
ABSTRACT
Pathogens frequently use multivalent binding to sialic acid to infect cells or to modulate immunity through interactions with human sialic acid-binding immunoglobulin-type lectins (Siglecs). Molecules that interfere with these interactions could be of interest as diagnostics, anti-infectives or as immune modulators. This review describes the development of molecular scaffolds based on the crystallizable fragment (Fc) region of immunoglobulin (Ig) G that deliver high-avidity binding to innate immune receptors, including sialic acid-dependent receptors. The ways in which the sialylated Fc may be engineered as immune modulators that mimic the anti-inflammatory properties of intravenous polyclonal Ig or as blockers of sialic-acid-dependent infectivity by viruses are also discussed.
Introduction
Sialic acid-containing molecules play important roles in many fundamental immunological and pathological processes via carbohydrate–protein interactions that occur during the development of the immune system and during immune responses to pathogens.Citation1,Citation2 Examples include the regulation of fluid-phase innate immunity and modulation of leukocyte trafficking via sialylated selectin ligands.Citation3 Two members of the sialic acid family (Neu5Ac and Neu5Gc) commonly occur as the terminal constituents of carbohydrate chains and are attached through post-translational modifications to glycoproteins at N- and O-linked attachment sites within the protein ().
Figure 1. The crystal structure of human IgG1
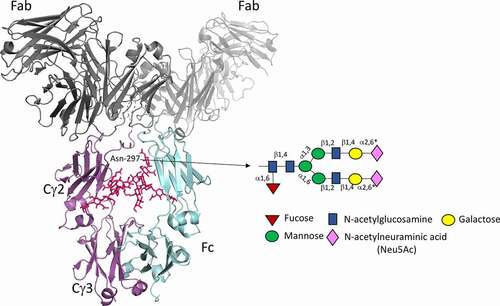
Immunoglobulin G (IgG) molecules, which are critical components of the immune system, have structures and functions that can be radically affected by changes to the glycan backbone (). Approximately 30–50 different glycan structures, with and without sialic acid, can be attached to N-linked attachment sites in IgG specified by the asparagine-X-threonine/serine (Asn-X-Thr/Ser) sequence, where X can be any amino acid other than proline.Citation5 Within IgG, one such conserved site at Asn-297 in the Fc is always occupied with a glycan, while 15–20% of variably occupied N-linked sites can also arise spontaneously in the antigen-binding fragments (Fabs) during antibody development ().Citation6
The oligosaccharides attached to Asn-297 are essential for the binding and activation of FcRs and complement component C1q as this functionality is abrogated or severely curtailed by enzymatic removal or site-directed mutagenesis of the Asn-X-Thr/Ser attachment site.Citation7–9 Numerous IgG-Fc crystal structures show Asn-297 glycans to be integral to the Fc structure, where they are buried within the internal cavity enclosed by the two CH2 domains (). This buried location makes it extremely difficult for the glycans to interact directly with glycan receptors but allows them to modify the overall conformation of the Fc peptide backbone,Citation10 thereby affecting the interactions with FcRs and complement component C1q ().
More detailed studies into the types of sugars involved in this functionality have shown enhanced FcRIIIA binding and antibody-dependent cell-mediated cytotoxicity (ADCC) of IgG1 in the absence of fucose;Citation11,Citation12 enhanced FcRIIIA binding but rapid clearance from the circulation of IgG1 enriched for oligomannose structures;Citation13–15 and improved solubility, anti-inflammatory activity, thermal stability, and circulatory half-life of terminally sialylated glycans from IgG1.Citation16–20 The sialylation of IgG Fc domains also impairs complement-dependent cytotoxicity.Citation21 The presence of Asn-297 terminal capping sialic acid may also prevent clearance of IgG by the asialoglycoprotein receptor that binds to terminal galactose residues of N-glycans.Citation22 Therefore, glycosylation, including terminal sialylation, is important for antibody function. Consequently, this review will explore how IgG, and in particular the Fc, may be engineered to enhance interactions with sialic acid-binding receptors that play a crucial role in the anti-inflammatory properties of IgG or as therapeutic blockers of sialic-acid dependent infectivity by viruses.
Sialic acid and the anti-inflammatory properties of IgG
Intravenously administered IgG (IVIG) is a highly successful biologic approved for treating several autoimmune diseases (ADs), including idiopathic thrombocytopenic purpura (ITP), chronic inflammatory demyelinating polyneuropathy, myasthenia gravis, and other neurological illnesses.Citation23 As ~70% of the global supply (worth ~$8.9 billion in 2017) of IVIG is now used to treat ADs, it can be unavailable to other patients who desperately need it, in particular individuals with primary immune deficiency where IVIG is used as replacement therapy.Citation24 Worldwide consumption of IVIG has increased over 400-fold since 1980 and currently >100 tons are consumed per annum. Global supplies of IVIG are critically limited, meaning that patients with an urgent need for the drug can have restricted access to it (https://www.alliedmarketresearch.com/intravenous-immunoglobulin-IVIG-market).
The therapeutic utility of IVIG has substantial limitations, including dependence on human donors for its manufacture, and from the fact that less than 5% of injected IVIG is therapeutically active, leading to a requirement for high doses (1–2 g/kg) when used in treating most ADs. Consequently, IVIG is expensive and adverse events due to excessive protein loading are not uncommon.Citation23,Citation24 There is thus an urgent clinical need to develop cheaper, safer, and more effective alternatives to IVIG that are effective at lower dosages, although efforts have been hindered by a lack of understanding of its likely pleiotropic mechanisms of action.Citation25
Many therapeutic modes of action have been attributed to IVIG. It is beyond the scope of this review to cover all such mechanisms in detail, but several excellent review articles on the topic are available.Citation26–29 Here, the focus is on the mechanisms in which sialic acid engineering approaches have been undertaken.
Changes in antibody sialylation have been associated with the evolution of autoimmune and inflammatory diseases, including rheumatoid arthritis and juvenile idiopathic arthritis, which are associated with decreased levels of IgG sialylation, and particularly of the pathogenic IgG found in inflamed joints of patients.Citation30,Citation31 Levels of sialylated IgG increase during pregnancy and correlate with remission from arthritis in pregnant women.Citation32 Furthermore, pathogenic anti-proteinase three autoantibodies are less sialylated in patients with active Wegener’s vasculitis.Citation33 The observation that increased levels of endogenous IgG sialylation increased the likelihood of successful treatment of Kawasaki disease in patients treated with IVIG, and that patients with severe Guillain-Barré syndrome also show lower levels of IgG sialylation despite IVIG treatment have stimulated research into converting such observations into therapies.Citation34,Citation35 These clinical observations have been substantiated in multiple mouse models of arthritis and other ADs where approximately 10-fold improvements in the severity of disease are seen with sialic-acid enriched IVIG.Citation20,Citation35 Approximately 30-fold improvements over IVIG in anti-inflammatory activity can also be demonstrated using sialylated Fc, recombinant antibodies or hypersialylated IVIG preparations (e.g., M254).Citation19,Citation20,Citation36,Citation37 Developed by Momenta Pharmaceuticals, Inc., M254 was shown to be well tolerated in healthy subjects,Citation38 and is now being investigated in patients with ITP (Phase 1/2 study NCT03866577).
