Abstract
Gibberellins (GAs) are well known for plant growth promotion. GAs production by fungi has received little attention, although substantial work has been carried out on other aspects of plant growth-promoting fungi (PGPF). We investigated GAs production and plant growth-promoting capacity of an endophytic fungus isolated from the roots of soil grown soybean plants. The endophytic fungus is reported as GAs producer and as PGPF for the first time in this study. Nine endophytic isolates were collected from the roots of soybean, and culture filtrates (CFs) obtained from their pure cultures were screened on Waito-C, a dwarf rice cultivar, for the presence of GAs. Of these, seven fungal isolates promoted shoot length as compared to control (distilled water), while one inhibited it. Three fungal isolates were selected on the basis of higher shoot elongation as compared to wild type Gibberella fujikuroi, which was used as positive control. The growth-prompting capacity of selected fungal isolates SB5-1, SB3-2, and SB3-3 was bio-assayed on soybean cv. Hwangkeumkong. Fungal isolate SB5-1 provided maximum plant height (31.6 cm), shoot length (21.1 cm), whole plant fresh biomass (2.41 g), shoot fresh biomass (1.99 g), and leaf area (24.37 cm2). The CF of isolate SB5-1 was analyzed for the presence of GAs, and it was found that all physiologically active GAs were present (GA1, 0.15 ng/ml, GA3, 1.2 ng/ml, GA4, 7.37 ng/ml, and GA7, 3.18 ng/ml) in conjunction with physiologically inactive GA5, GA9, GA15, GA19, GA20, and GA24. The fungal isolate SB5-1 was identified as a new strain of Cephalotheca sulfurea through molecular and phylogenetic approaches.
Introduction
Endophytic fungi have been shown to confer benefits to host plants. Fungi may improve the ecological adaptability of the host by enhancing their tolerance to biotic and abiotic stresses (Schulz and Boyle Citation2005), like tolerance to herbivory, heat, salt, disease, and drought, and an increased biomass both above and below the ground (Waller et al. Citation2005; Márquez et al. Citation2007). Cephalotheca is a saprophytic fungus inhabiting soil, wood, and mushrooms. Cephalotheca is a little known genus, which exhibits both anamorph (asexual reproduction) and teleomorph (sexual reproduction). Cephalotheca sulfurea living on wood and mushrooms is characterized by nonciliated cleistothecia, oval-shaped ascospores and it has not yet reported to cause human infections (Suh et al. Citation2006).
Gibberellins (GAs) are diterpenoid plant hormones, first detected in the 1920s from culture filtrates (CFs) of Gibberella fujikuroi, a known pathogen of rice plants (Ogas Citation2000). GAs appear to be involved in every aspect of plant growth and development, but their most typical (and spectacular) property is the enhancement of stem growth (Nishijima et al. Citation1995). GAs may modify the sex expression of flowers, induce the parthenocarpic development of fruit, and delay senescence. They obviate the need for exposure to red light in the germination of seeds and spores, and the need for vernalization in the growth of bulbs and tubers. They are associated with the breaking of winter dormancy and stimulate the formation of hydrolytic enzymes in germinating cereal grain (Martin Citation1983). Currently, 136 GAs have been identified, while 12 fungi, pathogenic and non-pathogenic, associated with plants and/or soil has been reported as GA producers (MacMillan Citation2002; Kawaide Citation2006; Vandenbussche et al. Citation2007). New strains of Penicillium citrinum, Cladosporium, Phoma, and Aspergillus fumigatus had also been reported as GA producers (Khan et al. Citation2008; Hamayun et al. Citation2010a, Citation2010b; Khan et al. Citation2011).
There is an increasing concern about the excessive use of fertilizer and pesticides in agricultural fields and their subsequent negative impact on environment. Use of plant growth-promoting fungi for higher crop production, while reducing the inputs of fertilizer and pesticides may possess great potential for crops, most of which respond very well to fungal inoculation. Current study was aimed to identify potential fungal inoculums for plant growth promotion to decrease the use of fertilizer in agricultural fields.
