
ABSTRACT
The carob tree (Ceratonia siliqua L., Fabaceae) is a forest spontaneous and cultivated species, and grows in the Mediterranean countries and climates. It occupies a socioeconomic rank increasingly growing, both for the local population and national economies. Depending on tree characteristics and its components, authors often use geographical and/or local names especially in countries that market the carob pods. In Morocco, four intra-specific categories of carob tree were defined primarily in terms of reproductive traits and grafting, namely, “Lanta,” productive “Dkar,” unproductive “Dkar,” and “Sterile.” This work summarizes the state of knowledge on these categories and compares them with categories from other Mediterranean countries and climates, based on various descriptions and characteristics.
Introduction
The carob tree (Ceratonia siliqua L.) belongs to the family of Leguminosae (Fabaceae) (Fennane et al., Citation2007). It was described by Abi AL Khaïr Al Ichbili in the 12th century as a tree of Sulaiman or “Kharrouba” (El Khattabi, Citation1990). In Morocco, it is referred to with terms of Arab or Berber origin, namely, “slaghwa,” “tasliroua,” “sloiouig,” “tislîwha,” “kharrub,”and “tikidit” (Bellakhdar, Citation1997; INRA, Citation1965). In the region of Toudma (SW of Morocco), male and female trees are named by the respective words: “tikida” and “ikidw” (Gharnit, Citation2003).
Although its origin is controversial, it has been considered as originating from a xerotropical Indo-Malesian flora and placing the origin of its genus on the Arabian peninsula (Batlle and Tous, Citation1997). The species progeny currently present in the Western Mediterranean would have an origin like a germaplasm introduced by the Arabs in the Iberian Peninsula during the medieval domination (Cairati, Citation2013). The spontaneous forms of carob are particularly common in lower altitudes around the Mediterranean (Batlle and Tous, Citation1997; Rejeb et al., Citation1991); its cultivation in marginal and calcareous soils is very important from an environmental and economic point of view (Batlle and Tous, Citation1997).
The carob tree holds an important place for local populations; it is appropriated by the private sector, preserved by local communities, or exploited from the forest domain (Gharnit et al., Citation2001; Cairati, Citation2013). The carob tree and its components (leaf, flower, fruit, wood, bark, and root) are thus useful in human and animal food, ornamentation, in many industrial uses, carpentry, apiculture, and traditional medicines (Batlle and Tous, Citation1997; Cairati, Citation2013; Correia and Martins-Loução, Citation2005; Gharnit, Citation2003).
According to the literature and previous works, carob species is recommended for the reforestation of degraded areas threatened by desertification and disadvantaged rural areas (locust bean production, apiculture, etc.), ornamentation (aisles planting and urban green spaces), as windbreak (e.g., Gharnit et al., Citation2003, Vekiari et al., Citation2011).
The carob tree occupies an economic rank that is increasingly growing, representing a potential of development and diversification of agricultural and industrial income for both the local population and national economies (Cairati, Citation2013; Correia and Martins-Loução, Citation2005; Gharnit et al., Citation2001, Citation2003, Citation2006a). The main producers of the carob bean are Spain, Italy, Morocco, Portugal, Greece, Turkey, and Cyprus (Jorge et al., Citation2006; Matthaus and Ozcan, Citation2011).
Moreover, most of the carob cultivars are of unknown origin and represent a typical germaplasm of each country (Batlle and Tous, Citation1997). Depending on the characteristics of the tree and its components, geographical and/or local designations are often used, especially in countries that commercialize locust beans. Thus, more than 110 categories, cultivars, types, varieties, or clones of carob species have been reported in the literature in 13 countries; Italy, Spain, and Portugal have 26, 21, and 13 categories, respectively (Barracosa et al., Citation2007, Citation2008; Biner et al., Citation2007; Gharnit, Citation2003; Mahmoud Sidina et al., Citation2009; Naghmouchi et al., Citation2009; Tetik et al., Citation2011; Vekiari et al., Citation2011, Citation2012).
Intraspecific diversity of carob species from Morocco is partly genetic, and the provenance (accession or ecoregion) also has an effect in this regard. Generally, the morphological relationship between accessions of carob differs profoundly from their molecular one (Konaté et al., Citation2007; Mahmoud Sidina et al., Citation2009). In Lebanon, Talhouk et al. (Citation2005) reported a significant difference between and among carob populations, thereby constituting a valuable genetic material. Assessment of the genetic diversity of the carob species from Italy, Malta, and Spain demonstrated a high degree of mixing between the gene pools of the three countries. The low level of diversification of geographic areas reveals that the asexual propagation of selected clones played an important role in the spread of the species in the Western Mediterranean (La Malfa et al., Citation2014).
In Morocco, four intra-specific categories of the carob tree can be defined mainly in terms of reproductive traits and grafting, namely, “Lanta,” productive “Dkar,” unproductive “Dkar,” and “Sterile” (Gharnit et al., Citation2001). This work aims specifically to synthesize recent works on intra-specific categories of carob species from Morocco, in comparison with their counterparts in other geographical areas, based on morphometric and agro-economic descriptors.
