: PMC2916962.
- Wells SE, Hughes JM, Igel AH, Ares M Jr. Use of dimethyl sulfate to probe RNA structure in vivo. Methods Enzymol. 2000;318:479–493. doi: 10.1016/s0076-6879(00)18071-1 PubMed PMID: 10890007.
- Siegfried NA, Busan S, Rice GM, Nelson JA, Weeks KM. RNA motif discovery by SHAPE and mutational profiling (SHAPE-MaP). Nat Methods. 2014;11(9):959–965. doi: 10.1038/nmeth.3029 Epub 20140713. PubMed PMID: 25028896; PubMed Central PMCID: PMC4259394.
- Zubradt M, Gupta P, Persad S, et al. DMS-MaPseq for genome-wide or targeted RNA structure probing in vivo. Nat Methods. 2017;14(1):75–82. doi: 10.1038/nmeth.4057 Epub 20161107. PubMed PMID: 27819661; PubMed Central PMCID: PMC5508988.
- Lu Z, Zhang QC, Lee B, et al. RNA duplex map in living cells reveals higher-order transcriptome structure. Cell. 2016;165(5):1267–1279. doi: 10.1016/j.cell.2016.04.028 Epub 20160512. PubMed PMID: 27180905; PubMed Central PMCID: PMC5029792.
- Ziv O, Gabryelska MM, Lun ATL, et al. COMRADES determines in vivo RNA structures and interactions. Nat Methods. 2018;15(10):785–788. doi: 10.1038/s41592-018-0121-0 Epub 20180910. PubMed PMID: 30202058; PubMed Central PMCID: PMC6168409.
- Morandi E, Manfredonia I, Simon LM, et al. Genome-scale deconvolution of RNA structure ensembles. Nat Methods. 2021;18(3):249–252. doi: 10.1038/s41592-021-01075-w Epub 20210222. PubMed PMID: 33619392.
- Lee E, Bujalowski PJ, Teramoto T, et al. Structures of flavivirus RNA promoters suggest two binding modes with NS5 polymerase. Nat Commun. 2021;12(1):2530. doi: 10.1038/s41467-021-22846-1 Epub 20210505. PubMed PMID: 33953197; PubMed Central PMCID: PMC8100141.
- Osawa T, Aoki M, Ehara H, Sekine SI. Structures of dengue virus RNA replicase complexes. Mol Cell. 2023;83(15):2781–91 e4. doi: 10.1016/j.molcel.2023.06.023 Epub 20230720. PubMed PMID: 37478848.
- Spitale RC, Incarnato D. Probing the dynamic RNA structurome and its functions. Nat Rev Genet. 2023;24(3):178–196. doi: 10.1038/s41576-022-00546-w Epub 20221108. PubMed PMID: 36348050; PubMed Central PMCID: PMC9644009.
- Mazeaud C, Freppel W, Chatel-Chaix L. The multiples fates of the flavivirus RNA genome during pathogenesis. Front Genet. 2018;9:595. doi: 10.3389/fgene.2018.00595 Epub 20181204. PubMed PMID: 30564270; PubMed Central PMCID: PMC6288177.
- Filomatori CV, Lodeiro MF, Alvarez DE, Samsa MM, Pietrasanta L, Gamarnik AV. A 5’ RNA element promotes dengue virus RNA synthesis on a circular genome. Genes Dev. 2006;20(16):2238–2249. doi: 10.1101/gad.1444206 Epub 2006 Aug 03. PubMed PMID: 16882970; PubMed Central PMCID: PMC1553207.
- Liu ZY, Li XF, Jiang T, Deng YQ, Ye Q, Zhao H, et al. Viral RNA switch mediates the dynamic control of flavivirus replicase recruitment by genome cyclization. Elife. 2016;5:e17636. doi: 10.7554/eLife.17636 Epub 2016 Oct 04. PubMed PMID: 27692070; PubMed Central PMCID: PMC5101012.
- Alvarez DE, Lodeiro MF, Luduena SJ, Pietrasanta LI, Gamarnik AV. Long-range RNA-RNA interactions circularize the dengue virus genome. J Virol. 2005;79(11):6631–6643. doi: 10.1128/JVI.79.11.6631-6643.2005 Epub 2005 Jun 14. PubMed PMID: 15890901; PubMed Central PMCID: PMC1112138.
- Clyde K, Barrera J, Harris E. The capsid-coding region hairpin element (cHP) is a critical determinant of dengue virus and West Nile virus RNA synthesis. Virology. 2008;379(2):314–323. doi: 10.1016/j.virol.2008.06.034 Epub 20080803. PubMed PMID: 18676000; PubMed Central PMCID: PMC2628549.