Materials and methods
Growth conditions, sample collection, and isolation of endophytic fungi
Endophytic fungi were isolated from the roots of soybean cultivar Hwangkeumkong, which was grown in soil under green-house conditions. For isolation of endophytic fungi, soybean plants were randomly selected and the fine roots close to root tips were washed out. The root samples were then suspended in Tween 80 solution (2–3 drops in 50 ml of distilled water) and placed in a shaking incubator set at 120 rpm for 5 min at room temperature. The roots were then rinsed in distilled water to remove any residual Tween 80 (detergent). The cleaned samples were then surface sterilized by suspending them in 50 ml of 1% perchloric acid, and placed on shaking incubator (120 rpm for 5 min). The roots were then washed with autoclaved distilled H2O, dried with sterile filter paper, cut into pieces (0.5 cm each), cultured on Hagem media plates, and incubated at 25°C until the emergence of fungal cells (Khan et al. Citation2008). The Hagem media plates were supplemented with 80 µg/ml streptomycin (Yamada et al. Citation2001), to avoid bacterial growth in the medium. To obtain true endophytic fungi, the effectiveness of the surface sterilization procedure was tested by the imprinting technique (Schulz et al. Citation1999), and sterilized root pieces placed on Hagem media plates. Absence of any microbial growth on imprinted media plates after 4–7 days of incubation confirmed the effectiveness of surface sterilization (Khan et al. Citation2008). Pure fungal cultures were isolated, grown on potato dextrose agar (PDA) media plates, and slants. The PDA slants were used for storage purpose. Czapek broth medium, containing 1% glucose and peptone was used for GA production (Hasan Citation2002) by incubating the fungal isolates at 30°C at 120 rpm for 7 days. The wild type strain of G. fujikuroi was used as a positive control for GAs production during this experiment. Pure cultures of G. fujikuroi were grown on Czapek broth. G. fujikuroi was provided by the Korean Culture Center of Microorganisms (KCCM).
Bioassay on Waito-C and soybean
The CFs of fungal isolates were bioassayed on Waito-C sprouts for the presence of plant growth-promoting metabolites. Seeds of Waito-C were surface sterilized and treated with 20 µg/ml uniconazol for 24 h, to check the GA biosynthesis. The treated seeds were washed thoroughly and soaked in autoclaved distilled H2O for germination. The young seedlings were transplanted in glass tubes containing a 0.8% water-agar medium and kept in a growth chamber. Forty milliliter of culture fluid was centrifuged at 5000×g at 4°C for 15 min and the resulting pellet and supernatant were immediately stored at –70°C and later lyophilized (ISE Bondiro Freeze dryer). The lyophilized supernatant was mixed with 1 ml autoclaved distilled water (DW) and 10 µl of supernatant solution was applied on apical meristem of 10 Waito-C rice seedlings at two leaf stage (Khan et al. Citation2008). The shoot and plant lengths were observed for 7 days after the application and compared with Waito-C seedlings treated either with DW or CF of G. fujikuroi. The screening experiment comprised three replicates.
In a separate experiment, seeds of soybean cv. Hwangkeumkong were surface sterilized with 5% NaClO for 15 min and then washed with DW. Seeds were sown in autoclaved perlite and 20 ml of Hoagland solution (Hoagland and Arnon Citation1950) was applied at germination time. Fungal isolates SB5-1, SB3-2, and SB3-3 were selected on the basis of maximum shoot length promotion of Waito-C, and 5 ml of fungal supernatant was applied to soybean seedlings at two leaf stage. The plant length, shoot length, plant fresh weight, and shoot fresh weight were measured after one week of CF application. Shoot length of intact plants was measured and the plants were then taken out of the perlite to measure the whole plant length with the help of a measuring tape. The leaf area was calculated with CI-203 Portable Laser Area Meter (CID Inc. USA), while the chlorophyll content of fully expanded leaves was analyzed with the help of chlorophyll meter (Minolta Co., Ltd., Japan).
