
ABSTRACT
Due to their ability to shuttle proteins, lipids and genetic material between distant cells, exosomes promote extensive phenotypic changes in recipient cells, modulating immune responses, cellular migration, cancer metastasis or the spreading of neurotoxic protein aggregates in neurodegenerative diseases. Besides intercellular communication, exosome biogenesis and secretion permit the rapid release of a selective repertoire of compounds, conferring cells with an additional mechanism to fight alterations in protein, lipid or RNA homeostasis during stress or pathological conditions. Here, we review the dual role of the different quality control mechanisms arising from the endolysosomal system and the diverse situations that control the decision between degradation or secretion. The crosstalk between exosome secretion and the different cellular degradation mechanisms confers an additional layer of protection to maintain cellular integrity and homeostasis in a number of physiological and pathological conditions.
Introduction
Throughout their entire lifetime, cells are exposed to different harmful conditions, including thermal, physical, chemical, and oxidative stresses, altogether contributing to molecular damage. Cells have evolved various and interconnected organelle-specific quality control mechanisms to recognize and respond to this damage at different subcellular locations of the cell, including the cytosol, the endoplasmic reticulum (ER), the nucleus, mitochondria and the plasma membrane.Citation1-3 A complex network of chaperones recognizes unfolded proteins and supports their refolding. However, when refolding fails, chaperones assist in protein degradation via the ubiquitin-proteasome (UPS) or autophagy-lysosomal pathways.Citation4 Notably, under some circumstances or when load of proteins destined for degradation saturates the capacity of the proteolytic systems, cells can defend themselves against proteotoxicity through the release of toxic protein products to the extracellular media associated or not to lipid vesicles or involving other types of unconventional secretion.Citation5-9
Exosomes are small lipid vesicles that are secreted to the extracellular environment upon the fusion of endosomal compartments with the plasma membrane.Citation10 Once in the extracellular milieu, exosomes can be taken up by nearby cells or, after reaching the blood stream, they can be taken up by distant cells, modulating the activity and fate of receptor cells.Citation11 Hence, exosomes are currently being recognized as important mediators for cell-to-cell communication in many physiologicalCitation12 and pathological situations, including immune response, cancer progression and metastasis, neuronal communication, cardiovascular diseases and progression of neurodegenerative diseases.Citation13-20
During the last years, a lot of effort has been placed trying to elucidate the role of exosomes as vehicles for cell-to-cell communication. However, less attention has been placed on the impact that the biogenesis and secretion of exosomes has in producing cells. Exosomes may participate in the control of cellular homeostasis by promoting the release of intracellular harmful components, including proteins, lipids or nucleic acids. In this review, we emphasize the role of exosomes as a quality control mechanism that maintains intracellular homeostasis by promoting the selective release of intracellular harmful components. Clearance of damaged or toxic material, including proteins, lipids and even nucleic acids through exosomes, might alleviate intracellular stress and contribute to the preservation of cellular homeostasis.Citation21
Here, we outline the evidence that selective incorporation and release of cellular compounds in exosomes is a quality-control strategy to alleviate intracellular stress and preserve cellular homeostasis. We highlight our current understanding on the physiological functions of exosomes and the endolysosomal system, and how they interact to preserve cellular homeostasis.
The endolysosomal pathway in the control of cellular homeostasis
The endolysosomal system is a highly dynamic compartment in which different membrane compartments modulate intracellular protein and lipid trafficking through regulated processes of internalization, sorting, recycling, degradation or secretion. Endocytosis allows the internalization of adhesion receptors, growth-factor receptor–ligand complexes, nutrient transporters, lipids, extracellular material and pathogens. Vesicles formed from the plasma membrane fuse and deliver their membrane and protein content to Rab5- and EEA1-positive early endosomes, which later undergo conversion from Rab5- to Rab7-positive endosomes.Citation22 During this conversion, a significant amount of the internalized content is recycled back to the plasma membrane through Rab11-positive recycling endosomes, while the remaining material is sequestered in intraluminal vesicles (ILVs) in late endosomes, also known as multivesicular bodies (MVBs).Citation23 Ubiquitinated membrane proteins are recruited to endosomes by the ESCRT machinery. ESCRT-0, ESCRT-I and ESCRT-II directly bind to ubiquitinated membrane proteins, while ESCRT-III and the ATPase Vps4 drive membrane-remodeling reactions that result in ILV invagination and scission. ILV biogenesis can also occur through ESCRT-independent mechanisms.Citation24 Tetraspanin proteins, such as CD63 and CD81, are proposed to act as regulators of ILVs formation.Citation25-28 The accumulation of ceramide in the endosomal membrane is also relevant for the formation of ILV.Citation29 The effect of ceramide on ILV formation has been proposed to be mediated by the local production of its downstream metabolite sphingosine-1-phosphate (S1P).Citation30 Recently, it has been described that not only membrane proteins but also cytoplasmic proteins can be selectively packaged into ILVs through a process called endosomal microautophagy (MA). MA depends on the ESCRT machinery and the electrostatic binding of Hsc70 to endosomal acidic phospholipids. Hsc70 interacts and introduce into ILVs cytoplasmic proteins containing KFERQ-motifs.Citation31
Once ILV are formed, MVBs can degrade their cargo by fusing with lysosomes or, alternatively, MVBs can secrete their ILVs by fusing with the plasma membrane, eliminating the incorporated components through secretion to the extracellular environment. The molecular mechanisms and cellular situations that regulate the fate of MVBs are not completely understood. MVB are decorated with tethering complexes (CORVET, HOPS) and SNAREs (e.g. Stx7, Stx8, VTI1b and Vamp7) that facilitate the fusion with lysosomes or with the plasma membrane.Citation32 Hence, 2 quality-control mechanisms converge at the endolysosomal compartment for dealing with damaged, unwanted or toxic intracellular components. It can mediate the degradation and recycling of damaged or toxic intracellular components through lysosomal degradation, or allow the sequestration and release of these compounds in exosomes.
Role of exosomes in the preservation of intracellular homeostasis
Exosomes were first described in the 80s as a cellular mechanism by which reticulocytes get rid of the transferrin receptor (TfR) during their maturation into erythrocytes. TfR molecules were found in small vesicles inside endosomal compartments, which were released to the extracellular medium upon exocytosis of these endosomes, supporting the role of exosomes as vehicles to eliminate unwanted cellular compounds.Citation33,34 The protein composition of exosomes supports the function of exosomes as a cellular mechanism to get rid of obsolete, and toxic material. Misfolded and prion proteins are released in exosomesCitation35 and have been involved in the propagation of neurodegenerative diseases, such as Huntington disease, Alzheimer disease, and Parkinson disease. Several self-aggregating neurotoxic proteins including amyloid β, APP C-terminal fragments, Tau, α-synuclein, SOD1 and the prion protein (PrP) can be released from cells in exosomes.Citation36-41 These findings delineate a protein quality control pathway that, unlike degradation-based mechanisms, promotes protein homeostasis by exporting misfolded proteins through exosomal route.
The extracellular release of RNAs or microRNAs in exosomes may be a rapid way to regulate gene expression during cellular activation or transformation. For example, during an induced muscle atrophy process, myotubes secrete miR-23 and miR-182 to the extracellular environment through exosomes. miR-182 represses FoxO expression and its extracellular release in exosomes allows FoxO expression and the transcriptional program required for the acquisition of atrophy phenotype.Citation42,43 Similarly, upon activation, lymphocytes down-modulate the intracellular levels of miR-150 , a key repressor of lymphocyte differentiation and function, by releasing it in exosomes.Citation44 let-7 miRNAs generally play a tumor-suppressive role targeting oncogenes such as RAS or HMGA2. It has been proposed that cancer cells release let-7 miRNAs via exosomes into the extracellular environment to maintain their oncogenesis.Citation45 Inhibition of exosome secretion by silencing Rab27 proteins leads to impaired microRNA release and increased miRNA activity in the parental cell.Citation46 Interestingly, exosomal miRNA secretion is a mechanism whereby cells rapidly dispose miRNAs in excess of their targets to adjust miRNA:mRNA ratio. Physiological (cell-activation dependent) or artificial overexpression of miRNA target sequences promotes a relative miRNA enrichment in P-bodies and depletion from MVBs and exosomes. Conversely, artificial overexpression of a miRNA enriches it in MVBs and exosomes.Citation47
Exosomes participate in the regulation of intracellular RNA homeostasis by promoting the release of misfolded or degraded RNA products. yRNAs are involved in the degradation of structured and misfolded RNAs. Recent analysis of the RNA content of exosomes by deep-sequencing techniques has shown a remarkable enrichment of yRNA fragments and mRNA degradation products in exosomes.Citation48 Interestingly, exosomes may be also involved in the release of toxic RNAs since expanded trinucleotide repeat RNAs, such as the CAG repeats that underlie RNA toxicity in Huntington disease, are released from the cell in exosomes.Citation49 Other evidences support the relation between RNA degradation and its export to extracellular vesicles. In this regard, proteins involved in RNA processing are abundant in exosomes, and secreted RNAs have almost twice shorter half-life times than intracellular mRNAs.Citation50,51 Altogether, all these reports support that cells maintain intracellular RNA homeostasis through the release of distinct RNA species in extracellular vesicles.
Although less known, exosomes are involved in the control of lipid homeostasis. Exosomes have been shown to alleviate cholesterol accumulation in Niemann-Pick type C disease, a lysosomal storage disease in which cells accumulate unesterified cholesterol and sphingolipids within the endosomal and lysosomal compartment.Citation52 Antipsychotics drugs carry serious side effects such as the disruption of lipid homeostasis. Antipsychotics induces intracellular accumulation of LDL and impair intracellular cholesterol trafficking. When curcumin is administrated to cells under the effects of antipsychotics drugs, LDL is released into exosomes improving lipid homeostasis.Citation53
Genomic and mitochondrial DNA have been found in exosomes. Evidence shows that genomic DNA is secreted in exosomes by cancer cells. One of the fragments of double stranded genomic DNA (dsDNA) secreted correspond to mutations of the suppressor gene p53.Citation54 Other works also showed that exosomes released from tumor cells lines contain high levels of ssDNA. Mitochondrial DNA (mtDNA) has been found in glioblastoma and astrocyte exosomes.Citation55 Telomeric repeat-containing RNA (TERRA) is release into exosomes by cells that present telomere dysfunction.Citation56,57 Although several works have reported the presence of DNA in exosomes, the function of exosomes in the regulation of DNA homeostasis is far from been elucidated.