- Clyde K, Harris E. RNA secondary structure in the coding region of dengue virus type 2 directs translation start codon selection and is required for viral replication. J Virol. 2006;80(5):2170–2182. doi: 10.1128/JVI.80.5.2170-2182.2006 Epub 2006 Feb 14. PubMed PMID: 16474125; PubMed Central PMCID: PMC1395379.
- Friebe P, Harris E. Interplay of RNA elements in the dengue virus 5‘and 3’ ends required for viral RNA replication. J Virol. 2010;84(12):6103–6118. doi: 10.1128/JVI.02042-09 Epub 20100331. PubMed PMID: 20357095; PubMed Central PMCID: PMC2876622.
- Villordo SM, Carballeda JM, Filomatori CV, et al. RNA structure duplications and flavivirus host adaptation. Trends Microbiol. 2016;24(4):270–283. doi: 10.1016/j.tim.2016.01.002 Epub 2016 Feb 07. PubMed PMID: 26850219; PubMed Central PMCID: PMC4808370.
- Filomatori CV, Carballeda JM, Villordo SM, et al. Dengue virus genomic variation associated with mosquito adaptation defines the pattern of viral non-coding RNAs and fitness in human cells. PLOS Pathog. 2017;13(3):e1006265. doi: 10.1371/journal.ppat.1006265 Epub 2017 Mar 07. PubMed PMID: 28264033; PubMed Central PMCID: PMC5354447.
- Villordo SM, Alvarez DE, Gamarnik AV. A balance between circular and linear forms of the dengue virus genome is crucial for viral replication. RNA. 2010;16(12):2325–2335. doi: 10.1261/rna.2120410 Epub 2010 Oct 29. PubMed PMID: 20980673; PubMed Central PMCID: PMC2995394.
- Filomatori CV, Iglesias NG, Villordo SM, et al. RNA sequences and structures required for the recruitment and activity of the dengue virus polymerase. J Biol Chem. 2011;286(9):6929–6939. doi: 10.1074/jbc.M110.162289 Epub 2010 Dec 25. PubMed PMID: 21183683; PubMed Central PMCID: PMC3044948.
- Xie X, Zou J, Zhang X, Zhou Y, Routh AL, Kang C, et al. Dengue NS2A protein orchestrates virus assembly. Cell Host Microbe. 2019;26(5):606–22 e8. doi: 10.1016/j.chom.2019.09.015 Epub 20191017. PubMed PMID: 31631053.
- Zhang X, Xie X, Xia H, Zou J, Huang L, Popov VL, et al. Zika Virus NS2A-Mediated Virion assembly. MBio. 2019;10(5):10–128. doi: 10.1128/mBio.02375-19 Epub 20191029. PubMed PMID: 31662457; PubMed Central PMCID: PMC6819661.
- Boerneke MA, Gokhale NS, Horner SM, et al. Structure-first identification of RNA elements that regulate dengue virus genome architecture and replication. Proc Natl Acad Sci U S A. 2023;120(15):e2217053120. doi: 10.1073/pnas.2217053120 Epub 20230403. PubMed PMID: 37011200; PubMed Central PMCID: PMC10104495.
- Boon PLS, Martins AS, Lim XN, Enguita FJ, Santos NC, Bond PJ, et al. Dengue virus capsid protein facilitates genome compaction and packaging. Int J Mol Sci. 2023;24(9):8158. doi: 10.3390/ijms24098158 Epub 20230502. PubMed PMID: 37175867; PubMed Central PMCID: PMC10179140.
- Huber RG, Lim XN, Ng WC, et al. Structure mapping of dengue and Zika viruses reveals functional long-range interactions. Nat Commun. 2019;10(1):1408. doi: 10.1038/s41467-019-09391-8 Epub 20190329. PubMed PMID: 30926818; PubMed Central PMCID: PMC6441010.
- Li P, Wei Y, Mei M, Tang L, Sun L, Huang W, et al. Integrative analysis of Zika Virus Genome RNA structure reveals critical determinants of viral infectivity. Cell Host Microbe. 2018;24(6):875–86 e5. doi: 10.1016/j.chom.2018.10.011 Epub 20181121. PubMed PMID: 30472207.
- Fajardo T, Sanford TJ, Mears HV, et al. The flavivirus polymerase NS5 regulates translation of viral genomic RNA. Nucleic Acids Res. 2020;48(9):5081–5093. doi: 10.1093/nar/gkaa242 PubMed PMID: 32313955; PubMed Central PMCID: PMC7229856.
