Abstract
In Gabon, two major species of Loranthaceae, Phragmanthera capitata (Sprengel) Balle and Globimetula braunii (Engl.) Danser to a lesser extent, parasitize Hevea brasiliensis (Müll.) Arg. The behavior of different clones of rubber trees facing the Loranthaceae parasitism has been studied in the large industrial plantation of Mitzic and in three neighboring smallholding plantations. The clones PR 261, RRIM 600 and PB 260, although never totally resistant, are far less susceptible to Loranthaceae than the clones AVROS 2037, PB 217, PB 235 and GT 1. The parasite development kinetics was more rapid on the most susceptible clones. Phellem thickness, cortical parenchyma thickness and tannin cell density were independent of susceptibility. The number of tree ramifications was significantly different between the clones. Interestingly, on the least susceptible trees, being also the most ramified, the parasite showed the slowest development.
Introduction
In Gabon, the cultivation of rubber tree, Hevea brasiliensis (Müll.) Arg., was first introduced during World War II to supply rubber for the Allied nations (Serier Citation1985), and then given up soon after the war. It was reintroduced in the 1980s to diversify the economy of the country, which was mainly depending on its oil reserves (Chambrier Citation1990).
The rubber tree, grown for its latex, has been introduced in various regions of the world where it is a target for specific parasitic plants belonging to the Loranthaceae family, e.g. Taxillus chinensis and Dendrophthoe pentandra in China (Zhiwei et al. Citation1995), and Phragmanthera capitata in most African plantations, namely in Cameroon (Sallé Citation1994; HEVECAM Citation1995), Nigeria (Gill and Onybé Citation1990; Akinlabi and Aigbodion Citation2005) and Gabon (Guyot and Ntawanga Citation1998; Engone Obiang Citation2006; Engone Obiang and Sallé Citation2006).
The Loranthaceae, thoroughly studied by Balle and Hallé (Citation1961) and Polhill and Wiens (Citation1998), consist of small chlorophyllous epiphytic shrubs living as hemiparasites on branches of wild or cultivated trees. These hemiparasites look like clumps anchored in the wood of the host through a specialized organ, the haustorium, which establishes functional connections with the host vascular tissues. Through the haustorium, the parasite pumps in water, mineral salts and the organic matter required for its development from the host (Kuijt Citation1969; Sallé et al. Citation1998).
The infestation of rubber trees by Loranthaceae is one of the main phytosanitary problems encountered in Gabon, especially within the country's main plantation located in Mitzic where Phragmanthera capitata is by far the most common parasite (Guyot and Ntawanga Citation1998; Engone Obiang Citation2006; Engone Obiang and Sallé Citation2006).
Previous research carried out on Viscaceae showed a susceptibility gradient to parasitic epiphytes on hardwood trees. For example, studies concerning the associations of poplar/Viscum album (Frochot et al. Citation1978) and oak/Viscum album (Grazi and Urech Citation1983) led to the identification of some resistant cultivars to the European mistletoe. Parameters involved in this resistance are now well documented, i.e., anatomical structures and phenolic compounds (Armillotta Citation1984; Sallé et al. Citation1984; Hariri Citation1989; Hariri et al. Citation1991, Citation1992; Sallé et al. Citation1993). Nevertheless, these studies did not take into account the effect of the morphology of the host's tree crown.
Phragmanthera capitata and Globimetula braunii are the major species of Loranthaceae among the nine species identified on rubber trees in Mitzic's zone (Engone Obiang Citation2006). Phragmanthera capitata is the most invasive as it parasitizes 95% of the rubber trees, whereas G. braunii attacks only 5% of the trees.
In this article, we put forward a methodology applying the anatomical and histochemical methods mentioned above but also involving the influence of the morphology of the host's ramification. The aim of this work is to establish a relationship between susceptibility to Loranthaceae, and either anatomical (phellem and cortical parenchyma thicknesses), or histochemical (tannin cell density) or morphological (the number of primary branches) characteristics.