Extraction and quantification of fungal gibberellins
Gibberellins were extracted from the CF of SB5-1 following the protocol of Lee et al. (Citation1998). GAs were chromatographed on a 3.9×300 mm Bondapak, C18 column (Waters Corp., Milford, MA, USA), and eluted at 1.5 ml min−1 with the following gradient: 0–5 min, isocratic 28% MeOH in 1% aqueous acetic acid; 5–35 min, linear gradient from 28 to 86% MeOH; 35–36 min, 86–100% MeOH; 36–40 min, isocratic 100% MeOH. Forty-eight fractions of 1.5 ml each were collected. The fractions were then prepared for gas chromatograph/mass spectrometer (GC/MS) with selected ion monitoring (SIM) (6890N network GC system, and 5973 network mass selective detector; Agilent Technologies, Palo Alto, CA, USA). For each GA, 1 µl of sample was injected in a 30 m×0.25 mm i.d., 0.25-µm film thickness DB-1 capillary column (J & W Scientific Co., Folsom, CA, USA). The GC oven temperature was programmed for a 1-min hold at 60°C, then to rise at 15°C min−1 to 200°C followed by 5°C min−1 to 285°C. Helium carrier gas was maintained at a head pressure of 30 kPa. The GC was directly interfaced to a Mass Selective Detector with an interface and source temperature of 280°C, an ionizing voltage of 70 eV, and a dwell time of 100 ms. Full scan mode (the first trial) and three major ions of the supplemented [2H2] GAs internal standards (obtained from Prof. Lewis N. Mander, Australian National University, Canberra, Australia) and the fungal GAs were monitored simultaneously. The retention time was determined using hydrocarbon standards to calculate the Kovats Retention Index (KRI) value, while the GAs quantification was based on peak area ratios of nondeuterated (extracted) GAs to deuterated GAs.
Genomic DNA extraction and fungal identification
Genomic DNA isolation and PCR was performed according to established protocol (Khan et al. Citation2008). Universal primers for internal transcribed spacer (ITS) region (ITS1/4) as well as D1/D2 region primers of 28S rDNA, LR0R(F) (ACCCGCTGAACTTAAGC) (http://www.botany.duke.edu/fungi/mycolab/primers.htm) and TW13(R) (GGTCCGTGTTTCAAGACG) (Taylor and Bruns Citation1999) were used to identify the fungal isolate. The BLAST search program (http://www.ncbi.nlm.nih.gov/BLAST/) was used to look for nucleotide sequence homology. The sequences obtained were then aligned by ClustalW using MEGA version 4 software (Tamura et al. Citation2007), and the neighbor-joining (NJ) tree was generated using the same software. The bootstrap replications (1K) were used as a statistical support for the nodes in the phylogenetic tree.
Statistical analysis
The data were analyzed statistically for standard deviation by using MS-EXCEL. The mean values were compared, using Duncan's multiple range test (DMRT) at P<0.05 (ANOVA SAS release 9.1; SAS, Cary, NC, USA).
Results
Screening of endophytic fungi on Waito-C
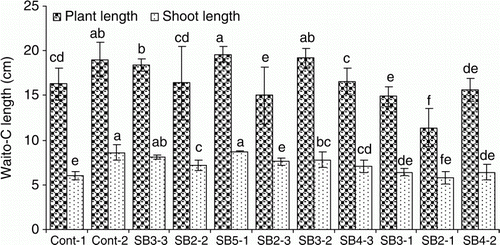
The CFs of nine endophytic fungi were screened for plant growth-promoting metabolites by applying them on Waito-C rice. Eight fungal isolates promoted shoot length of Waito-C rice while one inhibited it (). The fungal isolates SB5-1, SB3-2, and SB3-3 provided maximum shoot length as compared to control and were further investigated.
Figure 1. Screening of fungal isolates on Waito-C rice for their plant growth-promoting capacity. Cont-1 and Cont-2 stands for negative control (d·H2O) and positive control (G. fujikuroi), respectively. Data bars having a common letter(s) are not significantly different at the 5% level by DMRT. Error bars show standard deviations.