- Sanford TJ, Mears HV, Fajardo T, et al. Circularization of flavivirus genomic RNA inhibits de novo translation initiation. Nucleic Acids Res. 2019;47(18):9789–9802. doi: 10.1093/nar/gkz686 PubMed PMID: 31392996; PubMed Central PMCID: PMC6765113.
- Mishra B, Balaji A, Beesetti H, Swaminathan S, Aduri R. The RNA secondary structural variation in the cyclization elements of the dengue genome and the possible implications in pathogenicity. Virusdisease. 2020;31(3):299–307. doi: 10.1007/s13337-020-00615-w Epub 20200730. PubMed PMID: 32904896; PubMed Central PMCID: PMC7458965.
- Samsa MM, Mondotte JA, Caramelo JJ, et al. Uncoupling cis-Acting RNA elements from coding sequences revealed a requirement of the N-terminal region of dengue virus capsid protein in virus particle formation. J Virol. 2012;86(2):1046–1058. doi: 10.1128/JVI.05431-11 Epub 20111109. PubMed PMID: 22072762; PubMed Central PMCID: PMC3255831.
- Patkar CG, Jones CT, Chang YH, et al. Functional requirements of the yellow fever virus capsid protein. J Virol. 2007;81(12):6471–6481. doi: 10.1128/JVI.02120-06 PubMed PMID: 17526891; PubMed Central PMCID: PMC1900127.
- Berzal-Herranz A, Berzal-Herranz B, Ramos-Lorente SE, Romero-Lopez C. The genomic 3’ UTR of flaviviruses is a translation initiation enhancer. Int J Mol Sci. 2022;23(15):8604. doi: 10.3390/ijms23158604 Epub 20220803. PubMed PMID: 35955738; PubMed Central PMCID: PMC9369090.
- Edgil D, Polacek C, Harris E. Dengue virus utilizes a novel strategy for translation initiation when cap-dependent translation is inhibited. J Virol. 2006;80(6):2976–2986. doi: 10.1128/JVI.80.6.2976-2986.2006 PubMed PMID: 16501107; PubMed Central PMCID: PMC1395423.
- Wang T, Merits A, Wu Y, et al. cis-acting sequences and secondary structures in untranslated regions of duck tembusu virus RNA are important for cap-independent translation and viral proliferation. J Virol. 2020;94(16):10–128. doi: 10.1128/JVI.00906-20 Epub 20200730. PubMed PMID: 32522848; PubMed Central PMCID: PMC7394898.
- Fernandez-Garcia L, Angulo J, Ramos H, Barrera A, Pino K, Vera-Otarola J, et al. The internal ribosome entry site of the Dengue virus mRNA is active when cap-dependent translation initiation is inhibited. J Virol. 2021;95(5):10–128. doi: 10.1128/JVI.01998-20 Epub 20201209. PubMed PMID: 33298544; PubMed Central PMCID: PMC8092825.
- Song Y, Mugavero J, Stauft CB, Wimmer E. Dengue and Zika Virus 5’ untranslated regions harbor internal ribosomal entry site functions. MBio. 2019;10(2):10–128. doi: 10.1128/mBio.00459-19 Epub 20190409. PubMed PMID: 30967466; PubMed Central PMCID: PMC6456755.
- Roth H, Magg V, Uch F, Mutz P, Klein P, Haneke K, et al. Flavivirus infection uncouples translation suppression from cellular stress responses. MBio. 2017;8(1):10–128. doi: 10.1128/mBio.02150-16 Epub 20170110. PubMed PMID: 28074025; PubMed Central PMCID: PMC5225315.
- Sagan SM, Weber SC. Let’s phase it: viruses are master architects of biomolecular condensates. Trends Biochem Sci. 2023;48(3):229–243. doi: 10.1016/j.tibs.2022.09.008 Epub 20221019. PubMed PMID: 36272892.
- Barton DJ, Morasco BJ, Flanegan JB. Translating ribosomes inhibit poliovirus negative-strand RNA synthesis. J Virol. 1999;73(12):10104–10112. doi: 10.1128/JVI.73.12.10104-10112.1999 PubMed PMID: 10559325; PubMed Central PMCID: PMC113062.
- De Falco L, Silva NM, Santos NC, Huber RG, Martins IC. The Pseudo-circular genomes of flaviviruses: structures, mechanisms, and functions of circularization. Cells. 2021;10(3):642. doi: 10.3390/cells10030642 Epub 20210313. PubMed PMID: 33805761; PubMed Central PMCID: PMC7999817.