Materials and methods
Study area and experimental design
Our data were mainly collected in the largest industrial rubber tree plantation of Mitzic and in three neighboring smallholding plantations named 1M2, M05 and M07.
The industrial plantation of Mitzic (lat. 0°50′ N, long. 11°50′ E, alt. 550 m) is divided into 14 sectors separated by swamps. Every sector consists of a variable number of plots. Generally, a plot is a surface comprising trees of a single clone, planted in the same year. However, AA01 and AA02 are clone trials, i.e. plots consisting of four replications obtained from one another by permuting six clones. Every clone of AA01 (AVROS 2037, GT 1, PB 217, PB 235, PR 261 and RRIM 600) contains 80 individuals, and every clone of AA02 (i.e. GGT 1, IRCA 109, IRCA 111, IRCA 130, RRIC 100 and RRIM 712), 108 individuals.
Whatever the type of plantation, trees are arranged in rows whose number varies according to the plot. However, distances between rows and between trees on a single row are constant, 7.5 m and 2.4 m, respectively.
Estimation of the parasitism level
To estimate the parasitism level of the rubber tree clones, a survey was performed in various plots (5/12, 4/2, 3/1, 7/5, 1/0 3/9, AA01 and AA02) of the industrial plantation of Mitzic and nearby smallholding plantations 1M2, M05 and M07. The sample thus obtained comprised 15,739 trees; for each of them, the number of clumps and the names of the Loranthaceae species were recorded. For a given clone, the parasitism rate (Pr) is defined as follows:
According to this measure of host susceptibility to the parasite, trees were distributed into three classes: non-parasitized (without visible clumps), moderately parasitized (with one to five clumps) and highly parasitized (with more than five clumps).
In the industrial plantation, the trees thus classed belonged to the six following clones: PB 217, GT 1, PB 235, AVROS 2037, PR 261 and RRIM 600. The smallholding plantation M07 comprises three clone types, PB 235, PB 260 and GT 1; whereas, the other smallholding plantations, 1M2 and M05, are just composed of one single clone type, GT 1.
First, a test for equality of proportions was performed to show whether susceptibility to Loranthaceae significantly differed from one clone to another. Then, a correspondence analysis (CA) was carried out to visualize the relationships between clones and susceptibility to Loranthaceae.
Artificial infestations
Artificial infestations were performed according to the protocol proposed by Frochot and Sallé (Citation1980), to follow the germination of P. capitata seeds and the attachment on the host, as well as to study the development kinetics of P. capitata on different rubber tree clones. Mature seeds of P. capitata were laid on branches of clones of variable susceptibility like AVROS 2037, GT 1, RRIM 600 and PB 260. After being submitted to outdoor drying, the viscin around the seeds hardens and allows the attachment on the host branches.
By means of one way-ANOVA tests, we compared – between the six clones – the percentage of germination and attachment on the one hand, and the length of the first leaves of the parasite, four months after infestation, on the other hand. This measure gave an idea about the parasite growth rate.
Morphometric, anatomical and histochemical study
For the morphometric study, the number of the ramifications of the trunk was established on seven clones: PB 217, PB 235, AVROS 2037, GT 1, PR 261, RRIM 600 and PB 260. The first six clones were in the AA01 clone trial and the last one in the farmer plantation M07. For each clone, 300 trees were measured.
For the statistical analysis, Friedman's test was used to estimate the role of the clone factor independently of the rank which each clone occupies in a row, using the formula:
A correspondence analysis enabled us to look for possible relationships between the clones and the number of branches, using Sturgess's criterion for establishing the K intervals of the discretized of the number of branches. The number K depends on N, the number of branches, according to the formula:
The anatomical and histochemical studies were carried out on non-parasitized branch fragments of 22-year-old rubber trees belonging to the six clones, PB 217, AVROS 2037, GT 1, PR 261, RRIM 600 and PB 260, each clone containing three individuals. Bark samples were fixed with a Regaud solution, which precipitates phenolic compounds that turn then into a brownish color.
Thirty µm thick sections were performed using a frozen microtome Jung AG Heibeberg type. For each individual, ten samples of about 25 mm2 section were cut. Thin sections were mounted in glycerin.