Sialic acid engineering approaches to the Fc glycans
The controlling influence of Asn-297 oligosaccharides on Fc-mediated effector functions of antibodies has driven experimental approaches to modify them, either through glycoengineering/chemoenzymatic means,Citation19,Citation39 by mutagenesis programs on the Fc protein backbone that disrupt the protein-Asn-297-carbohydrate interface,Citation40 or by expression in glycosidase-adapted transgenic cell lines.Citation41 For example, the marketed humanized antibody mogamulizumab, used to treat lymphoma, is manufactured in Chinese hamster ovary (CHO) cells in which the α(1-6)-fucosyltransferase (FUT8) gene is removed, resulting in an afucosylated IgG1 with enhanced FcRIIIA-dependent tumor cell killing by ADCC.Citation42 Although similar approaches have yielded enhanced sialylation of IgG, with zero to moderate improvements in binding to FcRs,Citation19,Citation40,Citation43,Citation44 these have not led to significant enhancements in binding to inhibitory Siglecs that are important in controlling unwanted inflammation,Citation44–46 a finding that others have attributed to the buried location of the Asn-297-attached glycan within the Fc.Citation10,Citation47
Approaches to enhance the sialylation of IgG have to date focussed on modifications to the known preexisting N-linked glycosylation sites.Citation19,Citation40,Citation48,Citation49 My colleagues and I took an alternative approach to enhancing the sialic acid content of the Fc of IgG1 by adding the 18 amino acid tailpiece from IgM that contains an N-linked glycosylation site at Asn-563 to the C-terminus of the IgG1 Fc,Citation50–53 and into which a cysteine-to-alanine substitution at Cys-575 may be introduced to prevent covalent multimerization (). A further N-linked glycosylation site can also be introduced, if desired, to the N terminus at position Asn-221.Citation50 By inserting or removing in different combinations the Asn-221, Asn-297 and Asn-563 glycosylation sites, a panel of variably glycosylated Fc monomers can be generated.Citation51 As a result, tetravalent, octavalent, and dodecavalent Fc monomers can be made with respect to the attached terminal sialic acid (). Both non-covalent or covalently bonded higher ordered multimers, e.g., with possible sialic acid valences up to 72 for the hexamers depicted in , can then be generated from the basic Fc unit by the addition or removal of cysteines (Cys-309/Cys-575), either alone or in combination with the tailpiece Asn-563 glycan, that radically increase the available oligosaccharide combining sites (). Citation47,Citation51,Citation53
Figure 2. A model of sialylated Fc

Hexamers of the IgG1-Fc with no additional N-glycosylation sites introduced, other than the glycan found at Asn-297, are currently in clinical development as they have been shown to block cytotoxicity and pathological changes in experimental in vitro and rat models of neuromyelitis optica through mechanisms that involve interference with complement activation.Citation56–58 However, potential drawbacks to the clinical use of hexamers include their large size (~350 kDa) and presence of multiple disulfide bonds that may combine to limit manufacture to scale by commercially available cell lines.
We therefore created a large panel of cysteine- and N-glycan-adapted mutants from the parent hexamer, including molecules with additional N-link attachment sites at Asn-221 and Asn-563.Citation50–52 Because the Asn-297 glycan is largely buried within the Fc,Citation10,Citation47 () the location of Asn-221 and Asn-563 at the tips of the Fc imparts alternative functional attributes to these molecules (). Citation51 Five combinations of glycosylation and cysteine substitution mutants that formed either monomers or multimers and possessed different binding characteristics for FcRs, C- and I-type lectins and complement components were shortlisted.Citation55 As sialylation of IgG-Fc domains is believed to be important for the anti-inflammatory effects of IVIG,Citation35,Citation59,Citation60 molecules containing simple mono-antennary sialylation and larger more complex tri- and tetra-antennary sialylation were selected for study in an ex vivo model of antibody-mediated demyelination of the nervous system.Citation55 The relative abundance of complex sialylated structures on mutants that did not protect (e.g., D221N/C309L/N297A/C575A), compared to the paucity of sialylated structures on mutants that did protect, argues against a direct role for sialic acid in this particular model of neurological disease.Citation55 This observation is supported by studies in a number of AD models that have shown the protective effects of IVIG to be largely independent of sialylation or interactions with DC-SIGN.Citation44,Citation61 However, the sialylation state of the Fc may become critical in vivo, especially for neurological diseases, where the influx and efflux of IgG through the blood–brain barrier has been shown to be dependent on glycan- and sialic acid-dependent mechanisms.Citation62,Citation63
Additional heterogeneity to the glycan profiles with dramatic functional consequences can occur by expressing the molecules in either CHO or human endothelial kidney (HEK) cell lines.Citation51 Unlike CHO cells, HEK cells have an active α2,6-sialyltransferase. As such, CHO-derived Fcs can only be sialylated through α2,3 linkages, whereas both α2,3 and α2,6 linkages can be found on molecules expressed by HEK cells.Citation51 The potential clinical and therapeutic applications of these variably glycosylated Fc molecules are discussed below.
Modulation of Siglecs
Through their capacity to activate or inhibit immune responses, Siglecs have become attractive therapeutic targets.Citation64–66 Humans possess 14 different Siglecs that bind sialic acid. Siglec-3 (also known as CD33) belongs to a group of related Siglecs that include Siglec-5,-6, −7, −8, −9, −10, −11.Citation64 These inhibitory Siglecs carry immunoreceptor tyrosine-based inhibitory motifs (ITIMs) and/or ITIM-like motifs in their cytoplasmic domains that lead to signaling cascades that suppress the activity of immune cells leading to anti-inflammatory effects.Citation67 Consequently, synthetic sialylated ligands, such as sialo-polymers, sialo-nanoparticles, sialylated RNAs, and sialo-liposomes, that can bind Siglecs are being developed.Citation68–72 Other promising strategies for enhancing sialylation use exogenous sialyl-transferases and donors, as well as blockade of glycosphingolipid biosynthesis.Citation49,Citation73
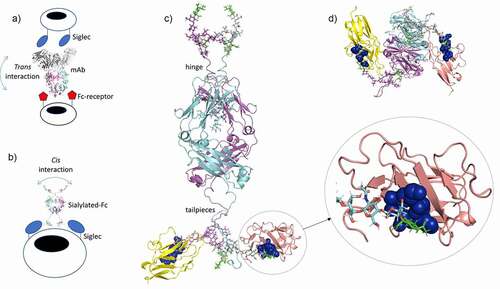
Many of these, including anti-CD33 monoclonal antibodies (mAbs) in clinical development, will interact with Siglecs in trans (), meaning that these synthetic ligands engage cell-surface Siglecs from the surrounding medium. Cross-linking CD33 on monocytes via antibodies induces pro-inflammatory effects, while cis binding of sialic acids to CD33 represses IL-1β production by monocytes.Citation75 Consequently, cis ligands for Siglecs that reside on the same cell membrane may be superior at maintaining inhibitory signals that increase the threshold for immune activation, and this may make them useful for anti-inflammatory therapy (). Citation67 Indeed, the depletion of cis Siglec ligands has been shown to increase the activity of both macrophages and microglia, and other studies have shown that the metabolic blockade of sialic acid renders phagocytes more prone to activation.Citation67 Based on these observations, it is anticipated that small high-affinity multivalent sialic acid-ligands that are better at diffusing in and around membranes than larger synthetic sialic acid-liganded molecules would have less potential for trans-mediated binding. Compared to other sialylated scaffolds that may bind CD33 in trans, e.g., antibodies, the sialylated Fc offers a number of attractive biophysical and therapeutic properties over chemically generated non-natural sialoside polymerization platforms (or even mAbs) for targeting Siglecs therapeutically in cis ().
Table 1. Different carbohydrate polymer approaches to target sialic acid-binding ligands using influenza virus as an example
Figure 3. A model for the known interaction of sialylated Fc with Siglec-3 (CD33)
We previously identified sialylated Fc candidates that bound CD33 yet showed minimal binding to Fc-receptors and complement C1q.Citation52 Siglec-3 is a validated target for acute myeloid leukemia, and versions of the receptor that cannot bind sialic acid correlate with susceptibility to Alzheimer’s disease.Citation77,Citation78 Siglec-3 is also believed to be a common determinant of SARS-CoV-2 infection and severe respiratory distress syndrome mediated by cytokine storms.Citation79
The Fc glycovariant D221N/C309L/C575A (shown in ) bound CD33 when expressed by CHO-K1 but did not bind when expressed by HEK cells,Citation52 suggesting that α2,3-linked sialosides are more important to CD33 binding as these are the only linkages attached by CHO cells. This is in line with CD33 having been reported to have a preference for α2,3- over α2,6-linked sialosides with a reported weak binding affinity (EC50 = 2–5 mM).Citation80 The corresponding mutant in which the hinge Asn-221 glycan was removed (C309L/C575A) is still capable of binding CD33 indicating that the primary CD33 interacting glycans are located in the tailpiece at Asn-563 ().