Categorizing descriptors
Domestication
The carob tree is domesticated within orchards in the presence of other fruit trees or cereals. The grafting of “Sterile” and unproductive “Dkar” types of carob species is very common, favoring the “Lanta” category. Other domestication aspects, such as culture (plantations, application of manure and fertilizers, etc.), maintenance (coppicing, pruning, etc.) and/or exploitation (grazing, lopping, etc.) have been reported in this species (Gharnit et al., Citation2001, Citation2006a; Talhouk et al., Citation2005).
Grafting of male or female trees with low production of carob pod in domesticated areas disrupts the sex ratio. The partial grafting of rejections and/or branches of a single stem might suggest, wrongly, to cases of monoecy in this species. The “Lanta” category is most common in orchards, while the unproductive “Dkar” characterizes more marginal areas, hedgerows, marabouts, and cemeteries (Gharnit, Citation2003; Gharnit et al., Citation2004).
Carob trees are between 31 and 115 years old in the “Lanta” category and from 21 to 150 years old in the productive “Dkar” category. The maximum age was observed in productive “Dkar” type, more vigorous, followed by the “Lanta” type, more preserved but sometimes falling, and unproductive “Dkar” one, apparently because of intensive practice of grafting and/or lopping ().
Figure 1. Tree features in carob species categories. Corpus: Gharnit (Citation2003), unpublished data.
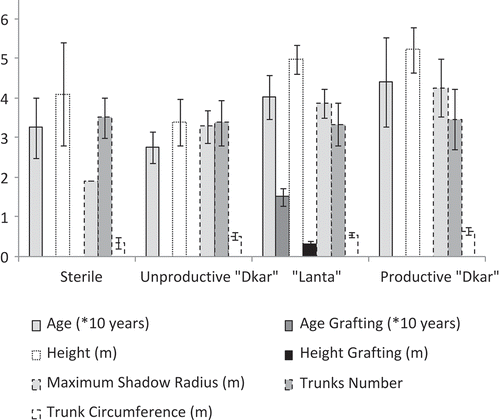
In general, the productive “Dkar” type grows in height and is less affected by falling in comparison with the “Lanta” one, given that this phenomenon is related to the grafting level. Trees of “Lanta” type are usually grafted at an early stage not exceeding 30 years. The height of grafting rarely exceeds 1 m in height with respect to ground level, depending on the quality of the rejection (crevices, secondary branches, subject to lopping, etc.) (). Grafting techniques practiced in carob trees are either escutcheon (63.8%) for small-circumference stems, and graft (27.7%) for older stems, generally with larger circumference, or both escutcheon and scion (8.50%) in multi-rejections trees with variable circumference (Gharnit, Citation2003).
The carob tree has a broad-based crown whose diameter is often greater than the tree height. The middle maximal ray of carob-tree shade varies from 1.90 to 4.26 m according to the species category (). The tree shade is especially symmetrical in pruned trees of productive “Dkar” (80.0%) and “Lanta” (69.6%) types (versus 53.3% and 50%, respectively, for unproductive “Dkar” and “Sterile” types), and in areas cleared of undergrowth. Asymmetrical port trees are often found in aerial congestion with other fruit trees in orchards or wild species in forest domain (Gharnit, Citation2003; unpublished data).
Carob trees have one main rejection or multiple rejections of variable circumference (). The multi-stem shrub forms in productive “Dkar” type are confined to certain very unfavorable habitats or due to preservation lack; while the average number of rejections observed in “Lanta” type is explained more by the recovery aptitude after trees falling (Gharnit, Citation2003).
Leaf
The relative development of the photosynthetic layer is more pronounced in trees of unproductive “Dkar” type when compared to productive “Dkar” and “Lanta” ones. Given the small number of trees in the “Sterile” type, it was not selected for biometric study of leaves. The unproductive “Dkar” category has longer and wider leaves, and longer petiole. Leaflets are also longer, wider, and rounder than those of productive “Dkar” and “Lanta” categories. The average width of leaves is similar in productive “Dkar” and “Lanta” types ().
Figure 2. Leaf and leaflets characteristics in carob species categories (Gharnit et al., Citation2005).
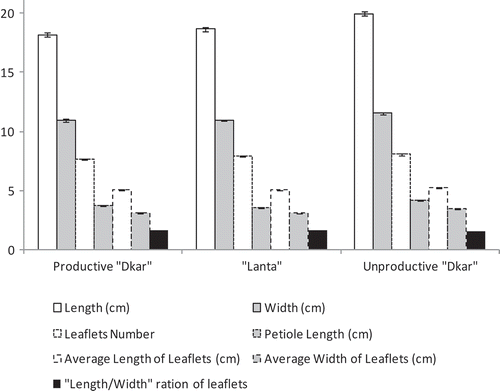
Leaflets in unproductive “Dakar” type are more numerous (leaves with 9–10 leaflets), reaching 40 leaflets per bipinnate or doubly composed leaf, depending on the degree of leaf division and branching of primary rachis (Gharnit, Citation2003; Gharnit et al., Citation2005). Basal leaflets enable the most complete discrimination between these types of carob species and reach their maximum in unproductive “Dkar” type: 5.08 cm of length and 3.46 cm of width. Basal leaflets are shorter and apical leaflets are narrowest in the three types of carob species (Gharnit et al., Citation2005).