To estimate the tannin cell density in the ten branch fragments of the three individuals belonging to each clone studied, 35 sectors of 260 µm side length were delineated on a tracing paper. The number of tannin cells was estimated for each sector on the screen of a Reichert Visopan (Austria). A comparative study on the bark pertaining to the six rubber tree clones was performed to detect a possible anatomical variability in relation with susceptibility to Loranthaceae. Phellem and cortical parenchyma thicknesses were measured using a Reichert (Austria) micrometer objective for the ten sections observed.
Three one-way ANOVAs tested whether the susceptibility of the six clones differed significantly, each analysis being carried out for a given variable: phellem thickness, cortical parenchyma thickness or tannin cell density.
All the statistical analyses were performed by means of the R statistical processor (R Development Core Team Citation2005).
Results
Infestation level by Loranthaceae in the industrial and smallholding plantations of Mitzic
During our survey of the plantations studied, more than 15,000 trees scattered in the observed clones were studied and yielded the following information (): Parasitism rates varied between 19% and 91%, and differed significantly between the clones (F = 460, p<0.001).
Table 1. Parasitism rate (%) on rubber trees in the clones and various plot types in Mitzic's industrial and smallholding plantations. The identification of every plot meets the denomination applied by the managers of Mitzic's industrial and smallholding plantations. Every plot comprises only one clone type except for AA01 and AA02 (polyclonal trials).
Parasitism level remained very high for AVROS 2037, PB 235 and GT 1 in all the plantations prospected so far. Remarkably, no rubber tree clone was totally devoid of parasites. In these plantations, there existed no rubber tree clone completely resistant to one or another form of Loranthaceae. At the opposite of clone trial AA01 characterized by a very high parasitism rate (76%, see ), clone trial AA02 showed a much smaller one (26%). This discrepancy is easily explained; AA02 was subjected to a mechanical eradication of Loranthaceae in 1996, an event prior to which parasitism was rated low on young trees (3%).
The distribution of trees in three classes of susceptibility to the parasite (see Materials and methods section) was very useful for correspondence analyses (CA) to visualize the differences in the behavior of the clones facing the parasite. For the two types of plantation (the industrial and smallholding ones), correspondence analyses were carried out ( and ) and built up a good representation of the sets of clones and their corresponding susceptibility levels.
Figure 1. Correspondence analysis visualizing the responses of rubber tree clones to the Loranthaceae in the plots from Mitzic's industrial plantation. The indices a and b represent, respectively, clones planted in the plots not belonging to the clones trials, and clones planted in clone trial AA01. Triangles represent the susceptibility levels and circles, the clones. Respective inertia rates of the factorial axes F1 and F2: 87.2% and 12.8%. n.p., not parasitized; m.p., moderately parasitized; h.p., highly parasitized.
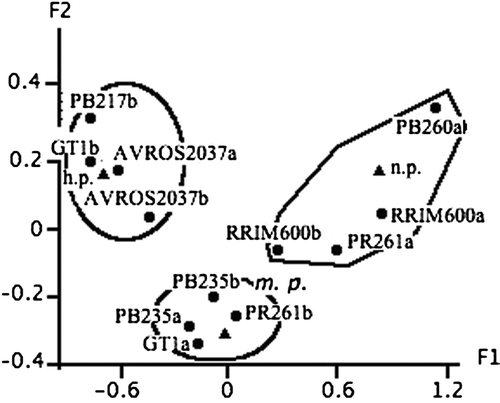
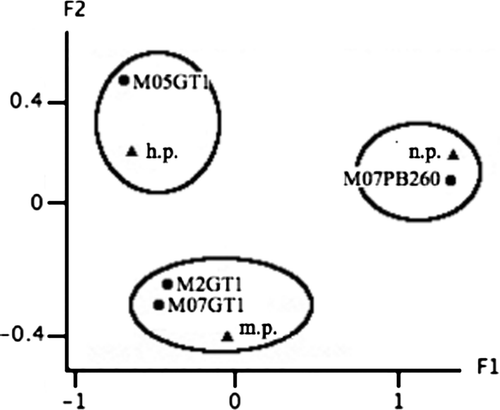
Figure 2. Correspondence analysis visualizing the responses of the clones to the Loranthaceae in the smallholding plantations. The triangles represent the susceptibility levels; and circles, the clones. Respective inertia rates of the factorial axes F1 and F2: 90.8% and 9.2%. n.p., not parasitized; m.p., moderately parasitized; h.p., highly parasitized.