- de Borba L, Villordo SM, Marsico FL, Carballeda JM, Filomatori CV, Gebhard LG, et al. RNA structure duplication in the dengue virus 3′ UTR: redundancy or host specificity? MBio. 2019;10(1). doi: 10.1128/mBio.02506-18 Epub 2019 Jan 10. PubMed PMID: 30622191; PubMed Central PMCID: PMC6325252.
- Meyer A, Freier M, Schmidt T, Rostowski K, Zwoch J, Lilie H, et al. An RNA thermometer activity of the West Nile Virus Genomic 3′-terminal stem-loop element modulates viral replication efficiency during host switching. Viruses. 2020;12(1):104. doi: 10.3390/v12010104 Epub 20200115. PubMed PMID: 31952291; PubMed Central PMCID: PMC7019923.
- Wang S, Chan KWK, Tan MJA, Flory C, Luo D, Lescar J, et al. A conserved arginine in NS5 binds genomic 3’ stem-loop RNA for primer-independent initiation of flavivirus RNA replication. RNA. 2022;28(2):177–193. doi: 10.1261/rna.078949.121 Epub 20211110. PubMed PMID: 34759006; PubMed Central PMCID: PMC8906541.
- Hodge K, Tunghirun C, Kamkaew M, et al. Identification of a conserved RNA-dependent RNA polymerase (RdRp)-RNA interface required for flaviviral replication. J Biol Chem. 2016;291(33):17437–17449. doi: 10.1074/jbc.M116.724013 Epub 20160622. PubMed PMID: 27334920; PubMed Central PMCID: PMC5016140.
- Livingston NM, Kwon J, Valera O, Saba JA, Sinha NK, Reddy P, et al. Bursting translation on single mRnas in live cells. Mol Cell. 2023;83(13):2276–89.e11. doi: 10.1016/j.molcel.2023.05.019 Epub 20230616. PubMed PMID: 37329884; PubMed Central PMCID: PMC10330622.
- Cerikan B, Goellner S, Neufeldt CJ, Haselmann U, Mulder K, Chatel-Chaix L, et al. A non-replicative role of the 3’ terminal sequence of the dengue virus genome in membranous replication organelle formation. Cell Rep. 2020;32(1):107859. doi: 10.1016/j.celrep.2020.107859 PubMed PMID: 32640225; PubMed Central PMCID: PMC7351112.
- Mazeaud C, Pfister S, Owen JE, et al. Zika virus remodels and hijacks IGF2BP2 ribonucleoprotein complex to promote viral replication organelle biogenesis. bioRxiv. 2023:2023–12. doi: 10.1101/2023.12.08.570783
- Burke JM, Ratnayake OC, Watkins JM, et al. G3BP1-dependent condensation of translationally inactive viral RNAs antagonizes infection. Sci Adv. 2024;10(5):eadk8152. doi: 10.1126/sciadv.adk8152 Epub 20240131. PubMed PMID: 38295168; PubMed Central PMCID: PMC10830107.
- Barnard TR, Wang AB, Sagan SM. A highly sensitive strand-specific multiplex RT-qPCR assay for quantitation of Zika virus replication. J Virol Methods. 2022;307:114556. doi: 10.1016/j.jviromet.2022.114556 Epub 20220530. PubMed PMID: 35654259.
- Selisko B, Potisopon S, Agred R, et al. Molecular basis for nucleotide conservation at the ends of the dengue virus genome. PLOS Pathog. 2012;8(9):e1002912. doi: 10.1371/journal.ppat.1002912 Epub 20120913. PubMed PMID: 23028313; PubMed Central PMCID: PMC3441707.
- Upstone L, Colley R, Harris M, Goonawardane N. Functional characterization of 5’ untranslated region (UTR) secondary RNA structures in the replication of tick-borne encephalitis virus in mammalian cells. PLOS Negl Trop Dis. 2023;17(1):e0011098. doi: 10.1371/journal.pntd.0011098 Epub 20230123. PubMed PMID: 36689554; PubMed Central PMCID: PMC9894543.
- Lodeiro MF, Filomatori CV, Gamarnik AV. Structural and functional studies of the promoter element for dengue virus RNA replication. J Virol. 2009;83(2):993–1008. doi: 10.1128/JVI.01647-08 Epub 20081112. PubMed PMID: 19004935; PubMed Central PMCID: PMC2612346.
- Sun YT, Varani G. Structure of the dengue virus RNA promoter. RNA. 2022;28(9):1210–1223. doi: 10.1261/rna.079197.122 Epub 20220624. PubMed PMID: 35750488; PubMed Central PMCID: PMC9380747.
- Bujalowski PJ, Bujalowski W, Choi KH. Identification of the viral RNA promoter stem loop a (SLA)-binding site on Zika virus polymerase NS5. Sci Rep. 2020;10(1):13306. doi: 10.1038/s41598-020-70094-y Epub 20200806. PubMed PMID: 32764551; PubMed Central PMCID: PMC7413259.
