Abstract
Invasion of poorly palatable grasses due to abandonment or improper grazing management decreases pastures feeding value and biodiversity. With the aim to control their spread, we assessed the relationship between sheep foraging behavior and changes in aboveground phytomass, leaf traits, and chemical features of the tall grass Brachypodium rupestre and evaluated the effects of a B. rupestre-based diet on epithelium keratinization of rumen. Our results demonstrated that sheep became less selective throughout the experimental trials and B. rupestre decreased its aboveground phytomass. Some leaf traits showed significant changes (LDMC, LA, and ADL were higher in ungrazed areas; leaf nitrogen content was higher in the grazed ones). In addition, we detected an increase of the degree of epithelium keratinization of sheep. Thus high grazing pressure can be used to control the spread of B. rupestre, but negative effects on animal welfare due to the increase of rumen keratinization might be expected.
1. Introduction
Throughout Europe, abandonment or improper management threaten calcareous grasslands (Riecken et al. Citation2002; Sebastià et al. Citation2008). Conservation of pasture biodiversity depends on key elements such as density of herbivores, type of management, and animal feeding behavior (Biondini et al. Citation1998; Adler et al. Citation2001; de Bello et al. Citation2006, Citation2007; Kahmen & Poschlod Citation2008; Catorci et al. Citation2011b, Citation2012a; Kiss et al. Citation2011). The decrease in herbage use due to pasture abandonment or improper management results in a taller and less digestible vegetation (Louault et al. Citation2002). Indeed, in such conditions dominant tall plants tend to spread (e.g. Poschlod et al. Citation1998; Grime Citation2001; Bonanomi & Allegrezza Citation2004), because of specific plant traits (i.e. extensive lateral clonal spread), litter deposition (Sydes & Grime Citation1981a, Citation1981b; Grubb et al. Citation1982; Campbell et al. Citation1992), and micro-scale influence on environmental features (Hurst & John Citation1999; Catorci et al. Citation2011a). The Brachypodium genus is one of the most widespread groups of dominant grasses in Europe and, within this group, Brachypodium rupestre (Host) Roem. et Schult., is the most common species in sub-Mediterranean pastures. It is a perennial grass (30–50 cm tall), with strong lateral clonal spread, semi-persistent green leaves, and high rate of litter deposition (Pignatti Citation1982). Because of its silica-rich and hairy leaves, it is also a poorly palatable plant (Roggero et al. Citation2002). In fact, in extensive farming systems, B. rupestre is disliked by sheep (Catorci et al. Citation2012b), especially if its tussocks form large patches. This promotes its invasive behavior, causing a change in species composition and a loss of biodiversity (Bonanomi et al. Citation2006, Citation2009; Catorci et al. Citation2011c). Thus, the control of the B. rupestre spread is a key issue for biodiversity conservation and the maintenance or restoration of pasture feeding value.
To reach these goals, the involvement of local stakeholders is a key tool. Thus, as sheep rearing was the major form of animal husbandry and today is still a key economic activity in Mediterranean mountains, we chose to use sheep for the experimental trial, though horses are more effective in controlling the spread of B. rupestre (Catorci et al. Citation2012a).
Sheep are able to select their diet from a large number of plant species and vegetation structures, which confronts them with a multitude of stimuli when deciding what to eat and what to avoid (Ginane & Dumont Citation2006). The dominance of unpalatable species may deter the use of patches (Arnold Citation1987), while the ingestion of poorly palatable species increases at high stocking rates (Kausrud et al. Citation2006). Indeed, herbivores foraging behavior and food items selection change depending on the intensity of grazing, namely the number of herbivores in relation to the availability of forage (Skarpe Citation1986; Heitschmidt et al. Citation1987).
Plants respond to herbivory by changing tissue chemistry, growth rate, morphology, and resources allocation (Belsky Citation1986; Bryant et al. Citation1992; Rooke & Bergström Citation2007), and by producing secondary metabolites and protective coatings (Alba et al. Citation2011). Moreover, grazing changes the plant ability to acquire resources, influencing the competitive interactions and thus the plant community composition (Louda et al. Citation1990) and the forage quantity/quality (Pavlů et al. Citation2006).
Changes in forage features, in turn, produce morphological and functional modifications at different levels of the digestive apparatus (Hofmann Citation1989; Scocco et al. Citation2011, Citation2012) in which rumen epithelial lining is a basic element. It is a stratified and keratinized flat epithelium, whose main functions are protection from exogenous mechanical stimuli (Dellmann & Eurell Citation1998; Dyce et al. Citation2002) and nourishment absorption. The outer keratinized layer of rumen epithelium creates a protective barrier, while the deeper layers are able to metabolize the volatile fatty acids produced from cellulose digestion (Dyce et al. Citation2002). Rumen epithelium works in a perfect balance between protective and absorptive functions (Bortolami et al. Citation2000), thus, when the degree of keratinization increases, the absorptive ability decreases (Hofmann Citation1989) and animal welfare may suffer (Malfatti et al. Citation2010). Moreover Ceccarelli et al. (Citation2009) demonstrated that during the pasture vegetative cycle, changes in forage agronomic features are related to significant morphometric modifications of rumen and of animal body state. Indeed low quality and/or quantity of forage directly alter the rumen absorptive surface area and cause a decrease in the absorption of volatile fatty acids (Hofmann et al. Citation1988; Hofmann Citation1989).
Because of this, it might be helpful to think in terms of ‘pasture-rumen interaction’ in deciding how pasture can be best managed and converted into sustainable profit (Dove Citation1996). Though the importance of using holistic approaches for planning pastoral systems is recognized, there is a dearth of data on the cumulative effect of grazing on plant aboveground phytomass and leaf traits of poorly palatable grasses, as well as on rumen features and foraging behavior of sheep. Considering this knowledge gap and with the goal of promoting forage quality and pasture biodiversity, we hypothesized that periodic high grazing pressure may prove useful in controlling the spread of B. rupestre if it does not lead to worsening of morpho-structural aspects of rumen.