We therefore generated a model of D221N/C309L/C575A to which sialic acid was attached via α2,3-linkages to Asn-221 and Asn-563 (). These structures were then docked to the known sialoside binding site of CD33 (). Only one arm of the α2,3-sialylated tailpiece glycan could be accommodated within the shallow-binding pocket of the N-terminal V-set domain, with contacts made to the conserved Arg-119, Lys-126, Lys-130, and Phe-117 residues seen in the ligand-bound crystal (). Superimposition of the docked CD33 V-set domain with the other available crystal structures from Siglecs-1, 2, and 4, offered possible models to explain why this Fc glycovariant could also bind these receptors.Citation51,Citation52
We have yet to test binding of D221N/C309L/C575A, or indeed any of the other Fc mutants, to Siglecs-6, −7, −8 and −9 largely because the commercially available Siglecs are direct fusions to the Fc. In a similar vein, we were unable to test binding to human Siglec-5 because we observed significant direct binding of Fab′2 detecting reagents to this receptor. The Fab′2-mediated binding to Siglec-5 was dependent on glycans, because treatment of the Fab′2 detecting reagent with neuraminidase abrogated binding to Siglec-5.Citation50,Citation81 Siglec-5 may therefore be a target for Fab glycans that have also been associated with the anti-inflammatory activity of IVIG.Citation6 As the described Fc mutants can contain many different sialylated structures,Citation51,Citation55 it will be important to precisely define Siglec binding to avoid potential off-target effects given the ubiquitous expression of Siglecs on many different types of cells and tissues. Greater homogeneity (leading to enhanced specificity) to the type of sialylated structures attached to Asn-221, Asn-297, or Asn-563 may be imparted using enzymatic and/or click-chemistry approaches described by others.Citation19,Citation49
The D221N/C309L/C575A glycovariant may target CD33 in cis (), as the negatively charged N-terminal hinge-located Asn-221 glycans may discourage interactions with FcRs in trans that are more likely to occur with mAb approaches (). Furthermore, the considerably smaller size of the Fc offers other advantages, including superior penetrability of, for example, the blood–brain barrier or of hard tumors, that may be more difficult to achieve with mAbs, sialo-polymers, sialo-nanoparticles, or sialo-liposomes (). Therefore, approaches that combine antibody Fc and glyco-mimetic targeting of Siglecs may offer advantages over mAb-only or glycomimetic-only strategies.
Sialic acid receptors of viruses
Sialic acids linked to glycoproteins and gangliosides are used by many viruses as a receptor for cell entry.Citation82,Citation83 Such viruses include significant human and animal pathogens, including influenza, parainfluenza, corona, mumps, noro, rota, adeno, and DNA tumor viruses.Citation82 Attachment to sialic acid is mediated through receptor-binding proteins that are exposed at the surface of non-enveloped viruses. Some of these viruses are also equipped with neuraminidase or a sialyl-O-acetyl-esterase, which are receptor-destroying enzymes that can promote virus release from infected cells and neutralize sialic acid-containing soluble proteins that interfere with cell surface binding of the virus.Citation84
One example is the influenza virus, which assembles hundreds of hemagglutinin (HA) trimers on its surface to recognize sialic acid-galactose linkages on target tissue.Citation84,Citation85 The monovalent interaction between HA and a typical sialylated lactosamine ligand is weak (mM range), but multivalency-enhanced interactions allow firmer adhesion at low concentration.Citation86,Citation87 Molecules that bind HA with high avidity may therefore be useful as diagnostics or anti-infective medicines.Citation68,Citation86,Citation87 Consequently, many studies have explored multivalent scaffolds to present sialic acid to HA with the aim of blocking the interaction between virus and host receptors.Citation86–93 Many different scaffolds have been used to chemically attach sialic acid, including antibodies, DNA, fullerenes, graphene, polyacrylamide, quantum dots, magnets, silver and gold nanoparticles, although biocompatibility and potential toxicological liabilities of all of these remain and none have progressed to approval for use in humans.Citation76,Citation94 Controlling the spatial distribution and number of ligand-bearing units in these oligomers can also be sub-optimal and/or ill-defined, leading to reduced binding or promiscuous binding to other sialic acid receptors (). The lack of target specificity can lead to faster in vivo clearance rates and may also explain reported toxicities for many polymeric inhibitors.Citation95,Citation96
This has driven the search for smaller, rationally designed sialylated scaffolds. Using a rigid self-assembled peptide nucleic acid complex, it has recently been shown that smaller bivalent displays of the natural sialyl-LacNAc ligand (50–68 Å between each sugar) are more effective at binding a single HA trimer and inhibiting hemagglutination by virus than longer scaffolds with sialic acid residues separated by distances >100 Å that may allow for inter-HA bridging, which may also be desirable.Citation86,Citation87 Electron micrograph analyses have shown the average distance between two adjacent HA trimers to be 101.7±0.6 Å.Citation86 ()
Figure 4. A model for the known interaction of the sialylated Fc with influenza hemagglutinin
We have previously shown that a sialylated Fc construct (molecule D221N/C309L/N297A/C575A) generated in CHO-K1 cells could potently inhibit binding to HA and blocked hemagglutination by influenza A and B viruses.Citation51,Citation52 Although D221N/C309L/N297A/C575 does not bind FcRs or C1q, other Fc mutants that partially blocked influenza B and retained FcR and C1q binding were also made, e.g., D221N/N563A/C575A.Citation51 The lack of binding to FcRs or complement may be therapeutically useful, as no interference with neutralizing antibodies raised by influenza vaccines would be expected. Through additional interactions with Siglecs, the sialylated Fc can mimic the known anti-inflammatory pathways of IgG that can protect patients from overt inflammatory responses, e.g., cytokine storms that can kill.Citation99,Citation100
In contrast to CD33, binding to HA is driven by the hinge-attached glycan located at Asn-221, as its removal in the C309L/N297A/C575A mutant abrogated binding.Citation52 The lower hemagglutination inhibition assay (HIA) activity seen by the C309L/N297A/C575A mutant, in which the tailpiece Asn-563 glycan is retained, was surprising as the tailpiece Asn-563 glycan can be required for the anti-influenza virus activity of human anti-HIV virus IgA.Citation101 All Fc constructs tested to date in the HIA assay contain adapted human IgM tails, but the findings of Maurer et al.,Citation101 suggest that different binding to HA may be acquired using the IgA backbone in which the tailpiece differs at seven of the 18 amino acids to the tailpiece found in IgM.
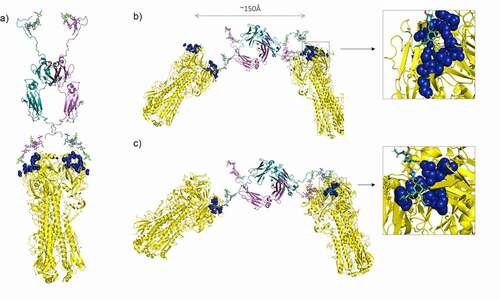
The narrowest and widest distances achievable between the sialic acid residues in our modeled Fc are ~30–70 Å and 110–150 Å, respectively (), suggesting that the sialic acid-adapted Fc may allow both intra-HA and inter-HA binding (). The hinge and tailpiece regions of the Fc are known to be highly flexible and the attached sialic acids may therefore reach more widely spaced HA trimers on the surface of more than one virus particle.Citation102 We previously observed that this octa-sialylated Fc was consistently more effective at inhibiting influenza B than influenza A viruses (EC50, ~30 nM vs. ~250 nM in HIA).Citation51 To investigate possible structural explanations for these observations, we evaluated the molecular interactions of the D221N/C309L/N297A/C575A sialylated Fc with known crystal structures of HA from both influenza A and B viruses ().