- Dong H, Zhang B, Shi PY. Terminal structures of West Nile virus genomic RNA and their interactions with viral NS5 protein. Virology. 2008;381(1):123–135. doi: 10.1016/j.virol.2008.07.040 Epub 20080916. PubMed PMID: 18799181.
- Bujalowski PJ, Bujalowski W, Choi KH, et al. Interactions between the dengue virus polymerase NS5 and Stem-Loop a. J Virol. 2017;91(11):10–128. doi: 10.1128/JVI.00047-17 Epub 20170512. PubMed PMID: 28356528; PubMed Central PMCID: PMC5432874.
- Yu L, Nomaguchi M, Padmanabhan R, Markoff L. Specific requirements for elements of the 5‘and 3’ terminal regions in flavivirus RNA synthesis and viral replication. Virology. 2008;374(1):170–185. doi: 10.1016/j.virol.2007.12.035 Epub 20080129. PubMed PMID: 18234265; PubMed Central PMCID: PMC3368002.
- Lott WB, Doran MR. Do RNA viruses require genome cyclisation for replication? Trends Biochem Sci. 2013;38(7):350–355. doi: 10.1016/j.tibs.2013.04.005 Epub 20130612. PubMed PMID: 23768999.
- Cong Y, Ulasli M, Schepers H, et al. Nucleocapsid protein recruitment to replication-transcription complexes plays a crucial role in coronaviral life cycle. J Virol. 2020;94(4). doi: 10.1128/JVI.01925-19 Epub 20200131. PubMed PMID: 31776274; PubMed Central PMCID: PMC6997762.
- Gottipati K, McNeme SC, Tipo J, White MA, Choi KH. Structural basis for cloverleaf RNA-initiated viral genome replication. Nucleic Acids Res. 2023;51(16):8850–8863. doi: 10.1093/nar/gkad618 PubMed PMID: 37486760; PubMed Central PMCID: PMC10484678.
- Lo CY, Tsai TL, Lin CN, Lin CH, Wu HY. Interaction of coronavirus nucleocapsid protein with the 5’- and 3’-ends of the coronavirus genome is involved in genome circularization and negative-strand RNA synthesis. FEBS J. 2019;286(16):3222–3239. doi: 10.1111/febs.14863 Epub 20190508. PubMed PMID: 31034708; PubMed Central PMCID: PMC7164124.
- Pflug A, Guilligay D, Reich S, et al. Structure of influenza a polymerase bound to the viral RNA promoter. Nature. 2014;516(7531):355–360. doi: 10.1038/nature14008 Epub 20141119. PubMed PMID: 25409142.
- Chen CJ, Kuo MD, Chien LJ, et al. RNA-protein interactions: involvement of NS3, NS5, and 3’ noncoding regions of Japanese encephalitis virus genomic RNA. J Virol. 1997;71(5):3466–3473. doi: 10.1128/JVI.71.5.3466-3473.1997 PubMed PMID: 9094618; PubMed Central PMCID: PMC191493.
- Tilgner M, Deas TS, Shi PY. The flavivirus-conserved penta-nucleotide in the 3’ stem-loop of the West Nile virus genome requires a specific sequence and structure for RNA synthesis, but not for viral translation. Virology. 2005;331(2):375–386. doi: 10.1016/j.virol.2004.07.022 PubMed PMID: 15629780.
- Villordo SM, Gamarnik AV. Genome cyclization as strategy for flavivirus RNA replication. Virus Res. 2009;139(2):230–239. doi: 10.1016/j.virusres.2008.07.016 Epub 20080909. PubMed PMID: 18703097; PubMed Central PMCID: PMC5440119.
- Ferrero DS, Ruiz-Arroyo VM, Soler N, Uson I, Guarne A, Verdaguer N. Supramolecular arrangement of the full-length Zika virus NS5. PLOS Pathogens. 2019;15(4):ARTN e1007656. doi: 10.1371/journal.ppat.1007656 PubMed PMID: WOS:000466742700014.
- Klema VJ, Ye MY, Hindupur A, Teramoto T, Gottipati K, Padmanabhan R, Rey FA, et al. Denguevirus Nonstructural Protein 5 (NS5) assembles into a dimer with a unique methyltransferase and polymerase interface. PLOS Pathogens. 2016;12(2):ARTN e1005451. doi: 10.1371/journal.ppat.1005451 PubMed PMID: WOS:000378152900044.