It was demonstrated that extensive grazing promotes selective patch defoliation, which results in an uneven distribution of grazing pressure and in a patchwork in sward structure (Rook et al. Citation2004), that may promote the establishment of poorly palatable plants. Moreover, Catorci et al. (Citation2012b) argued that problems with B. rupestre invasion might be due to a combination of facts (i.e. livestock composition and facilitation processes between small thorny shrubs and juvenile individuals of B. rupestre) that broadens the effect of low disturbance intensity. Thus, we posited that fenced areas (useful in keeping disturbance intensity high) might help to overcome these management problems.
Considering these issues, we tested the following hypotheses. (1) Sheep item selection changes over the long run in overstocked systems. (2) Leaf removal affects the leaf traits of B. rupestre. (3) Leaf removal reduces the aboveground phytomass of B. rupestre and thus its dominant role in the plant community. (4) The degree of keratinization of the rumen epithelial lining is affected by a diet mainly composed of B. rupestre leaves.
2. Materials and methods
2.1. Study area
The study area, part of the ‘Torricchio Mountain Natural Reserve’ (central Italy; central coordinates 42°56′ N, 13°00′ E), is located at 900–1000 m a.s.l., on north-facing slightly steep slopes (5–15°), on limestone bedrock.
The vegetation consists of a mosaic of B. rupestre-dominated patches and of patches mostly composed of other grasses (e.g. Bromus erectus, Dactylis glomerata, Festuca sp. pl., and Koeleria lobata) and forbs (e.g. Salvia pratensis, Trifolium pratense, and Centaurea ambigua).
The mean annual rainfall is about 1200 mm, with a period of water scarcity during the summer. Winter cold stress is quite intense and lasts from December to February (Orsomando & Catorci Citation2000). In 2009 and 2010 (the years of experimentation), the annual rainfall was 1150 and 1400 mm year−1, while the summer rainfall was 200 and 310 mm, respectively (data gathered from the meteorological database of the ‘Torricchio Mountain Natural Reserve’).
Sheep traditionally graze pastures from June to October, but the number of grazing sheep has strongly decreased in recent decades.
2.2. Data collection
We conducted the research in a site of about 10 hectares covered by grasslands. The environmental features (altitude, slope aspect and angle, and soil depth) and management type (long-lasting grassland abandonment) were homogeneous throughout. The experimental design consisted of three phases, which are described below.
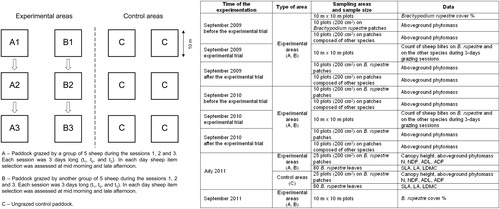
(1) To assess the foraging behavior of sheep, in September 2009 and 2010 the number of bites on B. rupestre and on the other species pool was collected in two groups of three paddocks (10 m×10 m each), while six other paddocks left ungrazed were used as control areas (see Appendix). In each experimental paddock, we visually estimated the B. rupestre cover (percent value) in September 2009 and again in September 2011 (before and after the experimental trials). In each group of experimental paddocks, five sheep were free to graze for three consecutive days (t1, t2, and t3), during which time they had no other food supply. This number of sheep was doubled than the optimal carrying capacity (2.5 sheep/plot×3 days) of the pasture (see Catorci et al. Citation2012b), so that it could be assumed as a high disturbance intensity.
Sheep item selection was assessed twice a day, at midmorning (tna) and late afternoon (tnb), for six sessions of observation per paddock (t1a, t1b, t2a, t2b, t3a, and t3b). During every session, we monitored each sheep three times, for five minutes each time, during which we recorded the number of bites on B. rupestre and on the other plant species. To evaluate the forage consumption during the experimental trials, inside each experimental paddock we randomly laid down 20 circular plots of 200 cm2 in patches entirely covered by B. rupestre and 20 in patches lacking B. rupestre. In 2009 and in 2010, we collected samples of the aboveground phytomass in half of these plots at the beginning of the three days of experimentation and in the second half of plots after its end (in the two years samples were collected from the same plots, which were marked by poles fixed in the ground at the plot center).
(2) To assess the cumulative effect on B. rupestre after the two consecutive years of foliage depletion (2009–2010), we collected samples of aboveground phytomass in patches entirely covered by B. rupestre in early July 2011 (i.e. in the phases of the growing period when this species was in bloom and had reached its complete development). Samples were collected in 50 randomly selected circular plots (200 cm2 each), 25 in the 6 paddocks grazed in 2009 and 2010, and 25 in the 6 control paddocks. All samples were oven-dried at 90°C to obtain dry matter (Whalley & Hardy Citation2000). These samples were used to evaluate whether the B. rupestre chemical features had changed as well. In the 200 cm2 plots, we sampled the sward canopy height (average value of five measurements for each plot) as well. To assess the effect of grazing on specific leaf area (SLA – the ratio of saturated leaf area to leaf dry mass), leaf area (LA – one-sided area of a fresh leaf), and leaf dry matter content (LDMC – the ratio of leaf dry mass to water-saturated fresh mass), 80 leaves of B. rupestre were randomly collected from both grazed and control paddocks. We used standardized procedures (Cornelissen et al. Citation2003) to preserve leaves.