We docked one neuraminic acid unit from each of the two hinges into the cleft specifying the receptor-binding domain (RBD) of HA from influenza A (), with one face of the pyranose ring toward the bottom of the depression, and the other face exposed to solution as in published structures.Citation97,Citation103,Citation104 All the relevant ring substituents of α-NeuAc can interact with known key residues including, Trp-153, Asp-190, Thr-136, Lys-155, Lys-221, and Glu-226 within the RBD (). In this model, adjacent N-acetylgalactosamine and N-acetylglucosamine can also form independent interactions with the peptide backbone of HA ().
Influenza B diverged from influenza A approximately 4,000 years agoCitation105 and we were interested in understanding how the octa-sialylated Fc inhibited hemagglutination by the B virus more effectively than the A virus. The influenza B RBD in the globular head of HA is noticeably longer and wider, allowing the sialic acid on the Fc to be more comfortably accommodated within the RBD.Citation98 ()
Sialic acid is also the substrate for influenza neuraminidase (NA). We do not yet know if sialylated Fcs are susceptible to cleavage by influenza NA. Although a decoy for NA may be a therapeutically attractive strategy,Citation106 we have not observed a direct decay in the HIA after prolonged incubation. This suggests that the high specific avidity of these molecules for HA may reduce their susceptibility to NA, a hypothesis that fits with the relatively low efficiency of NA (kcat = 30–155s−1), together with the asymmetric distribution of NA in relation to HA on the surface of filamentous influenza viruses.Citation76,Citation84,Citation107
To be useful, in compounds when administered intranasally or as an aerosol, the sialylated Fc needs to outcompete the sialylated mucins that viruses use, through ratchet-like interactions with HA and NA, to migrate to the underlying respiratory epithelium.Citation84 Of the 15 known human mucins in the human lung, only MUC5 has been shown to give protection from influenza.Citation108,Citation109 Most sialic acid found on human mucins are O-glycosylated, and where N-linked attachments do occur, these are mostly sialylated via α2,6-linkages.Citation109 Thus, we were surprised that none of the Fc leads inhibited influenza A or influenza B agglutination of human O+ erythrocytes when manufactured by HEK cells, which attach the more human type α2,6-linked sialic acid.Citation51 The apparent importance of α2,3-linked N-glycans to the inhibition of both influenza A and B by the CHO-K1 Fc mutants indicates that viruses can evolve away from inhibition by mucus, whose predominant O-linked glycans are mostly α2,6-linked.Citation109,Citation110 Our working hypothesis is that HEK-expressed sialylated Fc may inhibit influenza viruses that circulate in human populations or that are propagated in cell lines that attach more human-like α2,6-linked sialic acid. The findings suggest that sialylated Fcs may also be useful blockers of coronavirus S glycoproteins that mediate attachment to oligosaccharide receptors, such as MERS-CoV, which also binds α2,3-linked, and to a lesser extent, α2,6-linked sialic acids.Citation82,Citation111
Recent reports have demonstrated that the current H3N2 viruses no longer have a strict specificity toward human-type receptors, which may result in loss of binding by the sialylated Fc to these viruses.Citation112 It has become clear that H3N2 viruses maintain human-type specificity but have evolved a preference for a subset of sialylated receptors with branched glycans and extended poly-N-acetyl-lactosamine (poly-LacNAc) chainsCitation113 that are not attached to any of the inhibitory Fcs expressed in either CHO or HEK cells.Citation51 Because this specificity is also shared with the 2009 pandemic H1N1, enhanced Fc blockers may potentially be created by manufacturing these molecules in human respiratory cell lines where such sialic acid attachments are known to occur.Citation114 Alternatively, enzymatic and/or click-chemistry approaches may be used to build the required glycan structure onto the Asn-221 and Asn-563 acceptor sites as documented previously for IgG.Citation19,Citation49
In summary, multivalent sialylated IgG Fcs offer many advantages over existing approaches to deliver high-avidity blocking or triggering of sialic acid-dependent receptors, such as Siglecs (). The proven abilities of the Fc to be intravenously injectedCitation16 and of Fc-fusions to be delivered directly into the eyeCitation115 or as an aerosol to the respiratory tractCitation116 are particularly noteworthy. As the binding epitope for all hemagglutinins is sialic acid and is determined by the host, these ligands are less prone to viral escape by genetic drift compared to mAbs and, unlike mAbs, are more readily manufactured and improvable through click-chemistry approaches to the glycan backbone as a consequence of introducing additional N-linked glycosylation at exposed sites in the Fc.
Abbreviations
ADE, antibody-dependent enhancement; C1q, complement component 1q; CD, cluster designation; CH2, constant heavy 2; CHO, Chinese Hamster Ovary; Fc, fragment crystallizable; HA, hemagglutinin; HEK, human endothelial kidney; HIA, hemagglutination inhibition assay; IgG, immunoglobulin G; ITIM, Immunoreceptor tyrosine-based inhibitory motif; IVIG, intravenous immunoglobulin; Neu5Ac, N-acetylneuraminic acid; Neu5Gc, N-glycolylneuraminic acid; Siglec, sialic acid-binding immunoglobulin-type lectin.
Acknowledgments
The author thanks the Wellcome Trust for funding this work through an Innovator grant, 208938/Z/17/Z.
Disclosure statement
R.J.P. declares that the molecules discussed within are subject to ongoing patent applications. The author has no financial conflicts of interest.
Additional information
Funding
References
- Schauer R, Kamerling JP. Exploration of the Sialic Acid world. Adv Carbohydr Chem Biochem. 2018;75:1–12.
- Dalziel M, Crispin M, Scanlan CN, Zitzmann N, Dwek RA. Emerging principles for the therapeutic exploitation of glycosylation. Science. 2014;343(6166):1235681–87. doi:10.1126/science.1235681.
- Varki A, Gagneux P. Multifarious roles of sialic acids in immunity. Ann N Y Acad Sci. 2012;1253(1):16–36. doi:10.1111/j.1749-6632.2012.06517.x.
- Saphire EO, Parren PW, Pantophlet R, Zwick MB, Morris GM, Rudd PM, Dwek RA, Stanfield RL, Burton DR, Wilson IA. Crystal structure of a neutralizing human IGG against HIV-1: a template for vaccine design. Science. 2001;293(5532):1155–59. doi:10.1126/science.1061692.
- Mimura Y, Katoh T, Saldova R, O’Flaherty R, Izumi T, Mimura-Kimura Y, Utsunomiya T, Mizukami Y, Yamamoto K, Matsumoto T. Glycosylation engineering of therapeutic IgG antibodies: challenges for the safety, functionality and efficacy. Protein Cell. 2018;9(1):47–62. doi:10.1007/s13238-017-0433-3.
- van de Bovenkamp FS, Hafkenscheid L, Rispens T, Rombouts Y. The emerging importance of IgG Fab glycosylation in immunity. J Immunol. 2016;196(4):1435–41. doi:10.4049/jimmunol.1502136.
- Lund J, Takahashi N, Pound JD, Goodall M, Jefferis R. Multiple interactions of IgG with its core oligosaccharide can modulate recognition by complement and human Fcy receptor I and influence the synthesis of its oligosaccharide chains. J Immunol. 1996;157:4963–69.
- Wright A, Morrison SL. Effect of altered CH2-associated carbohydrate structure on the functional properties and in vivo fate of chimeric mouse-human immunoglobulin G1. J Exp Med. 1994;180(3):1087–96. doi:10.1084/jem.180.3.1087.