In “Lanta” type, leaf development is optimal for all parameters studied in age classes from 10 to 20 years. The age classes “20–40 years,” “20–40 years and 80–160 years,’ and “20–80 years” also promote, respectively, the length, leaflets number, and petiole length. Given the characters average of leaflets, the age range of classes from 40 to 80 years and/or those of 80 to 160 years old have generally a slower development in leaflet length. The last tree class also disadvantages the leaflet width. As for the ratio of length to width of the leaflet, the average minimum is observed in the age class from 40 to 80 years, coinciding with the end of the optimum development of the leaflet and giving it a more rounded shape (Gharnit et al., Citation2005).
Flowers
Except for the “Sterile” type, very rare and not blooming, the flower is present in the other three types of the carob species from Morocco: “Lanta” (usually female flowers), productive “Dkar” (usually female flowers and sometimes hermaphrodites ones), and unproductive “Dkar” (flowers usually male, sometimes female or hermaphrodite) (Gharnit et al., Citation2001; unpublished data).
Inflorescences are simple, singular, or grouped in whorls, or sometimes branched. The leaflets can sometimes alternate the flowers at the base of the inflorescence axis. The indefinite inflorescences characterize more female trees (91.7% versus 88.0%); while the defined inflorescences characterize more male trees (12.0% versus 8.33%). However, the rate of defined inflorescences is approximately 10% (Gharnit et al., Citation2004), as the inflorescences of Leguminosae are mostly indefinite growth (Tucker, Citation1992).
The average length of the inflorescence is 4 to 6 cm, having from 32 to 40 flowers per inflorescence, with a flower density from 6 to 8 flowers/cm. The inflorescence length is higher in male trees when compared with female ones (5.71 cm versus 4.45 cm). Similarly, the flowers number per inflorescence is greater in male trees than in female ones (42 flowers versus 30 flowers in the same order) (Gharnit et al., Citation2004).
According to the flower sex, four inflorescence types are recognized: male, female, polygamous, and hermaphrodite. The floral mosaic in carob species marks an instability generating several types of male flowers, differing by the sepal’s number, the stamen’s number, and the length of stamen filament, and female flowers, which differ by the pistil shape (Gharnit et al., Citation2004). In addition, seven types of male flowers can be distinguished. The flower type with [5 sepals (S) + 5 stamens (St)] is the most represented with a percentage of 71.6%; it is followed primarily by types with [5 S + 6 St) and (6 S + 5-6 St] with percentages of 11.3% and 15.8% in the same order. In male flowers, the pistil is rudimentary and considered non-functional. Further, the female flowers are of three types, with 5 or 6 sepals and differing by the pistil shape: 35% with curved pistil, 34.8% with slightly curved pistil, and 30.2% with straight pistil. The presence of a non-functional stamen is one of the particularities of the female flowers. The hermaphrodite flowers are of the following floral formulas: [5 S + 5 St + 2 Carpels (C)] or [5 S+ 6 St + 2 C]. In this case, the stamens (anthers and filaments) persist after pollination until the mature stage of fruits (Gharnit et al., Citation2004).
In the terminal flower of male inflorescences, sometimes with nectariferous spatulate-disc, the sepals and stamens numbers vary respectively from 5 to 8 St and from 5 to 14 St, of which two at least are sometimes welded. The terminal flower of female inflorescences, sometimes spatulate, present 4, 5, 8, or 10 sepals and 1, 2, or 4 pistils partially welded or free (Gharnit et al., Citation2004).
Carob bean
The production of locust bean characterizes two categories of carob species: “Lanta” and productive “Dkar” (Gharnit et al., Citation2001). The “Lanta” type has an alternate and moderate fruiting. The production regularity is more important and more marked in productive “Dkar” type. The percentage of aborted flowers is high for both productive “Dkar” (83%) and “Lanta” (81.8%) categories. The percentage of aborted fruit remains low: 6.58% and 7.03%, respectively, for “Lanta” and productive “Dkar” types. The developed fruit rate varies from 9.97% for the productive “Dkar” category to 11.5% for the “Lanta” one ().
Figure 3. Pod production in carob categories from Morocco (Gharnit et al., Citation2010).
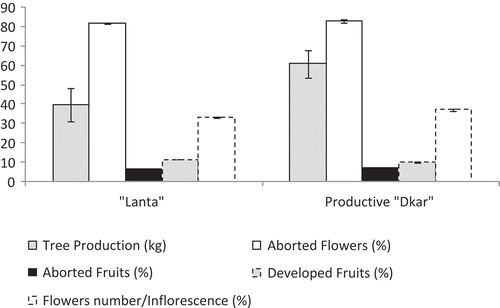
The carob pods of productive “Dkar” type present the best kernel yield (20.5%) and are straighter than those of “Lanta” type; its curvature degree is inversely proportional to the tree age of “Lanta” type. Those older (≥50 years) have longer (14.2 cm), wider (2.01 cm), heavier (11.5 g), and more fleshy (9.35 g) locust beans, with more numerous seeds (10.1). The sufficiently old trees show a more significant seed yield of 19.3%. In productive “Dkar” type, more pods are long; more of them are thin and have high yield kernels. This is roughly the same for the pod of “Lanta” type. The age effect could not be demonstrated in trees of productive “Dkar” type because the strong domestication of the carob-tree rarefies this type (low numbers) on one hand and the most identified specimens are significantly aged on the other hand (Gharnit et al., Citation2006b).