(3) To test the effects of a B. rupestre-based diet on the degree of rumen keratinization, in September 2010 we selected 18 Comisana×Appenninica yearling sheep that had previously grazed together on grasslands where B. rupestre was almost absent. Six of them were killed at the beginning of the experiment; the remaining 12 were free to graze until the forage was totally consumed (15 days) within a paddock of 2500 m2 (50 m×50 m) established on the B. rupestre-dominated grassland. Inside the paddock the visually estimated cover of B. rupestre was of about 70%. Another group of six sheep was killed at the end of the experiment. Animals were slaughtered at the Visso (MC-Italy) slaughterhouse; samples from ruminal atrium and ventral sac wall were quickly removed, fixed in Bouin's fluid, and embedded in paraffin wax. In order to perform morphological evaluation and highlight changes in the degree of the rumen keratinization, tissue sections 5 µm thick were subjected to the following staining: hematoxylin/eosin, Mallory's trichromic, and Floxin B/Orange G/Alcian blue (Melis et al. Citation1992). To evaluate changes of the degree of keratinization of the rumen mucosa, we observed the sections under a light microscope (Eclipse E800, Nikon Corporation, Tokyo, Japan) and photographed them with a digital camera (Dxm 1200, Nikon Corporation). We used the images to measure the keratin layer. We directly measured the thickness of the epithelium and of the nucleated layers, while the thickness of the keratin layer was calculated as the difference. We converted these values into percentages with respect to the total epithelial thickness. For each animal, we randomly selected 10 fields of 0.35×0.27 mm and carried out five measurements in each of them. The images were processed by image analysis software (Lucia Measurement, Nikon Corporation).
2.3. Data analysis
We assessed the normality distribution and variance homogeneity of data using the Kolmogorov–Smirnov and Levene tests, respectively, to choose the appropriate statistical test (parametric/nonparametric). When data met the assumptions required for parametric tests, paired samples or independent t-tests were performed; when they did not, Wilcoxon or Mann–Whitney U-tests were run.
We calculated the mean cover of B. rupestre (±standard deviation) in the experimental paddocks before and after the two years of experimentation, and tested the significance of differences using the Wilcoxon test.
To describe the trend of food items selection, we calculated the percentage of bites on B. rupestre tussocks and on other plants for each sheep in each session of observation (t1a − 3b). To test whether there were significant differences in forage selection between 2009 and 2010, the mean percentages of bites of each sheep on B. rupestre tussock or on the patches composed of other plants at each session were compared using the paired samples t-test. We performed the same analyses to compare the bite percentages on B. rupestre tussock and on patches composed of other plants, at each session in 2009 and in 2010. To assess whether there was a significant correlation between forage selection and time, linear regression analyses were performed for both years, using time as the independent variable and bite percentage as dependent variable.
To assess the depletion of aboveground phytomass by sheep, we calculated the mean (±standard deviation) of dry matter of B. rupestre and of all the other plant species, collected in the 200 cm2 plots at the beginning of the first day (t1a) and at the end of the last day of grazing (t3b). The significance of differences was assessed using the Mann–Whitney U-test and Wilcoxon test.
To evaluate the effect of defoliation on B. rupestre tussocks and on the other species set, we used the formula TAP=A×S, where TAP is the total aboveground phytomass, A is the amount of dry matter per square meter (g m−2), and S is the surface area (m2). We calculated the TAP values for B. rupestre (TAP br ) and for the other species set (TAP os ). Finally, we calculated the percentage contribution of B. rupestre to the total aboveground phytomass of the experimental paddocks.
To ascertain whether there were significant differences in aboveground phytomass (dry weight) of B. rupestre between samples collected in plots of the experimental (grazed) and of the control (ungrazed) paddocks, we performed the independent t-test; to assess the differences in the sward canopy height we performed Mann–Whitney U-test. Mean percent values (±standard deviation) of dry weight and canopy height were calculated as well.
Detergent analysis was performed to determine neutral detergent fiber (NDF – hemicelluloses, cellulose, lignin, and silica ash), acid detergent fiber (ADF – lignin and silica ash), and acid detergent lignin (ADL) (Goering & Van Soest Citation1970). We calculated the total nitrogen content (N) starting from the crude protein values of the dry matter divided by the coefficient 6.25 (Bittante et al. Citation1990). LDMC, LA, and SLA were calculated for each sample recorded in the experimental and control areas following the procedures standardized by Cornelissen et al. (2003). To assess whether grazing on B. rupestre affected its leaf traits, the statistical significance of differences in NDF, ADF, ADL, N, LDMC, LA, and SLA between groups of sampling units collected in control and experimental paddocks (samples of aboveground phytomass and B. rupestre leaves) was tested using independent t-test or Mann–Whitney U-test. For each leaf trait, basic descriptive statistics (mean, standard deviation, minimum, maximum, and median) were calculated.
Mann–Whitney U-tests were run to assess statistically significant differences in keratinization degree in the rumen atrium and ventral sac, between sheep that, before having been killed, had grazed for 15 days in the 2500 m2 B. rupestre-dominated paddock and those that had not. Mean percent values (±standard deviation) of keratinization were calculated as well.
We used the SPSS 8.0 software to perform statistical analyses (version 8.0, SPSS Inc., Chicago, IL, USA).
3. Results
The average cover percentage of B. rupestre in the experimental paddocks was significantly higher (P=0.016) in 2009, before the start of the experimental activities (30.8±0.04) than in 2011, after the end of the experimental activities (25.0±0.03).
The item selection data highlighted differences throughout the experimental sessions of each year and between the two years. In 2009, the mean percentage of bites on B. rupestre increased throughout the experimental period, while that on the other species had the opposite trend (). The differences were statistically significant in t1a (P=0.000), which showed the greatest difference (90.2% vs. 9.8% of bites), t1b (P=0.000), and t2a (P=0.005). In 2010, the mean percentage of sheep bites on the patches composed by the two vegetation types remained almost constant during the experimental trial (), and this was consistent with the average cover value of patches composed by B. rupestre or other species. Differences between the percentage of bites on B. rupestre and on the other species remained statistically significant throughout most of the phases of the experimental period (t1a, P=0.017; t2a, P=0.014; t2b, P=0.003; t3a, P=0.018; t3b, P=0.002).