- Pincetic A, Bournazos S, DiLillo DJ, Maamary J, Wang TT, Dahan R, Fiebiger B-M, Ravetch JV. Type I and type II Fc receptors regulate innate and adaptive immunity. Nat Immunol. 2014;15(8):707–16. doi:10.1038/ni.2939.
- Subedi GP, Hanson QM, Barb AW. Restricted motion of the Conserved Immunoglobulin G1 N-glycan is essential for efficient FcγRIIIa binding. Structure. 2014;22(10):1478–88. doi:10.1016/j.str.2014.08.002.
- Shields RL, Lai J, Keck R, O’Connell LY, Hong K, Meng YG, Weikert SHA, Presta LG. Lack of fucose on human IgG1 N-linked oligosaccharide improves binding to human FcγRIII and antibody-dependent cellular toxicity. J Biol Chem. 2002;277(30):26733–40. doi:10.1074/jbc.M202069200.
- Shinkawa T, Nakamura K, Yamane N, Shoji-Hosaka E, Kanda Y, Sakurada M, Uchida K, Anazawa H, Satoh M, Yamasaki M. The absence of fucose but not the presence of galactose or bisecting N-acetylglucosamine of human IgG1 complex-type oligosaccharides shows the critical role of enhancing antibody-dependent cellular cytotoxicity. J Biol Chem. 2003;278(5):3466–73. doi:10.1074/jbc.M210665200.
- Kanda Y, Yamada T, Mori K, Okazaki A, Inoue M, Kitajima-Miyama K, Kuni-Kamochi R, Nakano R, Yano K, Kakita S. Comparison of biological activity among nonfucosylated therapeutic IgG1 antibodies with three different N-linked Fc oligosaccharides: the high-mannose, hybrid, and complex types. Glycobiology. 2007;17(1):104–18. doi:10.1093/glycob/cwl057.
- Yu M, Brown D, Reed C, Chung S, Lutman J, Stefanich E, Wong A, Stephan JP, Bayer R. Production, characterization and pharmacokinetic properties of antibodies with N-linked mannose-5 glycans. MAbs. 2012;4(4):475–87. doi:10.4161/mabs.20737.
- Goetze AM, Liu YD, Zhang Z, Shah B, Lee E, Bondarenko PV, Flynn GC. High-mannose glycans on the Fc region of therapeutic IgG antibodies increase serum clearance in humans. Glycobiology. 2011;21(7):949–59. doi:10.1093/glycob/cwr027.
- Debre M, Bonnet MC, Fridman WH, Carosella E, Philippe N, Reinert P, Vilmer E, Kaplan C, Teillaud JL, Griscelli C. Infusion of Fc gamma fragments for treatment of children with acute immune thrombocytopenic purpura. Lancet. 1993;342(8877):945–49. doi:10.1016/0140-6736(93)92000-J.
- Liu L. Antibody glycosylation and its impact on the pharmacokinetics and pharmacodynamics of monoclonal antibodies and Fc-fusion proteins. J Pharm Sci. 2015;104(6):1866–84. doi:10.1002/jps.24444.
- Zhang G, Massaad CA, Gao T, Pillai L, Bogdanova N, Ghauri S, Sheikh KA. Sialylated intravenous immunoglobulin suppress anti-ganglioside antibody mediated nerve injury. Exp Neurol. 2016;282:49–55. doi:10.1016/j.expneurol.2016.05.020.
- Washburn N, Schwab I, Ortiz D, Bhatnagar N, Lansing JC, Medeiros A, Tyler S, Mekala D, Cochran E, Sarvaiya H. Controlled tetra-Fc sialylation of IVIg results in a drug candidate with consistent enhanced anti-inflammatory activity. Proc Natl Acad Sci U S A. 2015;112(11):E1297–306. doi:10.1073/pnas.1422481112.
- Anthony RM, Nimmerjahn F, Ashline DJ, Reinhold VN, Paulson JC, Ravetch JV. Recapitulation of IVIG anti-inflammatory activity with a recombinant IgG Fc. Science. 2008;320(5874):373–76. doi:10.1126/science.1154315.
- Quast I, Keller CW, Maurer MA, Giddens JP, Tackenberg B, Wang L-X, Münz C, Nimmerjahn F, Dalakas MC, Lünemann JD. Sialylation of IgG Fc domain impairs complement-dependent cytotoxicity. J Clin Invest. 2015;125(11):4160–70. doi:10.1172/JCI82695.
- Ashwell G, Harford J. Carbohydrate-specific receptors of the liver. Annu Rev Biochem. 1982;51(1):531–54. doi:10.1146/annurev.bi.51.070182.002531.
- Orange JS, Hossny EM, Weiler CR, Ballow M, Berger M, Bonilla FA, Buckley R, Chinen J, El-Gamal Y, Bd M. Use of intravenous immunoglobulin in human disease: a review of evidence by members of the primary immunodeficiency committee of the American academy of allergy, asthma and Immunology. J Allergy Clin Immunol. 2006;117:525–53.
- Ballow M. The IgG molecule as a biological immune response modifier: mechanisms of action of intravenous immune serum globulin in autoimmune and inflammatory disorders. J Allergy Clin Immunol. 2011;127(2):315–23. doi:10.1016/j.jaci.2010.10.030.
- Stephen-Victor E, Bayry J. Multimerized IgG1 Fc molecule as an anti-inflammatory agent. Nat Rev Rheumatol. 2018;14(7):390–92. doi:10.1038/s41584-018-0013-9.
- Lünemann JD, Nimmerjahn F, Dalakas MC. Intravenous immunoglobulin in neurology--mode of action and clinical efficacy. Nat Rev Neurol. 2015;11(2):80–89. doi:10.1038/nrneurol.2014.253.
- Schwab I, Nimmerjahn F. Intravenous immunoglobulin therapy: how does IgG modulate the immune system?. Nat Rev Immunol. 2013;13(3):176–89. doi:10.1038/nri3401.
- Shock A, Humphreys D, Nimmerjahn F. Dissecting the mechanism of action of intravenous immunoglobulin in human autoimmune disease: lessons from therapeutic modalities targeting Fcγ receptors. J Allergy Clin Immunol. 2020;146(3):492–500. doi:10.1016/j.jaci.2020.06.036.
- Li D, Lou Y, Zhang Y, Liu S, Li J, Tao J. Sialylated immunoglobulin G: a promising diagnostic and therapeutic strategy for autoimmune diseases. Theranostics. 2021;11(11):5430–46. doi:10.7150/thno.53961.
- Parekh RB, Dwek RA, Sutton BJ, Fernandes DL, Leung A, Stanworth D, Rademacher TW, Mizuochi T, Taniguchi T, Matsuta K. Association of rheumatoid arthritis and primary osteoarthritis with changes in the glycosylation pattern of total serum IgG. Nature. 1985;316(6027):452–57. doi:10.1038/316452a0.
- Scherer HU, Van Der Woude D, Ioan-Facsinay A, El Bannoudi H, Trouw LA, Wang J, Häupl T, Burmester GR, Deelder AM, Huizinga TWJ. Glycan profiling of anti-citrullinated protein antibodies isolated from human serum and synovial fluid. Arthritis Rheum. 2010;62(6):1620–29. doi:10.1002/art.27414.
- Van De Geijn FE, Wuhrer M, Selman MHJ, Willemsen SP, De Man YA, Deelder AM, Hazes JMW, Dolhain RJEM. Immunoglobulin G galactosylation and sialylation are associated with pregnancy-induced improvement of rheumatoid arthritis and the postpartum flare: results from a large prospective cohort study. Arthritis Res Ther. 2009;11(6):1–10. doi:10.1186/ar2892.