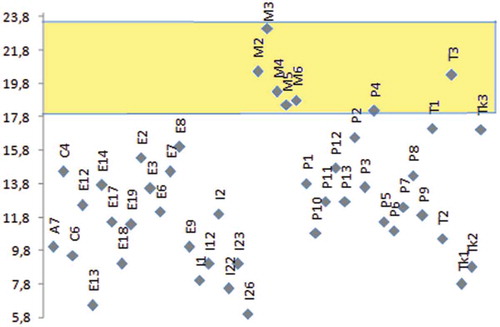
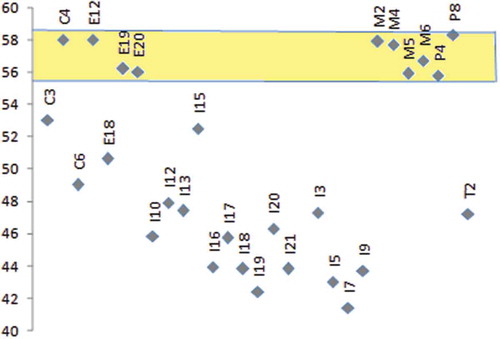
In comparison with other Mediterranean countries and climates, it turns out that the carob bean from Morocco ranks among the categories with very high kernels yield (“Canela” from Portugal and “Wild population” from Tunisia; ). The high seed yield in the “Domesticated” type, denomination reported by Mahmoud Sidina et al. (Citation2009), (M3, ) could correspond, among others, to spontaneous types, not grafted and/or preserved after clearing of primary forests (productive “Dkar” category in our case).
Figure 4. Kernels yield (%) in carob categories from Morocco (M2, productive “Dkar”; M3, “Domesticated type” (Naghmouchi et al., Citation2009); M4, 25–50-year-old “Lanta”; M5, ≥50-year-old “Lanta”; M6, ≤25-year-old “Lanta”); Spain (Ex), Portugal (Px), Tunisia (Tx), Italy (Ix), Cyprus (Cx), Turkey (Tkx), and USA (Ax). Corpus: Gharnit (Citation2003), Barracosa et al. (Citation2007, Citation2008), Biner et al. (Citation2007), Mahmoud Sidina et al. (Citation2009), Naghmouchi et al. (Citation2009), and Tetik et al. (Citation2011).
Locust bean from Morocco is among the shortest (“Tantillo,” “Melara,” “Femminella,” “Racemosa,” and “Orleans 1” from Italy; “Bolser” and “Clifford” from USA), moderately large (“Rojal” and “De le Mel” from Spain; “Spargale,” “Alfarroba Burro,” “Galhosa,” and “Gasparinha” from Portugal; “Sfax” from Tunisia; “Impollinatrice” from Italy), thinnest (“Nera di Cefalù” from Italy; “Wild Type” from Turkey), more lightweight pods (“Canela” from Portugal; “Wild population” from Tunisia), those with low seeds number (“Mulata” from Portugal), and low rate of aborted seeds (“Wild population” from Tunisia) (Barracosa et al., Citation2007, Citation2008; Biner et al., Citation2007; Gharnit, Citation2003; Mahmoud Sidina et al., Citation2009; Naghmouchi et al., Citation2009; Tetik et al., Citation2011).
Carob kernel
In productive “Dkar” type, endosperm yield of seeds is very highly correlated with the thickness (4.2 mm) and negatively with the weight (0.17–0.21 g) of the kernel. Therefore, the thick and lightweight seeds of this type of carob-tree have a high gum yield. In seeds of “Lanta” type, the endosperm percentage is highly negatively correlated with the length (9.3–9.48 mm) and highly negatively correlated with the weight (0.23–0.42 g) of the kernel. Then, shorter and more lightweight seeds of this carob type generate a relatively high gum yield. For other seed components, the highest values were observed in older “Lanta” type: 23.0% for the seed coat and 21.0% for the embryo-cotyledons (Gharnit, Citation2003; Gharnit et al., Citation2006b).
The productive “Dkar” category inserted among those with more elevated endosperm yield (“Rojal” from Spain and “Koundourka” from Cyprus; ) and with shorter (“Lagoinha” and “Costela Canela” from Portugal), moderately wide (“Alfarroba Burro” and “Galhosa” from Portugal), fairly thick (“Costa Loulé” and “Lagoinha” from Portugal), and lightweight seeds (“Gasparinha” and “Costela Canela” from Portugal). The “Lanta” type is among categories presenting a high endosperm yield (“Rojal” and “Sayalonga” from Spain, “Canela” from Portugal, and “Koundourka” from Cyprus; ) and having fairly long (“Spargale,” “Galhosa,” “Lagoinha,” and “Costela Canela” from Portugal), larger (“Negra” from Spain), thicker (“Alfarroba Burro” and “Mulata” from Portugal), and heavier kernels (“Galhosa” from Portugal) (Barracosa et al., Citation2007, Citation2008; Biner et al., Citation2007; Gharnit, Citation2003; Naghmouchi et al., Citation2009; Tetik et al., Citation2011).