Bioassay of selected fungal isolates on soybean
The CFs of fungal isolates SB5-1, SB3-2, and SB3-3 were bioassyed on soybean cultivar Hwangkeumkong. The three fungal isolates promoted growth characteristics of soybean. However, maximum growth promotion was observed in plants treated with CF of SB5-1. The CF of SB5-1 markedly enhanced plant height (31.6 cm), shoot length (21.1 cm), whole plant fresh biomass (2.41 g), shoot fresh biomass (1.99 g), and leaf area (24.37 cm2) as compared to all other treatments (). The chlorophyll content per unit area was lower in SB5-1-treated plants. However, the overall chlorophyll contents may be higher due to higher leaf area in such treatments ().
Table 1. Influence of CFs of fungal isolates SB5-1, SB3-2, and SB3-3 on mean growth characteristics of soybean.
Analysis of CF of SB5-1 for gibberellins
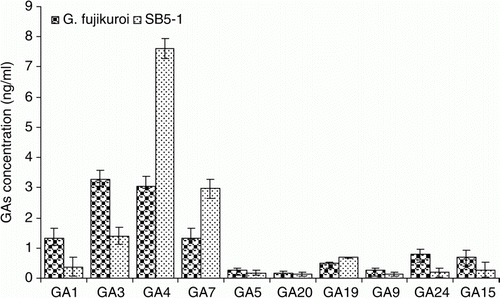
Gibberellins analysis of SB5-1 CF showed the presence of GA1 (0.15 ng/ml), GA3 (1.2 ng/ml), GA4 (7.37 ng/ml), GA5 (0.23 ng/ml), GA7 (3.18 ng/ml), GA9 (0.09 ng/ml), GA15 (0.46 ng/ml), GA19 (0.68 ng/ml), GA20 (0.19 ng/ml), and GA24 (0.09 ng/ml) in CF of SB5-1. Among them, GA1, GA3, GA4, and GA7 are physiologically active GAs. Fungal isolate SB5-1 produced significantly higher amounts of GA4 and GA7 than wild type G. fujikuroi during the current investigation ().
Figure 2. Production of various GAs by SB5-1 and G. fujikuroi. Of the four bioactive GAs, GA4 and GA7 were produced in higher amounts by SB5-1 compared to G. fujikuroi. Error bars show standard deviations.
Molecular and phylogenetic identification of SB5-1
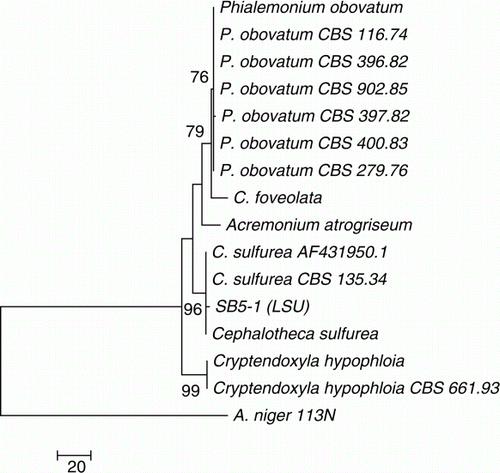
The phylogenetic analysis of fungal isolate SB5-1 was carried out by NJ and maximum parsimony (MP) methods. Consensus trees were constructed from 16 (15 references and 1 clone) aligned 28S rDNA sequences with 1000 bootstrap replications. These strains were selected through BLAST search showing maximum sequence homology percentage and query coverage, and lowest E values. Aspergillus niger was used as an out group. For D1/D2 region sequence of 28S rDNA, BLAST search showed that fungal isolate SB5-1 has 99% sequence homology with C. sulfurea. In the NJ dendrogram, fungal isolate SB5-1 formed a sub-clade (98% bootstrap support) with three strains of C. sulfurea (), while in MP dendrogram, fungal isolate SB5-1 formed a clade (96% bootstrap support) with the same strains of C. sulfurea (). On the basis of sequence homology and phylogenetic analysis, isolate SB5-1 was thus identified as a new strain of C. sulfurea. The 28S rDNA sequence was submitted to NCBI GenBank and was allotted accession number FJ808681.