It is noteworthy to specify that in the industrial plantation, clones planted in the monoclonal plots were labeled ‘a’, while the others belonging to the AA01 clone trials were labelled ‘b’. One of the CAs () showed that the clones RRIM 600a, PB 260a, PR 261a and RRIM 600b were lowly parasitized (meaning in this case that most of the trees from the clone are healthy), whereas PB235b, PB 235a, PR 261b and GT 1a were moderately parasitized, and AVROS 2037a, AVROS 2037b, PB 217b and GT 1b were highly parasitized.
The same statistical analysis performed in the smallholding plantations (), showed that the clone PB 260 was lowly parasitized, whereas GT 1 was moderately or highly parasitized.
The two analyses yielded very consistent results. In both cases, PB 260 was lowly parasitized and GT 1, moderately or highly parasitized. Thus, a susceptibility gradient could already be put forward, ranging from the most susceptible to the least susceptible: PB 217, AVROS 2037, GT 1, PB 235, PR 261, RRIM 600 and PB 260.
Development kinetics of Loranthaceae on several rubber tree clones
shows the development kinetics of Loranthaceae after artificial infestations performed on four rubber tree clones listed according to their decreasing susceptibility level (AVROS 2037, GT 1, RRIM 600 and PB 260). For all the clones, the percentage of germination was high, exceeding 75%, whereas the percentages of attachment on host branches were much lower, close to 30%. No significant difference was observed between the clones concerning the success of germination or attachment (the F values being, respectively, F = 1.4 (dfs = 3 and 710), p>0.05) and F = 0.4 (dfs = 3 and 208, p>0.05). Hence, no significant clone effect was detectable on germination or attachment success.
Table 2. Results from artificial infestations with seeds of P. capitata. Germination and attachment rates (% in brackets) are given by the following formulae:
However, when the development of the 212 plantlets (see ) was followed during four months ( and ), a significant ‘clone’ effect could be shown regarding the length of the first leaves (F = 54.9; df = 3 and 208; p<0.05). The plantlets developing on AVROS 2037 and GT 1 had longer leaves, respectively 38.8 and 22.1 mm, than those growing on PB 260 and RRIM 600, respectively 16.1 and 17.0 mm.
Figure 3. Development of the Phragmanthera capitata plantlets on branches belonging to four rubber tree clones. The circles represent the length in mm of the first leaves of the plantlets; and the vertical bars, the 95% confidence intervals of the mean values. Abscissae = the different clones, ordinates = First leaves lengths (FLL).
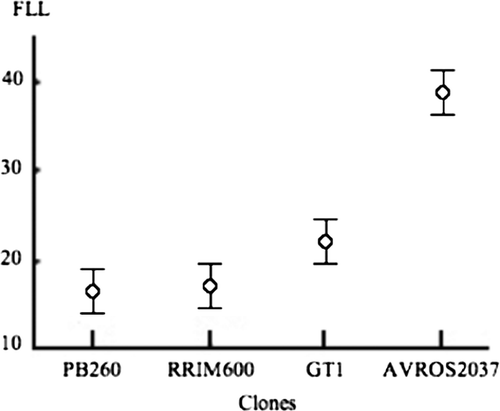
Table 3. Clone assignment to groups (Neuman-Keuls test). Each clone, characterized by a mean leaf length value, is assigned to one and only one among three labels, A, B or C. Each assignment corresponds to the clone's membership to a single specific interval whose leaf mean length varies between two bounds; the three intervals are adjacent, and their reunion equal to the interval of all the values taken on by the clones. The bounds are fixed by a criterion of the test.