Degradation vs secretion to preserve intracellular homeostasis
Whereas extracellular components and plasma membrane receptors are transported to the degradation/secretion pathway by the endosomal/exosomal pathway, intracellular components are transported to the lysosome by the process of autophagy, a ‘self-eating’ catabolic pathway that is used by cells to capture their own cytoplasmic components destined for degradation and recycling.Citation58 Autophagy can handle degradation not only of cytosolic macromolecules, but also of much larger structures such as excess or dysfunctional organelles, protein aggregates and intracellular pathogens.
Autophagy begins when double-membraned structures called phagophores engulf portions of cytosol that can include aggregates, lipids, carbohydrates, damaged organelles or invading pathogens. The phagophore expands and grows into a double-membrane compartment, known as the autophagosome.Citation59 Autophagosomes must undergo a series of maturation steps in part by fusing with endocytic vesicles, including early and late endosomes and MVBs.Citation60,61 The resulting hybrid organelles, called amphisomes, are more acidic and fuse with lysosomes to form degradative autolysosomes. Proper maturation of the autophagosome requires an intact endocytic trafficking pathway, components of the endosomal sorting complex required for transport (ESCRT) pathway,Citation62-64 and components involved in endocytic vesicle fusion.Citation65,66 Autophagosomes traffic along microtubules toward the microtubule-organizing center, where lysosomes are concentrated, enabling fusion and degradation of the contents by lysosomal hydrolases ().
Figure 1. Degradation and secretion converge at the endolysosomal system. To ensure efficient function in the regulation of intracellular homeostasis, the endolysosomal system orchestrates endocytosis, MVB formation, exosome secretion, autophagy induction and lysosomal degradation, coordinating the balance between the degradation and secretion mechanisms. Clearance of damaged or toxic material, including proteins, lipids and nucleic acids through exosomes, secretory autophagy or secretory lysosomes might alleviate intracellular stress and contribute to the preservation of cellular homeostasis. ER: Endoplasmic reticulum, MVB: Multivesicular body; MTOC: Microtubule-organizing center.
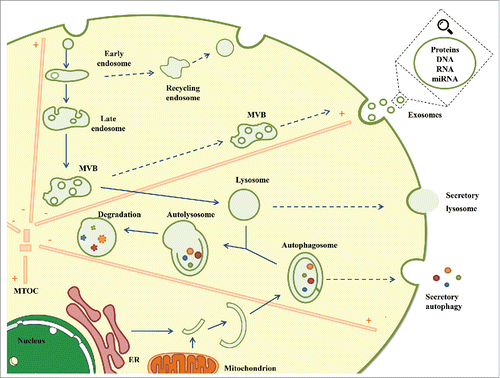
In contrast to degradative autophagy, autophagy has other role in unconventional secretion by a mechanism called secretory autophagy.Citation67 One of the best studied example of secretory autophagy is the release of the proinflammatory cytokine IL-1β that depends on the autophagy factor Atg5 and the membrane-associated small GTPase Rab8.Citation68 Rab8a has been involved in the vectorial sorting to plasma membrane of others secretory autophagic cargo proteins like α-synuclein. In this regard, Ejlerskov and colleagues showed evidence that secretion of α-synuclein requires Atg5 and is increased by lysosomal dysfunction.Citation69 Moreover, silencing of deacetylase HDAC6 increase the levels of acetylated tubulin, impairing the fusion of autophagosomes with lysosome and increasing the secretion of α-synuclein. Interestingly, α -synuclein secretion has been reported to be meditated by exosomes and the secretory autophagy pathways, underlining the crosstalk between both pathways to maintain cellular homeostasis. Autophagic cargo, depending on certain circumstances can be destined for autophagic degradation or secretion. Controlling autophagosomal motility along microtubules toward minus-end or plus-end could by an approach to direct autophagosomal organelles toward degradation or secretion.
Lysosomes are the principal degradative organelle of the cell.Citation70 Lysosomes degrade and recycle unwanted or damaged proteins and organelles to guarantee the continuous renewal of cellular constituents and to prevent the accumulation of toxic components. Besides their catabolic function, lysosomes contribute to different physiological processes including cell signaling, energy metabolism, plasma membrane repair, inflammation, or cell death.Citation71
Lysosomes contain a single membrane that isolates and protects the cell from the acidic environment of the lumen by an internal thick glycocalyx. Degradation in the lysosomes is mediated by multiple acid hydrolases, which include members of the proteases, peptidases, sulphatases, glycosidase, lipase and nuclease protein families; this wide repertoire allows degradation of different biological substrates including proteins, nucleic acids and lipids.
It is well known how the flux of cargo through the endolysosomal system and the induction of autophagy/lysosomal degradation are adjusted by mTOR signaling that integrates signaling from nutrients, growth factors, and energy availability.Citation72 Lysosomes respond to nutrients availability by moving toward the plasma membrane, whereas starvation results in a tighter perinuclear localization.Citation73 Mitochondrial respiration is absolutely required for lysosomal degradation since in cells that relay on glycolysis as a consequence of mitochondrial dysfunction, there is an impairment of lysosomal degradation and function.Citation74
Lysosomes may also act as secretory organelles in multiples cell types.Citation75 Lysosome release their content to plasma membrane as a mechanism for defense against parasites and for plasma membrane repair.Citation76,77 When proteins destined for degradation saturates the capacity of proteolytic system of the cell, as occurs in lysosome storage diseases, cells can defend themselves against proteotoxicity releasing dysfunctional lysosomes.Citation78,79 Other possibility is that cells compensate for lysosome malfunction by disposal of potentially toxic cargos in exosomes. Hence, it starts to become evident that the functional state of the lysosome affects the release of exosomes.Citation80-82
Concluding remarks
To ensure efficient function in the regulation of intracellular homeostasis, the endolysosomal system orchestrates endocytosis, MVB formation, exosome secretion, autophagy induction and lysosomal degradation, and coordinates the balance between degradation and secretion mechanisms (). The decision point between the 2 fates might be regulated by the cellular metabolic state, the nutrient availability or by external signals. Cellular status may affect the positioning of MVBs toward plasma membrane or perinuclear regions by switching microtubule motor proteins. Kinesin proteins may direct MVBs to plus end microtubule releasing MVBs as exosomes and dyneins family could guide MVBs toward lysosomes.Citation83 Understanding the signaling and the molecular clues that determine the traffic of cargo toward the lysosomes or the plasma membrane will allow us to manipulate the flux between degradation and secretion and could have immense relevance in the control of different pathological situations such as neurodegenerative or inflammatory diseases.