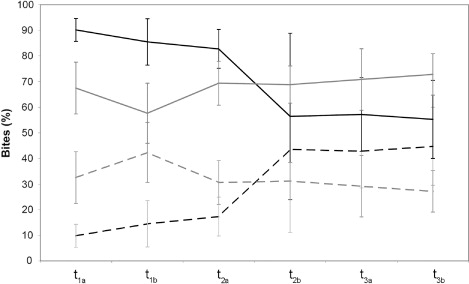
As regards the comparison of the percentage of bites on B. rupestre between the two years of experimentation, t-tests indicated significant differences in t1a (P=0.023), t1b (P=0.001), and t2a (P=0.026), while for the other species, differences were significant only in t1a (P=0.025) and t1b (P=0.013).
Linear regression analysis indicated significant correlations between bite percentage and time with regard to B. rupestre (R 2=0.385, P<0.001) and the other species (R 2=0.372, P<0.001) in 2009. No significant correlations were found in 2010.
Dry matter of B. rupestre and of other plant species decreased significantly (P<0.001) from t1a to t3b both in 2009 and in 2010 (). In 2010, B. rupestre dry matter was significantly lower than in 2009 in both sampling periods (P<0.001).
Table 1. Aboveground phytomass (g of dry matter [d.m.] m−2) of B. rupestre and of other species (± standard deviation) at the beginning of the first session (t1a) and at the end of the last session (t3b) of experimentation in 2009 and 2010.
Considering the surface covered by B. rupestre tussocks and by the patches of other species, the estimated average values of dry matter per paddock were of 1.8 kg and 1.4 kg, respectively, in 2009, and of 0.5 kg and 1.0 kg in 2010. The contribution of B. rupestre (as percentage of dry matter over the total dry matter characterizing the plots) was of about 56% in 2009 and 33% in 2010. The differences in dry matter production between the two years were −73% for the B. rupestre tussocks and −31% for the patches of other species.
As regards the cumulative effect of sheep grazing on B. rupestre, the experimental paddocks had lower values of aboveground dry matter and sward canopy height than control ones (48.8 g m−2±19.5 g m−2 vs. 62.2 g m−2±24.4 g m−2 of dry matter, P=0.04; 19.6 cm±4.2 cm vs. 31.0 cm±4.0 cm of canopy height, P<0.001).
The overview of mean, median, and ranges of leaf traits values in control and experimental paddocks (July 2011) is shown in . Mean values of LDMC and LA of B. rupestre were greater in the control paddocks than in the experimental ones. Differences were statistically significant (P<0.001). SLA did not show significant differences between control areas and experimental paddocks. The mean leaf nitrogen content was significantly higher in the grazed treatment (P<0.001). Instead, ADL values were significantly lower in the grazed treatment (P<0.001). Differences in NDF and ADF were not statistically recognizable.
Table 2. LDMC, LA, SLA, N, NDF, ADF, and ADL statistics.
The mean percentages of keratinization in the mucosa of the rumen atrium and of the ventral sac in sheep that had browsed for 15 days in high B. rupestre-covered grassland were 31.3%±2.3% in the atrium, and 35.1%±4.2% in the ventral sac. In sheep sacrificed before the beginning of the experiment, the degree of keratinization was of 17.3%±2.2% in the atrium and 20.2%±3.1% in the ventral sac. Percentages of keratinization in the two groups of sheep were significantly different (P<0.001).
4. Discussion
In accordance with previous studies (e.g. Biondini et al. Citation1998; Kausrud et al. Citation2006), we found that in 2009 sheep were less selective when food availability decreased. Indeed, at the beginning of the experimentation, the mean percent of bites on the leaves of B. rupestre was lower than that expected, considering the mean cover value of B. rupestre (about 30%, while the mean percentage of sheep bites was lower than 10%). After the first day, the number of bites on B. rupestre increased and the percentage of B. rupestre selection became higher than its average cover value. Change in item selection throughout the experimental sessions was also highlighted by linear regression analysis. The low values of the coefficient of determination are probably due to the small differences in item selection in t3a and t3b, when the availability of dry matter of B. rupestre and of other species was low and similar (as indicated by the lack of significant differences in t3a and t3b).
Despite the increase in percentage of bites on B. rupestre after t2a, in the last day of the experimentation, the frequency of bites on the other species was still high. This might be ascribed to the total consumption in the first phases of the highly palatable ‘other species,’ so that in the second half of the experimental trial, differences in the selection between the poorly palatable ones and B. rupestre were very small. We may also hypothesize that sheep were less selective because less satiated, due to the lower amount of aboveground phytomass in the second half of the experimentation.
Sheep foraging behavior changed dramatically in the second year. Indeed, the percentage of bites remained mostly similar to the species cover value (30% B. rupestre; 70% ‘other species’) throughout the experimental trial. Hence, as also indicated by linear regression analysis (no correlation was found between bites percentage and time), it is possible to argue that there was no patch selection even in t1a/t1b.
Differences between the two years seem to indicate that after the first year, the dislike for B. rupestre strongly decreased (as mainly highlighted by the different percentage of B. rupestre leaves selection that was less than 10% at the beginning of the first year and about 30% at the beginning of the second year). The significant differences recorded in the percent of bites on B. rupestre 2009 vs. B. rupestre 2010 in t1a, t1b, and t2a, supported this hypothesis. A number of factors may explain this change in foraging strategy. It was argued that dietary experience modulates feeding behavior and diet selection of sheep (Provenza Citation1995; Villalba & Provenza Citation2009). Thus, the use of the same group of animals in both years might partially explain the observed differences in the foraging behavior between 2009 and 2010. Nevertheless, we also detected changes in the leaf traits of B. rupestre, most of which could play a key role in the modification of the sheep foraging behavior. Indeed, as indicated by several researchers (e.g. Illius et al. Citation1987; Duncan et al. Citation1994; Crofts & Jefferson Citation1999), the increase in nitrogen content and the decrease in lignin content observed at the end of the second year of experimentation (as demonstrated by chemical analyses performed on B. rupestre leaves) seem to highlight a trend leading to the improvement of the B. rupestre palatability. Moreover, the absence of old and dead leaves (owing to the total consumption of the aboveground phytomass in the first year) as well as the decrease in canopy height may foster the selection of the B. rupestre-dominated patches (Mingo & Oesterheld Citation2009). The absence of senescent/dead material has also a high influence on digestibility of organic matter (Søegaard Citation2002).