The Neuman-Keuls test () identified three different groups of plantlets, which were assigned ‘A’, ‘B’ and ‘C’, the alphabetic order of these chosen letters being in parallel with the increasing order of the length of their first leaves, in turn in parallel with the increasing order of development speediness. Hence, we inferred that in the least susceptible clones RRIM 600 and PB 260 (the members of the A group), the development of the plantlets was slow, in comparison to the plantlet behavior in the other clones. Interestingly, the leaf development was more rapid in a moderately susceptible clone, GT 1 (member of the B group) and still faster in the most susceptible clone like AVROS 2037 (member of the C group).
This kinetic approach suggests that the more the rubber tree is susceptible to P. capitata, the more rapid the development of the parasite tends to be.
Ramification effect on the susceptibility of the different clones
This morphometric study treats the effect of the ramification of the first order on the susceptibility of six clones – AVROS 2037, GT 1, PB 235, PB 217, PR 261 and RRIM 600 – from the AA01 clone trial. The numbers of branches of the relevant ramifications were estimated on the clones just mentioned above and ranged from 4.6 to 16.3 (). Individuals of RRIM 600 and PR 261 were always much more ramified than the others. The Friedman's test confirmed these observations and indicated the existence of a ‘ramification’ effect on the susceptibility of the clones (Fr = 8.4, p<0.05).
Table 4. Mean number of branches per clone in the various plots. Clones RRIM 600 and PR 261 are lowly susceptible to the Loranthaceae, and clones AVROS 2037, PB 217 and GT 1 highly susceptible.
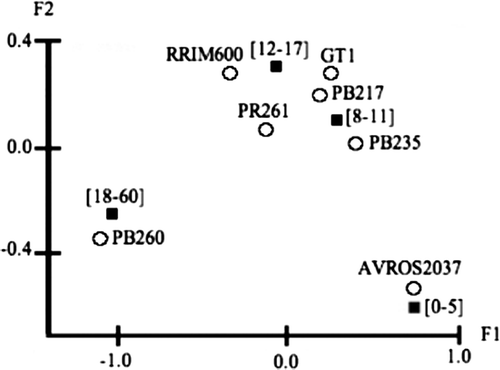
To carry out correspondence analysis on clones and ramification, the numbers of the observed branches of the first order were distributed in different classes according to the criterion of Sturgess. For this study, a seventh clone was added, PB 260, because of its very low level of parasitism (see ) and its absence in the AA01 clone trial. So far, four classes of ramification levels were determined: [0–5], [6–11], [12–17] and [18–60].
The analysis showed that trees belonging to the most susceptible clones like AVROS 2037, PB 235, PB 217 and GT 1 (), displayed very little ramification since they belonged to the [0–5] and the [6–11] classes. On the contrary, most individuals from the least susceptible clones like RRIM 600, PR 261 and PB 260, corresponded to the intervals including the highest ramification values [12–17] and [18–60]. Therefore, a relationship could be established between susceptibility and ramification, high ramification corresponding to a low parasitism rate.
Figure 4. Correspondence analysis visualizing the clone effect on the number of branches. The filled squares represent the number of branches (intervals in square brackets next to the symbols); and circles, the clones. Respective inertia rates of the factorial axes F1 and F2: 77.4% and 21.4%.
Location and density of the tannin cells
The location of tannin cells was studied on longitudinal radial sections of branches pertaining to six rubber tree clones. These compounds are located in the vacuole of the cortical parenchyma cells. Observations showed no differences in the density of tannin cells according to the clones.
A one-way ANOVA () was applied to each of the three following variables, i.e. phellem thickness, cortical parenchyma thickness and tannin cell density. According to the data set, no significant difference could be shown between each of the variables just mentioned and the clones. This statistical analysis is illustrated by the empirical diagram where a number of overlapping 95% confidence intervals has been displayed ().