Figure 3. Phylogenetic tree constructed by neighbor-joining method, using the 28S rDNA sequence (D1/D2 domains) of Cephalotheca sulfurea and related fungi. SB5-1 formed a sub-clade (98% bootstrap support) with three strains of C. sulfurea, which identify SB5-1 as a new strain of C. sulfurea.

Figure 4. Phylogenetic tree constructed by the maximum parsimony method, using the 28S rDNA sequence (D1/D2 domains) of Cephalotheca sulfurea and related fungi. SB5-1 formed sub-clade (96% bootstrap support%) with three strains of C. sulfurea, which identify SB5-1 as a new strain of C. sulfurea.
Discussion
Endophytism offers a new area of research based on the benefits of mutualistic interactions between host crops and nonpathogenic fungi. The advantages conferred by endophytic fungi include their ability to promote plant growth and tolerance to abiotic and biotic stresses. As such, the practical applications of endophytes as potential sources of bio-organic nutrients and as bio-control agents can significantly improve yields in an environment friendly manner (Diene and Narisawa Citation2009). Fungal endophytes are well known for their symbiotic relationship with their host plant, but information on their GAs production and plant growth-promotion capacity is still limited. We investigated GAs production for fungal endophytes isolated from soybean plant. The isolated fungi were screened for the presence of GAs, as screening of microbial CFs is an efficient technique for the identification of biologically active molecules (Higgs et al. Citation2001), and the fungal CFs provide a major source of novel secondary metabolites (Cragg et al. Citation1997). CFs obtained from pure fungal cultures were applied on Waito-C, to check their effects on plant growth, especially shoot length. An increase in shoot length provides a good idea about the presence of GAs, as these hormones are specialized for stem elongation (Khan et al. Citation2008). The usage of water-agar media in current experiment aided in determining the sole effect of CF on rice seedling growth, as water-agar media was devoid of nutrients. We selected Waito-C for the screening experiment, because Waito-C is a dwarf and GA-deficient rice cultivar, due to blocked C13 hydroxylation pathway, which leads to the formation of bioactive GA1 (Khan et al. Citation2008). The CFs of three fungal isolates, which provided maximum shoot length promotion of Waito-C, were bio-assayed on soybean. We were interested to investigate the effect of these selected fungi on soybean growth promotion as the fungal isolates were initially collected from the roots of soybean plant. It was observed that growth characteristics of soybean were promoted by such an application. For soybean bioassay experiment, soybean plants were grown in perlite growth medium and the pots were placed in a programmed growth chamber, to provide controlled environmental conditions for the plant growth. Our current experimental arrangements and results are in agreement with an earlier report on the subject (Khan et al. Citation2008).
Gibberellin analysis of the CF of SB5-1 showed the presence of 10 different GAs, which also included bioactive GA1, GA3, GA4, and GA7. Bioactive GA4 and GA7 production was higher in SB5-1 than in the wild type G. fujikuroi, which demonstrates the potential of our fungal isolate as plant growth promoter of its host plant. The CF of isolate SB5-1 was analyzed the presence of GAs through GC/MS SIM. GC/MS apparatus was used for quantification of GAs due to its unbiased character, as GC/MS analysis imparts interesting and unexpected new findings about a particular extract (Franck et al. Citation2005).
The fungal isolate SB5-1 was identified by sequencing highly conserved regions of its genomic DNA. ITS regions has gained immense importance in fungal identification in recent times (Lee et al. Citation2001; Sugita and Nishikawa Citation2003), although the ITS sequences sometimes fail to produce good phylogenetic comparisons at species level and thus make species-level identification difficult. However, the 5′ end of large-sub-unit (LSU) contains variable regions D1 and D2 of about 600 bps, which are important in the identification of closely related species (Wagner and Fischer Citation2002; Daniel and Meyer Citation2003). In the current study, ITS region failed to provide proper identification for our fungal isolate, while good sequence homology and phylogenetic tree was obtained for D1/D2 region of 28S rDNA gene. On the basis of sequence homology and phylogenetic analysis results, isolate SB5-1 was identified as a new strain of C. sulfurea.