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
Acknowledgments
MM is a researcher from the Miguel Servet Program from Fondo de Investigación Sanitaria del Instituto de Salud Carlos III. We thank Dr. F. Baixauli for his advice and dedication.
Funding
This work is supported by the research grant CP 14/00219 from Fondo de Investigación Sanitaria del Instituto de Salud Carlos III, and co-funding by Fondo Europeo de Desarrollo Regional (FEDER).
Related Research Data
References
- Brodsky JL. Cleaning up: ER-associated degradation to the rescue. Cell 2012; 151:1163-7; PMID:23217703; http://dx.doi.org/10.1016/j.cell.2012.11.012
- MacGurn JA. Garbage on, garbage off: New insights into plasma membrane protein quality control. Curr Opin Cell Biol 2014; 29:92-8; PMID:24908345; http://dx.doi.org/10.1016/j.ceb.2014.05.001
- Babst M. Quality control: quality control at the plasma membrane: one mechanism does not fit all. J Cell Biol 2014; 205:11-20; PMID:24733583; http://dx.doi.org/10.1083/jcb.201310113
- McClellan AJ, Tam S, Kaganovich D, Frydman J. Protein quality control: chaperones culling corrupt conformations. Nat Cell Biol 2005; 7:736-41; PMID:16056264; http://dx.doi.org/10.1038/ncb0805-736
- Chen B, Retzlaff M, Roos T, Frydman J. Cellular strategies of protein quality control. Cold Spring Harb Perspect Biol 2011; 3:a004374; PMID:21746797; http://dx.doi.org/10.1101/cshperspect.a004374
- Tyedmers J, Mogk A, Bukau B. Cellular strategies for controlling protein aggregation. Nat Rev Mol Cell Biol 2010; 11:777-88; PMID:20944667; http://dx.doi.org/10.1038/nrm2993
- Vilchez D, Saez I, Dillin A. The role of protein clearance mechanisms in organismal ageing and age-related diseases. Nat Commun 2014; 5:5659; PMID:25482515; http://dx.doi.org/10.1038/ncomms6659
- Wolff S, Weissman JS, Dillin A, Vilchez D, Saez I, Dillin A, Tyedmers J, Mogk A, Bukau B, Chen B, et al. Differential scales of protein quality control. Cell 2014; 5:52-64; http://dx.doi.org/10.1016/j.cell.2014.03.007
- Lee J-G, Takahama S, Zhang G, Tomarev SI, Ye Y. Unconventional secretion of misfolded proteins promotes adaptation to proteasome dysfunction in mammalian cells. Nat Cell Biol 2016; 18:765-76; PMID:27295555; http://dx.doi.org/10.1038/ncb3384
- Colombo M, Raposo G, Thery C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu Rev Cell Dev Biol 2014; 30:255-89; PMID:25288114; http://dx.doi.org/10.1146/annurev-cellbio-101512-122326
- Mittelbrunn M, Sanchez-Madrid F. Intercellular communication: diverse structures for exchange of genetic information. Nat Rev Mol Cell Biol 2012; 13:328-35; PMID:22510790
- Yanez-Mo M, Siljander PR-M, Andreu Z, Zavec AB, Borras FE, Buzas EI, Buzas K, Casal E, Cappello F, Carvalho J, et al. Biological properties of extracellular vesicles and their physiological functions. J Extracell vesicles 2015; 4:27066; PMID:25979354; http://dx.doi.org/10.3402/jev.v4.27066
- Kalluri R. The biology and function of exosomes in cancer. J Clin Invest 2016; 126:1208-15; PMID:27035812; http://dx.doi.org/10.1172/JCI81135
- Lawson C, Vicencio JM, Yellon DM, Davidson SM. Microvesicles and exosomes: new players in metabolic and cardiovascular disease. J Endocrinol 2016; 228:R57-71; PMID:26743452; http://dx.doi.org/10.1530/JOE-15-0201
- Bellingham SA, Coleman BM, Hill AF. Small RNA deep sequencing reveals a distinct miRNA signature released in exosomes from prion-infected neuronal cells. Nucleic Acids Res 2012; 40:10937-49; PMID:22965126; http://dx.doi.org/10.1093/nar/gks832
- Fruhbeis C, Frohlich D, Kuo WP, Kramer-Albers E-M. Extracellular vesicles as mediators of neuron-glia communication. Front Cell Neurosci 2013; 7:182; PMID:24194697; http://dx.doi.org/10.3389/fncel.2013.00182
- Gutierrez-Vazquez C, Villarroya-Beltri C, Mittelbrunn M, Sanchez-Madrid F. Transfer of extracellular vesicles during immune cell-cell interactions. Immunol Rev 2013; 251:125-42; PMID:23278745; http://dx.doi.org/10.1111/imr.12013