- Espy C, Morelle W, Kavian N, Grange P, Goulvestre C, Viallon V, Chéreau C, Pagnoux C, Michalski JC, Guillevin L. Sialylation levels of anti-proteinase 3 antibodies are associated with the activity of granulomatosis with polyangiitis (wegener’s). Arthritis Rheum. 2011;63(7):2105–15. doi:10.1002/art.30362.
- Fokkink W-JR, Selman MHJ, Dortland JR, Durmuş B, Kuitwaard K, Huizinga R, van Rijs W, Tio-Gillen AP, Van Doorn PA, Deelder AM. IgG Fc N-Glycosylation in guillain–barré syndrome treated with immunoglobulins. J Proteome Res. 2014;13(3):1722–30. doi:10.1021/pr401213z.
- Kaneko Y, Nimmerjahn F, Ravetch JV. Anti-inflammatory activity of immunoglobulin G resulting from Fc sialylation. Science. 2006;313(5787):670–73. doi:10.1126/science.1129594.
- Schwab I, Biburger M, Krönke G, Schett G, Nimmerjahn F. IVIg-mediated amelioration of ITP in mice is dependent on sialic acid and SIGNR1. Eur J Immunol. 2012;42:826–30.
- Schwab I, Mihai S, Seeling M, Kasperkiewicz M, Ludwig RJ, Nimmerjahn F. Broad requirement for terminal sialic acid residues and FcγRIIB for the preventive and therapeutic activity of intravenous immunoglobulins in vivo. Eur J Immunol. 2014;44(5):1444–53. doi:10.1002/eji.201344230.
- Arroyo S, Tiessen RG, Denney WS, Jin J, Van Iersel MP (Thijs), Zeitz H, Am M, Mr S, JB B. Hyper-Sialylated IgG M254, an innovative therapeutic candidate, evaluated in healthy volunteers and in patients with immune thrombocytopenia purpura: safety, tolerability, pharmacokinetics, and pharmacodynamics. Blood. 2019;134(Supplement_1):1090. doi:10.1182/blood-2019-125993.
- Dekkers G, Plomp R, Koeleman CAM, Visser R, von Horsten HH, Sandig V, Rispens T, Wuhrer M, Vidarsson G. Multi-level glyco-engineering techniques to generate IgG with defined Fc-glycans. Sci Rep. 2016;6(1):36964. doi:10.1038/srep36964.
- Yu X, Baruah K, Harvey DJ, Vasiljevic S, Alonzi DS, Song BD, Higgins MK, Bowden TA, Scanlan CN, Crispin M. Engineering hydrophobic protein-carbohydrate interactions to fine-tune monoclonal antibodies. J Am Chem Soc. 2013;135(26):9723–32. doi:10.1021/ja4014375.
- Cymer F, Beck H, Rohde A, Reusch D. Therapeutic monoclonal antibody N-glycosylation - structure, function and therapeutic potential. Biologicals. 2018;52:1–11. doi:10.1016/j.biologicals.2017.11.001.
- Makita S, Tobinai K. Mogamulizumab for the treatment of T-cell lymphoma. Expert Opin Biol Ther. 2017;17(9):1145–53. doi:10.1080/14712598.2017.1347634.
- Huang W, Giddens J, Fan SQ, Toonstra C, Wang LX. Chemoenzymatic glycoengineering of intact IgG antibodies for gain of functions. J Am Chem Soc. 2012;134(29):12308–18. doi:10.1021/ja3051266.
- Yu X, Vasiljevic S, Mitchell DA, Crispin M, Scanlan CN. Dissecting the molecular mechanism of IVIg therapy: the interaction between serum IgG and DC-SIGN is independent of antibody glycoform or Fc domain. J Mol Biol. 2013;425(8):1253–58. doi:10.1016/j.jmb.2013.02.006.
- Mahajan VS, Pillai S. Sialic acids and autoimmune disease. Immunol Rev. 2016;269(1):145–61. doi:10.1111/imr.12344.
- Lübbers J, Rodríguez E, Van Kooyk Y. Modulation of immune tolerance via siglec-Sialic Acid interactions. Front Immunol. 2018;9:2807. doi:10.3389/fimmu.2018.02807.
- Czajkowsky DM, Andersen JT, Fuchs A, Wilson TJ, Mekhaiel D, Colonna M, He J, Shao Z, Mitchell DA, Wu G. Developing the IVIG biomimetic, Hexa-Fc, for drug and vaccine applications. Sci Rep. 2015;5(1):9526. doi:10.1038/srep09526.
- Crispin M, Yu X, Bowden TA. Crystal structure of sialylated IgG Fc: implications for the mechanism of intravenous immunoglobulin therapy. Proc Natl Acad Sci. 2013;110(38):3544–46. doi:10.1073/pnas.1310657110.
- Tang F, Wang L-X HW. Chemoenzymatic synthesis of glycoengineered IgG antibodies and glycosite-specific antibody–drug conjugates. Nat Protoc. 2017;12(8):1702–21. doi:10.1038/nprot.2017.058.
- Blundell PA, Le NPL, Allen J, Watanabe Y, Pleass RJ. Engineering the fragment crystallizable (Fc) region of human IgG1 multimers and monomers to fine-tune interactions with sialic acid-dependent receptors. J Biol Chem. 2017;292(31):12994–3007. doi:10.1074/jbc.M117.795047.
- Blundell PA, Lu D, Dell A, Haslam S, Pleass RJ. Choice of host cell line is essential for the functional glycosylation of the Fc Region of human IgG1 inhibitors of influenza B viruses. J Immunol. 2020;204(4):1022–34. doi:10.4049/jimmunol.1901145.
- Blundell PA, Lu D, Wilkinson M, Dell A, Haslam S, Pleass RJ. Insertion of N-terminal hinge glycosylation enhances interactions of the Fc Region of human IgG1 monomers with glycan-dependent receptors and blocks hemagglutination by the influenza virus. J Immunol. 2019;202:1595–611.
- Mekhaiel DNA, Czajkowsky DM, Andersen JT, Shi J, El-Faham M, Doenhoff M, McIntosh RS, Sandlie I, He J, Hu J. Polymeric human Fc-fusion proteins with modified effector functions. Sci Rep. 2011;1:124. 10.1038/srep00124
- Böhm M, Bohne-Lang A, Frank M, Loss A, Rojas-Macias MA, Lütteke T. Glycosciences.DB: an annotated data collection linking glycomics and proteomics data (2018 update). Nucleic Acids Res. 2019;47(D1):D1195–201. doi:10.1093/nar/gky994.
- Baksmeier C, Blundell P, Steckel J, Schultz V, Gu Q, Da Silva Filipe A, Kohl A, Linnington C, Lu D, Dell A. Modified recombinant human IgG1-Fc is superior to natural IVIG at inhibiting immune-mediated demyelination. Immunology. 2021;00:1–16.
- Tradtrantip L, Felix CM, Spirig R, Morelli AB, Verkman AS. Recombinant IgG1 Fc hexamers block cytotoxicity and pathological changes in experimental in vitro and rat models of neuromyelitis optica. Neuropharmacology. 2018;133:345–53. doi:10.1016/j.neuropharm.2018.02.002.
- Spirig R, Campbell IK, Koernig S, Chen C-G, Lewis BJB, Butcher R, Muir I, Taylor S, Chia J, Leong D. rIgG1 Fc hexamer inhibits antibody-mediated autoimmune disease via effects on complement and FcγRs. J Immunol. 2018;200(8):2542–53. doi:10.4049/jimmunol.1701171.
- Zuercher AW, Spirig R, Baz Morelli A, Rowe T, Käsermann F. Next-generation Fc receptor–targeting biologics for autoimmune diseases. Autoimmun Rev. 2019;18(10):102366. doi:10.1016/j.autrev.2019.102366.