Figure 5. Endosperm yield (%) in carob categories from Morocco (M2, productive “Dkar”; M4, 25–50-years-old “Lanta”; M5, ≥50-years-old “Lanta”; M6, ≤25-years-old “Lanta”), Spain (Ex), Portugal (Px), Tunisia (Tx), Italy (Ix) and Cyprus (Cx). Corpus: Gharnit (Citation2003), Barracosa et al. (Citation2007, Citation2008).
Chemical and biochemical aspects
The carob tree is an important resource of minerals, especially calcium, iron, and potassium, with correspondences between the mineral composition (quantitative variables) on the one hand and plant parts (leaves, bark, pulp, and seed coat), the collection year, and intra-specific category on the other hand (El Hajaji et al., Citation2013).
The carob-tree bark contains high quantities of polyphenolic compounds; the unproductive “Dkar” type has generally a higher concentration of polyphenols and a higher antioxidant activity in comparison with the productive “Dkar” and “Lanta” types (El Hajaji et al., Citation2011). Leaves of the three carob categories studied exhibit potent antioxidant properties and contain important quantities of phenolic compounds; the “Lanta” type generally has an elevated concentration of polyphenols when compared with unproductive and productive “Dkar” types (El Hajaji et al., Citation2010).
Otherwise, in Southern Greece, pods of the fleshy variety of carob species exceed those of the wild variety in concentrations of proteins, sugars, and fatty acids, while the wild variety passes the fleshy one in concentration of polyphenols (Vekiari et al., Citation2012). Comparing the fleshy variety from Turkey with the fleshy and wild ones from Greece, the Turkish locust bean appears to have a higher level of total sugars, fructose, and sucrose in the first stage of pod growth. While a higher ratio of fatty total-acids “unsaturated/saturated” was found in the fleshy mature pods from Greece (Vekiari et al., Citation2011).
In Turkey, locust beans of the cultivated variety have a high concentration in total sugar than those of the wild type. However, this difference is due to the highest concentration of sucrose in the cultivated variety, which does not differ from the wild type in terms of concentrations of fructose or glucose (Biner et al., Citation2007).
The study of the oil content, fatty acids, tocopherols, and sterols in kernels of cultivated and wild carob tree from Turkey underlines generally an exceeding in wild carob tree for oil yield, the main fatty acids (except among others oleic acid), major tocopherols (except among others β-tocopherol), and sterols (Matthaus and Ozcan, Citation2011). The highest average percentages of the total ash content and total nitrogen (%) were observed in carob seeds of the grafted type, compared with the wild one (Oziycia et al., Citation2014). Besides, the average soluble solids content of the carob pulp is more marked in the grafted genotype compared to the wild one (Tetik et al., Citation2011).
Abiotic factors
The carob tree is a xerophyte that is not very demanding as to soil type (except for hydromorphic or clayey-impermeable soils) while also showing some other particular features, such the rusticity, the resistance to drought, soil fertilization via fixing atmospheric nitrogen, soil conservation against erosion, etc. (Gharnit, Citation2003).
By focusing on one ecoregion or accession, as defined by Konaté et al. (Citation2007) and Mahmoud Sidina et al. (Citation2009) and with an homogenous macroclimate, we have highlighted the effect of the location (especially exposure and soil type) on morphometric characters of the carob tree (Gharnit, Citation2003; Gharnit et al., Citation2005, Citation2006b).
Trees from preserved locations, subject to fertilization, and growing on shady exposure and wetter or highly developed soils show maxima of average heights and have the most developed leaves. Leaflet length and width show also their maxima, and leaflets are more elongated. Basal leaflets are shortest and apical ones are narrowest. The leaf rachis presents an appreciable elongation (Gharnit et al., Citation2005).
However, the location circumstances do not explain very significant differences of floral quantitative parameters (inflorescence length, flowers density, etc.). The pollen diameter shows a marked variability even at the intra-individual level (Gharnit et al., Citation2004). The study of fruiting phenology of productive “Dkar” and “Lanta” types revealed that the site conditions have no significant effect on rates of aborted flowers and fruits or on the percentage of developed fruits (Gharnit et al., Citation2010).
Preservation and/or edaphic conditions promote pod width and weight, pulp weight, and seed weight. The maximum values of the chord, corresponding to straighter locust beans, are observed in locations moderately domesticated (preservation and manure application) and in the Northern exposures. However, these factors apparently have no effect on other studied parameters of carob bean, namely, length, thickness, number of seeds, seed yield, and aborted seeds rate. The highly developed soil would advantageous for seed thickness and weight. However, endosperm yield of seeds does not seem to be related to site conditions (Gharnit et al., Citation2006b).
Regeneration
The carob tree can be propagated by seeds, cuttings, layering, and grafting; the most efficient technique in the natural environment is the multiplication by seedling. Some farmers assert that carob seeds rejected by cows and jackals or impregnated with cow dung regenerate easily and can vegetate efficiently; its growth depends on the intra-specific category (Gharnit, Citation2003; unpublished data).