Figure 5. Simultaneous representation of three variables. The filled squares represent the mean parenchyma thickness (µm); the empty circles, phellem thickness (µm); the filled square, cell density per 260×260 µm sector. Vertical bars represent the 95% confidence intervals of the mean lengths per clone.
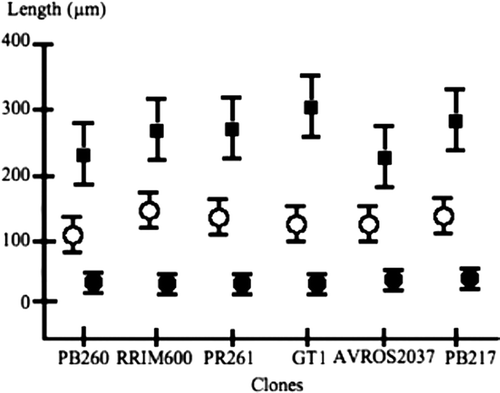
Table 5. One-way ANOVAs on the three variables.
Discussion
Data collected in the area of Mitzic, from the industrial and smallholding plantations of rubber trees, showed that Loranthaceae were very common and that among them Phragmanthera capitata was, by far, the most widespread (95%). The level of parasitism by P. capitata, estimated on more than 15,000 rubber trees, widely varied according to the clone. The clones AVROS 2037, PB 217, PB 235 and GT 1 were highly parasitized whereas PR 261, RRIM 600 and PB 260 were less attacked. This gradient of susceptibility is fully consistent with the previous observations of Guyot and Ntawanga (Citation1998) who found, in experimental plots of the C.A.T.H. (Centre d'Appui Technique à l'Hévéaculture), that whatever the type of plantation (industrial or smallholding), RRIM 600 was always less susceptible than PB 217 and GT 1. In our study, another clone, PB 260, turned out to be slightly susceptible to P. capitata.
It is noteworthy that rubber trees never displayed a complete resistance to mistletoe in contrast with some cultivars of poplars, such as Blanc de Garonne and Vereecken, totally resistant to Viscum album (Frochot et al. Citation1978; Armillotta Citation1984; Hariri Citation1989; Hariri et al. Citation1992), which never hosted mistletoe in natural conditions or after artificial infestations.
In the associations poplar/V. album and oak/V. album, several parameters involved in resistance were well established (Hariri Citation1989; Hariri et al. Citation1991): Anatomical features (phellem thickness, cortical parenchyma thickness), or biochemical characteristics (tannin-rich cell density). Moreover, it was shown on poplars that a positive correlation existed between these parameters and the behavior of mistletoe (Hariri Citation1989; Hariri et al. Citation1991, Citation1992): the higher the values of these three variables, the more resistant the cultivar was. As far as rubber tree is concerned, no relation between these parameters and the susceptibility to Loranthaceae could be observed. This is consistent with previous data showing the absence of a true resistance in rubber trees (Guyot and Ntawanga Citation1998; Engone Obiang Citation2006).
On the other hand, the ramification level is significantly correlated to the susceptibility to Loranthaceae. We established statistically that the most susceptible clones like AVROS 2037, PB 217, PB 235 and GT 1 showed few ramifications whereas the least susceptible like PR 261, RRIM 600 and PB 260 were the most branched. These results are in agreement with those of Combes and Du Plessix (Citation1974) who observed that RRIM 600 had the highest number of lateral buds, well known to generate branches. In addition, results concerning the development kinetics of P. capitata on different clones also support this idea. Plantlets developed much more rapidly on susceptible clones, like AVROS 2037. Accessibility of light to parasites could easily account for these observations. As it is well known that light is crucial for mistletoe development (Lamont and Perry Citation1977), there is evidence that rubber trees with few branches, therefore with sparse foliage, allow a better access of sunlight to parasites. We must notice that the high incidence of Loranthaceae in Mitzic plantations is the consequence of Colletotrichum leaf disease: The canopy of rubber trees has been strongly affected for many years with a harmful incidence on the foliage density.