- Yang J, Jing X, Yi W, et al. Crystal structure of a tick-borne flavivirus RNA-dependent RNA polymerase suggests a host adaptation hotspot in RNA viruses. Nucleic Acids Res. 2021;49(3):1567–1580. doi: 10.1093/nar/gkaa1250 PubMed PMID: 33406260; PubMed Central PMCID: PMC7897508.
- Xu S, Ci Y, Wang L, et al. Zika virus NS3 is a canonical RNA helicase stimulated by NS5 RNA polymerase. Nucleic Acids Res. 2019;47(16):8693–8707. doi: 10.1093/nar/gkz650 PubMed PMID: 31361901; PubMed Central PMCID: PMC6895266.
- Herod MR, Ward JC, Tuplin A, et al. Positive strand RNA viruses differ in the constraints they place on the folding of their negative strand. RNA. 2022;28(10):1359–1376. doi: 10.1261/rna.079125.122 Epub 20220802. PubMed PMID: 35918125; PubMed Central PMCID: PMC9479745.
- Homan PJ, Favorov OV, Lavender CA, et al. Single-molecule correlated chemical probing of RNA. Proc Natl Acad Sci USA. 2014;111(38):13858–13863. doi: 10.1073/pnas.1407306111 Epub 20140909. PubMed PMID: 25205807; PubMed Central PMCID: PMC4183288.
- Choi KH. The role of the stem-loop a RNA promoter in flavivirus replication. Viruses. 2021;13(6):1107. doi: 10.3390/v13061107 Epub 20210609. PubMed PMID: 34207869; PubMed Central PMCID: PMC8226660.
- Friebe P, Bartenschlager R. Role of RNA structures in genome terminal sequences of the hepatitis C virus for replication and assembly. J Virol. 2009;83(22):11989–11995. doi: 10.1128/JVI.01508-09 Epub 20090909. PubMed PMID: 19740989; PubMed Central PMCID: PMC2772684.
- Schult P, Nattermann M, Lauber C, et al. Evidence for internal initiation of RNA synthesis by the Hepatitis C virus RNA-Dependent RNA Polymerase NS5B in Cellulo. J Virol. 2019;93(19):10–128. doi: 10.1128/JVI.00525-19 Epub 20190912. PubMed PMID: 31315989; PubMed Central PMCID: PMC6744235.
- Chahal J, Gebert LFR, Camargo C, MacRae IJ, Sagan SM. miR-122-based therapies select for three distinct resistance mechanisms based on alterations in RNA structure. Proc Natl Acad Sci, USA. 2021;118(33). doi: 10.1073/pnas.2103671118 PubMed PMID: 34385308; PubMed Central PMCID: PMC8379925.
- Kim YG, Yoo JS, Kim JH, Kim CM, Oh JW. Biochemical characterization of a recombinant Japanese encephalitis virus RNA-dependent RNA polymerase. BMC Mol Biol. 2007;8(1):59. doi: 10.1186/1471-2199-8-59 Epub 2007 Jul 12. PubMed PMID: 17623110; PubMed Central PMCID: PMC1934914.
- Calmels C, Metifiot M, Andreola ML. Template requirements of Zika RNA polymerase during in vitro RNA synthesis from the 3’-end of virus minus-strand RNA. Biochimie. 2022;195:71–76. doi: 10.1016/j.biochi.2021.11.003 Epub 20211113. PubMed PMID: 34780840.
- Dong H, Ray D, Ren S, et al. Distinct RNA elements confer specificity to flavivirus RNA cap methylation events. J Virol. 2007;81(9):4412–4421. doi: 10.1128/JVI.02455-06 Epub 20070214. PubMed PMID: 17301144; PubMed Central PMCID: PMC1900168.
- Zhao Y, Soh TS, Lim SP, Chung KY, Swaminathan K, Vasudevan SG, et al. Molecular basis for specific viral RNA recognition and 2’-O-ribose methylation by the dengue virus nonstructural protein 5 (NS5). Proc Natl Acad Sci U S A. 2015;112(48):14834–14839. doi: 10.1073/pnas.1514978112 Epub 20151117. PubMed PMID: 26578813; PubMed Central PMCID: PMC4672796.
- Nicholls CMR, Sevvana M, Kuhn RJ. Structure-guided paradigm shifts in flavivirus assembly and maturation mechanisms. Adv Virus Res. 2020;108:33–83. doi: 10.1016/bs.aivir.2020.08.003 Epub 20200923. PubMed PMID: 33837721; PubMed Central PMCID: PMC7510438.
- Barnard TR, Abram QH, Lin QF, et al. Molecular determinants of flavivirus virion assembly. Trends Biochem Sci. 2021;46(5):378–390. doi: 10.1016/j.tibs.2020.12.007 Epub 20210107. PubMed PMID: 33423940.