The high rate of seed germination was obtained in the productive “Dkar” type, in comparison with the “Lanta” one. Although seedlings transplanting is easily successful in the field, seedlings of productive “Dkar” grow more in height, reaching 2.10 m after 4 years, while having a more important root-system since the juvenile stage. Macro-cuttings tests in vivo of both carob-tree types show difficulty rooting, despite the high rate of bud burst, especially in the productive “Dkar” type (Gharnit et al., Citation2008).
In other countries, such as Portugal, Spain, and Egypt, authors obtained an elevated rooting rate using cuttings in carob cultivars, including De la Mel, Galhosa, and Mulata. But transplanting in the field of rooted cuttings has been successful in Spain (in Batlle and Tous, Citation1997). In California, the experience of several nurserymen showed the difficulty to propagate this species by cuttings (Coit, Citation1949).
The micropropagation of the carob tree, from nodes of plantlets derived of germination has been successful until the acclimatization (Belaizi et al., Citation1994). The micropropagation of the carob tree from Morocco is possible from the juvenile material (Gharnit, Citation1997; Gharnit and Ennabili, Citation2009; Saidi et al., Citation2007).
Nevertheless, the plantlets on which is practiced the vitro-culture derived from carob-kernels germination can generate various intra-specific categories. Moreover, the in vitro establishment of several carob cultivars from Italy, Portugal, Spain, and Morocco, from buds collected on mature plants, is possible, but adjustments are needed depending on the variety, explant type and season (Brugaletta et al., Citation2009). In this respect, Hsina and El Mtili (Citation2009) developed a micrografting technique using in vitro germinated seedlings as rootstock and shoots, from a mature genotype of productive “Dkar” type, as grafts.
Concluding remarks
The carob tree from Morocco is dioecious, rarely polygamous or trimonoecious, but not hermaphrodite. The intensive practice of grafting disrupts the sex ratio in this species, and is the origin of some cases of “artificial” monoecy.
Depending on reproductive traits and grafting, four intra-specific categories of the carob tree can be defined, namely: (i) “Lanta” or grafted type, female, producing locust bean; (ii) productive “Dkar” or spontaneous type, generally female, producing locust bean; (iii) unproductive “Dkar” or spontaneous type, usually male, sometimes polygamous, but not producing locust bean; and (iv) “Sterile” type, very rare, not flowering.
According to the circumstances, one or the other type of carob tree can be recommended in plantations of the farming and/or urban zones. For example, the “Lanta” type is widely known by its high carob pulp yield. The productive “Dkar” type could be recommended for large-scale reforestation in disadvantaged rural areas both for its socio-economic interest (regular production, high seed and endosperm yields, etc.) and its ecological and environmental importance (adaptation to poor soils, drought tolerance, fight against erosion, etc.). Since the carob bean is sought in particular for its industrial use, morphometric characters, such as length, width, and thickness, can help to select seeds with high agro-industrial potential.
In urban areas, the planting of the productive “Dkar” type could also be recommended due to its very dense leaf layer, slow growth, and not requiring frequent fertilization, and can generate supplementary returns. However, falls of mature pods and aborted flowers can cause a fairly frequent cleaning of streets and gardens. The unproductive “Dkar” type is not recommended in this environment because of its strong flower odor and cases of pollen allergy. The “Sterile” type would be advisable for ornamentation.
On the other hand, it would be interesting to finalize a program of Research/Development involving the intra-specific characterization of the carob-tree on a national scale. In this regard, it would be very useful to give importance to its typology, and not be limited to non-accurate appellations (spontaneous or wild/domesticated or cultivated). An orchard (domesticated area) could include both spontaneous carob-trees, preserved after clearing of primary forests, grafted ones, and others derived of transplantation, etc. A pre-forest or forest zone can also include orchards abandoned, formerly domesticated.
Seeing the genetic erosion mainly due to grafting, the conservation of this species requires an evaluation in domesticated areas (socio-agro-economic variables) and in pre-forest and forest domains (natural variables). A collection reconstitution from the recognized carob-tree categories would contribute to preservation of the current species diversity and serve as a gene bank to develop the carob-tree propagation.
Furthermore, the efficient way of multiplying the carob species from Morocco would be from the seedlings systematically grafted after 1 year according to the required category, because the recognition of these types of carob tree when young remains impossible, and the farmer opts for categories with reliable and rapid production. Research on regeneration in vivo and/or in vitro of this species should target an effective and reproducible procedure according the intra-specific category, from mature or rejuvenated vegetative organs with known agro-economic characteristics.
The participatory approach, involving the local population, local and Sectional Administrations and Institutions, and Research Centers, is sine qua non to develop this Research/Development program: land management and conservation of plant diversity and local know-how, extension and development of carob-tree cultivation especially in disadvantaged areas, creating processing cooperatives and valorization of locust bean.
Literature cited
- Barracosa, P., J. Osório, and A. Cravador. 2007. Evaluation of fruit and seed diversity and characterization of carob (Ceratonia siliqua L.) cultivars in Algarve region. Sci. Hort. 114:250–257.
- Barracosa, P., M.B. Lima, and A. Cravador. 2008. Analysis of genetic diversity in Portuguese Ceratonia siliqua L. cultivars using RAPD and AFLP markers. Sci. Hort. 118:189–199.