During artificial infestations performed on different rubber tree clones, it was noted that percentages of germination and attachment on the host were not significantly different from one clone to another. These results match well with our knowledge of the biological cycle of Loranthaceae and Viscaceae: they germinate whatever the substratum (Sallé Citation1975; Clay et al. Citation1985). However, further development of plantlets requires a live and compatible host. Some of the reasons for the resistance lie in the development during the early growth of the haustorium (Hariri et al Citation1990, Citation1991; Sallé et al. Citation1993). In all poplar cultivars, it was shown that the onset of development of the primary haustorium provoked a breakdown of phellem and a division of the phellogen, the inner part giving rise to an inner phellem which enclosed the young primary haustorium. The perihaustorial zone formed at that stage contained polyphenolic substances. On the behavior of this mechanico-chemical barrier depends the success or the failure of the association. In very susceptible cultivars like Fritzy Pauley, the young primary haustorium passed rapidly through the inner phellem and the host secreted few polyphenols. On the contrary, in resistant cultivars such as Blanc de Garonne and Vereecken, the host reacted more intensively against the parasitic attack: The inner phellem was never broken and the synthesis of polyphenols was enhanced. In these conditions, the parasite never reached live host tissues and consequently the plantlet died.
In rubber trees, even in the least susceptible clones such as PB 260, histological sections performed on four-month-old plantlets (Engone Obiang Citation2006), showed that the young primary haustorium always reached the host live tissues, so that the plantlet could keep on growing.
Conclusion
Our study carried out on several rubber tree clones leads to four important conclusions:
-
In the association Hevea brasiliensis/Phagmanthera capitata, total resistance to Loranthaceae does not exist. A gradient of susceptibility can be proposed ranging from highly susceptible clones (AVROS 2037, GT 1, PB 217 and PB 235) to lowly susceptible ones (PR 261, RRIM 600 and PB 260).
-
The development of mistletoe is significantly and positively correlated to the susceptibility of the clone: the slower the parasite development, the less susceptible the clone is. However, the haustorium develops in all clones.
-
No positive correlation can be established significantly between susceptibility and the anatomical or biochemical characteristics studied here.
-
Susceptibility of rubber trees to Loranthaceae is negatively correlated to host ramification: the more ramified the rubber tree is, the slower the parasite develops and the less susceptible the clone is.
Acknowledgements
This study was supported by a grant from the Gabonese government and from the French Ministry of Foreign Affairs. We deeply thank S.I.A.T. (Gabon) and the French Rubber Institute for their financial help during our stay in the plantations of Mitzic.
References
- Akinlabi , AK and Aigbodion , I . 2005 . Characterisation of natural rubber latex from mistletoe-infested Hevea trees of NIG 804 clones . J Rub Res. , 8 : 153 – 159 .
- Armillotta A. 1984 . Etude de la résistance au gui (Viscum album L.). [Thesis dissertation] Paris : Université Pierre et Marie Curie .
- Balle , S and Hallé , N . 1961 . Les Loranthaceæ de la côte d'Ivoire . Adansonia. , 1 : 209 – 265 .
- Chambrier , HA . 1990 . L’économie du Gabon , Paris : Flammarion .
- Clay , K , Dement , D and Rejmanek , M . 1985 . Experimental evidence for host races in mistletoes (Phorandendron tomentosum) . Am J Bot. , 72 : 1225 – 1231 .
- Combes , JCL and du Plessix , CJ . 1974 . Etude du dévelopement morphologique de la couronne d’Hevea brasiliensis (Müll. Arg. Euphorbiacées-Crotonoïdées) . Ann Sci Forest. , 31 : 207 – 228 .
- Engone Obiang NL . 2006 . Biologie et écologie des Loranthaceæ parasites au Gabon. Impact de Phragmanthera capitata sur la productivité des hévéas [Doctorate thesis] . Paris, Université Pierre et Marie Curie .
- Engone Obiang , NL and Sallé , G . 2006 . Faut-il éradiquer Phragmanthera capitata, parasite des hévéas en Afrique? . C R Biol. , 329 : 185 – 195 .