- Karsten CM, Pandey MK, Figge J, Kilchenstein R, Taylor PR, Rosas M, McDonald JU, Orr SJ, Berger M, Petzold D. Anti-inflammatory activity of IgG1 mediated by Fc galactosylation and association of FcγRIIB and dectin-1. Nat Med. 2012;18(9):1401–06. doi:10.1038/nm.2862.
- Lux A, Yu X, Scanlan CN, Nimmerjahn F. Impact of immune complex size and glycosylation on IgG binding to human FcγRs. J Immunol. 2013;190(8):4315–23. doi:10.4049/jimmunol.1200501.
- Campbell IK, Miescher S, Branch DR, Mott PJ, Lazarus AH, Han D, Maraskovsky E, Zuercher AW, Neschadim A, Leontyev D. Therapeutic effect of IVIG on inflammatory arthritis in mice is dependent on the Fc portion and independent of sialylation or basophils. J Immunol. 2014;192(11):5031–38. doi:10.4049/jimmunol.1301611.
- JM F, Kr A, Rp B, Ha H, EE W, Wa B. Antibody blood-brain barrier efflux is modulated by glycan modification. Biochim Biophys Acta - Gen Subj. 2017;1861(9):2228–39. doi:10.1016/j.bbagen.2017.06.008.
- Zhang G, Massaad CA, Gao T, Pillai L, Bogdanova N, Ghauri S, Sheikh KA. Sialylated intravenous immunoglobulin suppress anti-ganglioside antibody mediated nerve injury. Exp Neurol. 2016;282:49–55. doi:10.1017/j.expneurol.2016.05.020.
- MacAuley MS, Crocker PR, Paulson JC. Siglec-mediated regulation of immune cell function in disease. Nat Rev Immunol. 2014;14(10):653–66. doi:10.1038/nri3737.
- Movsisyan LD, Macauley MS. Structural advances of siglecs: insight into synthetic glycan ligands for immunomodulation. Org Biomol Chem. 2020;18(30):5784–97. doi:10.1039/D0OB01116A.
- Crocker PR, Redelinghuys P. Siglecs as positive and negative regulators of the immune system. Biochem Soc Trans. 2008;36(6):1467–71. doi:10.1042/BST0361467.
- Delaveris CS, Chiu SH, Riley NM, Bertozzi CR. Modulation of immune cell reactivity with cis-binding siglec agonists. Proc Natl Acad Sci U S A. 2021;118(3):e2012408118. doi: 10.1073/pnas.2012408118
- Bondioli L, Ruozi B, Belletti D, Forni F, Vandelli MA, Tosi G. Sialic acid as a potential approach for the protection and targeting of nanocarriers. Expert Opin Drug Deliv. 2011;8(7):921–37. doi:10.1517/17425247.2011.577061.
- Courtney AH, Puffer EB, Pontrello JK, Yang ZQ, Kiessling LL. Sialylated multivalent antigens engage CD22 in trans and inhibit B cell activation. Proc Natl Acad Sci U S A. 2009;(8). doi:10.1073/pnas.0807207106.
- Rillahan CD, Schwartz E, McBride R, Fokin VV, Paulson JC. Click and pick: identification of sialoside analogues for siglec-based cell targeting. Angew Chemie. 2012;51(44):11014–18. doi:10.1002/anie.201205831.
- Flynn RA, Pedram K, Malaker SA, Batista PJ, Smith BAH, Johnson AG, George BM, Majzoub K, Villalta PW, Carette JE. Small RNAs are modified with N-glycans and displayed on the surface of living cells. Cell. 2021;(12). doi:10.1016/j.cell.2021.04.023.
- Spence S, Greene MK, Fay F, Hams E, Saunders SP, Hamid U, Fitzgerald M, Beck J, Bains BK, Smyth P. Targeting siglecs with a sialic acid-decorated nanoparticle abrogates inflammation. Sci Transl Med. 2015;7(303):1–13. doi:10.1126/scitranslmed.aab3459.
- Kilcoyne M, Joshi L. Translational Glycobiology in Human Health and Disease. Academic Press,London; 2020. ISBN 9780128196557.
- Miles LA, Hermans SJ, Crespi GAN, Gooi JH, Doughty L, Nero TL, Markulić J, Ebneth A, Wroblowski B, Oehlrich D. Small molecule binding to alzheimer risk factor CD33 promotes Aβ phagocytosis. iScience. 2019;19:110–18. doi:10.1016/j.isci.2019.07.023.
- Lajaunias F, Dayer JM, Chizzolini C. Constitutive repressor activity of CD33 on human monocytes requires sialic acid recognition and phosphoinositide 3-kinase-mediated intracellular signaling. Eur J Immunol. 2005;35(1):243–51. doi:10.1002/eji.200425273.
- Waldmann M, Jirmann R, Hoelscher K, Wienke M, Niemeyer FC, Rehders D, Meyer B. A nanomolar multivalent ligand as entry inhibitor of the hemagglutinin of avian influenza. J Am Chem Soc. 2014;136(2):783–88. doi:10.1021/ja410918a.
- Cartellieri M, Feldmann A, Koristka S, Arndt C, Loff S, Ehninger A, von Bonin M, Bejestani EP, Ehninger G, Bachmann MP. Switching CAR T cells on and off: a novel modular platform for retargeting of T cells to AML blasts. Blood Cancer J. 2016;6(8):e458. doi:10.1038/bcj.2016.61.
- Estus S, Bc S, Devanney N, Katsumata Y, EE P, DW F. Evaluation of CD33 as a genetic risk factor for alzheimer’s disease. Acta Neuropathol. 2019;138:187–99.
- Murch SH. Common determinants of severe covid-19 infection are explicable by SARS-CoV-2 secreted glycoprotein interaction with the CD33-related siglecs, siglec-3 and siglec-5/14. Med Hypotheses. 2020;144:110168. doi:10.1016/j.mehy.2020.110168.
- Rodrigues E, Jung J, Park H, Loo C, Soukhtehzari S, Kitova EN, Mozaneh F, Daskhan G, Schmidt EN, Aghanya V. A versatile soluble siglec scaffold for sensitive and quantitative detection of glycan ligands. Nat Commun. 2020;11(1):5091. doi:10.1038/s41467-020-18907-6.
- Blundell P, Pleass R. A method to detect the binding of hyper-glycosylated fragment crystallizable (Fc) region of human IgG1 to glycan receptors. Methods Mol Biol. 2019;1904:417–21.
- Matrosovich M, Herrler G, Klenk HD. Sialic acid receptors of viruses. SialoGlyco Chem Biol II. 2015;367:1–28.
- Stencel-Baerenwald JE, Reiss K, Reiter DM, Stehle T, Dermody TS. The sweet spot: defining virus-sialic acid interactions. Nat Rev Microbiol. 2014;12(11):739–49. doi:10.1038/nrmicro3346.
- Vahey MD, Fletcher DA. Influenza A virus surface proteins are organized to help penetrate host mucus. Elife. 2019;8:e43764. doi:10.7554/eLife.43764.
- Jj S, Dc W. Receptor binding and membrane fusion in virus entry: the influenza hemagglutinin. Annu Rev Biochem. 2000;69(1):531–69. doi:10.1146/annurev.biochem.69.1.531.
- Bandlow V, Lauster D, Ludwig K, Hilsch M, Reiter-Scherer V, Rabe JP, Böttcher C, Herrmann A, Seitz O. Sialyl-LacNAc-PNA⋅DNA concatamers by rolling-circle amplification as multivalent inhibitors of influenza A virus particles. ChemBioChem. 2019;20:1–8.
- Bandlow V, Liese S, Lauster D, Ludwig K, Netz RR, Herrmann A, Seitz O. Spatial screening of hemagglutinin on influenza A virus particles: sialyl-LacNAc displays on DNA and PEG scaffolds reveal the requirements for bivalency enhanced interactions with weak monovalent binders. J Am Chem Soc. 2017;139(45):16389–97. doi:10.1021/jacs.7b09967.
- Tang S, Puryear WB, Seifried BM, Dong X, Runstadler JA, Ribbeck K, Olsen BD. Antiviral agents from multivalent presentation of sialyl oligosaccharides on brush polymers. ACS Macro Lett. 2016;5(3):413–18. doi:10.1021/acsmacrolett.5b00917.
- Kwon S-J, Na DH, Kwak JH, Douaisi M, Zhang F, Park EJ, Park J-H, Youn H, Song C-S, Kane RS. Nanostructured glycan architecture is important in the inhibition of influenza A virus infection. Nat Nanotechnol. 2017;12(1):48–54. doi:10.1038/nnano.2016.181.
- Papp I, Sieben C, Sisson AL, Kostka J, Böttcher C, Ludwig K, Herrmann A, Haag R. Inhibition of influenza virus activity by multivalent glycoarchitectures with matched sizes. ChemBioChem. 2011;12(6):887–95. doi:10.1002/cbic.201000776.
- Sigal GB, Mammen M, Dahmann G, Whitesides GM. Polyacrylamides bearing pendant α-sialoside groups strongly inhibit agglutination of erythrocytes by influenza virus: the strong inhibition reflects enhanced binding through cooperative polyvalent interactions. J Am Chem Soc. 1996;118(16):3789–800. doi:10.1021/ja953729u.
- Weissleder R, Kelly K, Sun EY, Shtatland T, Josephson L. Cell-specific targeting of nanoparticles by multivalent attachment of small molecules. Nat Biotechnol. 2005;23(11):1418–23. doi:10.1038/nbt1159.
- Delaveris CS, Webster ER, Banik SM, Boxer SG, Bertozzi CR. Membrane-tethered mucin-like polypeptides sterically inhibit binding and slow fusion kinetics of influenza A virus. Proc Natl Acad Sci U S A. 2020;117(23):12643–50. doi:10.1073/pnas.1921962117.
- Ghosh S. Nanotechnology and sialic acid biology. Academic Press, London; 2020. ISBN 9780128161265.
- Nel A, Xia T, Mädler L, Li N. Toxic potential of materials at the nanolevel. Science. 2006;311(5761):622–27. doi:10.1126/science.1114397.
- Rathor S, Bhatt DC, Aamir S, Singh SK, Kumar V. A comprehensive review on role of nanoparticles in therapeutic delivery of medicine. Pharm Nanotechnol. 2017;5:263–75.
- Xu R, Ekiert DC, Krause JC, Hai R, Crowe JE, Wilson IA. Structural basis of preexisting immunity to the 2009 H1N1 pandemic influenza virus. Science. 2010;328(5976):357–60. doi:10.1126/science.1186430.
- Wang Q, Cheng F, Lu M, Tian X, Ma J. Crystal structure of unliganded influenza B virus hemagglutinin. J Virol. 2008;82(6):3011–20. doi:10.1128/JVI.02477-07.
- Ahmed AA, Giddens J, Pincetic A, Lomino JV, Ravetch JV, Wang LX, Bjorkman PJ. Structural characterization of anti-inflammatory immunoglobulin G Fc proteins. J Mol Biol. 2014;426(18):3166–79. doi:10.1016/j.jmb.2014.07.006.
- Liu Q, Zhou YH, Yang ZQ. The cytokine storm of severe influenza and development of immunomodulatory therapy. Cell Mol Immunol. 2016;13(1):3–10. doi:10.1038/cmi.2015.74.
- Maurer MA, Meyer L, Bianchi M, Turner HL, Le NPL, Steck M, Wyrzucki A, Orlowski V, Ward AB, Crispin M. Glycosylation of human IgA directly inhibits influenza A and other Sialic-Acid-binding viruses. Cell Rep. 2018;23(1):90–99. doi:10.1016/j.celrep.2018.03.027.
- Sandin S, Öfverstedt LG, Wikström AC, Ö W, Skoglund U. Structure and flexibility of individual immunoglobulin G molecules in solution. Structure. 2004;12(3):409–15. doi:10.1016/j.str.2004.02.011.
- Weis W, Jh B, Cusack S, JC P, Jj S, Dc W. Structure of the influenza virus haemagglutinin complexed with its receptor, sialic acid. Nature. 1988;333(6172):426–31. doi:10.1038/333426a0.
- Hong M, Lee PS, Hoffman RMB, Zhu X, Krause JC, Laursen NS, Yoon S-I, Song L, Tussey L, JE C. Antibody recognition of the pandemic H1N1 influenza virus hemagglutinin receptor binding site. J Virol. 2013;87(22):12471–80. doi:10.1128/JVI.01388-13.
- Suzuki Y, Nei M. Origin and evolution of influenza virus hemagglutinin genes. Mol Biol Evol. 2002;19(4):501–09. doi:10.1093/oxfordjournals.molbev.a004105.
- Ernst B, Magnani JL. From carbohydrate leads to glycomimetic drugs. Nat Rev Drug Discov. 2009;8(8):661–77. doi:10.1038/nrd2852.
- Gao Z, Niikura M, Withers SG. Ultrasensitive fluorogenic reagents for neuraminidase titration. Angew Chemie - Int Ed. 2017;56(22):6112–16. doi:10.1002/anie.201610544.
- Roy MG, Livraghi-Butrico A, Fletcher AA, McElwee MM, Evans SE, Boerner RM, Alexander SN, Bellinghausen LK, Song AS, Petrova YM. Muc5b is required for airway defence. Nature. 2014;505(7483):412–16. doi:10.1038/nature12807.
- Zanin M, Baviskar P, Webster R, Webby R. The interaction between respiratory pathogens and mucus. Cell Host Microbe. 2016;19(2):159–68. doi:10.1016/j.chom.2016.01.001.
- Bansil R, Turner BS. The biology of mucus: composition, synthesis and organization. Adv Drug Deliv Rev. 2018;124:3–15. doi:10.1016/j.addr.2017.09.023.
- Alejandra Tortorici M, Walls AC, Lang Y, Wang C, Li Z, Koerhuis D, Boons GJ, Bosch BJ, Rey FA, de Groot RJ. Structural basis for human coronavirus attachment to sialic acid receptors. Nat Struct Mol Biol. 2019;26(6):481–89. doi:10.1038/s41594-019-0233-y.
- Chambers BS, Li Y, Hodinka RL, Hensley SE. Recent H3N2 influenza virus clinical isolates rapidly acquire hemagglutinin or neuraminidase mutations when propagated for antigenic analyses. J Virol. 2014;88(18):10986–89. doi:10.1128/JVI.01077-14.
- Peng W, De Vries RP, Grant OC, Thompson AJ, McBride R, Tsogtbaatar B, Lee PS, Razi N, Wilson IA, Woods RJ. Recent H3N2 viruses have evolved specificity for extended, branched human-type receptors, conferring potential for increased avidity. Cell Host Microbe. 2017;21(1):23–34. doi:10.1016/j.chom.2016.11.004.
- Broszeit F, van Beek RJ, Unione L, Bestebroer TM, Chapla D, Yang JY, Moremen KW, Herfst S, Fouchier RAM, De Vries RP. Glycan remodeled erythrocytes facilitate antigenic characterization of recent A/H3N2 influenza viruses. bioRxiv. 2020. doi.10.1101/2020.12.18.423398
- Trichonas G, Kaiser PK. Aflibercept for the treatment of age-related macular degeneration. Ophthalmol Ther. 2013;2(2):89–98. doi:10.1007/s40123-013-0015-2.
- Faghihi H, Najafabadi AR, Daman Z, Ghasemian E, Montazeri H, Vatanara A. Respiratory administration of infliximab dry powder for local suppression of inflammation. AAPS PharmSciTech. 2019;20(3):128. doi:10.1208/s12249-019-1308-0.