The current study presents valuable information on the relationship of GAs production capacity of C. sulfurea AGH07 and its potential role and application in plant growth and development. Our studies indicate that C. sulfurea AGH07 is a novel Gas-producing fungus secreting high amounts in CF when compared with G. fujikuroi. As such, this fungal isolate may in future, be developed as a potential bio-fertilizer with minimal environmental risks. For future research, it is suggested that C. sulfurea AGH07 mycelia may be applied to the crops in the field, to give an insight on their usage with crop plants under field conditions, albeit additional studies on the identification and characterization of GAs encoding gene cluster, and optimization of Gas-producing media for this fungus need to be addressed.
Acknowledgements
This study was funded by Korea Research Foundation (KRF), Republic of Korea.
Additional information
Notes on contributors
Sumera Afzal Khan
M. Hamayun and S.A. Khan have contributed equally to the paperRelated Research Data
References
- Cragg , GM , Newman , DJ and Snader , KM . 1997 . Natural products in drug discovery and development . J Nat Prod. , 60 : 52 – 60 .
- Daniel , HM and Meyer , W . 2003 . Evaluation of ribosomal RNA and actin gene sequences for the identification of ascomyceteous yeasts . Int J Food Microbiol. , 86 : 61 – 78 .
- Diene , O and Narisawa , K . 2009 . The use of symbiotic fungal associations with crops in sustainable agriculture . J Dev Sus Agr. , 4 : 50 – 56 .
- Franck C , Lammertyn J , Nicolaï B. 2005 . Metabolic profiling using GC-MS to study biochemical changes during long-term storage of pears . In : F. Mencarelli and P. Tonutti , Proceedings of 5th international postharvest symposium . Acta Hort . 682 : 1991 – 1998 .
- Hamayun , M , Khan , SA , Khan , AL , Rehman , G , Kim , YH , Iqbal , I , Hussain , J , Sohn , EY and Lee , IJ . 2010a . Gibberellins production and plant growth promotion by pure cultures of Cladosporium sp. MH-6 isolated from Cucumber (Cucumis sativus L.) . Mycologia. , 102 : 989 – 995 .
- Hamayun , M , Khan , SA , Khan , AL , Tang , DS , Hussain , J , Ahmad , B , Anwar , Y and Lee , IJ . 2010b . Growth promotion of cucumber by pure cultures of gibberellin-producing Phoma sp. GAH7 . World J Microbiol Biotechnol. , 26 : 889 – 894 .
- Hasan , HAH . 2002 . Gibberellin and auxin production plant root fungi and their biosynthesis under salinity–calcium interaction . Rostlinná Výroba. , 48 : 101 – 106 .
- Higgs , RE , James , AZ , Jeffrey , DG and Matthew , DH . 2001 . Rapid method to estimate the presence of secondary metabolites in microbial extracts . Appl Environ Microbiol. , 67 : 371 – 376 .
- Hoagland DR , Arnon DI. 1950 . The water-culture method for growing plants without soil . Berkley : University of California Agricultural Experiment Station , Circ 347 .
- Kawaide , H . 2006 . Biochemical and molecular analysis of gibberellin biosynthesis in fungi . Biosci Biotechnol Biochem. , 70 : 583 – 590 .
- Khan , AL , Hamayun , M , Kim , HY , Kang , SM , Lee , JH and Lee , IJ . 2011 . Gibberellins producing endophytic Aspergillus fumigatus sp. LH02 influenced endogenous phytohormonal levels, isoflavonoids production and plant growth in salinity stress . Process Biochem. , 46 : 440 – 447 .