- Mittelbrunn M, Gutierrez-Vazquez C, Villarroya-Beltri C, Gonzalez S, Sanchez-Cabo F, Gonzalez MA, Bernad A, Sanchez-Madrid F. Unidirectional transfer of microRNA-loaded exosomes from T cells to antigen-presenting cells. Nat Commun 2011; 2:282; PMID:21505438; http://dx.doi.org/10.1038/ncomms1285
- Peinado H, Aleckovic M, Lavotshkin S, Matei I, Costa-Silva B, Moreno-Bueno G, Hergueta-Redondo M, Williams C, Garcia-Santos G, Ghajar C, et al. Melanoma exosomes educate bone marrow progenitor cells toward a pro-metastatic phenotype through MET. Nat Med 2012; 18:883-91; PMID:22635005; http://dx.doi.org/10.1038/nm.2753
- Robbins PD, Morelli AE. Regulation of immune responses by extracellular vesicles. Nat Rev Immunol 2014; 14:195-208; PMID:24566916; http://dx.doi.org/10.1038/nri3622
- Baixauli F, Lopez-Otin C, Mittelbrunn M. Exosomes and autophagy: coordinated mechanisms for the maintenance of cellular fitness. Front Immunol 2014; 5:403; PMID:25191326; http://dx.doi.org/10.3389/fimmu.2014.00403
- Rink J, Ghigo E, Kalaidzidis Y, Zerial M. Rab conversion as a mechanism of progression from early to late endosomes. Cell 2005; 122:735-49; PMID:16143105; http://dx.doi.org/10.1016/j.cell.2005.06.043
- Hanson PI, Cashikar A. Multivesicular body morphogenesis. Annu Rev Cell Dev Biol 2012; 28:337-62; PMID:22831642; http://dx.doi.org/10.1146/annurev-cellbio-092910-154152
- Villarroya-Beltri C, Baixauli F, Gutierrez-Vazquez C, Sanchez-Madrid F, Mittelbrunn M. Sorting it out: regulation of exosome loading. Semin Cancer Biol 2014; 28:3-13; PMID:24769058; http://dx.doi.org/10.1016/j.semcancer.2014.04.009
- Bari R, Guo Q, Xia B, Zhang YH, Giesert EE, Levy S, Zheng JJ, Zhang XA. Tetraspanins regulate the protrusive activities of cell membrane. Biochem Biophys Res Commun 2011; 415:619-26; PMID:22079629; http://dx.doi.org/10.1016/j.bbrc.2011.10.121
- Edgar JR, Eden ER, Futter CE. Hrs- and CD63-dependent competing mechanisms make different sized endosomal intraluminal vesicles. Traffic 2014; 15:197-211; PMID:24279430; http://dx.doi.org/10.1111/tra.12139
- Perez-Hernandez D, Gutierrez-Vazquez C, Jorge I, Lopez-Martin S, Ursa A, Sanchez-Madrid F, Vazquez J, Yanez-Mo M. The intracellular interactome of tetraspanin-enriched microdomains reveals their function as sorting machineries toward exosomes. J Biol Chem 2013; 288:11649-61; PMID:23463506; http://dx.doi.org/10.1074/jbc.M112.445304
- van Niel G, Charrin S, Simoes S, Romao M, Rochin L, Saftig P, Marks MS, Rubinstein E, Raposo G. The tetraspanin CD63 regulates ESCRT-independent and -dependent endosomal sorting during melanogenesis. Dev Cell 2011; 21:708-21; PMID:21962903; http://dx.doi.org/10.1016/j.devcel.2011.08.019
- Trajkovic K, Hsu C, Chiantia S, Rajendran L, Wenzel D, Wieland F, Schwille P, Brugger B, Simons M. Ceramide triggers budding of exosome vesicles into multivesicular endosomes. Science 2008; 319:1244-7; PMID:18309083; http://dx.doi.org/10.1126/science.1153124
- Kajimoto T, Okada T, Miya S, Zhang L, Nakamura S. Ongoing activation of sphingosine 1-phosphate receptors mediates maturation of exosomal multivesicular endosomes. Nat Commun 2013; 4:2712; PMID:24231649; http://dx.doi.org/10.1038/ncomms3712
- Sahu R, Kaushik S, Clement CC, Cannizzo ES, Scharf B, Follenzi A, Potolicchio I, Nieves E, Cuervo AM, Santambrogio L. Microautophagy of cytosolic proteins by late endosomes. Dev Cell 2011; 20:131-9; PMID:21238931; http://dx.doi.org/10.1016/j.devcel.2010.12.003
- Huotari J, Helenius A. Endosome maturation. EMBO J 2011; 30:3481-500; PMID:21878991; http://dx.doi.org/10.1038/emboj.2011.286
- Harding C, Heuser J, Stahl P. Endocytosis and intracellular processing of transferrin and colloidal gold-transferrin in rat reticulocytes: demonstration of a pathway for receptor shedding. Eur J Cell Biol 1984; 35:256-63; PMID:6151502
- Johnstone RM, Adam M, Hammond JR, Orr L, Turbide C. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J Biol Chem 1987; 262:9412-20; PMID:3597417
- Guo BB, Bellingham SA, Hill AF. The neutral sphingomyelinase pathway regulates packaging of the prion protein into exosomes. J Biol Chem 2015; 290(6):3455-67.
- Rajendran L, Honsho M, Zahn TR, Keller P, Geiger KD, Verkade P, Simons K. Alzheimer's disease beta-amyloid peptides are released in association with exosomes. Proc Natl Acad Sci U S A 2006; 103:11172-7; PMID:16837572; http://dx.doi.org/10.1073/pnas.0603838103
- Perez-Gonzalez R, Gauthier SA, Kumar A, Levy E. The exosome secretory pathway transports amyloid precursor protein carboxyl-terminal fragments from the cell into the brain extracellular space. J Biol Chem 2012; 287:43108-15; PMID:23129776; http://dx.doi.org/10.1074/jbc.M112.404467
- Saman S, Kim W, Raya M, Visnick Y, Miro S, Saman S, Jackson B, McKee AC, Alvarez VE, Lee NCY, et al. Exosome-associated tau is secreted in tauopathy models and is selectively phosphorylated in cerebrospinal fluid in early Alzheimer disease. J Biol Chem 2012; 287:3842-9; PMID:22057275; http://dx.doi.org/10.1074/jbc.M111.277061
- Emmanouilidou E, Melachroinou K, Roumeliotis T, Garbis SD, Ntzouni M, Margaritis LH, Stefanis L, Vekrellis K. Cell-produced alpha-synuclein is secreted in a calcium-dependent manner by exosomes and impacts neuronal survival. J Neurosci 2010; 30:6838-51; PMID:20484626; http://dx.doi.org/10.1523/JNEUROSCI.5699-09.2010
- Grad LI, Yerbury JJ, Turner BJ, Guest WC, Pokrishevsky E, O'Neill MA, Yanai A, Silverman JM, Zeineddine R, Corcoran L, et al. Intercellular propagated misfolding of wild-type Cu/Zn superoxide dismutase occurs via exosome-dependent and -independent mechanisms. Proc Natl Acad Sci U S A 2014; 111:3620-5; PMID:24550511; http://dx.doi.org/10.1073/pnas.1312245111
- Fevrier B, Vilette D, Archer F, Loew D, Faigle W, Vidal M, Laude H, Raposo G. Cells release prions in association with exosomes. Proc Natl Acad Sci U S A 2004; 101:9683-8; PMID:15210972; http://dx.doi.org/10.1073/pnas.0308413101
- Hudson MB, Woodworth-Hobbs ME, Zheng B, Rahnert JA, Blount MA, Gooch JL, Searles CD, Price SR. miR-23a is decreased during muscle atrophy by a mechanism that includes calcineurin signaling and exosome-mediated export. Am J Physiol Cell Physiol 2014; 306:C551-8; PMID:24336651; http://dx.doi.org/10.1152/ajpcell.00266.2013
- Hudson MB, Rahnert JA, Zheng B, Woodworth-Hobbs ME, Franch HA, Russ Price S. miR-182 attenuates atrophy-related gene expression by targeting FoxO3 in skeletal muscle. Am J Physiol Cell Physiol 2014; 307:C314-9; PMID:24871856; http://dx.doi.org/10.1152/ajpcell.00395.2013
- de Candia P, Torri A, Gorletta T, Fedeli M, Bulgheroni E, Cheroni C, Marabita F, Crosti M, Moro M, Pariani E, et al. Intracellular modulation, extracellular disposal and serum increase of MiR-150 mark lymphocyte activation. PLoS One 2013; 8:e75348; PMID:24205408; http://dx.doi.org/10.1371/journal.pone.0075348
- Ohshima K, Inoue K, Fujiwara A, Hatakeyama K, Kanto K, Watanabe Y, Muramatsu K, Fukuda Y, Ogura S, Yamaguchi K, et al. Let-7 microRNA family is selectively secreted into the extracellular environment via exosomes in a metastatic gastric cancer cell line. PLoS One 2010; 5:e13247; PMID:20949044; http://dx.doi.org/10.1371/journal.pone.0013247
- Ostenfeld MS, Jeppesen DK, Laurberg JR, Boysen AT, Bramsen JB, Primdal-Bengtson B, Hendrix A, Lamy P, Dagnaes-Hansen F, Rasmussen MH, et al. Cellular disposal of miR23b by RAB27-dependent exosome release is linked to acquisition of metastatic properties. Cancer Res 2014; 74:5758-71; PMID:25261234; http://dx.doi.org/10.1158/0008-5472.CAN-13-3512
- Squadrito ML, Baer C, Burdet F, Maderna C, Gilfillan GD, Lyle R, Ibberson M, De Palma M. Endogenous RNAs modulate microRNA sorting to exosomes and transfer to acceptor cells. Cell Rep 2014; 8:1432-46; PMID:25159140; http://dx.doi.org/10.1016/j.celrep.2014.07.035
- van Balkom BWM, Eisele AS, Pegtel DM, Bervoets S, Verhaar MC. Quantitative and qualitative analysis of small RNAs in human endothelial cells and exosomes provides insights into localized RNA processing, degradation and sorting. J Extracell vesicles 2015; 4:26760; PMID:26027894; http://dx.doi.org/10.3402/jev.v4.26760
- Zhang X, Abels ER, Redzic JS, Margulis J, Finkbeiner S, Breakefield XO. Potential Transfer of Polyglutamine and CAG-Repeat RNA in Extracellular Vesicles in Huntington's Disease: Background and Evaluation in Cell Culture. Cell Mol Neurobiol 2016; 36:459-70; PMID:26951563; http://dx.doi.org/10.1007/s10571-016-0350-7