- Pong WL, Huang ZS, Teoh PG, et al. RNA binding property and RNA chaperone activity of dengue virus core protein and other viral RNA-interacting proteins. FEBS Lett. 2011;585(16):2575–2581. doi: 10.1016/j.febslet.2011.06.038 Epub 20110719. PubMed PMID: 21771593; PubMed Central PMCID: PMC7164067.
- Xie X, Zou J, Puttikhunt C, et al. Two distinct sets of NS2A molecules are responsible for dengue virus RNA synthesis and virion assembly. J Virol. 2015;89(2):1298–1313. doi: 10.1128/JVI.02882-14 Epub 20141112. PubMed PMID: 25392211; PubMed Central PMCID: PMC4300643.
- Byk LA, Gamarnik AV. Properties and functions of the dengue virus capsid protein. Annu Rev Virol. 2016;3(1):263–281. doi: 10.1146/annurev-virology-110615-042334 Epub 20160803. PubMed PMID: 27501261; PubMed Central PMCID: PMC5417333.
- Tan TY, Fibriansah G, Kostyuchenko VA, Ng TS, Lim XX, Zhang S, et al. Capsid protein structure in Zika virus reveals the flavivirus assembly process. Nat Commun. 2020;11(1):895. doi: 10.1038/s41467-020-14647-9 Epub 20200214. PubMed PMID: 32060358; PubMed Central PMCID: PMC7021721.
- Reid CR, Hobman TC. The nucleolar helicase DDX56 redistributes to West Nile virus assembly sites. Virology. 2017;500:169–177. doi: 10.1016/j.virol.2016.10.025 Epub 20161104. PubMed PMID: 27821284.
- Xu Z, Hobman TC. The helicase activity of DDX56 is required for its role in assembly of infectious West Nile virus particles. Virology. 2012;433(1):226–235. doi: 10.1016/j.virol.2012.08.011 Epub 20120825. PubMed PMID: 22925334; PubMed Central PMCID: PMC7119007.
- Overwijn D, Hondele M. DEAD-box ATPases as regulators of biomolecular condensates and membrane-less organelles. Trends Biochem Sci. 2023;48(3):244–258. doi: 10.1016/j.tibs.2022.10.001 Epub 20221104. PubMed PMID: 36344372.
- Krishnan MN, Ng A, Sukumaran B, et al. RNA interference screen for human genes associated with West Nile virus infection. Nature. 2008;455(7210):242–245. doi: 10.1038/nature07207 PubMed PMID: 18690214; PubMed Central PMCID: PMC3136529.
- Michalski D, Ontiveros JG, Russo J, et al. Zika virus noncoding sfRnas sequester multiple host-derived RNA-binding proteins and modulate mRNA decay and splicing during infection. J Biol Chem. 2019;294(44):16282–16296. doi: 10.1074/jbc.RA119.009129 Epub 20190913. PubMed PMID: 31519749; PubMed Central PMCID: PMC6827284.
- Paranjape SM, Harris E. Y box-binding protein-1 binds to the dengue virus 3’-untranslated region and mediates antiviral effects. J Biol Chem. 2007;282(42):30497–30508. doi: 10.1074/jbc.M705755200 Epub 20070828. PubMed PMID: 17726010.
- Bonenfant G, Williams N, Netzband R, et al. Zika virus subverts stress granules to promote and restrict viral gene expression. J Virol. 2019;93(12):10–128. doi: 10.1128/JVI.00520-19 Epub 20190529. PubMed PMID: 30944179; PubMed Central PMCID: PMC6613768.
- Diosa-Toro M, Kennedy DR, Chuo V, Popov VL, Pompon J, Garcia-Blanco MA, Coyne CB. Y-Box binding protein 1 interacts with dengue virus nucleocapsid and mediates viral assembly. MBio. 2022;13(1):e0019622. doi: 10.1128/mbio.00196-22 Epub 20220222. PubMed PMID: 35189699; PubMed Central PMCID: PMC8903895.
- Sanz MA, Almela EG, Garcia-Moreno M, et al. A viral RNA motif involved in signaling the initiation of translation on non-AUG codons. RNA. 2019;25(4):431–452. doi: 10.1261/rna.068858.118 PEpub 20190118. ubMed PMID: 30659060; PubMed Central PMCID: PMC6426287.
- Toribio R, Diaz-Lopez I, Boskovic J, et al. An RNA trapping mechanism in Alphavirus mRNA promotes ribosome stalling and translation initiation. Nucleic Acids Res. 2016;44(9):4368–4380. doi: 10.1093/nar/gkw172 Epub 20160316. PubMed PMID: 26984530; PubMed Central PMCID: PMC4872096.
- Kozak M. Downstream secondary structure facilitates recognition of initiator codons by eukaryotic ribosomes. Proc Natl Acad Sci U S A. 1990;87(21):8301–8305. doi: 10.1073/pnas.87.21.8301 PubMed PMID: 2236042; PubMed Central PMCID: PMC54943.
- Vogt DA, Andino R, Diamond MS. An RNA element at the 5′-End of the Poliovirus genome functions as a general promoter for RNA synthesis. PLOS Pathog. 2010;6(6):e1000936. doi: 10.1371/journal.ppat.1000936 Epub 20100603. PubMed PMID: 20532207; PubMed Central PMCID: PMC2880563.
- Tomescu AI, Robb NC, Hengrung N, et al. Single-molecule FRET reveals a corkscrew RNA structure for the polymerase-bound influenza virus promoter. Proc Natl Acad Sci USA. 2014;111(32):E3335–42. doi: 10.1073/pnas.1406056111 Epub 20140728. PubMed PMID: 25071209; PubMed Central PMCID: PMC4136606.
- Chen SC, Olsthoorn RC. Group-specific structural features of the 5’-proximal sequences of coronavirus genomic RNAs. Virology. 2010;401(1):29–41. doi: 10.1016/j.virol.2010.02.007 Epub 20100304. PubMed PMID: 20202661; PubMed Central PMCID: PMC7111916.
- Hurst KR, Koetzner CA, Masters PS. Identification of in vivo-interacting domains of the murine coronavirus nucleocapsid protein. J Virol. 2009;83(14):7221–7234. doi: 10.1128/JVI.00440-09 Epub 20090506. PubMed PMID: 19420077; PubMed Central PMCID: PMC2704785.
- Terasaki K, Narayanan K, Makino S, et al. Identification of a 1.4-kb-Long sequence located in the nsp12 and nsp13 coding regions of SARS-CoV-2 genomic RNA that mediates efficient viral RNA packaging. J Virol. 2023;97(7):e0065923. doi: 10.1128/jvi.00659-23 Epub 20230627. PubMed PMID: 37367225; PubMed Central PMCID: PMC10373556.
- Miyamoto S, Muramoto Y, Shindo K, et al. Contribution of RNA-RNA interactions mediated by the genome packaging signals for the selective genome packaging of influenza a virus. J Virol. 2022;96(6):e0164121. doi: 10.1128/JVI.01641-21 Epub 20220119. PubMed PMID: 35044211; PubMed Central PMCID: PMC8941900.
- Martinez-Salas E, Francisco-Velilla R, Fernandez-Chamorro J, Embarek AM. Insights into structural and mechanistic features of viral IRES elements. Front Microbiol. 2017;8:2629. doi: 10.3389/fmicb.2017.02629 Epub 20180104. PubMed PMID: 29354113; PubMed Central PMCID: PMC5759354.
- Dilweg IW, Gultyaev AP, Olsthoorn RC. Structural features of an Xrn1-resistant plant virus RNA. RNA Biol. 2019;16(6):838–845. doi: 10.1080/15476286.2019.1592070 Epub 20190405. PubMed PMID: 30951405; PubMed Central PMCID: PMC6546385.
- Villordo SM, Filomatori CV, Sanchez-Vargas I, et al. Dengue virus RNA structure specialization facilitates host adaptation. PLOS Pathog. 2015;11(1):e1004604. doi: 10.1371/journal.ppat.1004604 Epub 2015 Jan 31. PubMed PMID: 25635835; PubMed Central PMCID: PMC4311971.
- Long S. SARS-CoV-2 Subgenomic RNAs: Characterization, utility, and perspectives. Viruses. 2021;13(10):1923. doi: 10.3390/v13101923 Epub 20210924. PubMed PMID: 34696353; PubMed Central PMCID: PMC8539008.
- Boerneke MA, Ehrhardt JE, Weeks KM. Physical and functional analysis of viral RNA genomes by SHAPE. Annu Rev Virol. 2019;6(1):93–117. doi: 10.1146/annurev-virology-092917-043315 Epub 20190723. PubMed PMID: 31337286; PubMed Central PMCID: PMC6768749.
- Cao C, Cai Z, Xiao X, et al. The architecture of the SARS-CoV-2 RNA genome inside virion. Nat Commun. 2021;12(1):3917. doi: 10.1038/s41467-021-22785-x Epub 20210624. PubMed PMID: 34168138; PubMed Central PMCID: PMC8225788.