- Khan , SA , Hamayun , M , Yoon , HJ , Kim , HY , Suh , SJ , Hwang , SK , Kim , JM , Lee , IJ , Choo , YS Yoon , UH . 2008 . Plant growth promotion and Penicillium citrinum . BMC Microbiol. , 8 : 231
- Lee , IJ , Foster , K and Morgan , PW . 1998 . Photoperiod control of gibberellin levels and flowering in sorghum . Plant Physiol. , 116 : 1003 – 1011 .
- Lee , HG , Lee , JY and Lee , DH . 2001 . Cloning and characterization of the ribosomal RNA gene from Gonyaulax polyerdra . J Microbiol Biotechnol. , 11 : 515 – 523 .
- Márquez , LM , Redman , RS , Rodriguez , RJ and Roossinck , MJ . 2007 . A virus in a fungus in a plant—three way symbioses required for thermal tolerance . Science. , 315 : 513 – 515 .
- MacMillan , J . 2002 . Occurrence of gibberellins in vascular plants, fungi and bacteria . J Plant Growth Reg. , 20 : 387 – 442 .
- Martin GC. 1983 . Commercial uses of gibberellins Biochemistry and physiology of gibberellins In: Crozier , A . New York , NY : Praeger 395 – 444 .
- Nishijima , T , Koshioka , M , Yamazaki , H and Mander , LN . 1995 . Endogenous gibberellins and bolting in cultivars of Japanese radish . Acta Hort. , 394 : 199 – 206 .
- Ogas , J . 2000 . Gibberellins . Cur Biol. , 10 : R48
- Schulz , B and Boyle , C . 2005 . The endophytic continuum . Mycol Res. , 109 : 661 – 686 .
- Schulz , B , Römmert , AK , Damman , U , Aust , HJ and Strack , D . 1999 . The endophyte-host interaction: a balanced antagonism? . Mycol Res. , 103 : 1275 – 1283 .
- Sugita , T and Nishikawa , A . 2003 . Fungal identification method based on DNA sequence analysis. Reassessment of the methods of the pharmaceutical society of Japan and the Japanese pharmacopoeia . J Health Sci. , 49 : 531 – 533 .
- Suh , MK , Lim , JK , Lee , YH , Ha , GY , Kim , H , Kim , JR , Yaguchi , T and Nishimura , K . 2006 . Subcutaneous hyalohyphomycosis due to Cephalotheca foveolata in an immunocompetent host . Br J Dermatol. , 154 : 1184 – 1189 .
- Tamura , K , Dudley , J , Nei , M and Kumar , S . 2007 . MEGA4: molecular evolutionary genetics analysis (MEGA) software version 4.0 . Mol Biol Evol. , 24 : 1596 – 1599 .
- Taylor , DL and Bruns , TD . 1999 . Community structure of ectomycorrhizal fungi in a Pinus muricata forest: minimal overlap between the mature forest and resistant propagule communities . Mol Ecol. , 8 : 1837 – 1850 .
- Vandenbussche , F , Fierro , AC , Wiedemann , G , Reski , R and Van Der Straeten , D . 2007 . Evolutionary conservation of plant gibberellin signalling pathway components . BMC Plant Biol. , 7 : 65
- Wagner , T and Fischer , M . 2002 . Proceedings towards a natural classification of worldwide taxa Phellinus s.l. and Inonotus s.l., and phylogenetic relationships of allied genera . Mycologia. , 94 : 998 – 1016 .
- Waller , F , Achatz , B , Baltruschat , H , Fodor , J , Becker , K , Fischer , M , Heier , T , Huckelhoven , R , Neumann , C Von Wettstein , D . 2005 . The endophytic fungus Piriformis indica reprograms barley to salt-stress tolerance, disease resistance, and higher yield . Proc Natl Acad Sci. , 102 : 13386 – 13391 .
- Yamada , A , Ogura , T , Degawa , Y and Ohmasa , M . 2001 . Isolation of Tricholoma matsutake and T. bakamatsutake cultures from field-collected ectomycorrhizas . Mycoscience. , 42 : 43 – 50 .