- Frochot , H and Sallé , G . 1980 . Modalités de dissémination et d'implantation du gui . Rev Fr Forest. XXXII , 6 : 505 – 518 .
- Frochot H , Pitsch M , Wehrlen L . 1978 . 103e Congrès national des sociétés savantes, Nancy , France : Sciences, facs . I , p. 371 – 380 . Différence de sensibilité au gui (Viscum album L.) de quelques clones de peuplier (Populus sp.).
- Gill , LS and Onybé , HI . 1990 . Mistletoes on rubber trees in Nigeria . Haustorium , 23 : 1 – 2 .
- Grazi G , Urech K . 1983 . La susceptibilité des chênes, des ormes et des mélèzes au gui (Viscum album L.) . Rev scient. Bourbonnais : 6 – 12 .
- Guyot J , Ntawanga E . 1998 . Note sur la sensibilité de six clones d'hévéa à Phragmanthera capitata . Plant Rech Dév : 356 – 361 .
- Hariri , EB . 1989 . Polyphénols et résistance au gui. Etude histochimique et analytique [doctorate thesis] , Paris : Université Pierre et Marie Curie .
- Hariri , EB , Sallé , G and Andary , C . 1990 . Mécanismes de résistance de quatre cultivars de peupliers en réponse à l'attaque du gui (Viscum album L.) . C R Acad Sci Paris. , 311 : 439 – 444 .
- Hariri , EB , Sallé , G and Andary , C . 1991 . Involvement of flavonoids in the resistance of the poplar cultivars to mistletoe (Viscum album L.) . Protoplasma. , 162 : 20 – 26 .
- Hariri , EB , Jeune , B , Baudino , S , Urech , K and Sallé , G . 1992 . Elaboration d'un coefficient de résistance au gui chez le chêne . Can J Bot. , 70 : 1239 – 1246 .
- HEVECAM . 1995 . Rapport de la Direction Exploitation Agricole: Bilan de l'essai Loranthus (12-AGRO/DEA du 17/02/95) , p 9 .
- Kuijt , JL . 1969 . The biology of parasitic flowering plants , Berkeley and Los Angeles : University of California Press .
- Lamont , B and Perry , M . 1977 . The effects of light, osmotic potential and atmospheric gases on germination of mistletoe (Amyema preissii) . Ann Bot. , 41 : 203 – 209 .
- Polhill , R and Wiens , D . 1998 . Mistletoes of Africa , London : The Royal Botanic, Kew .
- Sallé , G . 1975 . Etude cytologique, cytochimique et histoautoradiographique du Viscum album L. (Loranthacée). I. La graine, sa germination et les modalités de fixation sur l'hôte . Rev Cyt Biol Vég. , 38 : 1 – 110 .
- R development Core Team . 2005 . R: A language and environment for statistical computing . Vienna, , Austria : R foundation for Statistical Computing .
- Sallé G . 1994 . Rapport de la mission «Loranthaceæ» réalisée au Cameroun . p 19 .
- Sallé , G , Armillotta , A and Frochot , H . 1984 . “ Proceeding of the 3rd International Symposium of Parasitic Weeds. Aleppo, Syria ” . In Mechanisms of resistance of four cultivars of poplar against Viscum album Edited by: Parker , C , Musselman , LJ , Polhill , RM and Wilson , AK . 21 – 30 .
- Sallé , G , Hariri , EB and Andary , C . 1993 . Bases structurales et biochimiques de la résistance au gui dans le genre Populus . Acta Bot Gallica. , 140 : 633 – 648 .
- Sallé , G , Tuquet , C and Raynal-Roques , A . 1998 . Biologie des Phanérogames parasites . C R Soc Biol. , 192 : 9 – 36 .
- Serier , JB . 1985 . L'hévéaculture au Gabon . Rev. Caoutchouc et Plastiques , 5 : 99 – 100 .
- Zhiwei , F , Xingguo , D , Yufang , Z and Yide , S . 1995 . Chemical control of Chinese Taxillus on rubber trees . The Planter, Kuala Lumpur. , 71 : 459 – 468 .