- Batlle, I., and J. Tous. 1997. Carob tree. Ceratonia siliqua L. promoting the conservation and use of underutilized and neglected crops. Institute of Plant Genetics and Crop plant Research, Gatersleben/International Plant Genetic Ressources Institute, Rome, Italy.
- Belaizi, M., M.R. Bolen, and P. Boxus. 1994. Régénération in vitro et acclimatation du caroubier (Ceratonia siliqua L.), p. 227–232. In: AUPELF-UREF (ed.). Quel avenir pour l’amélioration des plantes? John Libbey Eurotext, Paris, France.
- Bellakhdar, J. 1997. La pharmacopée marocaine traditionnelle. Médecine arabe ancienne et savoirs populaires. IBIS Press, Paris, France.
- Biner, B., H. Gubbuk, M. Karhan, M. Aksu, and M. Pekmezci. 2007. Sugar profiles of the pods of cultivated and wild types of carob bean (Ceratonia siliqua L.) in Turkey. Food Chem. 100:1453–1455.
- Brugaletta, M., S. La Malfa, A. Gentile, R. Almeida, and A. Romano. 2009. In vitro culture establishment of Ceratonia siliqua (L.) mature trees from cultivars of different Mediterranean countries. Acta Hort. 812:113–120.
- Cairati, E. 2013. Historia cultural del algarrobo, desde la cuenca del Mediterráneo hasta la Costa Norte de Perú. Other Modernities. 10:186–204.
- Coit, J.E. 1949. Carob culture in the semi-arid southwest. Walter Rittenhouse, San Diego, CA.
- Correia, P.J., and M.A. Martins-Loução. 2005. The use of macronutrients and water in marginal Mediterranean areas: The case of carob-tree. Field Crops Res. 91:1–6.
- El Hajaji, H., A. Farah, A. Ennabili, D. Bousta, H. Greche, B. El Bali, and M. Lachkar. 2013. Etude comparative de la composition minérale des constituants de trois catégories de Ceratonia siliqua L. J. Mater. Environ. Sci. 4(2):165–170.
- El Hajaji, H., N. Lachkar, K. Alaoui, Y. Cherrah, A. Farah, A. Ennabili, B. El Bali, and M. Lachkar. 2010. Antioxidant properties and total phenolic content of three varieties of carob tree leaves from Morocco. Rec. Nat. Prod. 4(4):193–204.
- El Hajaji, H., N. Lachkar, K. Alaoui, Y. Cherrah, A. Farah, A. Ennabili, B. El Bali, and M. Lachkar. 2011. Antioxidant activity, phytochemical screening and total phenolic content of extracts from three genders of carob tree barks growing in Morocco. Arab. J. Chem. 4:321–324.
- El Khattabi, M.A. 1990. Umdat Attabib fi maârifat annabat d’Abi AL Khaïr Al Ichbili [Doctor’s support for plants recognition]. Series “Attourat,” Academy Editions of the Kingdom of Morocco.
- Fennane, M., M. Ibn Tattou, A. Ouyahya, and J. El Oualidi. 2007. Flore pratique du Maroc. Vol. 2. Série Botanique 38, Travaux de l’Institut Scientifique, Rabat, Morocco.
- Gharnit, N. 1997. Le caroubier (Ceratonia siliqua L.), Essais de propagation in vitro et intérêt socio-économique au cercle de Mokrisset (NW Maroc). Mémoire D.E.S.A., no. 576.5 GHA. Faculté des Sciences, Tétouan, Morocco.
- Gharnit, N. 2003. Caractérisation du caroubier (Ceratonia siliqua L.) originaire de la Province de Chefchaouen (Nord-ouest du Maroc). Faculté des Sciences et Techniques, Tanger, Morocco, Thèse de Doctorat.
- Gharnit, N. and A. Ennabili. 2009. Essais préliminaires de culture in vitro du caroubier (Ceratonia siliqua L.) originaire du Nord ouest du Maroc. Biomatec Echo 3(6):18–25.
- Gharnit, N., N. El Mtili, A. Ennabili, and F. Sayah. 2004. Floral characterization of carob tree (Ceratonia siliqua L.) from the Province of Chefchaouen (NW of Morocco). Moroccan J. Biol. 1:41–51.
- Gharnit, N., N. El Mtili, A. Ennabili, and F. Sayah. 2005. Caractérisation foliaire du caroubier (Ceratonia siliqua L.) originaire de la province de Chefchaouen (Nord-ouest du Maroc). J. Bot. Soc. Bot. France, 31:75–84.
- Gharnit, N., N. El Mtili, A. Ennabili, and F. Sayah. 2006a. Importance socio-économique du caroubier (Ceratonia siliqua L.) dans la Province de Chefchaouen (Nord-ouest du Maroc). J. Bot. Soc. Bot. France 33:43–48.
- Gharnit, N., N. El Mtili, A. Ennabili, and F. Sayah. 2006b. Pomological characterization of carob tree (Ceratonia siliqua L.) from the Province of Chefchaouen (NW of Morocco). Moroccan J. Biol. 2–3:1–11.
- Gharnit, N., N. El Mtili, A. Ennabili, and F. Sayah. 2008. Essais de culture in vivo du caroubier (Ceratonia siliqua L.) originaire du Nord-ouest du Maroc. Rev. AFN Maroc. 2–3:107–115.
- Gharnit, N., N. El Mtili, A. Ennabili, and F. Sayah. 2010. Aspects de la phénologie fructifère du caroubier (Ceratonia siliqua L.) originaire de la Province de Chefchaouen (nord-ouest du Maroc). J. Bot. Soc. Bot. France 49:9–12.
- Gharnit, N., N. El Mtili, A. Toubi Ennabili, and A. Ennabili. 2001. Social characterisation and exploitation of carob tree (Ceratonia siliqua L.) from Mokrisset and Bab Taza (NW of Morocco). Sci. Lett. 3(2):1–10.
- Gharnit, N., N. El Mtili, A. Toubi Ennabili, and A. Ennabili. 2003. Exploitation du caroubier (Ceratonia siliqua L.) dans la Commune d’Aïn Beïda, Province de Chefchaouen, p. 93–100. In: A. Boukroute (ed.). Arbres et espaces verts urbains: Du chercheur au gestionnaire. Actes Editions, Rabat, Morocco.
- Hsina, T., and N. El Mtili. 2009. In vitro micrografting of mature carob tree (Ceratonia siliqua L.). Open Hort. J. 2:44–48.
- INRA (Institut National de la Recherche Agronomique). 1965. Les cultures fourragères irriguées au Maroc. Collection Technique et Productions Agricoles, Rabat, Morocco.
- Canhoto, J.M., S.C. Rama, and G.S. Cruz. 2006. Somatic embryogenesis and plant regeneration in carob (Ceratonia siliqua L.). In Vitro Cell. Dev. Biol.—Plant 42:514–519.
- Konaté, I., A. Filali-Maltouf, and E. Berraho. 2007. Diversity analysis of Moroccan carob (Ceratonia siliqua L.) accessions using phenotypic traits and RAPD markers. Acta Bot. Malac. 32:79–90.
- La Malfa, S., S. Currò, A.B. Douglas, M. Brugaletta, M. Caruso, and A. Gentile. 2014. Genetic diversity revealed by EST-SSR markers in carob tree (Ceratonia siliqua L.). Biochem. Sys. Ecol. 55:205–211.
- Mahmoud Sidina, M., M. El Hansali, N. Wahid, A. Ouatmane, A. Boulli, and A. Haddioui. 2009. Fruit and seed diversity of domesticated carob (Ceratonia siliqua L.) in Morocco. Sci. Hort. 123:110–116.
- Matthaus, B., and M.M. Ozcan. 2011. Lipid evaluation of cultivated and wild carob (Ceratonia siliqua L.) seed oil growing in Turkey. Sci. Hort. 130:181–184.
- Naghmouchi, S., M.L. Khouja, A. Romero, J. Tous, and M. Boussaid. 2009. Tunisian carob (Ceratonia siliqua L.) populations: Morphological variability of pods and kernel. Sci. Hort. 121:125–130.
- Oziycia, H.R., N. Tetika, I. Turhana, E. Yatmaza, K. Ucgunb, H. Akgulb, H. Gubbukc, and M. Karhana. 2014. Mineral composition of pods and seeds of wild and grafted carob (Ceratonia siliqua L.) fruits. Sci. Hort. 167:149–152.
- Rejeb, M.N., D. Laffray, and P. Louguet. 1991. Physiologie du caroubier (Ceratonia siliqua L.) en Tunisie, p. 417–426. In: Physiologie des arbres et arbustes en zones arides et semi-arides. Groupe d’Etude de l’Arbre, Paris.
- Saidi, R., A. Lamarti, and A. Badoc. 2007. Micropropagation du caroubier (Ceratonia siliqua) par culture de bourgeons axillaires issus de jeunes plantules. Bull. Soc. Pharm. Bordeaux 146:113–129.
- Talhouk, S.N., P. Van Breugel, R. Zurayk, A. Al-Khatib, J. Estephan, A. Ghalayini, N. Debian, and D. Lychaa. 2005. Status and prospects for the conservation of remnant semi-natural carob Ceratonia siliqua L. populations in Lebanon. For. Ecol. Mgt. 206:49–59.
- Tetik, N., I. Turhan, H.R. Oziyci, H. Gubbuk, M. Karhan, and S. Ercisli. 2011. Physical and chemical characterization of Ceratonia siliqua L. germplasm in Turkey. Sci. Hort. 129:583–589.
- Tucker, S.C. 1992. The role of floral development in studies of legume evolution. Can. J. Bot. 70:692–700.
- Vekiari, A.S., G. Ouzounidou, G. Gork, M. Ozturk, and M. Asfi. 2012. Compositional changes of major chemical compounds in Greek carob pods during development. Bull. Chem. Soc. Ethiop. 26(3):343–351.
- Vekiari, S.A., G. Ouzounidou, M. Ozturk, and G. Görk. 2011. Variation of quality characteristics in Greek and Turkish carob pods during fruit development. Proc. Soc. Behav. Sci. 19:750–755.