With regard to the response of pasture to grazing, in 2010 a lower value of the aboveground phytomass was detected at the beginning of the experimentation both for B. rupestre tussocks and for the pool of the other species. Many studies have demonstrated that variations in annual rainfall drive the inter-annual changes in plant biomass production (Fay et al. Citation2002, Citation2003; Knapp et al. Citation2002; Nippert et al. Citation2006), especially in Mediterranean and sub-Mediterranean climates (Bugalho & Milne Citation2003). Nevertheless, in the study case, rainfall was greater in 2010 than it was in 2009, while aboveground phytomass decreased from 2009 to 2010, and therefore it is possible to state that inter-annual rainfall variations do not explain the observed changes in aboveground phytomass. Moreover, the contribution of B. rupestre (as percentage of dry matter over the total dry matter present in the paddocks) was of about 56% in 2009 and only of 33% in 2010. Thus, one can conclude that the effect of defoliation was greater on B. rupestre than on the other species pool. These results, as well as the comparison of dry matter between the six paddocks grazed in 2009/2010 and the six ungrazed control paddocks, indicate that it is possible to control the spread of B. rupestre by high disturbance intensity. It is a key goal because, as argued by Grime (2001) and demonstrated by Bonanomi et al. (Citation2006), the weakening of potential dominant species is a basic tool for grassland biodiversity conservation. Furthermore, the decrease of the B. rupestre aboveground phytomass, together with the observed changes in leaf traits, improves the pasture feeding value.
The unexpected lack of significant differences in the ADF and NDF values could be due to a number of factors. Bryant et al. (Citation1991) demonstrated that after severe defoliation, different species have different types of responses. Instead, Dannel et al. (Citation1994) stated that seasonal timing is a key factor in determining leaf chemical response after browsing, while Zheng et al. (Citation2010) demonstrated that the effect of grazing on leaf traits may change with the site condition (soil water and nutrient availability).
With regard to rumen response, we observed that the mucosa of both ruminal atrium and ventral sac wall showed a statistically significant increase in the degree of epithelium keratinization after the sheep grazed on paddock with high coverage of B. rupestre. This means that the intake of the B. rupestre leaves, rich in silica, hairs, and fibers, induces a defense response that fosters the defensive traits of rumen and probably lowers its absorptive ability (Hofmann Citation1989; Bortolami et al. Citation2000). Several studies demonstrated that changes in food quality, as in the case of forage feed value variations during the seasonal pasture vegetative cycle, may induce morphometric modifications of rumen absorptive surface and thus reduce its absorptive ability (e.g. Hofmann et al. Citation1988; Jiang et al. Citation2003; Ceccarelli et al. Citation2009). In this study, we demonstrated another type of rumen response to intake of poorly palatable grasses that contributes to improve our understanding of sheep foraging behavior. This knowledge also helps elucidate the defense mechanisms and strategies adopted by invasive grasses to avoid the herbivore bite.
5. Conclusions
Our findings indicated that sheep foraging with high stock density is useful for the depletion of the B. rupestre tussocks and improvement of forage quality. Indeed, B. rupestre strongly decreased its phytomass in only two years of grazing. As hypothesized by Endresz et al. (Citation2005), it may be due to the depletion of the reserves of perennating organs, and ultimately might lead to the individual's death. After the first year, the management of B. rupestre populations became less problematic, because sheep foraging behavior changed and the aboveground phytomass of the other species became higher than that of B. rupestre. This is consistent with Belsky (Citation1992), Mitchley (Citation1988), and Pavlů et al. (Citation2006), who demonstrated that the depletion of tall grasses owing to frequent defoliation results in an increase of species diversity.
The observed variation in leaf trait features and aboveground phytomass is in accordance with what would be expected of a stress-tolerant competitive species (Grime Citation2001), and seems to indicate that the plant defense mechanisms and strategies against herbivory tend to weaken when disturbance has high intensity and frequency. Moreover, assessment of the interaction between rumen and poorly palatable grass helps to understand the interplay of the different elements that compose a pastoral system and thus its response to management variation.
Actually, in accordance with Laca and Demment (Citation1996), we stated that grazers respond to changes in their resources by modifying their grazing strategy. The strategy modification affects, in turn, the plant community structure (by depletion of dominant species in the study case), in a feedback loop that also involves the morpho-structural aspects of rumen.
Moreover, our results indicated that in sub-Mediterranean pastoral systems, biodiversity conservation and forage quality improvement are not conflicting objectives, unlike in temperate regions, where the needs of farmers and of nature conservation are often in contrast (White et al. Citation2004; Pavlů et al. Citation2006). Nevertheless, the observed high and sudden increase in rumen keratinization has to be considered in pasture restoration planning, because animal welfare could suffer as consequence of the lower absorption of nutrients. In fact, Ceccarelli et al. (Citation2009) and Malfatti et al. (Citation2010) highlighted a negative relation between rumen keratinization degree and sheep body state. This may be a key factor in farming system management, because the proper balance between conserving biodiversity, protecting the livelihood of farmers, and safeguarding animal welfare is the central concern in governance of modern pastoralism (Ronchi & Nardone Citation2003; Hogan & Phillips Citation2008).
Acknowledgments
The Department for the Environment of the Marche Region and the Torricchio Natural Reserve funded this research; the grant was assigned to Prof. Andrea Catorci. The Authors would like to thank Mara Orsini for livestock management and friendly collaboration and Sheila Beatty for editing the English usage of the manuscript.
Related Research Data
References
- Adler, P, Raff, R, Lauenroth, WK. 2001. The effect of grazing on the spatial heterogeneity of vegetation. Oecologia. 128:465–479. 10.1007/s004420100737
- Alba, JM, Glas, JJ, Schimmel, BCJ, Kant, MR. 2011. Avoidance and suppression of plant defenses by herbivores and pathogens. J Plant Interact. 6:221–227. 10.1080/17429145.2010.551670
- Arnold, GW. 1987. Influence of the biomass, botanical composition and sward height of annual pastures on foraging behaviour by sheep. J Appl Ecol. 24:759–772. 10.2307/2403979
- Belsky, AJ. 1986. Does herbivory benefit plants? A review of the evidence. Am Nat. 127:870–892. 10.1086/284531
- Belsky, AJ. 1992. Effects of grazing, competition, disturbance and fire on species composition and diversity in grassland communities. J Veg Sci. 3:187–200. 10.2307/3235679
- Biondini, ME, Patton, BD, Nyren, PE. 1998. Grazing intensity and ecosystem processes in a northern mixed-grass prairie, USA. Ecol Appl. 8:469–479. 10.1890/1051-0761(1998)008[0469:GIAEPI]2.0.CO;2
- Bittante G, Andrighetto I, Ramanzin M. 1990. Fondamenti di zootecnica [Fundamentals of animal husbandry]. Torino: Liviana Ed. Italian.
- Bonanomi G, Allegrezza M. 2004. Effetti della colonizzazione di Brachypodium rupestre (Host) Roemer et Schultes sulla diversità di alcune fitocenosi erbacee dell'Appennino centrale [Effects of Brachypodium rupestre (Host) Roemer et Schultes colonisation on the diversity of some herbaceous coenoses of central Apennines]. Fitosociologia 41:51–69. Italian.
- Bonanomi, G, Caporaso, S, Allegrezza, M. 2006. Short-term effects of nitrogen enrichment, litter removal and cutting on Mediterranean grassland. Acta Oecol. 30:419–425. 10.1016/j.actao.2006.06.007
- Bonanomi, G, Caporaso, S, Allegrezza, M. 2009. Effects of nitrogen enrichment, plant litter removal and cutting on species rich Mediterranean calcareous grassland. Plant Biosyst. 143:443–455. 10.1080/11263500903172128
- Bortolami R, Callegari E, Beghelli V. 2000. Anatomia e Fisiologia degli animali domestici [Anatomy and physiology of domestic animals]. Bologna: Calderini Edagricole Ed. Italian.
- Bryant, JP, Heitkonig, I, Kuropat, PJ, Owen-Smith, N. 1991. Effect of severe defoliation on the long-term resistance to insect attack and leaf chemistry in six woody species of the southern African savanna. Am Nat. 137:50–63. 10.1086/285145
- Bryant, JPB, Reichardt, PB, Clausen, TP. 1992. Chemically mediated interactions between woody plants and browsing mammals. J Rangel Manag. 45:18–24. 10.2307/4002520
- Bugalho, MN, Milne, JA. 2003. The composition of the diet of red deer (Cervus elaphus) in a mediterranean environment: a case of summer nutritional constraint?. For Ecol Manag. 181:23–29. 10.1016/S0378-1127(03)00125-7
- Campbell, BD, Grime, JP, Mackey, JML. 1992. Shoot thrust and its role in plant competition. J Ecol. 80:633–641. 10.2307/2260855
- Catorci, A, Cesaretti, S, Gatti, R, Ottaviani, G. 2011a. Abiotic and biotic changes due to spread of Brachypodium genuense (DC.) Roem. & Schult. in sub-Mediterranean meadows. Community Ecol. 12:117–125. 10.1556/ComEc.12.2011.1.14
- Catorci, A, Gatti, R, Cesaretti, S. 2012a. Effect of sheep and horse grazing on species and functional composition of sub-Mediterranean grasslands. Appl Veg Sci. 15:459–469. 10.1111/j.1654-109X.2012.01197.x
- Catorci, A, Ottaviani, G, Ballelli, S, Cesaretti, S. 2011b. Functional differentiation of central apennine grasslands under mowing and grazing disturbance regimes. Pol J Ecol. 59:115–128.
- Catorci, A, Ottaviani, G, Cesaretti, S. 2011c. Functional and coenological changes under different long-term management conditions in Apennine meadows (central Italy). Phytocoenologia. 41:45–58. 10.1127/0340-269X/2011/0041-0481
- Catorci, A, Ottaviani, G, Vitasović Kosić, I, Cesaretti, S. 2012b. Effect of spatial and temporal patterns of stress and disturbance intensities in a sub-Mediterranean grassland. Plant Biosyst. 146:352–367.
- Ceccarelli, P, Scocco, P, Malfatti, A, Vitali, F, Catorci, A. 2009. Changes of sheep ruminal mucosae related to seasonal plant growth: when the anatomy is involved in the management of pastoral systems [Abstract]. Ital J Anat Embryol. 114:72.
- Cornelissen, JHC, Lavorel, S, Garnier, E, Díaz, S, Buchmann, N, Gurvich, DE, Reich, PB, ter Steege, H, Morgan, HD, van der Heijden, MGA. 2003. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust J Bot. 51:335–380. 10.1071/BT02124
- Crofts, A, Jefferson, RG. 1999. The Lowland grassland management handbook. Peterborough (UK): English Nature–The Wildlife Trusts.
- Dannel, K, Bergstrom, R, Edenius, L. 1994. Effects of large mammalian browsers on architecture, biomass and nutrient of woody plants. J Mammal. 75:833–844. 10.2307/1382465
- de Bello, F, Lepš, J, Sebastià, MT. 2006. Variations in species and functional plant diversity along climatic and grazing gradients. Ecography. 29:801–810. 10.1111/j.2006.0906-7590.04683.x
- de Bello, F, Lepš, J, Sebastià, MT. 2007. Grazing effect on species and functional diversity along a climatic gradient. J Veg Sci. 18:25–34.
- Dellmann, HD, Eurell, JA. 1998. Textbook of veterinary histology. 4th ed. Baltimore (MD): William and Wilkins Eds.
- Dove, H. 1996. The Ruminant, the rumen and the pasture resource: nutrient interactions in the grazing animals. In: Hodgson J, Illius AW, editors. The ecology and management of grazing systems. Wallingford (UK): CAB International; p. 219–246.
- Duncan, AJ, Hartley, SE, Iason, GR. 1994. Fine-scale discrimination of forage quality by sheep offered a soyabean meal or barley supplement while grazing a nitrogen-fertilized heather (Calluna vulgaris) mosaic. J Agric Sci. 123:363–370. 10.1017/S0021859600070374
- Dyce, KM, Sack, WO, Wensing, CJG. 2002. Textbook of veterinary anatomy. 3rd ed. Philadelphia (PA): WB Saunders.
- Endresz, G, Zöld-Balogh, Á, Kalapos, T. 2005. Local distribution pattern of Brachypodium pinnatum (Poaceae) – field experiments in xeric loess grassland in N. Hungary. Phyton. 45:249–265.
- Fay, PA, Carlisle, JD, Danner, BN, Lett, MS, McCarron, JK, Stewart, C, Knapp, AK, Blair, JM, Collins, SL. 2002. Altered rainfall patterns, gas exchange, and growth in grasses and forbs. Int J Plant Sci. 163:549–557. 10.1086/339718
- Fay, PA, Carlisle, JD, Knapp, AK, Blair, JM, Collins, SL. 2003. Productivity responses to altered rainfall patterns in a C4-dominated grassland. Oecologia. 137:245–251. 10.1007/s00442-003-1331-3
- Ginane, C, Dumont, B. 2006. Generalization of conditioned food aversions in grazing sheep and its implications for food categorization. Behav Process. 73:178–186. 10.1016/j.beproc.2006.05.006
- Goering HK, Van Soest PJ. 1970. Forage fiber analysis (apparatus, reagents, procedures, and some applications). ARS/USDA Agr. Handbook no. 379. Washington: US Government Printing Office.
- Grime, JP. 2001. Plant strategies, vegetation processes and ecosystems properties. Chichester (UK): Wiley.
- Grubb, PJ, Kelly, D, Mitchley, J. 1982. The control of relative abundance in communities of herbaceous plants. In: Newman E, editor. The plant community as a working mechanism. Oxford (UK): Blackwell. p. 79–97.
- Heitschmidt, RK, Dowhower, SL, Walker, JW. 1987. Some effects of a rotational grazing treatment on quantity and quality of available forage and amount of ground litter. J Rangel Manag. 40:318–321. 10.2307/3898728
- Hofmann, RR. 1989. Evolutionary steps of ecophysiological adaptation and diversification of ruminants: a comparative view of their digestive system. Oecologia. 78:443–457. 10.1007/BF00378733
- Hofmann, RR, Kock, RA, Ludwig, J, Axmacher, H. 1988. Seasonal changes in rumen papillary development and body condition in free ranging Chinese water deer (Hydropotes inermis). J Zool. 216:103–117. 10.1111/j.1469-7998.1988.tb02418.x
- Hogan, JP, Phillips, CJC. 2008. Nutrition and welfare of ruminants. Annu Rev Biomed Sci. 10:33–50. 10.5016/1806-8774.2008.v10pT33
- Hurst, A, John, E. 1999. The biotic and abiotic changes associated with Brachypodium pinnatum dominance in chalk grassland in south-east England. Biol Conserv. 88:75–84. 10.1016/S0006-3207(98)00089-5
- Illius, AW, Wood-Bush, DGM, Eddison, JC. 1987. A study of the foraging behaviour of cattle grazing patchy swards. Biol Behav. 12:33–44.
- Jiang, Z, Takatsuki, S, Wang, W, Li, J, Jin, K, Gao, Z. 2003. Seasonal changes in parotid and rumen papillary development in Mongolian gazelle (Procapra gutturosa Pallas). Ecol Res. 18:65–72. 10.1046/j.1440-1703.2003.00533.x
- Kahmen, S, Poschlod, P. 2008. Effects of grassland management on plant functional trait composition. Agric Ecosyst Environ. 128:137–145. 10.1016/j.agee.2008.05.016
- Kausrud, K, Mysterud, A, Rekdal, Y, Holand, Ø, Austrhein, G. 2006. Density-dependent foraging behaviour of sheep on alpine pastures: effects of scale. J Zool. 270:63–71.
- Kiss, T, Lévai, P, Ferencz, Á, Szentes, S, Hufnagel, L, Nagy, A, Balogh, Á, Pinter, O, Saláta, D, Házi, J, et al. 2011. Change of composition and diversity of species and grassland management between different grazing intensity in pannonian dry and wet grasslands. Appl Ecol Environ Res. 9:197–230.
- Knapp, AK, Fay, PA, Blair, JM, Collins, SL, Smith, MD, Carlisle, JD, Harper, CW, Danner, BT, Lett, MS, McCarron, JK. 2002. Rainfall variability, carbon cycling, and plant species diversity in a mesic grassland. Science. 298:2202–2205. 10.1126/science.1076347
- Laca, EA, Demment, MW. 1996. Foraging strategies of grazing animals. In: Hodgson J, Illius AW, editors. The ecology and management of grazing systems. Wallingford (UK): CAB International; p. 137–158.
- Louault, F, Soussana, JF, Perrodin, M. 2002. Long-term effects of a reduced herbage use in a semi-natural grassland. I. Plant functional traits and plant response groups. Grassl Sci Eur. 7:338–339.
- Louda, SM, Keeler, KH, Holt, RD. 1990. Herbivore influences on plant performance and competitive interactions. In: Grace J, Tilman D, editors. Perspectives in plant competition. San Diego (CA): Academic Press. p. 414–444.
- Malfatti, A, Scocco, P, Belardinelli, C, Gatti, R, Mercati, F, Dall'Aglio, C, Catorci, A. 2010. Rumen keratinization and BCS in sheep grazing in badland for wood fire prevention. Large Anim Rev. 5:88.
- Melis M, Carpino F, Di Tondo U. 1992. Tecniche in anatomia patologica [Techniques in pathological anatomy]. Milan: Edi-Ermes. Italian.
- Mingo, A, Oesterheld, M. 2009. Retention of dead leaves by grasses as a defence against herbivores. A test on the palatable grass Paspalum dilatatum. Oikos. 118:753–757. 10.1111/j.1600-0706.2008.17293.x
- Mitchley, J. 1988. Control of relative abundance of perennials in chalk grassland in southern England. II. Vertical canopy structure. J Ecol. 76:341–350. 10.2307/2260597
- Nippert, JB, Knapp, AK, Briggs, JM. 2006. Intra-annual rainfall variability and grassland productivity: can the past predict the future?. Plant Ecol. 184:65–74. 10.1007/s11258-005-9052-9
- Orsomando, E, Catorci, A. 2000. The phytoclimate of Umbria. Parlatorea. 6:5–24.
- Pavlů, V, Hejcman, M, Pavlů, L, Gaisler, J, Nežerková,, P. 2006. Effect of continuous grazing on forage quality, quantity and animal performance. Agric Ecosyst Environ. 113:349–355. 10.1016/j.agee.2005.10.010
- Pignatti, S. 1982. Flora d'Italia [Flore of Italy]. I-III. Bologna: Edagricole. Italian.
- Poschlod, P, Kiefer, S, Tränkle, U, Fischer, SF, Bonn, S. 1998. Plant species richness in calcareous grasslands as affected by dispersability in space and time. Appl Veg Sci. 1:75–90. 10.2307/1479087
- Provenza FD. 1995. Role of learning in food preferences of ruminants: Greenhalgh and Reid revisited. In: von Engelhardt W. Ruminant physiology: digestion, metabolism, growth and reproduction. Proceedings of the eighth International Symposium on Ruminant Physiology; 1994 September25–30; Willingen, Hesse. Stuttgard: Ferdinand Enke.
- Riecken U, Finck P, Schroder E. 2002. Significance of pasture landscape for nature conservation and extensive agriculture. In: Redecker B, Finck P, Hardtle W, Riecken U, Schroder E, editors. Pasture landscape and nature conservation. Berlin, Heidelberg: Springer, Verlag; 423–435.
- Roggero S, Bagella PP, Farina R. 2002. Un archivio dati di Indici specifici per la valutazione integrata del valore pastorale [A database of specific indices for the integrated assessment of grazing value]. Riv Agron. 36:149–156. Italian.
- Ronchi, B, Nardone, A. 2003. Contribution of organic farming to increase sustainability of Mediterranean small ruminant livestock systems. Livest Prod Sci. 80:17–31. 10.1016/S0301-6226(02)00316-0
- Rook, AJ, Dumont, B, Isselstein, J, Osoro, K, Wallis De Vries, MF, Parente, G, Mills, J. 2004. Matching type of livestock to desired biodiversity outcomes in pastures – a review. Biol Conserv. 119:137–150. 10.1016/j.biocon.2003.11.010
- Rooke, T, Bergström, R. 2007. Growth, chemical responses and herbivory after simulated leaf browsing in Combretum apiculatum. Plant Ecol. 189:201–212. 10.1007/s11258-006-9177-5
- Scocco, P, Aralla, M, Catorci, A, Belardinelli, C, Arrighi, S. 2011. Immunodetection of aquaporin 5 in sheep salivary glands related to pasture vegetative cycle. Folia Histochem Cytobiol. 49:458–464. 10.5603/FHC.2011.0065
- Scocco, P, Brusaferro, A, Catorci, A. 2012. Comparison between two different methods for evaluating rumen papillae measures related to different diets. Microsc Res Tech. 75:884–889. 10.1002/jemt.22008
- Sebastià, MT, de Bello, F, Puig, L, Taull, M. 2008. Grazing as a factor structuring grasslands in the Pyrenees. Appl Veg Sci. 11:215–222. 10.3170/2008-7-18358
- Skarpe, C. 1986. Plant community structure in relation to grazing and environmental changes along a north-south transect in the western Kalahari. Vegetatio. 68:3–18.
- Søegaard, K. 2002. The effect of grazing management on botanical composition and herbage quality on field level. Grassl Sci Eur. 7:162–163.
- Sydes, C, Grime, JP. 1981a. Effects of tree leaf litter on herbaceous vegetation in deciduous woodland. I Field investigations. J Ecol. 69:237–248. 10.2307/2259828
- Sydes, C, Grime, JP. 1981b. Effects of tree leaf litter on herbaceous vegetation in deciduous woodland. II An experimental investigation. J Ecol. 69:249–262. 10.2307/2259829
- Villalba, JJ, Provenza, FD. 2009. Learning and dietary choice in herbivores. Rangel Ecol Manag. 62:399–406. 10.2111/08-076.1
- Whalley, RDB, Hardy, MB. 2000. Measuring botanical composition of grasslands. In: Mannetje L.'t, Jones RM, editors. Field and laboratory methods for grassland and animal production research, 67–102. Wallingford (UK): CAB International.
- White, TA, Barker, DJ, Moore, KJ. 2004. Vegetation diversity, growth, quality and decomposition in managed grasslands. Agric Ecosyst Environ. 80:213–226.
- Zheng, SX, Ren, HY, Lan, ZC, Li, WH, Wang, KB, Bai, YF. 2010. Effects of grazing on leaf traits and ecosystem functioning in Inner Mongolia grasslands: scaling from species to community. Biogeosciences. 7:1117–1132. 10.5194/bg-7-1117-2010
Appendix: Explanation of the experimental design