- Zhu Y, Chen X, Pan Q, Wang Y, Su S, Jiang C, Li Y, Xu N, Wu L, Lou X, et al. A Comprehensive Proteomics Analysis Reveals a Secretory Path- and Status-Dependent Signature of Exosomes Released from Tumor-Associated Macrophages. J Proteome Res 2015; 14:4319-31; PMID:26312558; http://dx.doi.org/10.1021/acs.jproteome.5b00770
- Batagov AO, Kuznetsov VA, Kurochkin IV. Identification of nucleotide patterns enriched in secreted RNAs as putative cis-acting elements targeting them to exosome nano-vesicles. BMC Genomics 2011; 3(12 Suppl):S18; http://dx.doi.org/10.1186/1471-2164-12-S3-S18
- Strauss K, Goebel C, Runz H, Möbius W, Weiss S, Feussner I, Simons M, Schneider A. Exosome secretion ameliorates lysosomal storage of cholesterol in Niemann-Pick type C disease. J Biol Chem 2010; 285:26279-88; PMID:20554533; http://dx.doi.org/10.1074/jbc.M110.134775
- Canfran-Duque A, Pastor O, Reina M, Lerma M, Cruz-Jentoft AJ, Lasuncion MA, Busto R. Curcumin Mitigates the Intracellular Lipid Deposit Induced by Antipsychotics In Vitro. PLoS One 2015; 10:e0141829; PMID:26517556; http://dx.doi.org/10.1371/journal.pone.0141829
- Kahlert C, Melo SA, Protopopov A, Tang J, Seth S, Koch M, Zhang J, Weitz J, Chin L, Futreal A, et al. Identification of double-stranded genomic DNA spanning all chromosomes with mutated KRAS and p53 DNA in the serum exosomes of patients with pancreatic cancer. J Biol Chem 2014; 289:3869-75; PMID:24398677; http://dx.doi.org/10.1074/jbc.C113.532267
- Guescini M, Genedani S, Stocchi V, Agnati LF. Astrocytes and Glioblastoma cells release exosomes carrying mtDNA. J Neural Transm 2010; 117:1-4; PMID:19680595; http://dx.doi.org/10.1007/s00702-009-0288-8
- Wang Z, Deng Z, Dahmane N, Tsai K, Wang P, Williams DR, Kossenkov AV, Showe LC, Zhang R, Huang Q, et al. Telomeric repeat-containing RNA (TERRA) constitutes a nucleoprotein component of extracellular inflammatory exosomes. Proc Natl Acad Sci U S A 2015; 112:E6293-300; PMID:26578789; http://dx.doi.org/10.1073/pnas.1505962112
- Wang Z, Lieberman PM. The crosstalk of telomere dysfunction and inflammation through cell-free TERRA containing exosomes. RNA Biol 2016; 13:690-5; PMID:27351774; http://dx.doi.org/10.1080/15476286.2016.1203503
- Ktistakis NT, Tooze SA. Digesting the Expanding Mechanisms of Autophagy. Trends Cell Biol 2016; 26:624-35; PMID:27050762; http://dx.doi.org/10.1016/j.tcb.2016.03.006
- Mizushima N, Levine B, Cuervo AM, Klionsky DJ. Autophagy fights disease through cellular self-digestion. Nature 2008; 451:1069-75; PMID:18305538; http://dx.doi.org/10.1038/nature06639
- Lamb CA, Yoshimori T, Tooze SA. The autophagosome: origins unknown, biogenesis complex. Nat Rev Mol Cell Biol 2013; 14:759-74; PMID:24201109; http://dx.doi.org/10.1038/nrm3696
- Lamb CA, Dooley HC, Tooze SA. Endocytosis and autophagy: Shared machinery for degradation. Bioessays 2013; 35:34-45; PMID:23147242; http://dx.doi.org/10.1002/bies.201200130
- Murrow L, Malhotra R, Debnath J. ATG12-ATG3 interacts with Alix to promote basal autophagic flux and late endosome function. Nat Cell Biol 2015; 17:300-10; PMID:25686249; http://dx.doi.org/10.1038/ncb3112
- Lee J-A, Gao F-B. Roles of ESCRT in autophagy-associated neurodegeneration. Autophagy 2008; 4:230-2; PMID:18094607; http://dx.doi.org/10.4161/auto.5384
- Fader CM, Colombo MI. Autophagy and multivesicular bodies: two closely related partners. Cell Death Differ 2009; 16:70-8; PMID:19008921; http://dx.doi.org/10.1038/cdd.2008.168
- Itakura E, Kishi-Itakura C, Mizushima N. The hairpin-type tail-anchored SNARE syntaxin 17 targets to autophagosomes for fusion with endosomes/lysosomes. Cell 2012; 151:1256-69; PMID:23217709; http://dx.doi.org/10.1016/j.cell.2012.11.001
- Takats S, Nagy P, Varga A, Pircs K, Karpati M, Varga K, Kovacs AL, Hegedus K, Juhasz G. Autophagosomal Syntaxin17-dependent lysosomal degradation maintains neuronal function in Drosophila. J Cell Biol 2013; 201:531-9; PMID:23671310; http://dx.doi.org/10.1083/jcb.201211160
- Ponpuak M, Mandell MA, Kimura T, Chauhan S, Cleyrat C, Deretic V. Secretory autophagy. Curr Opin Cell Biol 2015; 35:106-16; PMID:25988755; http://dx.doi.org/10.1016/j.ceb.2015.04.016
- Jiang S, Dupont N, Castillo EF, Deretic V. Secretory versus degradative autophagy: unconventional secretion of inflammatory mediators. J Innate Immun 2013; 5:471-9; PMID:23445716; http://dx.doi.org/10.1159/000346707
- Ejlerskov P, Rasmussen I, Nielsen TT, Bergstrom A-L, Tohyama Y, Jensen PH, Vilhardt F. Tubulin polymerization-promoting protein (TPPP/p25alpha) promotes unconventional secretion of alpha-synuclein through exophagy by impairing autophagosome-lysosome fusion. J Biol Chem 2013; 288:17313-35; PMID:23629650; http://dx.doi.org/10.1074/jbc.M112.401174
- Luzio JP, Hackmann Y, Dieckmann NMG, Griffiths GM. The biogenesis of lysosomes and lysosome-related organelles. Cold Spring Harb Perspect Biol 2014; 6:a016840; PMID:25183830; http://dx.doi.org/10.1101/cshperspect.a016840
- Settembre C, Fraldi A, Medina DL, Ballabio A. Signals from the lysosome: a control centre for cellular clearance and energy metabolism. Nat Rev Mol Cell Biol 2013; 14:283-96; PMID:23609508; http://dx.doi.org/10.1038/nrm3565
- Huber LA, Teis D. Lysosomal signaling in control of degradation pathways. Curr Opin Cell Biol 2016; 39:8-14; PMID:26827287; http://dx.doi.org/10.1016/j.ceb.2016.01.006
- Korolchuk VI, Saiki S, Lichtenberg M, Siddiqi FH, Roberts EA, Imarisio S, Jahreiss L, Sarkar S, Futter M, Menzies FM, et al. Lysosomal positioning coordinates cellular nutrient responses. Nat Cell Biol 2011; 13:453-60; PMID:21394080; http://dx.doi.org/10.1038/ncb2204
- Baixauli F, Acin-Perez R, Villarroya-Beltri C, Mazzeo C, Nunez-Andrade N, Gabande-Rodriguez E, Ledesma MD, Blazquez A, Martin MA, Falcon-Perez JM, et al. Mitochondrial Respiration Controls Lysosomal Function during Inflammatory T Cell Responses. Cell Metab 2015; 22:485-98; PMID:26299452; http://dx.doi.org/10.1016/j.cmet.2015.07.020
- Blott EJ, Griffiths GM. Secretory lysosomes. Nat Rev Mol Cell Biol 2002; 3:122-31; PMID:11836514; http://dx.doi.org/10.1038/nrm732
- Divangahi M, Chen M, Gan H, Desjardins D, Hickman TT, Lee DM, Fortune S, Behar SM, Remold HG. Mycobacterium tuberculosis evades macrophage defenses by inhibiting plasma membrane repair. Nat Immunol 2009; 10:899-906; PMID:19561612; http://dx.doi.org/10.1038/ni.1758
- Reddy A, Caler EV, Andrews NW. Plasma membrane repair is mediated by Ca(2+)-regulated exocytosis of lysosomes. Cell 2001; 106:157-69; PMID:11511344; http://dx.doi.org/10.1016/S0092-8674(01)00421-4
- Medina DL, Fraldi A, Bouche V, Annunziata F, Mansueto G, Spampanato C, Puri C, Pignata A, Martina JA, Sardiello M, et al. Transcriptional activation of lysosomal exocytosis promotes cellular clearance. Dev Cell 2011; 21:421-30; PMID:21889421; http://dx.doi.org/10.1016/j.devcel.2011.07.016
- Spampanato C, Feeney E, Li L, Cardone M, Lim J-A, Annunziata F, Zare H, Polishchuk R, Puertollano R, Parenti G, et al. Transcription factor EB (TFEB) is a new therapeutic target for Pompe disease. EMBO Mol Med 2013; 5:691-706; PMID:23606558; http://dx.doi.org/10.1002/emmm.201202176
- Alvarez-Erviti L, Seow Y, Schapira AH, Gardiner C, Sargent IL, Wood MJA, Cooper JM. Lysosomal dysfunction increases exosome-mediated alpha-synuclein release and transmission. Neurobiol Dis 2011; 42:360-7; PMID:21303699; http://dx.doi.org/10.1016/j.nbd.2011.01.029
- Vingtdeux V, Hamdane M, Loyens A, Gele P, Drobeck H, Begard S, Galas M-C, Delacourte A, Beauvillain J-C, Buee L, et al. Alkalizing drugs induce accumulation of amyloid precursor protein by-products in luminal vesicles of multivesicular bodies. J Biol Chem 2007; 282:18197-205; PMID:17468104; http://dx.doi.org/10.1074/jbc.M609475200
- Eitan E, Suire C, Zhang S, Mattson MP. Impact of lysosome status on extracellular vesicle content and release. Ageing Res Rev 2016; PMID:27238186; http://dx.doi.org/10.1016/j.arr.2016.05.001
- Granger E, McNee G, Allan V, Woodman P. The role of the cytoskeleton and molecular motors in endosomal dynamics. Semin Cell Dev Biol 2014; 31:20-9; PMID:24727350; http://dx.doi.org/10.1016/j.semcdb.2014.04.011