
ABSTRACT
Bispecific antibodies have recently attracted intense interest. CrossMab technology was described in 2011 as novel approach enabling correct antibody light-chain association with their respective heavy chain in bispecific antibodies, together with methods enabling correct heavy-chain association using existing pairs of antibodies. Since the original description, CrossMab technology has evolved in the past decade into one of the most mature, versatile, and broadly applied technologies in the field, and nearly 20 bispecific antibodies based on CrossMab technology developed by Roche and others have entered clinical trials. The most advanced of these are the Ang-2/VEGF bispecific antibody faricimab, currently undergoing regulatory review, and the CD20/CD3 T cell bispecific antibody glofitamab, currently in pivotal Phase 3 trials. In this review, we introduce the principles of CrossMab technology, including its application for the generation of bi-/multispecific antibodies with different geometries and mechanisms of action, and provide an overview of CrossMab-based therapeutics in clinical trials.
Introduction into bispecific antibodies
Antibodies, or so-called immunoglobulins, are Y-shaped proteins of ca. 150 kDa generated by B and plasma cells of the immune system as response to infection. They consist of two identical heavy and two identical light chains forming: 1) two variable antigen-binding sites within the antigen-binding fragments (Fabs) that serve for the specific recognition of (foreign) antigens; and 2) a constant Fc domain that serves for the recruitment of the human immune system.
Recombinant antibodies have been used therapeutically for over 30 years, and today more than 120 therapeutic antibodies are approved or under regulatory review by health authorities for use in humans (source: https://www.antibodysociety.org/resources/approved-antibodies/). Since the advent of recombinant antibody technologies, there has been substantial interest in the generation of engineered and bispecific antibodies that are characterized by having two independent specificities in the Fabs, resulting in novel mechanisms of action that typically cannot be achieved with conventional monospecific antibodies. More than 100 bispecific antibodies are currently being tested in clinical trials.Citation1–6
While uncountable approaches for the generation of bispecific antibodies have been described, Citation1–6 some of the most broadly applied technologies for the generation of bispecific antibodies include ART-Ig,Citation7–10 BEAT,Citation11 BiTE,Citation12,Citation13 common light chains,Citation9,Citation10,Citation14–16 DAF,Citation17 DART,Citation18 DuoBody,Citation19 DutaFab,Citation20 DVD-Ig,Citation21 Fab arm exchange,Citation22 Fcab,Citation23–25 FORCE,Citation26 half antibody assembly,Citation27 Hetero-Ig,Citation28,Citation29 IgG-scFv,Citation30Citation31,Citation131κλ-bodies,Citation32 Multiclonics,Citation14 orthogonal Fab interface,Citation33 Tandab,Citation34 XmAb,Citation35 VELOCI-Bi,Citation15 and WuxiBODY.Citation36
As of August 2021, three bispecific antibodies have been approved, the tandem single-chain variable fragment (Fv)-based CD19/CD3 Bispecific T-cell Engager (BiTE) blinatumomab developed by Amgen for the treatment of acute lymphocytic leukemia (ALL),Citation37 the heterodimeric ART-Ig-based coagulation factor IX/X bispecific IgG antibody emicizumab developed by Chugai and Roche for the treatment of hemophilia A,Citation7,Citation8,Citation10 and the heterodimeric DuoBody-based EGFR/c-Met bispecific IgG antibody amivantamab developed by Janssen for treatment of non-small cell lung cancer harboring EGFR exon 20 insertion mutations.Citation38–40
CrossMab technology for the generation of bispecific antibodies
We have developed an alternative technology, known as CrossMab technology, which together with methods enabling correct heavy-chain association such as the so-called knobs-into-holes technology (KiH),Citation16 that enables the correct association of the different antibody light chains with their respective counterparts. This is achieved in different antibody formats and geometries by the exchange or crossover of antibody domains.Citation41–43 Here, we give a brief overview of the basic principles of CrossMab technology and its application for the generation of various CrossMabs with different molecular formats and mechanisms of action.Citation42,Citation44,Citation45 In fact, back in 2011, this approach was the first technology described allowing the conversion of two pre-existing antibodies into heterodimeric bispecific antibodies of the bivalent IgG format without the need to rely on so-called common light-chain antibodies that have identical light chains in each Fab.Citation42
Since in bispecific antibodies the two heavy chains as well as the two light chains are different and can randomly associate, expression of these four chains leads to the formation of ten different antibody variants.Citation46 Correct heavy-chain association resulting in a heterodimeric Fc can be enforced using KiH technology by introducing a bulky tryptophan (Trp) residue in one Fc fragment and forming a corresponding cavity on the other Fc fragment that can accommodate the Trp residue.Citation16,Citation47,Citation48 More recently, multiple alternative approaches to enable correct heavy-chain association have been described, such as relying on charge interactions.Citation7–11,Citation14,Citation29,Citation49
Although KiH technology was developed in the late 1990s,Citation16 enabling correct light-chain association remained a major problem, and the only approach to achieve this at the time relied on the use of common light chains for both specificities.Citation9,Citation10,Citation14–16 However, the use of a common light chain requires the de novo identification of the corresponding antibody pairs, which can be challenging and/or time-consuming depending on the desired target, and restricts the availability and diversity of antibodies that can be used; thus, methods allowing the generation of bispecific antibodies from pre-existing antibody pairs were highly desired.
shows the basic principle of the domain crossover applied in CrossMab technology to enable correct light-chain association in bispecific antibodies.Citation41 By incorporating the original heavy chain VH-CH1 domains in the Fab of the second specificity of the bispecific antibody as the novel “light chain” and the original light chain VL-CL domains for the novel “heavy chain” by fusing them to the hinge region of the Fc fragment, correct light-chain association can be enforced in the CrossMabFab format. This format has recently also been described as Fabs-in-Tandem Ig (FIT-Ig).Citation50,Citation51
Figure 1. Principles of CrossMab technology: The four major CrossMab formats as applied to 1 + 1 heterodimeric bispecific antibodies are depicted as well as potential side products. On the bottom, the structure of mono- and duomabs is indicated. Heavy-chain domains are depicted in dark colors and respective light-chain domains are depicted with corresponding bright colors. Created with BioRender.com
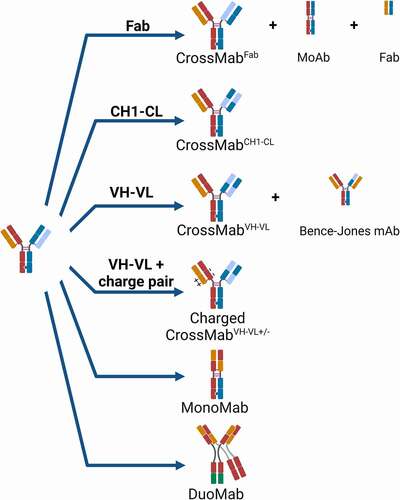
Alternatively, only the VH-VL or only the CH1-CL domains can be exchanged in the CrossMabVH-VL and CrossMabCHCitation1−CL formats (). In the case of the CH1-CL crossover, no theoretical side product due to domain crossover is expected and crystal structure analysis confirmed the structural integrity of the crossed Fab domain in the CrossMabCHCitation11−CL format.Citation52 In the case of the Fab crossover in the CrossMabFab, two heavy and light chain-based monovalent side products can be observed. However, the correct preferential formation of the CrossMabFab can be fostered by relative over-expression of the respective light chains so that the respective undesired monovalent and binding inactive side products do not form in significant amounts. Similarly, in the case of VH-VL crossover, a Bence-Jones-like side product based on VL-VL together with the CH1-CL interaction can be observed. In order to avoid formation of this side product, natural charge pairs in the Fab were identified and the respective orthogonal charge interactions were introduced into the non-exchanged antibody CH1-CL domains.Citation53 As a consequence, the undesired Bence-Jones-like side product does not form due to repulsive charge interactions, whereas the desired light-chain pairs correctly in the non-crossed Fab due to attractive charge interactions so that the corresponding CrossMabVH-VL± constructs can subsequently be obtained in high yields and purity without major side products.
Notably, these design principles can be applied not only to heterodimeric antibodies where one arm is directed to the first antigen, the other arm to the second antigen (1 + 1 format), but the CrossMab technology also allows generation of so-called MonoMabs, monovalent antibodies with one Fc portion, and DuoMabs, bivalent antibodies with two Fc portions ().Citation54 Furthermore, it can be applied to enable the correct light-chain association in hetero-/homodimeric bi-/multispecific antibody appended or tandem-Fab formats with, for example, 2 + 1, 2 + 2, 3 + 1, 4 + 1 or 4 + 2 valencies and in antibody fusion proteins ().Citation44,Citation45 In line with this, Wu and colleagues from Lilly applied Fab crossover to generate orthogonal Fab-based trispecific antibody formats termed “OrthoTsAbs”.Citation55,Citation56 Interestingly, domain crossover has also been described as a means to prevent mispairing of T-cell receptor (TCR) domains in adoptive T-cell therapy.Citation57
Figure 2. Major CrossMab formats: A) 1 + 1 CrossMab:CHCitation1−CL vanucizumab, faricimab, 10E8.4/iMab 1 + 1; B) 1 + 1 CrossMabVH-VL±: PD1-TIM3, PD1-LAG3; C) CrossMabCHCitation1−CL+/–based FAP-4-1BBL, CD19-4-1BBL fusion proteins; D) 2 + 1 CrossMab:CHCitation1−CL cibisatamab; E) 2 + 1 CrossMabVH-VL±: glofitamab, CC-93269, TYRP1-TCB, WT1-TCB, RG6123; F) 2 + 2 CrossMabCHCitation1−CL-based FIT-Ig EMB-01, EMB-02, EMB-06; G) 2 + 2 CrossMab:CHCitation1−CL FAP-DR5; H) 2 + 1 CrossMab VH-VL±: BS-GANT, FAP-CD40; I) 1 + 1 CrossMabCHCitation1−CL-based Nkp46-based NK cell engager (NKCE). Heavy-chain domains are depicted in dark colors and respective light-chain domains are depicted with corresponding bright colors. Fusion protein depicted in purple. Note: Differences in variable regions and/or isotype and Fc engineering are not depicted. Created with BioRender.com
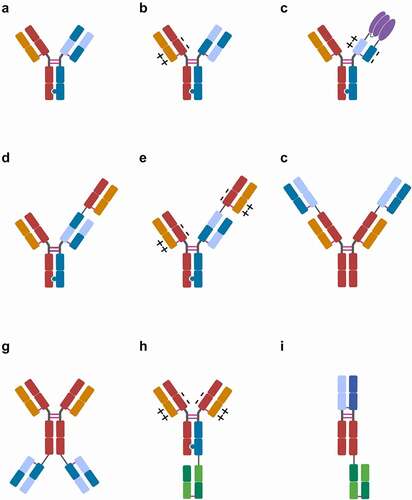
Because the CrossMab approach showed advantages in terms of production, stability, developability, and versatility over analogous formats based on either single-chain FvCitation58–60 or single-chain FabCitation61–67 building blocks, it was ultimately chosen as the antibody engineering approach of choice for the generation of various clinical development candidates. Obviously, in order to develop CrossMabs for therapeutic use, the establishment of various methods covering CMC (Chemistry, Manufacturing, and Controls) aspects, including upstream and downstream processing (USP, DSP) and the establishment of the respective bioassay and (bio-) analytical methods was and is essential.Citation68–80Citation81 When considering the formation of (undesired) side products, it has to be taken into account that, independent of CrossMab technology, other unrelated side products can occur, such as half- or ¾-antibodies missing two or one light chains, respectively, or hole-hole/knob-knob heavy-chain homodimers. In order to avoid the formation of these side products, achieving equal expression levels for the four heavy and light chains during transient expression and/or stable cell line generation by selecting suitable clones is advantageous. Based on the general advancement in the field of therapeutic antibody manufacturing, as well as considering these specific learnings, bispecific antibodies of different formats based on CrossMab technology can generally be manufactured in a consistent and reproducible fashion with volumetric yields in the several g/L range and in quality comparable to conventional therapeutic antibodies using established USP and DSP platforms.
Consequently, since the original description of this concept, the technology has evolved in the past decade into one of the most mature, versatile, and broadly applied technologies in the field for the generation of various bispecific antibody formats. As of mid-2021, at least 19 bispecific antibodies and fusion proteins based on CrossMab technology developed by Roche and others have entered clinical trials, of which 16 continue to be evaluated in active clinical trials ( and ). In the following sections, an overview of therapeutic bispecific antibodies and fusion proteins based on CrossMab technology is provided, with a focus on those in clinical trials.
Table 1. CrossMabs in clinical trials (status July 2021), FP: Fusion protein, FIT-Ig: Fabs-in-tandem Ig, EIH: Entry into human date
Applications in targeted cancer therapy: Angiogenesis, receptor tyrosine kinases, and death receptor signaling
For many years, anti-angiogenesis approaches blocking the vascular endothelial growth factor-A (VEGF-A) have been a major area of targeted cancer therapy.Citation95–Citation96 One of the first IgG-based antibodies and the first bispecific CrossMab to enter clinical trials, in 2012, was the heterodimeric 1 + 1 VEGF/Ang-2 CrossMabCHCitation1−CL vanucizumab (RG7221) () targeting the pro-angiogenic ligands VEGF-A and angiopoietin-2 (Ang-2), which are involved in (tumor) angiogenesis.Citation95,Citation96,Citation97 Vanucizumab, as well as a mouse-specific surrogate bispecific, mediated potent anti-tumoral and anti-angiogenic efficacy in various preclinical models as monotherapy and in combination with chemotherapy,Citation81,Citation98–101 as well as combined with PD-1 checkpoint inhibitionCitation102–104 and CD40 agonism.Citation105,Citation106 Vanucizumab was generally well tolerated as a monotherapy in a Phase 1 clinical trial and demonstrated promising anti-tumor efficacy, IgG-like pharmacokinetics and low immunogenicity,Citation107 as well as the anticipated pharmacodynamic mechanism of action.Citation108 Based on the negative outcome of the randomized McCAVE Phase 2 study, where it was compared to bevacizumab in combination with FOLFOX-6 chemotherapy in patients with untreated metastatic colorectal carcinoma, clinical development was discontinued.Citation109 Similarly, in spite of promising preclinical data, Phase 1b studies of vanucizumab in combination with the PD-L1 antibody atezolizumab (NCT01688206) and the CD40 antibody selicrelumab (NCT02665416) were ultimately discontinued. Recently, preclinical data demonstrated that dual inhibition of VEGF and Ang-2 by the vanucizumab mouse-specific surrogate bispecific in murine sepsis models improved the outcomes, making it a potential therapeutic against vascular barrier breakdown.Citation110 Similarly, Zhou and colleagues reported on an alternative anti-angiogenic approach for cancer therapy using a heterodimeric 1 + 1 VEGF/DLL4 CrossMabCHCitation1−CL called HB-32 that mediated potent anti-angiogenic activity in vitro, as well as in vivo anti-tumor activity in breast cancer xenograft models.Citation111
In addition to anti-angiogenesis, targeting receptor tyrosine kinases (RTKs) like EGFR, HER2 or c-Met has been a major area for cancer therapy during the past decades.Citation112 Accordingly, various preclinical-stage bispecific CrossMabs targeting RTKs have been developed during the past years, but none of these have advanced to clinical trials so far. Zhang and colleagues created a biparatopic HER2/HER2 1 + 1 CrossMabFab based on trastuzumab and an avidity-improved variant L56TY derived from pertuzumab called Tras-Permut CrossMab. Tras-Permut CrossMab mediated improved activity against trastuzumab-resistant breast cancer and enhanced calreticulin exposure, which may contribute to the induction of tumor-specific T-cell responses.Citation113 Lu and colleagues, in turn, generated a bispecific HER2/EGFR 1 + 1 CrossMabCHCitation1−CL based on the trastuzumab and cetuximab.Citation72 Interestingly, Hu and colleagues went a step further and generated so-called four-in-one antibodies that exhibited four different specificities against EGFR, HER2, HER3, and VEGF by generating a 1 + 1 CrossMAbCHCitation1−CL using dual-acting Fabs (DAF) as building blocks in the FL518 bispecific or by combining CrossMab and DVD-Ig technology in the tetraspecific, tetravalent antibody CRTB6 to enable correct light-chain association in the DVD format.Citation114 Not surprisingly, these tetraspecific antibodies showed superior efficacy as compared to the respective bispecific antibodies.Citation114 Finally, different bispecific EGFR/Notch CrossMabs were described to block EGFR signaling together with Notch signaling. The first of these antibodies, termed CT16, combined the EGFR antibody cetuximab and the Notch 2/3 antibody tarextumab using the prototypical heterodimeric 1 + 1 CrossMabCHCitation11−CL format, which served as a radiosensitizer and prevented acquisition of resistance to EGFR inhibitors and radiation in cell line models of non–small cell lung cancer and patient-derived xenograft tumors.Citation115 In a second publication from the same group, three heterodimeric bispecific 1 + 1 CrossMabCHCitation1−CL antibodies (PTG12, RTB3, MTJ16) were generated from panitumumab/tarextumab, RG7116/tarextumab, and MEHD7945A/tarextumab and were shown to increase the response to PI3K inhibition with GDC-0941 by inhibiting stem cell–like subpopulation, reducing tumor-initiating cell frequency, and downregulating mesenchymal gene expression.Citation116
Another major field in targeted cancer therapy has been and continues to be apoptosis induction through death receptor (DR) signaling.Citation117,Citation118 As conventional DR5 antibodies have not been successful in clinical trials, approaches for tumor-targeted DR5 agonism have been pursued. Expression of the fibroblast activation protein (FAP) on tumor fibroblasts is found in the majority of solid tumors, making FAP an attractive antigen for tumor targeting.Citation119,Citation120 Based on this rationale, FAP-targeted bispecific antibodies and fusion proteins have been created using CrossMab technology that rely on FAP binding with one moiety to induce, with their second moiety, hyper-clustering of tumor necrosis factor (TNF) receptor superfamily membersCitation121 like DR5 for apoptosis induction,Citation85 4–1BB/CD137 for T cell activation,Citation89 or CD40 for activation of antigen-presenting cells,Citation94,Citation122 as described below. The first of these conditional FAP-targeted TNFR agonistic antibodies entering Phase 1 clinical trials was the symmetric tetravalent C-terminally fused FAP/DR5 targeted 2 + 2 CrossMabCHCitation1−CL RG7386 (). Preclinical data demonstrated that RG7386 effectively triggered FAP-dependent, avidity-driven DR5 hyper-clustering and subsequent tumor cell apoptosis,Citation85 but ultimately, clinical development of RG7383 was not further continued after the completed Phase 1 study (NCT02558140) due to portfolio reprioritization.
Finally, Tung and colleagues described novel HER2 or CD19 tumor-targeted heterodimeric 1 + 1 CrossMabCHCitation1−CL antibodies that recognize with their second specificity PEGylated proteins, liposomes, and nanoparticles. Using these bispecific antibodies, cytotoxic cargo such as PEGylated liposomal doxorubicin can be delivered to tumor cells.Citation123
Applications in cancer immunotherapy: Dual checkpoint inhibitors, T and innate cell engaging bispecifics and tumor-targeted co-stimulation
With the advent of cancer immunotherapy and checkpoint inhibitor antibodies during the past decade, the development of bispecific antibodies for immunotherapy has attracted substantial attention in industry and academia, whereas the interest in anti-angiogenic and pro-apoptotic therapies has declined. In this context, bispecific monovalent dual checkpoint inhibitory PD-1 antibodies co-targeting the checkpoint inhibitory receptors TIM-3 or LAG-3 have been designed based on a bispecific 1 + 1 CrossMabVH-VL± format (), allowing avidity-mediated selectivity gain and thus enhanced selectivity for PD-1+ and PD-1+ TIM-3+/LAG-3+ double-positive T cells. Both of these bispecific dual checkpoint inhibitory antibodies, PD1-TIM3 (RG7769) Citation87 and PD1-LAG3 (RG6139),Citation91 are currently in Phase 1 and 2 clinical trials (NCT03708328, NCT04140500, NCT04785820).Citation124,Citation125 Preclinically, a heterodimeric 1 + 1 PD-1/RANKL CrossMabCHCitation1−CL was shown to demonstrate potent tumor growth inhibition as a monotherapy and combined with CTLA-4 antibodies, particularly in models showing checkpoint inhibitor resistance to PD-1 antibodies.Citation126
Many of bispecific antibodies currently being developed are bispecific T-cell engagers.Citation22,Citation127–131 One of the first IgG-based, and Roche’s first, T-cell bispecific antibody (TCB) to enter clinical trials was the heterodimeric and trivalent CEA/CD3ε 2 + 1 TCB cibisatamab (RG7802). It is a heterodimeric CEA/CD3ε bispecific antibody in the 1 + 1 CrossMabCHCitation1−CL format to which a single additional Fab targeting CEA is fused to the N-terminus of the knob-containing heavy chain ().Citation84,Citation132 FcγR and C1q binding are abolished by introduction of P329G LALA mutations.Citation133 This so-called 2 + 1 TCB format provides advantages over conventional heterodimeric 1 + 1 TCB formats through the highly flexible head-to-tail fusion in the tandem Fab arm and by being bivalent for the tumor antigen, allowing a better differentiation between tumor and normal cells due to avidity-mediated affinity tuning.Citation132 Cibisatamab demonstrated tumor targeting and in vitro and in vivo anti-tumor efficacy dependent on CEA over-expression due to the bivalent binding mode in models of colorectal and gastric cancer,Citation84,Citation134–137 which was further enhanced when combined with PD-L1 inhibition.Citation138 Based on these data and using a MABEL approach due to the lack of cross-reactive toxicology species,Citation139,Citation140 clinical studies were initiated in relapsed/refractory CEA-positive colorectal cancer patients. Cibisatamab is currently in Phase 1b clinical trials in combination with the PD-L1 antibody atezolizumab (NCT03866239) and with FAP-4-1-BBL (NCT04826003) (see below). Pre-treatment with obinutuzumab is being clinically explored to mitigate the potential development and impact of anti-drug antibodies that could be observed in patients treated with cibisatamab.
The most advanced 2 + 1 T cell bispecific antibody is glofitamab (RG6026), which, in contrast to cibisatamab, is based on a 2 + 1 CrossMabVH-VL format with charge interactions using variable regions derived from obinutuzumab (). Glofitamab showed potent tumor cell killing and antitumor efficacy in preclinical in vitro, ex vivo and in vivo lymphoma models, as well as superiority over the respective conventional heterodimeric 1 + 1 TCB formats as a consequence of its head-to-tail orientation and bivalent binding to CD20, allowing pre-treatment with obinutuzumab, in this case as a strategy to reduce the incidence of cytokine-release syndrome by glofitamab.Citation86,Citation141,Citation142 Based on the clinical efficacy and safety in the Phase 1 clinical trial in relapsed/refractory non-Hodgkin lymphoma (NHL) patients and particularly the high rate of durable complete responses,Citation143,Citation144 glofitamab is currently being evaluated in multiple clinical trials in lymphoma patients, including trials in patients relapsed after CAR-T cell therapy (NCT04703686) and in Phase 3 clinical trials in relapsed/refractory diffuse large B cell lymphoma patients (NCT04077723, NCT04408638).Citation145 No anti-drug antibodies recognizing glofitamab were detected in the Phase 1 clinical study.Citation143
Additional analogous 2 + 1 T cell bispecific antibodies using this technology have entered early clinical Phase 1 trials, including the BCMA-TCE CC-93269 for the treatment of multiple myeloma (NCT03486067)Citation88 and the TYRP1-TCB (RG6232) for the treatment of TYRP1-expressing melanoma (NCT04551352).Citation92 Recently, the WT1-peptide-MHC-specific TCR-like WT1-TCB (RG6007) for the treatment of acute myeloid leukemia (AML) became the first TCR-like bispecific antibody to enter a clinical trial (NCT04580121).Citation93 While Immunocore pioneered the field of targeting peptide-MHC complexes with recombinant TCR-based bispecific T-cell engagers, the so-called ImmTACs,Citation146 WT1-TCB is based on a TCR-like antibody fragment recognizing the RMF WT1 peptide-HLA-A*02 complex. WT1-TCB can mediate specific killing of AML cell lines and primary AML cells, and it has anti-tumor activity in humanized mice bearing SKM-1 tumors.Citation93
Additional preclinical stage 2 + 1 TCBs based on this format have been described, including ones that target HER2Citation147,Citation148 or the p95 HER2 fragment.Citation149 The p95 HER2 fragment is only found on a portion of ~ 30–40% of HER2+++ tumor cells, and as such can be considered a highly tumor-specific neoantigen. Thus, 2 + 1 TCBs targeting specifically p95 HER2 are of particular interest as they do not mediate T cell killing of normal cells that express HER2, such as cardiomyocytes or breast epithelial cells, as opposed to conventional HER2-TCBs.Citation149 An alternative approach to overcome the on-target off-tumor killing of normal cells is tumor-specific activation of TCBs by protease expressed in the tumor. For this purpose, a protease-activated mesothelin-TCB using CrossMab technology has been described that is blocked by an anti-CD3 anti-idiotypic mask that is cleaved in the tumor-microenvironment.Citation150 Alternatively, to counteract and manage any undesired T cell activation, it was shown that the Src/lck inhibitor dasatinib is able to reversibly switch off cytokine release and T-cell cytotoxicity following stimulation with different 2 + 1 TCBs targeting CEA, CD19 and WT1.Citation151 Finally, related to the TCB approach, 2 + 1 bispecific antibodies designed based on CrossMab technology have been developed specifically for the recruitment of synthetic agonistic receptor transduced T-cells (SAR-T) in adoptive T-cell therapy together with Kobold and colleagues.Citation152–154
In order to further boost the potency of T-cell bispecific antibodies, the tumor (stroma)-targeted FAP-4-1BBL (RG7827), CD19-4-1BBL (RG6076) and CEA-4-1BBL fusion proteins have been developed for solid tumors and NHL. These molecules are used to provide the co-stimulatory TNF receptor superfamily-mediated signal 2 to T cells in combination with the T-cell bispecific antibodies cibisatamab or glofitamab, which provide the signal 1.Citation89,Citation155,Citation156 These 4–1BBL fusion proteins contain a split trimeric 4–1BB ligand fused to the CH1 and CL domains, and constant chain mispairing is abolished by CH1-CL domain crossover in conjunction with the respective charge pairs ().Citation89 FAP-4-1BBL and CD19-4-1BBL have been designed to trigger 4–1BB/CD137 hyper-clustering specifically in the tumor microenvironment, but not in circulation or in the liver, with the goal to overcome typical 4–1BB antibody-mediated toxicities.Citation89 Tumor-targeted 4–1BBL fusion proteins were shown to mediate improved T-cell activation, superior tumor control in combination with TCBs and checkpoint inhibitors, and strong T-cell infiltration in preclinical models.Citation89,Citation155,Citation157 Clinical Phase 1b studies combining cibisatamab with FAP-4-1-BBL (NCT04826003) and glofitamab with CD19-4-1BBL (NCT04077723) are currently ongoing.
A similar rationale was applied to trigger the TNF receptor superfamily member CD40 on antigen-presenting cells (APCs) through a trivalent C-terminally fused FAP/CD40 2 + 1 bispecific antibody in a 2 + 1 CrossMabVH-VL± format with charges (). This design was chosen to make FAP-CD40 (RG6189), a FAP-targeted CD40 agonistic bispecific antibody, with the goal of abrogating systemic toxicity and enabling administration of doses sufficiently high to result in highly tumor- and lymph node-specific activation of APCs with subsequent induction of antitumor immunity.Citation94,Citation122 Phase 1 clinical trials have been initiated to validate this approach in the clinic (NCT04857138).
Notably, the domain crossover/CrossMab technology has also been used by researchers outside of Roche for the development of bispecific antibodies for cancer immunotherapy. This includes the so-called Fabs-in-tandem Ig (FIT-Ig) approach developed by Gong and colleagues from EpimAb, which relies on Fab crossover to enable correct light-chain association for the generation of symmetric tetravalent N-terminally fused bispecific antibodies in the 2 + 2 CrossMabFab format ().Citation50,Citation51 Three different bispecific FIT-Igs have reached clinical Phase 1 trials to date co-targeting: 1) EGFR/c-Met for receptor tyrosine kinase inhibition (EMB-01) (NCT03797391), 2) PD-1/LAG-3 for dual checkpoint inhibition (EMB-02) (NCT04618393), and 3) BCMA/CD3ε for T cell engagement in multiple myeloma (EMB-06) (NCT04735575).
In order to recruit innate immune cells for cancer cell killing, Gauthier and colleagues from Innate Pharma recently described an advanced preclinical approach to generate multifunctional natural killer cell engagers (NKCE) targeting a tumor antigen and the NK cell ligand NKp46 in a FcγRIII-binding competent monovalent C-terminally fused 1 + 1 antibody format using CH1-CL crossover to ensure correct light-chain association ().Citation158 Trifunctional NKCEs targeting CD19, CD20, or EGFR as tumor antigens triggered tumor killing by human primary NK cells in vitro and induced NK cell infiltration and anti-tumor efficacy, as well as protective tumor immunity in vivo.Citation158
Zhao and colleagues demonstrated that a bispecific heterodimeric CD20/HLA-DR 1 + 1 CrossMabCHCitation1−CL termed CD20–243 CrossMab for the treatment of NHL patients co-expressing CD20 and HLA-DR mediated strong complement-dependent cytotoxicity, antibody-dependent cell-mediated cytotoxicity and anti-proliferative activity.Citation159 Similarly, Rajendran and colleagues generated a bispecific heterodimeric CD30/CD137 1 + 1 CrossMabCHCitation1−CL to target specifically these two co-expressed antigens on Hodgkin and Reed-Sternberg cells without inducing CD137 signaling.Citation160
In an alternative approach to activate innate immunity, Du and colleagues devised a bispecific heterodimeric GPC3/CD47 1 + 1 CrossMabCHCitation1−CL to bind to GPC3 and CD47 on hepatocellular cancer cells, and at the same time inhibit the CD47 interaction with SIRP1α responsible for the “do-not-eat-me signal” to recruit myeloid cells for phagocytosis.Citation161 The GPC3/CD47 CrossMab induced enhanced Fc-mediated effector functions by both macrophages and neutrophils toward dual antigen-expressing hepatocellular carcinoma (HCC) cells in vitro, and strong in vivo efficacy against xenograft HCC tumors in a fashion superior to the respective monotherapies and combination thereof.Citation161
In order to further boost antigen presentation and foster the generation of a secondary anti-tumor immune response, again Zhao and colleagues created a novel CD20/Flt3 ligand antibody fusion protein, termed CD20-Flex BiFP using CrossMab technology.Citation162 CD20-Flex BiFP not only eliminated lymphoma temporarily but also potentiated tumor-specific T-cell immunity by expanding and fostering infiltration of antigen-presenting dendritic cells into the tumor tissue.Citation162
Most recently, Panina and colleagues described a novel bispecific heterodimeric HER2/IFNα-1 + 1 CrossMabCHCitation1−CL with the ultimate goal to deliver IFNα into HER2 expressing tumors.Citation163
Applications in therapy of viral infections and autoimmune diseases
The application of CrossMab technology has become quite popular for the generation of bispecific and multispecific antibodies targeting various viruses. During the past years, multiple highly potent bispecific antibodies targeting HIV-1 have been generated using CrossMab technology for the prevention and treatment of HIV-1.Citation164,Citation165 Examples of these approaches are: 1) four different 1 + 1 CrossMabCHCitation1−CL-based bispecific antibodies, of which the one based on VRC07 and PG9-16 displayed the most favorable neutralization profile and IgG-like pharmacokinetic properties in monkeys;Citation166 2) 1 + 1 CrossMabCHCitation1−CL-based bispecific antibodies that, however, did not allow intra-spike binding;Citation167 3) unique bispecific antibodies based on the broadly neutralizing antibodies (bNAbs) 3BNC117 and 10–1074 with a modified hinge region of human IgG3 isotype for increased Fab flexibility and improved neutralization potency based on a 1 + 1 CrossMabCHCitation1−CL format;Citation168 4) a 1 + 1 CrossMabCHCitation1−CL-based bispecific antibody targeting two non-competing epitopes on the HIV-1 co-receptor CCR5 based on RoAb13 and PRO 140 to increase avidity;Citation169 5) the 1 + 1 CrossMabCHCitation1−CL-based bispecific antibody iMab‑CAP256 comprising the highly potent CAP256.VRC26.25 bNAb and the host-directed CD4 antibody, ibalizumab (iMab);Citation170 and 6) the 1 + 1 CrossMabCHCitation1−CL-based bispecific antibody BICM-1A for simultaneous recognition of two critical V2-and V3-glycan epitopes of the single HIV-1 envelope glycoprotein.Citation171 Of all these approaches, the heterodimeric bispecific 1 + 1 CrossMabCHCitation1−CL antibody 10E8.4/iMab showed exquisite potency and breadth against various HIV-1 strains, including activity in HIV-1 in vivo treatment and prevention models,Citation90,Citation172 and compared very favorably to conventional antibodies and other bispecific bNAbs.Citation173 Based on these data, 10E8.4/iMab is currently being evaluated in a Phase 1 clinical trial (NCT03875209).Citation174
Wang and colleagues generated a symmetric and tetravalent FIT-Ig-based bispecific antibody against Zika virus that showed high in vitro and in vivo potency, and prevented viral escape, supporting its potential use for the therapy of Zika virus prevention or infections.Citation175
Interestingly, and most recently, De Gasparo and colleagues described the first bispecific antibody targeting SARS-CoV-2 based on a 1 + 1 CrossMabCHCitation1−CL format targeting two non-overlapping sites on the receptor binding domain of SARS-CoV-2 and blocking binding to angiotensin-converting enzyme 2 (ACE2).Citation176 The respective bispecific antibody CoV-X2 was designed using C121 and C135, two antibodies derived from donors who had recovered from COVID-19. Most notably, CoV-X2 neutralized wild-type SARS-CoV-2 and variants of concern and escape, protected mice from disease and suppressed viral escape.Citation176 Along these lines, Jette and colleagues described a subset of donor-derived neutralizing bispecific CrossMabs with broad cross-reactivity to sarbecoviruses.Citation177
Bispecific CrossMab-based antibodies have also been generated with the goal of treating autoimmune diseases.Citation178,Citation179 Fischer and colleagues showed that combined inhibition of TNFα and IL-17 was more effective in inhibiting the development of inflammation and bone and cartilage destruction in arthritic mice compared to the respective monotherapies. For this purpose, bispecific TNFα/IL-17 1 + 1 and 2 + 2 CrossMabCHCitation1−CL antibodies were prepared that showed superior efficacy in blocking cytokine and chemokine responses in vitro.Citation180 Similarly, Xu and colleagues showed that a tetravalent bispecific TNFα/IL-17 1 + 1 CrossMabVH-VL together with electrostatic steering for heavy-chain heterodimerization significantly decreased the expression level of neutrophil and Th17 chemokines, and the secretion of IL-6/IL-8 on fibroblast-like synoviocytes. Moreover, combined inhibition of both cytokines by the bispecific antibody was superior to inhibition of either cytokine alone.Citation181 Based on these data, dual-targeting bispecific antibodies neutralizing pro-inflammatory cytokines may provide novel treatment options for autoimmune diseases. However, as they are not necessarily differentiated from the combination of the respective monotherapies, the benefit of using a bispecific antibody over a combination therapy needs to be assessed on a case-by-case basis.
Applications in ophthalmology and therapy of central nervous system diseases
The heterodimeric 1 + 1 VEGF/Ang-2 CrossMabCHCitation1−CL vanucizumab (RG7221) was the first anti-angiogenic bispecific antibody to enter clinical trials with the goal of suppressing tumor angiogenesis via simultaneous blockade of the pro-angiogenic ligands VEGF-A and Ang-2. VEGF and Ang-2 have also been shown to play an important role in ocular angiogenesis in diseases like wet age-related macular degeneration (wAMD) and diabetic macular edema (DME).96,Citation182–184 However, until now only the VEGF blocking antibody fragments ranibizumab and brolucizumab and the VEGFR1/2-ECD-Fc fusion protein aflibercept are approved for use in ophthalmology.Citation185
Faricimab (RG7716) is a heterodimeric 1 + 1 VEGF/Ang-2 CrossMabCHCitation1−CL specifically optimized for intraocular use and high concentration formulation in ophthalmology indications by use of optimized anti-VEGF and anti-Ang-2 Fabs, as compared to vanucizumab, and by the introduction of P329G LALA and Triple A mutations in the KiH-containing IgG1 Fc portion to abolish FcγR-mediated effector functions and FcRn recycling for low systemic exposure.Citation82,Citation83,Citation186–189 While faricimab neutralizes two soluble ligands, particularly in the field of ophthalmology the use of such a bispecific antibody provides advantages in terms of intraocular administration via a single injection due to the simultaneous inhibition of two different angiogenic pathways with a single agent. Importantly, as compared to VEGF inhibition alone, faricimab mediated improved anti-angiogenic activity in various preclinical models to limit pathological angiogenesis in the eye.Citation82,Citation83,Citation190,Citation191 Based on these data, faricimab was the first bispecific antibody worldwide entering Phase 1 clinical trials in ophthalmology, where it was well tolerated and exhibited a favorable safety profile with evidence of improvements in best-corrected visual acuity (BCVA) and anatomic parameters supporting further clinical investigation.Citation192 Subsequently, faricimab was compared head-to-head to ranibizumab in the BOULEVARD Phase 2 randomized clinical trial in patients with DME, where it met the primary end point and demonstrated statistically superior visual acuity gains versus ranibizumab, suggesting a benefit of simultaneous inhibition of angiopoietin-2 and VEGF-A.Citation193 In the AVENUE Phase 2 randomized clinical trial in patients with AMD, it did not meet the primary end point of superiority over ranibizumab in BCVA at week 36, but visual and anatomical gains observed with faricimab supported pursuing Phase 3 trials for an alternative to monthly anti-VEGF therapy.Citation194 This was taken into account together with the data from the STAIRWAY Phase 2 randomized clinical trial in AMD where faricimab dosed every 16 weeks or 12 weeks resulted in maintenance of initial vision and anatomic improvements comparable with monthly ranibizumab.Citation195,Citation196 Recently, positive outcomes were reported from four independent pivotal Phase 3 trials in wAMD and DME patients where faricimab was compared to aflibercept and met the primary endpoints (NCT03823287, NCT03823300, NCT03622580, NCT03622593). Based on these data, marketing applications for faricimab have been filed with health authorities for approval in DME and wAMD, with FDA granting it a priority review.Citation197
The treatment of central nervous system (CNS) diseases with monoclonal antibodies is hampered by the low penetration of antibodies through the blood-brain barrier, and the field still is in its infancy.Citation198 To overcome this limitation, Niewoehner and colleagues have generated transferrin receptor-targeted bispecific antibodies that allowed delivery of these antibodies through the blood-brain barrier and showed improved brain exposure and prevented plaque formation.Citation66,Citation67 Using this approach, BS-GANT (RG6102) was generated based on the amyloid-beta antibody gantenerumabCitation199 as a trivalent C-terminally fused amyloid-beta/TfR 2 + 1 bispecific antibody in a 2 + 1 CrossMabVH-VL± format with charges (). BS-GANT (RG6102) recently entered Phase 2 clinical trials in patients with prodromal or mild-to-moderate Alzheimer’s disease (NCT04639050).
Conclusions
During the past 20 years, numerous technologies have been developed to generate bispecific antibodies, and these molecules represent a rapidly growing class of biopharmaceuticals in clinical trials and on the market. CrossMab technology was first described in 2011 as a novel approach enabling correct antibody light-chain association with their respective heavy chain in bi-/multispecific antibodies, together with methods enabling correct heavy-chain association.
As briefly mentioned in the introduction, alternative technologies to achieve correct heavy-light-chain pairing are currently being applied for the generation of prototypical (heterodimeric) IgG-like bispecific antibodies. These include in vitro assembly approaches, where the two bispecific antibodies are produced separately and subsequently assembled in vitro like DuoBody,Citation19 Fab arm exchange,Citation22 FORCECitation26 or half antibody assembly,Citation27 as well as approaches allowing the production of bispecific antibodies in one cell line, for example via the use of common light chains or orthogonal Fab interfaces. Recently, several groups have also reported that the specific pairing preferences of selected heavy and light chain pairs can be used to drive the assembly of correct bispecific antibodies.Citation200,Citation201 In the field of common light chains, much progress has been made in the selection of suitable common light-chain antibodies from common light-chain-bearing animals or use of in vitro display technologies.Citation9,Citation10,Citation14–16,Citation202–205 Based on this progress, several bispecific common light-chain-based IgG antibodies are currently in clinical trials, including odronextamab, REGN4018, REGN5678, REGN7075, MCLA-145, MCLA-158, and others.Citation7,Citation8,Citation10,Citation15,Citation206–213 Alternatively, the correct light-chain-heavy-chain association can be enforced using orthogonal Fab interfaces by introduction of (several) mutations in the Fab interface.Citation28,Citation29,Citation33,Citation55,Citation56,Citation214,Citation215
CrossMab technology continues to represent a simple, straightforward and clinically validated antibody engineering solution to achieve correct light-chain association with minimal engineering using existing pairs of antibodies. In fact, since its original description, it has evolved into one of the most mature, versatile, and broadly applied technologies in industry and academia, in conjunction with the KiH technology. Until now ~20 bispecific antibodies and fusion proteins based on CrossMab technology developed by Roche and others have entered clinical trials. Based on the available clinical data, CrossMabs show favorable IgG-like properties in terms of pharmacokinetics and immunogenicity similar to conventional therapeutic monoclonal antibodies. The most advanced of these bispecific antibodies are: 1) the 1 + 1 heterodimeric Ang-2/VEGF bispecific antibody faricimab for the treatment of DME and wAMD, which is currently undergoing regulatory review, and 2) the 2 + 1 heterodimeric CD20/CD3 T-cell bispecific antibody glofitamab for the treatment of relapsed/refractory DLBCL or follicular NHL, which is currently in pivotal Phase 3 clinical trials.
Based on the progress in making bi- and multispecific antibodies, we anticipate that this class of therapeutics with novel mechanisms of actions as compared to conventional therapeutic antibodies will have a major impact on the treatment of various diseases, including oncology, infectious diseases, autoimmunity, CNS, and metabolic diseases. Taken together, CrossMab technology has proven to be very useful for the fast and straightforward generation of bispecific antibody formats to tackle novel biological challenges and help to develop novel therapeutic concepts for patients in need.
Abbreviations
Acknowledgments
The authors want to thank all contributors and project team members from Roche Innovation Centers Zurich, Munich and Basel, Roche pRED as well as all other collaborators within and outside of Roche contributing to the development of CrossMab technology and individual drug candidates for their support and effort. The authors apologize to those authors whose work could not be cited due to space restrictions.
Disclosure statement
MS declares employment with Roche, WS declares patents/royalties with Roche and CK declares employment, stock ownership, and patents/royalties with Roche. CROSSMAB® is a registered trademark by Genentech/Roche.
References
- Brinkmann U, Kontermann RE. The making of bispecific antibodies. MAbs. 2017;9(2):182–17. PubMed PMID: 28071970; PubMed Central PMCID: PMCPMC5297537. doi:10.1080/19420862.2016.1268307. Feb/Mar.
- Labrijn AF, Janmaat ML, Reichert JM, et al. Bispecific antibodies: a mechanistic review of the pipeline. Nat Rev Drug Discov. 2019 Aug;18(8):585–608. 10.1038/s41573-019-0028-1: PubMed PMID: 31175342
- Carter PJ, Lazar GA. Next generation antibody drugs: pursuit of the ‘high-hanging fruit’. Nat Rev Drug Discov. 2018 Mar;17(3):197–223. PubMed PMID: 29192287. doi:10.1038/nrd.2017.227.
- Sheridan C. Bispecific antibodies poised to deliver wave of cancer therapies. Nat Biotechnol. 2021 Mar;39(3):251–54. PubMed PMID: 33692520. doi:10.1038/s41587-021-00850-6.
- Suurs FV, Lub-de Hooge MN, De Vries EGE, et al. A review of bispecific antibodies and antibody constructs in oncology and clinical challenges. Pharmacol Ther. 2019 Sep;201:103–19. doi:10.1016/j.pharmthera.2019.04.006. PubMed PMID: 31028837.
- Zhang J, Yi J, Zhou P. Development of bispecific antibodies in China: overview and prospects. Antib Ther. 2020 Apr;3(2):126–45. doi:10.1093/abt/tbaa011. PubMed PMID: 33928227; PubMed Central PMCID: PMCPMC7990247.
- Kitazawa T, Igawa T, Sampei Z, et al. A bispecific antibody to factors IXa and X restores factor VIII hemostatic activity in a hemophilia A model. Nat Med. 2012 Oct;18(10):1570–74. 10.1038/nm.2942: PubMed PMID: 23023498
- Kitazawa T, Esaki K, Tachibana T, et al. Factor VIIIa-mimetic cofactor activity of a bispecific antibody to factors IX/IXa and X/Xa, emicizumab, depends on its ability to bridge the antigens. Thromb Haemost. 2017 Jun 28;117(7):1348–57. PubMed PMID: 28451690; PubMed Central PMCID: PMCPMC6292136. doi:10.1160/TH17-01-0030
- Igawa T. Next generation antibody therapeutics using bispecific antibody technology. Yakugaku Zasshi. 2017;137(7):831–36. doi:10.1248/yakushi.16-00252-3. PubMed PMID: 28674296
- Sampei Z, Igawa T, Soeda T, et al. Identification and multidimensional optimization of an asymmetric bispecific IgG antibody mimicking the function of factor VIII cofactor activity. PLoS One. 2013;8(2):e57479. PubMed PMID: 23468998; PubMed Central PMCID: PMCPMC3585358. doi:10.1371/journal.pone.0057479.
- Skegro D, Stutz C, Ollier R, et al. Immunoglobulin domain interface exchange as a platform technology for the generation of Fc heterodimers and bispecific antibodies. J Biol Chem. 2017 Jun 9;292(23):9745–59. PubMed PMID: 28450393; PubMed Central PMCID: PMCPMC5465497. doi:10.1074/jbc.M117.782433
- Einsele H, Borghaei H, Orlowski RZ, et al. The BiTE (bispecific T-cell engager) platform: development and future potential of a targeted immuno-oncology therapy across tumor types. Cancer. 2020 Jul 15;126(14):3192–201. PubMed PMID: 32401342. doi:10.1002/cncr.32909
- Wolf E, Hofmeister R, Kufer P, et al. BiTEs: bispecific antibody constructs with unique anti-tumor activity. Drug Discov Today. 2005 Sep 15;10(18):1237–44. PubMed PMID: 16213416. doi:10.1016/S1359-6446(05)03554-3
- De Nardis C, Hendriks LJA, Poirier E, et al. A new approach for generating bispecific antibodies based on a common light chain format and the stable architecture of human immunoglobulin G1. J Biol Chem. 2017 Sep 1;292(35):14706–17. PubMed PMID: 28655766; PubMed Central PMCID: PMCPMC5582861. doi:10.1074/jbc.M117.793497
- Smith EJ, Olson K, Haber LJ, et al. A novel, native-format bispecific antibody triggering T-cell killing of B-cells is robustly active in mouse tumor models and cynomolgus monkeys. Sci Rep. 2015 Dec 11;5(1):17943. PubMed PMID: 26659273; PubMed Central PMCID: PMCPMC4675964. doi:10.1038/srep17943
- Merchant AM, Zhu Z, Yuan JQ, et al. An efficient route to human bispecific IgG. Nat Biotechnol. 1998 Jul;16(7):677–81. 10.1038/nbt0798-677: PubMed PMID: 9661204
- Bostrom J, Yu SF, Kan D, et al. Variants of the antibody herceptin that interact with HER2 and VEGF at the antigen binding site. Science. 2009 Mar 20;323(5921):1610–14. PubMed PMID: 19299620. doi:10.1126/science.1165480
- Johnson S, Burke S, Huang L, et al. Effector cell recruitment with novel Fv-based dual-affinity re-targeting protein leads to potent tumor cytolysis and in vivo B-cell depletion. J Mol Biol. 2010 Jun 11;399(3):436–49. PubMed PMID: 20382161. doi:10.1016/j.jmb.2010.04.001
- Labrijn AF, Meesters JI, De Goeij BE, et al. Efficient generation of stable bispecific IgG1 by controlled Fab-arm exchange. Proc Natl Acad Sci U S A. 2013 Mar 26;110(13):5145–50. PubMed PMID: 23479652; PubMed Central PMCID: PMCPMC3612680. doi:10.1073/pnas.1220145110
- Beckmann R, Jensen K, Fenn S, et al. DutaFabs are engineered therapeutic Fab fragments that can bind two targets simultaneously. Nat Commun. 2021 Jan 29;12(1):708. PubMed PMID: 33514724; PubMed Central PMCID: PMCPMC7846786. doi:10.1038/s41467-021-20949-3
- Wu C, Ying H, Grinnell C, et al. Simultaneous targeting of multiple disease mediators by a dual-variable-domain immunoglobulin. Nat Biotechnol. 2007 Nov;25(11):1290–97. 10.1038/nbt1345: PubMed PMID: 17934452
- Strop P, Ho WH, Boustany LM, et al. Generating bispecific human IgG1 and IgG2 antibodies from any antibody pair. J Mol Biol. 2012 Jul 13;420(3):204–19. PubMed PMID: 22543237. doi:10.1016/j.jmb.2012.04.020
- Leung KM, Batey S, Rowlands R, et al. A HER2-specific modified fc fragment (fcab) induces antitumor effects through degradation of HER2 and Apoptosis. Mol Ther. 2015 Nov;23(11):1722–33. 10.1038/mt.2015.127: PubMed PMID: 26234505; PubMed Central PMCID: PMCPMC4817942
- Woisetschlager M, Antes B, Borrowdale R, et al. In vivo and in vitro activity of an immunoglobulin Fc fragment (Fcab) with engineered Her-2/neu binding sites. Biotechnol J. 2014 Jun;9(6):844–51. 10.1002/biot.201300387: PubMed PMID: 24806546
- Wozniak-Knopp G, Stadlmayr G, Perthold JW, et al. Designing Fcabs: well-expressed and stable high affinity antigen-binding Fc fragments. Protein Eng Des Sel. 2017 Sep 1;30(9):657–71. PubMed PMID: 28981753. doi:10.1093/protein/gzx042
- Dengl S, Mayer K, Bormann F, et al. Format chain exchange (FORCE) for high-throughput generation of bispecific antibodies in combinatorial binder-format matrices. Nat Commun. 2020 Oct 2;11(1):4974. PubMed PMID: 33009381; PubMed Central PMCID: PMCPMC7532213. doi:10.1038/s41467-020-18477-7
- Spiess C, Merchant M, Huang A, et al. Bispecific antibodies with natural architecture produced by co-culture of bacteria expressing two distinct half-antibodies. Nat Biotechnol. 2013 Aug;31(8):753–58. 10.1038/nbt.2621: PubMed PMID: 23831709
- Liu Z, Leng EC, Gunasekaran K, et al. A novel antibody engineering strategy for making monovalent bispecific heterodimeric IgG antibodies by electrostatic steering mechanism. J Biol Chem. 2015 Mar 20;290(12):7535–62. PubMed PMID: 25583986; PubMed Central PMCID: PMCPMC4367261. doi:10.1074/jbc.M114.620260
- Gunasekaran K, Pentony M, Shen M, et al. Enhancing antibody Fc heterodimer formation through electrostatic steering effects: applications to bispecific molecules and monovalent IgG. J Biol Chem. 2010 Jun 18;285(25):19637–46. PubMed PMID: 20400508; PubMed Central PMCID: PMCPMC2885242. doi:10.1074/jbc.M110.117382
- Coloma MJ, Morrison SL. Design and production of novel tetravalent bispecific antibodies. Nat Biotechnol. 1997 Feb;15(2):159–63. PubMed PMID: 9035142. doi:10.1038/nbt0297-159.
- Dong J, Sereno A, Aivazian D, et al. A stable IgG-like bispecific antibody targeting the epidermal growth factor receptor and the type I insulin-like growth factor receptor demonstrates superior anti-tumor activity. MAbs. 2011 May-Jun;3(3):273–88. 10.4161/mabs.3.3.15188: PubMed PMID: 21393993; PubMed Central PMCID: PMCPMC3149708
- Fischer N, Elson G, Magistrelli G, et al. Exploiting light chains for the scalable generation and platform purification of native human bispecific IgG. Nat Commun. 2015 Feb 12;6:6113. 10.1038/ncomms7113: PubMed PMID: 25672245; PubMed Central PMCID: PMCPMC4339886
- Lewis SM, Wu X, Pustilnik A, et al. Generation of bispecific IgG antibodies by structure-based design of an orthogonal Fab interface. Nat Biotechnol. 2014 Feb;32(2):191–98. 10.1038/nbt.2797: PubMed PMID: 24463572
- Cochlovius B, Kipriyanov SM, Stassar MJ, et al. Cure of Burkitt’s lymphoma in severe combined immunodeficiency mice by T cells, tetravalent CD3 x CD19 tandem diabody, and CD28 costimulation. Cancer Res. 2000 Aug 15;60(16):4336–41. PubMed PMID: 10969772.
- Moore GL, Bernett MJ, Rashid R, et al. A robust heterodimeric Fc platform engineered for efficient development of bispecific antibodies of multiple formats. Methods. 2019 Feb 1;154:38–50. 10.1016/j.ymeth.2018.10.006: PubMed PMID: 30366098
- Guo G, Han J, Wang Y, et al. A potential downstream platform approach for WuXiBody-based IgG-like bispecific antibodies. Protein Expr Purif. 2020 Sep;173:105647. doi:10.1016/j.pep.2020.105647. PubMed PMID: 32334139.
- Bargou R, Leo E, Zugmaier G, et al. Tumor regression in cancer patients by very low doses of a T cell-engaging antibody. Science. 2008 Aug 15;321(5891):974–77. PubMed PMID: 18703743. doi:10.1126/science.1158545
- Neijssen J, Cardoso RMF, Chevalier KM, et al. Discovery of amivantamab (JNJ-61186372), a bispecific antibody targeting EGFR and MET. J Biol Chem. 2021 Apr;8:100641. doi:10.1016/j.jbc.2021.100641. PubMed PMID: 33839159; PubMed Central PMCID: PMCPMC8113745.
- Vijayaraghavan S, Lipfert L, Chevalier K, et al. Amivantamab (JNJ-61186372), an Fc Enhanced EGFR/cMet Bispecific Antibody, Induces Receptor Downmodulation and Antitumor Activity by Monocyte/Macrophage Trogocytosis. Mol Cancer Ther. 2020 Oct;19(10):2044–56. 10.1158/1535-7163.MCT-20-0071: PubMed PMID: 32747419
- Yun J, Lee SH, Kim SY, et al. Antitumor Activity of Amivantamab (JNJ-61186372), an EGFR-MET bispecific antibody, in diverse models of EGFR Exon 20 Insertion-Driven NSCLC. Cancer Discov. 2020 Aug;10(8):1194–209. 10.1158/2159-8290.CD-20-0116: PubMed PMID: 32414908
- Schaefer W, Regula JT, Bahner M, et al. Immunoglobulin domain crossover as a generic approach for the production of bispecific IgG antibodies. Proc Natl Acad Sci U S A. 2011 Jul 5;108(27):11187–92. PubMed PMID: 21690412; PubMed Central PMCID: PMCPMC3131342. doi:10.1073/pnas.1019002108
- Klein C, Sustmann C, Thomas M, et al. Progress in overcoming the chain association issue in bispecific heterodimeric IgG antibodies. MAbs. 2012 Nov-Dec;4(6):653–63. 10.4161/mabs.21379: PubMed PMID: 22925968; PubMed Central PMCID: PMCPMC3502232
- Grote M, Haas AK, Klein C, et al. Bispecific antibody derivatives based on full-length IgG formats. Methods Mol Biol. 2012;901:247–63. PubMed PMID: 22723106. doi:10.1007/978-1-61779-931-0_16.
- Klein C, Schaefer W, Regula JT. The use of CrossMAb technology for the generation of bi- and multispecific antibodies. MAbs. 2016 Aug-Sep;8(6):1010–20. PubMed PMID: 27285945; PubMed Central PMCID: PMCPMC4968094. doi:10.1080/19420862.2016.1197457.
- Klein C, Schaefer W, Regula JT, et al. Engineering therapeutic bispecific antibodies using CrossMab technology. Methods. 2019 Feb 1;154:21–31. 10.1016/j.ymeth.2018.11.008: PubMed PMID: 30453028
- Schaefer W, Volger HR, Lorenz S, et al. Heavy and light chain pairing of bivalent quadroma and knobs-into-holes antibodies analyzed by UHR-ESI-QTOF mass spectrometry. MAbs. 2016;8(1):49–55. PubMed PMID: 26496506; PubMed Central PMCID: PMCPMC4966523. doi:10.1080/19420862.2015.1111498.
- Ridgway JB, Presta LG, Carter P. ‘Knobs-into-holes’ engineering of antibody CH3 domains for heavy chain heterodimerization. Protein Eng. 1996 Jul;9(7):617–21. doi:10.1093/protein/9.7.617. PubMed PMID: 8844834.
- Kuglstatter A, Stihle M, Neumann C, et al. Structural differences between glycosylated, disulfide-linked heterodimeric Knob-into-Hole Fc fragment and its homodimeric Knob-Knob and Hole-Hole side products. Protein Eng Des Sel. 2017 Sep 1;30(9):649–56. PubMed PMID: 28985438. doi:10.1093/protein/gzx041
- Stutz C, Blein S. A single mutation increases heavy-chain heterodimer assembly of bispecific antibodies by inducing structural disorder in one homodimer species. J Biol Chem. 2020 Jul 10;295(28):9392–408. PubMed PMID: 32404368; PubMed Central PMCID: PMCPMC7363136. doi:10.1074/jbc.RA119.012335.
- Gong S, Ren F, Wu D, et al. Fabs-in-tandem immunoglobulin is a novel and versatile bispecific design for engaging multiple therapeutic targets. MAbs. 2017 Oct 9;7:1118–28. doi:10.1080/19420862.2017.1345401. PubMed PMID: 28692328; PubMed Central PMCID: PMCPMC5627593.
- Gong S, Wu C. Generation of Fabs-in-tandem immunoglobulin molecules for dual-specific targeting. Methods. 2019 Feb 1;154:87–92. doi:10.1016/j.ymeth.2018.07.014. PubMed PMID: 30081078.
- Fenn S, Schiller CB, Griese JJ, et al. Crystal structure of an anti-Ang2 CrossFab demonstrates complete structural and functional integrity of the variable domain. PLoS One. 2013;8(4):e61953. PubMed PMID: 23613981; PubMed Central PMCID: PMCPMC3629102. doi:10.1371/journal.pone.0061953.
- Regula JT, Imhof-Jung S, Molhoj M, et al. Variable heavy-variable light domain and Fab-arm CrossMabs with charged residue exchanges to enforce correct light chain assembly. Protein Eng Des Sel. 2018 Jul 1;31(7–8):289–99. PubMed PMID: 30169707; PubMed Central PMCID: PMCPMC6277175. doi:10.1093/protein/gzy021
- Sustmann C, Dickopf S, Regula JT, et al. DuoMab: a novel CrossMab-based IgG-derived antibody format for enhanced antibody-dependent cell-mediated cytotoxicity. MAbs. 2019 Nov-Dec;11(8):1402–14. 10.1080/19420862.2019.1661736: PubMed PMID: 31526159; PubMed Central PMCID: PMCPMC6816436
- Wu X, Yuan R, Bacica M, et al. Generation of orthogonal Fab-based trispecific antibody formats. Protein Eng Des Sel. 2018 Jul 1;31(7–8):249–56. PubMed PMID: 29718394. doi:10.1093/protein/gzy007
- Wu X, Demarest SJ. Building blocks for bispecific and trispecific antibodies. Methods. 2019 Feb 1;154:3–9. doi:10.1016/j.ymeth.2018.08.010. PubMed PMID: 30172007.
- Bethune MT, Gee MH, Bunse M, et al. Domain-swapped T cell receptors improve the safety of TCR gene therapy. Elife. 2016 Nov;8:5. doi:10.7554/eLife.19095. PubMed PMID: 27823582; PubMed Central PMCID: PMCPMC5101000.
- Croasdale R, Wartha K, Schanzer JM, et al. Development of tetravalent IgG1 dual targeting IGF-1R-EGFR antibodies with potent tumor inhibition. Arch Biochem Biophys. 2012 Oct 15;526(2):206–18. PubMed PMID: 22464987. doi:10.1016/j.abb.2012.03.016
- Scheuer W, Thomas M, Hanke P, et al. Anti-tumoral, anti-angiogenic and anti-metastatic efficacy of a tetravalent bispecific antibody (TAvi6) targeting VEGF-A and angiopoietin-2. MAbs. 2016;8(3):562–73. PubMed PMID: 26864324; PubMed Central PMCID: PMCPMC4966847. doi:10.1080/19420862.2016.1147640.
- Schanzer J, Jekle A, Nezu J, et al. Development of tetravalent, bispecific CCR5 antibodies with antiviral activity against CCR5 monoclonal antibody-resistant HIV-1 strains. Antimicrob Agents Chemother. 2011 May;55(5):2369–78. 10.1128/AAC.00215-10: PubMed PMID: 21300827; PubMed Central PMCID: PMCPMC3088204
- Castoldi R, Ecker V, Wiehle L, et al. A novel bispecific EGFR/Met antibody blocks tumor-promoting phenotypic effects induced by resistance to EGFR inhibition and has potent antitumor activity. Oncogene. 2013 Dec 12;32(50):5593–601. PubMed PMID: 23812422; PubMed Central PMCID: PMCPMC3898114. doi:10.1038/onc.2013.245
- Castoldi R, Jucknischke U, Pradel LP, et al. Molecular characterization of novel trispecific ErbB-cMet-IGF1R antibodies and their antigen-binding properties. Protein Eng Des Sel. 2012 Oct;25(10):551–59. 10.1093/protein/gzs048: PubMed PMID: 22936109; PubMed Central PMCID: PMCPMC3449402
- Castoldi R, Schanzer J, Panke C, et al. TetraMabs: simultaneous targeting of four oncogenic receptor tyrosine kinases for tumor growth inhibition in heterogeneous tumor cell populations. Protein Eng Des Sel. 2016 Oct;29(10):467–75. 10.1093/protein/gzw037: PubMed PMID: 27578890; PubMed Central PMCID: PMCPMC5036864
- Schanzer JM, Wartha K, Croasdale R, et al. A novel glycoengineered bispecific antibody format for targeted inhibition of epidermal growth factor receptor (EGFR) and insulin-like growth factor receptor type I (IGF-1R) demonstrating unique molecular properties. J Biol Chem. 2014 Jul 4;289(27):18693–706. PubMed PMID: 24841203; PubMed Central PMCID: PMCPMC4081915. doi:10.1074/jbc.M113.528109
- Schanzer JM, Wartha K, Moessner E, et al. XGFR*, a novel affinity-matured bispecific antibody targeting IGF-1R and EGFR with combined signaling inhibition and enhanced immune activation for the treatment of pancreatic cancer. MAbs. 2016 May-Jun;8(4):811–27. 10.1080/19420862.2016.1160989: PubMed PMID: 26984378; PubMed Central PMCID: PMCPMC4966845
- Niewoehner J, Bohrmann B, Collin L, et al. Increased brain penetration and potency of a therapeutic antibody using a monovalent molecular shuttle. Neuron. 2014 Jan 8;81(1):49–60. PubMed PMID: 24411731. doi:10.1016/j.neuron.2013.10.061
- Weber F, Bohrmann B, Niewoehner J, et al. Brain shuttle antibody for alzheimer’s disease with attenuated peripheral effector function due to an inverted binding mode. Cell Rep. 2018 Jan 2;22(1):149–62. PubMed PMID: 29298417. doi:10.1016/j.celrep.2017.12.019
- Dengl S, Wehmer M, Hesse F, et al. Aggregation and chemical modification of monoclonal antibodies under upstream processing conditions. Pharm Res. 2013 May;30(5):1380–99. 10.1007/s11095-013-0977-8: PubMed PMID: 23322133
- Haberger M, Leiss M, Heidenreich AK, et al. Rapid characterization of biotherapeutic proteins by size-exclusion chromatography coupled to native mass spectrometry. MAbs. 2016;8(2):331–39. PubMed PMID: 26655595; PubMed Central PMCID: PMCPMC4966600. doi:10.1080/19420862.2015.1122150.
- Lee YF, Kluters S, Hillmann M, et al. Modeling of bispecific antibody elution in mixed-mode cation-exchange chromatography. J Sep Sci. 2017 Sep;40(18):3632–45. 10.1002/jssc.201700313: PubMed PMID: 28714211
- Meschendoerfer W, Gassner C, Lipsmeier F, et al. SPR-based assays enable the full functional analysis of bispecific molecules. J Pharm Biomed Anal. 2017 Jan 5;132:141–47. 10.1016/j.jpba.2016.09.028: PubMed PMID: 27721070
- Lu Y, Zhou Q, Han Q, et al. Inactivation of deubiquitinase CYLD enhances therapeutic antibody production in Chinese hamster ovary cells. Appl Microbiol Biotechnol. 2018 Jul;102(14):6081–93. 10.1007/s00253-018-9070-x: PubMed PMID: 29766242
- Gstottner C, Nicolardi S, Haberger M, et al. Intact and subunit-specific analysis of bispecific antibodies by sheathless CE-MS. Anal Chim Acta. 2020 Oct 16;1134:18–27. 10.1016/j.aca.2020.07.069: PubMed PMID: 33059862
- Gstottner C, Reusch D, Haberger M, et al. Monitoring glycation levels of a bispecific monoclonal antibody at subunit level by ultrahigh-resolution MALDI FT-ICR mass spectrometry. MAbs. 2020 Jan-Dec;12(1):1682403. 10.1080/19420862.2019.1682403: PubMed PMID: 31630606; PubMed Central PMCID: PMCPMC6927770
- Graf T, Heinrich K, Grunert I, et al. Recent advances in LC-MS based characterization of protein-based bio-therapeutics - mastering analytical challenges posed by the increasing format complexity. J Pharm Biomed Anal. 2020 Jul 15;186:113251. 10.1016/j.jpba.2020.113251: PubMed PMID: 32251978
- Filep C, Szigeti M, Farsang R, et al. Multilevel capillary gel electrophoresis characterization of new antibody modalities. Anal Chim Acta. 2021 Jun 29;1166:338492. 10.1016/j.aca.2021.338492: PubMed PMID: 34023000
- Chen SW, Zhang W. Current trends and challenges in the downstream purification of bispecific antibodies. Antib Ther. 2021 Apr;4(2):73–88. doi:10.1093/abt/tbab007. PubMed PMID: 34056544; PubMed Central PMCID: PMCPMC8155696.
- Register AC, Tarighat SS, Lee HY. Bioassay development for bispecific antibodies-challenges and opportunities. Int J Mol Sci. 2021 May 19;22(10). PubMed PMID: 34069573; PubMed Central PMCID: PMCPMC8160952. doi:10.3390/ijms22105350.
- Haberger M, Heidenreich AK, Hook M, et al. Multiattribute monitoring of antibody charge variants by cation-exchange chromatography coupled to native mass spectrometry. J Am Soc Mass Spectrom. 2021 Mar 9 [ PubMed PMID: 33687195]. doi:10.1021/jasms.0c00446.
- Wan Y, Wang Y, Zhang T, et al. Application of pH-salt dual gradient elution in purifying a WuXiBody-based bispecific antibody by MMC ImpRes mixed-mode chromatography. Protein Expr Purif. 2021 May;181:105822. doi:10.1016/j.pep.2021.105822. PubMed PMID: 33429037.
- Kienast Y, Klein C, Scheuer W, et al. Ang-2-VEGF-A CrossMab, a novel bispecific human IgG1 antibody blocking VEGF-A and Ang-2 functions simultaneously, mediates potent antitumor, antiangiogenic, and antimetastatic efficacy. Clin Cancer Res. 2013 Dec 15;19(24):6730–40. PubMed PMID: 24097868. doi:10.1158/1078-0432.CCR-13-0081
- Regula JT, Lundh von Leithner P, Foxton R, et al. Targeting key angiogenic pathways with a bispecific Cross MAb optimized for neovascular eye diseases. EMBO Mol Med. 2016 Nov 8;8(11):1265–88. PubMed PMID: 27742718; PubMed Central PMCID: PMCPMC5090659. doi:10.15252/emmm.201505889
- Regula JT, Lundh von Leithner P, Foxton R, et al. Targeting key angiogenic pathways with a bispecific CrossMAb optimized for neovascular eye diseases. EMBO Mol Med. 2019 May;11(5): 10.15252/emmm.201910666. PubMed PMID: 31040127; PubMed Central PMCID: PMCPMC6505574.
- Bacac M, Fauti T, Sam J, et al. A novel carcinoembryonic antigen T-cell bispecific antibody (CEA TCB) for the treatment of solid tumors. Clin Cancer Res. 2016 Jul 1;22(13):3286–97. PubMed PMID: 26861458. doi:10.1158/1078-0432.CCR-15-1696
- Brunker P, Wartha K, Friess T, et al. RG7386, a novel tetravalent FAP-DR5 antibody, effectively triggers fap-dependent, avidity-driven dr5 hyperclustering and tumor cell apoptosis. Mol Cancer Ther. 2016 May;15(5):946–57. 10.1158/1535-7163.MCT-15-0647: PubMed PMID: 27037412
- Bacac M, Colombetti S, Herter S, et al. CD20-TCB with obinutuzumab pretreatment as next-generation treatment of hematologic malignancies. Clin Cancer Res. 2018 Oct 1;24(19):4785–97. PubMed PMID: 29716920. doi:10.1158/1078-0432.CCR-18-0455
- Deak LLC, Seeber S, Perro M, et al. Abstract 2270: RG7769 (PD1-TIM3), a novel heterodimeric avidity-driven T cell specific PD-1/TIM-3 bispecific antibody lacking Fc-mediated effector functions for dual checkpoint inhibition to reactivate dysfunctional T cells. Cancer Research. 2020;80(16 Supplement):2270–2270. doi:10.1158/1538-7445.am2020-2270.
- Seckinger A, Delgado JA, Moser S, et al. Target expression, generation, preclinical activity, and pharmacokinetics of the BCMA-T cell bispecific antibody EM801 for multiple myeloma treatment. Cancer Cell. 2017 Mar 13;31(3):396–410. PubMed PMID: 28262554. doi:10.1016/j.ccell.2017.02.002
- Claus C, Ferrara C, Xu W, et al. Tumor-targeted 4-1BB agonists for combination with T cell bispecific antibodies as off-the-shelf therapy. Sci Transl Med. 2019 Jun 12;11(496):eaav5989. PubMed PMID: 31189721; PubMed Central PMCID: PMCPMC7181714. doi:10.1126/scitranslmed.aav5989
- Huang Y, Yu J, Lanzi A, et al. Engineered Bispecific Antibodies with Exquisite HIV-1-Neutralizing Activity. Cell. 2016 Jun 16;165(7):1621–31. PubMed PMID: 27315479; PubMed Central PMCID: PMCPMC4972332. doi:10.1016/j.cell.2016.05.024
- Deak LC, Weber P, Seeber S, et al. editors. A novel bispecific checkpoint inhibitor antibody to preferentially block PD-1 and LAG-3 on dysfunctional TILs whilst sparing Treg activation. In: JOURNAL FOR IMMUNOTHERAPY OF CANCER. ENGLAND: BMC CAMPUS, 4 CRINAN ST, LONDON N1 9XW; 2019.
- Nicolini VG, Waldhauer I, Freimoser-Grundschober A, et al. Abstract LB-389: combination of TYRP1-TCB, a novel T cell bispecific antibody for the treatment of melanoma, with immunomodulatory agents. Cancer Research. 2020;80(16Supplement):LB-389-LB-389. doi:10.1158/1538-7445.am2020-lb-389.
- Augsberger C, Hanel G, Xu W, et al. Targeting intracellular WT1 in AML with a novel RMF-peptide-MHC specific T-cell bispecific antibody. Blood. 2021 Jul 19 [ PubMed PMID: 34280257]. doi:10.1182/blood.2020010477.
- Sum E, Rapp M, Frobel P, et al. Fibroblast activation protein alpha-targeted CD40 agonism abrogates systemic toxicity and enables administration of high doses to induce effective antitumor immunity. Clin Cancer Res. 2021 Mar 26;27(14):4036–53. PubMed PMID: 33771854. doi:10.1158/1078-0432.CCR-20-4001
- Chung AS, Lee J, Ferrara N. Targeting the tumour vasculature: insights from physiological angiogenesis. Nat Rev Cancer. 2010 Jul;10(7):505–14. PubMed PMID: 20574450. doi:10.1038/nrc2868.
- Ferrara N, Adamis AP. Ten years of anti-vascular endothelial growth factor therapy. Nat Rev Drug Discov. 2016 Jun;15(6):385–403. PubMed PMID: 26775688. doi:10.1038/nrd.2015.17.
- Parmar D, Apte M. Angiopoietin inhibitors: a review on targeting tumor angiogenesis. Eur J Pharmacol. 2021 May 15;899:174021. doi:10.1016/j.ejphar.2021.174021. PubMed PMID: 33741382.
- Kloepper J, Riedemann L, Amoozgar Z, et al. Ang-2/VEGF bispecific antibody reprograms macrophages and resident microglia to anti-tumor phenotype and prolongs glioblastoma survival. Proc Natl Acad Sci U S A. 2016 Apr 19;113(16):4476–81. PubMed PMID: 27044098; PubMed Central PMCID: PMCPMC4843473. doi:10.1073/pnas.1525360113
- Baker LCJ, Boult JKR, Thomas M, et al. Acute tumour response to a bispecific Ang-2-VEGF-A antibody: insights from multiparametric MRI and gene expression profiling. Br J Cancer. 2016 Sep 6;115(6):691–702. PubMed PMID: 27529514; PubMed Central PMCID: PMCPMC5023775. doi:10.1038/bjc.2016.236
- Solecki G, Osswald M, Weber D, et al. Differential effects of ang-2/VEGF-A inhibiting antibodies in combination with radio- or chemotherapy in glioma. Cancers (Basel). 2019 Mar 6;11(3): 10.3390/cancers11030314. PubMed PMID: 30845704; PubMed Central PMCID: PMCPMC6468722.
- Mueller T, Freystein J, Lucas H, et al. Efficacy of a bispecific antibody co-targeting VEGFA and Ang-2 in combination with chemotherapy in a chemoresistant colorectal carcinoma xenograft model. Molecules. 2019 Aug 7;24(16): 10.3390/molecules24162865. PubMed PMID: 31394786; PubMed Central PMCID: PMCPMC6719918.
- Schmittnaegel M, Rigamonti N, Kadioglu E, et al. Dual angiopoietin-2 and VEGFA inhibition elicits antitumor immunity that is enhanced by PD-1 checkpoint blockade. Sci Transl Med. 2017 Apr 12;9(385): 10.1126/scitranslmed.aak9670. PubMed PMID: 28404865.
- Killock D. Immunotherapy: combine and conquer - antiangiogenic immunotherapy. Nat Rev Clin Oncol. 2017 Jun;14(6):327. PubMed PMID: 28466876. doi:10.1038/nrclinonc.2017.65.
- Schmittnaegel M, De Palma M. Reprogramming tumor blood vessels for enhancing immunotherapy. Trends Cancer. 2017 Dec;3(12):809–12. doi:10.1016/j.trecan.2017.10.002. PubMed PMID: 29198436.
- Kashyap AS, Schmittnaegel M, Rigamonti N, et al. Optimized antiangiogenic reprogramming of the tumor microenvironment potentiates CD40 immunotherapy. Proc Natl Acad Sci U S A. 2020 Jan 7;117(1):541–51. PubMed PMID: 31889004; PubMed Central PMCID: PMCPMC6955310. doi:10.1073/pnas.1902145116
- Ragusa S, Prat-Luri B, Gonzalez-Loyola A, et al. Antiangiogenic immunotherapy suppresses desmoplastic and chemoresistant intestinal tumors in mice. J Clin Invest. 2020 Mar 2;130(3):1199–216. PubMed PMID: 32015230; PubMed Central PMCID: PMCPMC7269598. doi:10.1172/JCI129558
- Hidalgo M, Martinez-Garcia M, Le Tourneau C, et al. First-in-human phase i study of single-agent vanucizumab, a first-in-class bispecific anti-angiopoietin-2/Anti-VEGF-A antibody, in adult patients with advanced solid tumors. Clin Cancer Res. 2018 Apr 1;24(7):1536–45. PubMed PMID: 29217526. doi:10.1158/1078-0432.CCR-17-1588
- Heil F, Babitzki G, Julien-Laferriere A, et al. Vanucizumab mode of action: serial biomarkers in plasma, tumor, and skin-wound-healing biopsies. Transl Oncol. 2021 Feb;14(2):100984. 10.1016/j.tranon.2020.100984: PubMed PMID: 33338877; PubMed Central PMCID: PMCPMC7749407
- Bendell JC, Sauri T, Gracian AC, et al. The McCAVE Trial: vanucizumab plus mFOLFOX-6 Versus Bevacizumab plus mFOLFOX-6 in patients with previously untreated metastatic colorectal carcinoma (mCRC). Oncologist. 2020 Mar;25(3):e451–e459. 10.1634/theoncologist.2019-0291: PubMed PMID: 32162804; PubMed Central PMCID: PMCPMC7066709
- Hauschildt J, Schrimpf C, Thamm K, et al. Dual Pharmacological Inhibition of Angiopoietin-2 and VEGF-A in murine experimental sepsis. J Vasc Res. 2020;57(1):34–45. PubMed PMID: 31726451. doi:10.1159/000503787.
- Zhou R, Wang S, Wen H, et al. The bispecific antibody HB-32, blockade of both VEGF and DLL4 shows potent anti-angiogenic activity in vitro and anti-tumor activity in breast cancer xenograft models. Exp Cell Res. 2019 Jul 15;380(2):141–48. PubMed PMID: 31034805. doi:10.1016/j.yexcr.2019.04.025
- Gschwind A, Fischer OM, Ullrich A. The discovery of receptor tyrosine kinases: targets for cancer therapy. Nat Rev Cancer. 2004 May;4(5):361–70. doi:10.1038/nrc1360. PubMed PMID: 15122207.
- Zhang F, Zhang J, Liu M, et al. Combating HER2-overexpressing breast cancer through induction of calreticulin exposure by Tras-Permut CrossMab. Oncoimmunology. 2015 Mar;4(3):e994391. 10.4161/2162402X.2014.994391: PubMed PMID: 25949918; PubMed Central PMCID: PMCPMC4404837
- Hu S, Fu W, Xu W, et al. