Abstract
How plant communities are structured, and the relative roles of gradients and disturbances in that structuring, has long been of interest. Here I use plots in replicate tallgrass, mixedgrass, and shortgrass prairies across Northern Oklahoma to address this issue by sampling plant percent cover three years after applying treatments realizing common prairie disturbances of burning, grazing, and soil turnover. I found (1) shortgrass plots had the least amount of aboveground biomass (AGB), with burning and soil turnover plots also having low AGB in the other two prairies, (2) tallgrass plots had the most total cover, with soil turnover plots having the least in the other two prairies, (3) tallgrass plots had the most species, with soil turnover plots having the least in the other two prairies, (4) control plots in mixedgrass had the smallest evenness and plots in shortgrass had the highest, and (5) a high degree of functional similarity in all three prairies. In addition to controlling these different aspects of population and community structure in prairies, results also show that the most severe disturbances can lead to a prairie plant composition and structure more similar to that found in the drier, most western prairie areas.
Keywords:
Introduction
Plants respond to environmental cues created both by landscape gradients (e.g. rainfall, temperature, elevation: Walter Citation1973; Whittaker Citation1975) and by disturbances (Gleason Citation1926; Pickett and White Citation1985). Indeed any current vegetation mosaic is largely influenced by the environmental variation on landscape gradients and by historical disturbances (Myster Citation2001). Grasslands are found throughout the world, and one of their most common types, prairies, form a major terrestrial ecosystem of the mid-continental United States (US) where they lie in a broad tract bordering deciduous forest on the east and desert/mountains on the west. These prairies have been intensely studied for many years (Collins and Adams Citation1983; Glenn and Collins Citation1993; Northup and Daniel 2002; Myster Citation2006; Haught and Myster Citation2008; Myster Citation2006; Weatherford and Myster Citation2011a, Citation2011b) and serve as flagship long-term ecological research sites of the National Science Foundation (e.g. Knapp et al. Citation1998; Myster Citation2011). Much of that research has focused on changes of resident prairie plants over space and time (Collins and Adams Citation1983; Polley and Collins Citation1984; Collins and Uno Citation1985; Glenn and Collins Citation1993; Risser et al. Citation1994; Briggs et al. Citation1998; Howe and Brown Citation1999; Silletti et al. Citation2004).
The vegetation in these prairies is greatly influenced by a decreasing rainfall gradient with increasing longitude across the Midwestern US landscape (Knapp et al. Citation1998). Across the northern part of Oklahoma, for example, tallgrass prairie gradually becomes mixedgrass prairie and finally shortgrass prairie [see Belsky (Citation1992) for a similar African savanna gradient]. This situation in Oklahoma presents an opportunity to combine all three prairie types in one study in order to compare and contrast how their plants respond to changes in precipitation and to common disturbances. Consequently, I set up replicate vegetative plots in all three prairie types across Northern Oklahoma and then sampled them three years after the application of the common prairie disturbances of burning, simulated grazing, and mammalian soil turnover (Blair Citation1997; Briggs et al. Citation1998).
Here I expand upon past studies that have examined how rainfall variation at different sites affects prairie structure and function (e.g. Vinton and Burke Citation1997; Gill and Burke Citation1999) and investigate the following questions: (1) How are the community structural parameters of aboveground biomass (AGB), total cover, species richness, and species evenness, affected by prairie-type defined by a precipitation gradient, affected by the application of common disturbances, and affected by the interaction of this gradient and these disturbances? (2) Which species and which disturbances are most important in defining variation in plant species responses among these plots? (3) How do the relative similarities among the plots and among species relate to their place along the rainfall gradient, and to the disturbance applied to them?, and (4) Do responses of prairie plant species correspond to common plant functional differences such as photosynthesis pathway, life form, and life-history?
Materials and methods
Six prairie remnant sites that span Northern Oklahoma were investigated, starting with two tallgrass prairie sites. The first tallgrass site was the Nature Conservancy's Tallgrass Prairie Preserve located in the Osage Hills of Northern Oklahoma outside Pawhuska, OK (36°75′N, 96°50′W). This site is 4000 acres in total size and characterized by shallow to deep soils over limestone and limy shale, which is often exposed (Coppedge et al. Citation1998). The average total annual precipitation for the area is 877 mm, most of which occurs between April and September (precipitation source for all study sites was Oklahoma Mesonet: www.mesonet.org). Whereas, management at the preserve includes grazing by bison (Bison spp.) and prescribed burning with a five-year return interval, neither disturbance occurred within 50 m of the study area during the time of the study (Coppedge et al. Citation1998). The second tallgrass site was the USDA-ARS Grazinglands Research Laboratory outside El Reno, Oklahoma (35°55′N, 97°96′W: Northup and Daniel Citation2000), with 32 acres set aside for research. The average annual precipitation there is 782 mm, and the soils are Norge silt loams: fine-silty, active thermic Udic Paleustolls (Daniel Citation2001). While prescribed burns are applied to pastures on a four-year cycle, the study plots were not within 50 m of any burning during the study. Common tallgrasses at both sites include big bluestem (Andropogon gerardii), little bluestem (Schizachyrium scoparium), indiangrass (Sorghastrum nutans), and switchgrass (Panicum virgatum).
Two mixedgrass prairie sites were also included. The first mixedgrass site was the USDA-ARS Southern Plains Range Research Station in Woodward, OK (36°25′N, 99°50′W), with 80 acres set aside for research. The second mixedgrass site was the Black Kettle National Grassland at Cheyenne, OK (35°64′N, 99°75′W) of a total size of 31,300 acres. For both sites, the mean March and September temperatures are between 4° and 6°C, 28° and 30°C with an average annual precipitation of 619 and 632 mm respectively. Soil at the Woodward site is a Carey silt loam from thermic Typic Argiustolls, and soil at the Cheyenne site is of the Woodward-Quinlan association, loamy and underlain by Udic Paleustolls. Grazing is not allowed on these lands but burning is prescribed and was performed five years before the experiment started, but not within 50 m of any plots. Mixedgrass prairies contain species from both tallgrass and shortgrass prairies.
Finally the two shortgrass prairie sites included the Optima Wildlife Refuge (36°50′N, 101°25′W) outside Hardesty, OK, with a total size of 4333 acres and an annual rainfall of 450 mm, and the Rita Blanca National Grassland (36°45′N, 102°75′W) outside Boise City, OK, with a total size of 230,000 acres and an annual rainfall of 401 mm. Neither grazing nor burning was performed on these lands. Common to the shortgrass prairie is the shrub sand sage (Artemisia filifolia) and the grasses blue grama (Bouteloua gracilis), western wheatgrass (Agropyron smithii), and buffalograss (Bouteloua dactyloides).
In spring 2002, 20 circular plots of radius 0.9 m (bigger than the quadrates used in Collins and Barber Citation1985; Collins Citation1987) were placed randomly, but away from any edges, in each of the six study sites for a total of 120 plots. Within each study site, the 20 plots were randomly assigned one of four treatments at the same time, so each treatment was replicated five times: (1) untreated control plots, (2) plots clipped once down to 5 cm (the natural grazing zone: Wallace Citation1981) with the biomass removed to simulate grazing without plant preference, (3) plots burned with the aid of a charcoal gas match in spring 2002 (mimicking the timing of natural lightning fires: Howe Citation1994; Bragg Citation1995), and (4) plots with the soil simply turned over to a depth of 15 cm (Belsky Citation1992) mimicking the burrowing action of mammals (Collins and Barber Citation1985) which can affect as much as 30% of a prairie's ground surface per year (Kyle et al. Citation2008). Plots in all three prairies were at least 6 m apart and burning was done by professionals on site.
The plots were harvested in August 2005, after allowing them over three years to respond to the treatments (the natural fire frequency in these areas: Fuhlendorf and Engle Citation2001). For each plot, percent cover of all plant species (visit http://plants.usda.gov for plant nomenclature ) was visually scored using a circular wooden plant frame. In addition AGB was collected in each plot by clipping, drying under room conditions in the laboratory for 30 days (Weatherford and Myster Citation2011a, Citation2011b), and then weighing to the nearest 0.1 g.
A two-way analysis of variance (ANOVA: SAS 1985) was used to analyze the data as a split-plot design with prairie-type (tallgrass, mixedgrass, shortgrass) and disturbance (control, clipped, burned, soil turned over), each treated as main effects. Interaction between these two main effects was also investigated. Plots were combined within each prairie type so that there were 10 replicates for each prairie type×disturbance cell. The response variables were AGB, total cover, species richness (the total number of species), and species evenness (using Pielou's J index: Ludwig and Reynolds Citation1988). If ANOVA results were significant, means testing was performed using the Ryan-Elinot-Gabriel-Welsch multiple range test (SAS Citation1985). In addition, a principal components analysis (PCA: SAS Citation1985; Myster and Walker Citation1997), suggested for grassland data-sets (Belsky Citation1992), was also performed. The data matrix was composed of the plots (defining the columns) and the plant species (defining the rows), with percent cover as the data in each cell of the matirx. Both R-type (plots preserved) and Q-type (species preserved) ordinations were performed using the correlation matrix for the computations. Q-type ordination species cluster patterns were then examined in order to explore possible plant functional types. This approach defined clusters and suggested important plant traits directly through the use of the sampled data, rather than selecting traits beforehand and investigating whether the data fit into them or not (see Grime Citation1995). Data were inspected before analysis and found to be normally distributed. All results are based on a single year of sampling.
Results
The control plots in tallgrass prairie were dominated by Bromus spp., Digitaria cognata, and Schizachyrium scoparium in mixedgrass prairie by Bouteloua curtipendula, and in shortgrass prairie by Eragrostis intermedia, Hordeum pusillum, and Boutelou spp. (). Burning in the tallgrass prairie led to more cover of Andropogon gerardii, Curtipendula gracilis, and Bothriochloa ischaemum but a decrease in Schizachyrium scoparium. All three disturbances in the mixedgrass prairie increased Panicum virgatum but decreased Bouteloua curtipendula. Clipping and burning in the shortgrass prairie increased the amount of Aristida spp., while burning increased Bothriochloa laguroides (). For the important exotic grass Bromus tectorum, it increased only after soil disturbance in the shortgrass prairie compared to controls ().
Table 1. Total cover for all plant species sampled in the plots (e.g., the sum of percent cover for all 5 replicate plots for each prairie type x disturbance type combination). Disturbance types are indicated as Co (control), Cl (clipping), Bu (burning), and St (soil turnover).
Aboveground biomass was significantly affected by both prairie type and disturbance type, and a significant interaction effect was also seen (). Both clipping and burning increased AGB in shortgrass plots (A) while burning and soil turnover reduced AGB in mixedgrass and tallgrass plots (B). The examination of the interaction effect showed that in the shortgrass plots both clipping and burning actually increased AGB (C). Total percent cover was significantly affected by prairie type and by disturbance type (). While the tallgrass prairie had the greatest total percent cover, the shortgrass and mixedgrass prairies had similar amounts (A). Plots that had their soil turned over had the least cover across all three prairie types (B). The other disturbance treatments were similar to the controls. Species richness was significantly affected by prairie type and by disturbance type (). The tallgrass prairie had the highest species richness (A), and the plots that had their soil turned over had the lowest regardless of prairie type (B). The highest species evenness was found after burning in the tallgrass and the lowest occurred on control plots in mixedgrass (A, 4B). All other treatment and prairie type combinations were similar. A significant interaction between prairie type and disturbance type was recorded for species evenness ().
Figure 1. Mean and standard error of levels of aboveground biomass by: (A) prairie type; (B) disturbance; and (C) interaction. Significantly different treatment levels are indicated by different lower case letters using the Ryan-Elinot-Gabriel-Welsch multiple range test (SAS Citation1985). Disturbances are indicated as control (co), clipping (cl), burning (bu), and soil turnover (st).
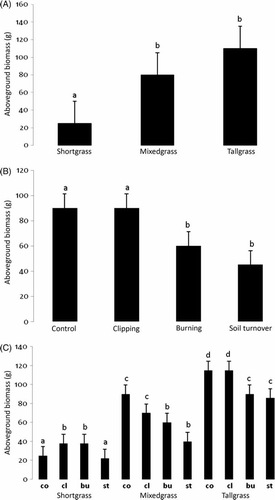
Figure 2. Mean and standard error of levels of total percent cover by: (A) prairie type and (B) disturbance. Significantly different treatment levels are indicated by different lower case letters using the Ryan-Elinot-Gabriel-Welsch multiple range test (SAS Citation1985).
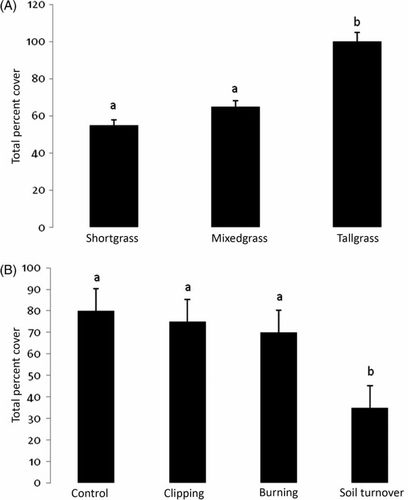
Figure 3. Mean and standard error of levels of species richness by: (A) prairie type and (B) disturbance. Significantly different treatment levels are indicated by different lower case letters using the Ryan-Elinot-Gabriel-Welsch multiple range test (SAS Citation1985).
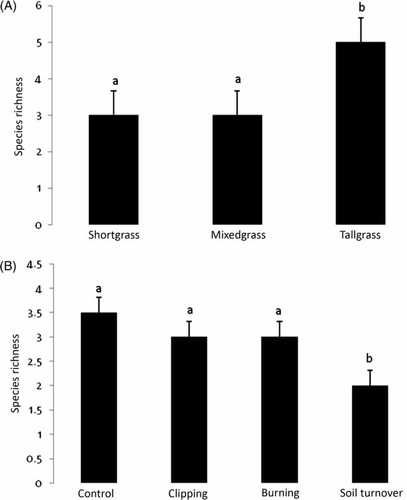
Figure 4. Mean and standard error of levels of species evenness by: (A) prairie type and (B) interaction. Significantly different treatment levels are indicated by different lower case letters using the Ryan-Elinot-Gabriel-Welsch multiple range test (SAS Citation1985). Disturbances are indicated as control (co), clipping (cl), burning (bu), and soil turnover (st).
Table 2. F statistic summary table for the two-way ANOVA performed with a main effect of prairie type, a main effect of disturbance, and an interaction of prairie type and disturbance. The response variables were above-ground biomass, total cover, species richness (total number of species), and species evenness (using Pielou's J index: Ludwig & Reynolds 1988).
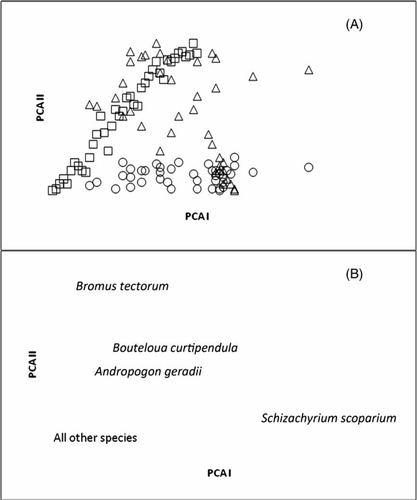
Within the R-type PCA ordination, the first two axes explained 58% of the original variation in the data matrix. PCA axis I was mainly defined by Schizachyrium scoparium (Pearson product-moment correlation coefficient [ppmcc] = − 0.895), and PCA axis II was also mainly defined by Schizachyrium scoparium (ppmcc = 0.308). The PCA separated shortgrass prairie and tallgrass prairie well. Some mixedgrass plots are located with the shortgrass plots; however, some mixedgrass plots are located with the tallgrass plots, and some stand alone (A). Those mixedgrass plots that were subjected to the most severe disturbance, soil turnover, are most located with the shortgrass plots, likewise, the tallgrass plots subjected to soil turnover were closest to the control mixedgrass plots. In the Q-type PCA ordination, the first two axes explained 57% of the original variation. PCA axis I was mainly defined by the mixedgrass control plots (ppmcc = 0.239) and the mixedgrass burned plots (ppmcc = 0.235). PCA axis II was mainly defined by the shortgrass clipped plots (ppmcc = 0.280), the burned plots (ppmcc = 0.273), and the soil turnover plots (ppmcc = 0.270). Most species formed a tight clump (B) but cheatgrass (Bromus tectorum), sideoats grama (Bouteloua curtipendula), big bluestem (Andropogon gerardii), and little bluestem (Schizachyrium scoparium) are located separately implying distinct functional significance for photosynthetic pathway (C3 vs. C4), life history (annual vs. perennial), and life form (forb vs. grass).
Figure 5. (A) PCA R-type ordination indicating shortgrass plots (open square), mixedgrass plots (open triangle) and tallgrass plots (open circle), and (B) PCA Q-type ordination where species are labeled with their scientific names.
Discussion
When the percent cover of plants in the plots was divided by prairie type and by disturbance type, there was agreement with these other grassland results: (1) burning results in lower B. tectorum cover (Collins Citation1987); (2) grazing reduces Schizachyrium scoparium (Penfound Citation1955); and (3) burning increases the growth of tall grasses, such as Andropogon gerardii (Towne and Owensby Citation1984; Collins Citation1987). The AGB, as a correlate of net primary productivity, was related to historical precipitation averages (also see Briggs and Knapp Citation1995) because production increased with greater annual rainfall that the mixedgrass and tallgrass prairies received. The total cover also increased as prairies received more rainfall. There was also some support in the data for grazing/defoliation increasing production, especially in shortgrass prairie (Kelting Citation1954; Collins and Barber Citation1985). There was support in the data for soil turnover having greater effects on cover than fire (Belsky Citation1992), but grazing did not lead to more cover (Kelting Citation1954). With the exception of shortgrass prairie, however, burning did not increase productivity, as predicted (Adams and Anderson Citation1978; Collins and Barber Citation1985; Hulbert Citation1988; Suding, Citation2001). In general, disturbances did not seem to ‘maintain’ productivity and diversity in grasslands (Blair Citation1997) where the severe disturbances often decrease richness, although some studies suggest that certain disturbances may increase it (Kelting Citation1954; Netherland Citation1979; Collins and Barber Citation1985).
The R-type PCA showed that plant responses to different disturbances within prairies change the relative location of plots within the PCA, which suggests that plants are presented with similar cues. The burned plots and the clipped plots covered the entire spectrum of prairie types, with the control plots staying close to the PCA center for each of the three prairie types. The plants’ response to the most severe disturbance of soil turnover shows that mixedgrass prairie can have the same plant species as shortgrass prairie if subjected to a high-intensity disturbance, and tallgrass have the same as mixedgrass. As the correlations in the R-type ordination show that Schizachyrium scoparium is a defining species among these plots. Also, Bouteloua curtipendula was found only in mixedgrass and shortgrass plots, and Andropogon gerardii was found exclusively in tallgrass plots (Kindscher and Wells Citation1995).
The Q-type PCA suggests that there are different and distinct plant functional types in prairies. By mapping these known morphological, anatomical, physiological, and ecological traits on the PCA patterns, certain plant types emerged (Kindscher and Wells Citation1995). Prairie species group mainly by metabolism and life form (Coppedge et al. Citation1998), so that Bromus tectorum is the only annual grass among the other perennial grasses and the only C3 grass among the other grasses which are all C4 grasses (Adams and Wallace Citation1984). In addition, B. tectorum is an exotic species that is rapidly invading many prairie areas and affecting numerous ecosystem properties (Ogle and Reiners Citation2003), including productivity and decomposition (Myster 2002; Ogle et al. Citation2004). B. tectorum dominates through superior competitive abilities against seedlings but, as shown in the results, may need disturbance to invade intact perennial plant communities (Humphrey and Schupp Citation2004). The results imply a large amount of functional redundancy among these species (Myster 2002).
Conclusion
To summarize the questions posed in the Introduction: (1) Both clipping and burning increased AGB in shortgrass plots while burning and soil turnover reduced AGB in mixedgrass and tallgrass plots, and plots that had their soil turned over had the least cover across all three prairie types. The tallgrass prairie had the highest species richness, and the plots that had their soil turned over had the lowest regardless of prairie type. The highest species evenness was found after burning in the tallgrass and the lowest occurred on control plots in mixedgrass, (2) While both PCA axis I and PCA axis II of the R-type ordination were mainly defined by Schizachyrium scoparium, PCA axis I of the Q-type ordination was mainly defined by mixedgrass control and burned plots, (3) Those mixedgrass plots that were subjected to the most severe disturbance, soil turnover, were mostly located with the shortgrass plots, likewise, the tallgrass plots subjected to soil turnover were closest to the control mixedgrass plots, and (4) While results implied a large amount of functional redundancy among these species, there was also functional significance for photosynthetic pathway, life-history, and life form.
Results suggest that these three prairie types may be in a constant state of flux, depending on the disturbance being applied and the structural parameter being investigated where yearly variation in precipitation could also be a determining factor. Such ‘fluidity’ may be particularly common when the dominant plants are wind-dispersed, as in these prairies, and can easily travel among different communities without being influenced by changes in animal behavior and animal population dynamics. Disturbances may change the set of cues presented to plants by gradients (Myster Citation2001) leading, in this case, to the growth of plants that belong to another prairie type. Severe disturbance, like soil turnover and burning as seen here, may most likely produce structural changes including ‘cold spots’ within prairies with reduced species richness and plant cover.
Acknowledgements
The author thanks B. Hamilton, B. Northup, C. Milner, R. Gillen, D. Maple, J. Young, N. Wallis, and J. Hall for their assistance in setting up experiments. The author also thanks J. Briggs, B. Northup, and R. Gillen for commenting on a previous draft of the manuscript.
References
- Adams , DE and Anderson , RC. 1978 . The response of a central Oklahoma grassland to burning . SW Nat. , 23 : 623 – 632 .
- Adams , DE and Wallace , LL. 1984 . Nutrient and biomass allocation in five grass species in an Oklahoma tallgrass prairie . Am Midl Nat. , 113 : 170 – 181 .
- Belsky , AJ. 1992 . Effects of grazing, competition, disturbance and fire on species composition and diversity in grassland communities . J Veg Sci. , 3 : 187 – 200 .
- Blair , JM. 1997 . Fire, N availability and plant response in grasslands: a test of the transient maxima hypothesis . Ecology. , 78 : 2539 – 2568 .
- Bragg , TB. 1995 . “ The physical environment of Great Plains grasslands ” . In The changing prairie , Edited by: Joern , J and Keeler , KH . 49 – 81 . Oxford , , UK : Oxford University Press .
- Briggs , JM and Knapp , AK. 1995 . Interannual variability in primary production in tallgrass prairie: climate, soil moisture, topographic position, and fire as determinants of aboveground biomass . Am J Bot. , 82 : 1024 – 1030 .
- Briggs , JM , Nellis , MD and Turner , CL. 1998 . “ A landscape perspective of patterns and processes in tallgrass prairie ” . In Grasslands dynamics: long-term ecological research in tallgrass prairie , Edited by: Knapp , AK , Briggs , JM , Hartnett , DC and Collins , SL . 265 – 279 . New York , NY : Oxford University Press .
- Collins , SL. 1987 . Interaction of disturbances in tallgrass prairie: a field experiment . Ecology. , 68 : 1243 – 1250 .
- Collins , SL and Adams , DE. 1983 . Succession in grasslands: thirty-two years of change in a central Oklahoma tall-grass prairie . Vegetatio. , 51 : 181 – 190 .
- Collins , SL and Barber , SC. 1985 . Effects of disturbance on diversity in mixedgrass prairie . Vegetatio. , 64 : 87 – 94 .
- Collins , SL and Uno , GE. 1985 . Seed predation, seed dispersal and disturbance in grasslands: a comment . Am Nat. , 125 : 866 – 872 .
- Coppedge , BR , Engle , DM , Toepfer , CS and Shaw , JH. 1998 . Effects of seasonal fire, bison grazing and climatic variation on tallgrass prairie vegetation . Plant Ecol. , 139 : 235 – 246 .
- Daniel JA. 2001 . The water resources and erosion watersheds, Fort Reno, OK . USDA-ARS Grazinglands Research Laboratory Report GRL1-01 .
- Fuhlendorf , SD and Engle , DM. 2001 . Restoring heterogeneity on rangelands: ecosystem management based on evolutionary grazing patterns . Bioscience. , 51 : 625 – 632 .
- Gill , RA and Burke , IC. 1999 . Ecosystem consequences of plant form changes at three sites in the semiarid United States . Oecologia. , 121 : 551 – 563 .
- Gleason , HA. 1926 . The individualistic concept of the plant association . Bull Torrey Bot Club. , 53 : 7 – 26 .
- Glenn , SM and Collins , SL. 1993 . Experimental analysis of patch dynamics in tallgrass prairie plant communities . J Veg Sci. , 4 : 157 – 162 .
- Grime , JP. 1995 . Functional types: a comment on Steneck and Dethier (1994) . Oikos. , 73 : 120 – 121 .
- Haught , JA and Myster , RW. 2008 . Effects of species, density, season and prairie-type on seed predation in Oklahoma . Am Midl Nat. , 159 : 482 – 488 .
- Howe , HF. 1994 . Response of early- and late-flowering plants to fire season in experimental prairies . Ecol Appl. , 4 : 121 – 133 .
- Howe , HF and Brown , JS. 1999 . Effects of birds and rodents on synthetic tallgrass communities . Ecology. , 80 : 1776 – 1781 .
- Hulbert , LC. 1988 . Causes of fire effects in tallgrass prairie . Ecology. , 69 : 46 – 58 .
- Humphrey , LD and Schupp , EW. 2004 . Competition as a barrier to establishment of a native perennial grass (Elymus elymoides) in alien annual grass (Bromus tectorum) communities . J Arid Env. , 58 : 405 – 422 .
- Kelting , RW. 1954 . Effects of moderate grazing on the composition and plant production of a native tallgrass prairie in central Oklahoma . Ecology. , 35 : 200 – 207 .
- Kindscher , K and Wells , PV. 1995 . Prairie plant guilds: a multivariate analysis of prairie species based on ecological and morphological traits . Vegetatio. , 117 : 29 – 50 .
- Knapp , AK , Briggs , JM and Hartnett , DC. 1998 . Grassland dynamics: long-term ecological research in Tallgrass prairie , Oxford , , UK : Oxford University Press .
- Kyle , GP , Kulmatiski , A and Beard , KH. 2008 . Influence of pocket gopher mounds on nonnative plant establishment in a shrub steppe ecosystem . W N Am Nat. , 68 : 374 – 383 .
- Ludwig , JA and Reynolds , JF. 1988 . Statistical ecology: a primer on methods and computing , New York , NY : Wiley & Sons .
- Myster , RW. 2001 . Mechanisms of plant response to gradients and after disturbances . Bot Rev. , 67 : 441 – 452 .
- Myster , RW. 2006 . Species-specific effects of grass litter mass and type on emergence of three tall grass prairie species . Ecoscience. , 13 : 95 – 99 .
- Myster , RW. 2011 . Above-ground vs. below-ground interactive effects of mammalian herbivory on tallgrass prairie plant and soil parameters . J plant Inter. , 8 : 1 – 8 .
- Myster , RW and Walker , LR. 1997 . Plant successional pathways on Puerto Rican landslides . J Trop Ecol. , 13 : 165 – 173 .
- Netherland , L. 1979 . The effect of disturbances in tallgrass prairie sites on an index of diversity and equitability . SW Nat. , 24 : 267 – 274 .
- Northup , BK and Daniel , JA. 2000 . The impacts of climate and management on species composition of a southern tallgrass prairie in Oklahoma . PNCGL. , 1 : 693 – 699 .
- Ogle , SM , Ojima , D and Reiners , WA. 2004 . Modeling the impact of exotic annual brome grasses on soil organic carbon storage in a northern mixedgrass prairie . Biol Inva. , 6 : 365 – 377 .
- Ogle , SM and Reiners , WA. 2003 . Impacts of exotic annual brome grass (Bromus spp.) on ecosystem properties of northern mixedgrass prairie . Am Midl Nat. , 149 : 46 – 58 .
- Penfound , WT. 1955 . The relation of grazing to plant succession in the tallgrass prairie . Ecology. , 36 : 86 – 94 .
- Pickett , STA and White , PS. 1985 . The ecology of natural disturbance and patch dynamics , New York , NY : Academic Press .
- Polley , HW and Collins , SL. 1984 . Relationships of vegetation and environment in Buffalo wallows . Am Midl Nat. , 112 : 178 – 186 .
- Risser , PG , Birney , EC , Blocker , HD , May , SW , Parton , WJ and Wiens , JA. 1994 . The true prairie ecosystem , Stroudsburg , PA : Hutchinson Ross .
- SAS 1985 . User's guide: statistics , 5th ed . Cary , NC : SAS Institute Inc .
- Silletti , AM , Knapp , AK and Blair , JM. 2004 . Competition and coexistence in grassland codominants: responses to neighbor removal and resource availability . Can J Bot. , 82 : 450 – 460 .
- Suding , KN. 2001 . The effect of spring burning on competitive ranking of prairie species . J Veg Sci. , 12 : 849 – 856 .
- Towne , G and Owensby , C. 1984 . Long-term effects of annual burning at different dates in ungrazed Kansas tallgrass prairie . J Range Manag. , 37 : 392 – 397 .
- Vinton , MA and Burke , IC. 1997 . Contingent effects of plant species on soils along a regional moisture gradient in the great plains . Oecologia. , 110 : 393 – 402 .
- Wallace , LL. 1981 . Growth, morphology and gas exchange of mycorrhizal and nonmycorrhizal Panicum coloratum L, a C4 grass species, under different clipping and fertilization regimes . Oecologia. , 49 : 272 – 278 .
- Walter , H. 1973 . Vegetation of the earth and the ecological systems of the geo-biosphere , New York , NY : Springer-Verlag .
- Weatherford JL , Myster RW . 2011b . Effects of species, water, and nitrogen on competition among three Prairie Grasses. Prairie Nat .
- Weatherford , JL and Myster , RW. 2011a . Interactive effects of simulated grazing, species and below-ground resources water on competitive outcome among three prairie grasses . J Torr Bot Soc. , 138 : 107 – 119 .
- Whittaker , RH. 1975 . Communities and ecosystems , New York , NY : Macmillan .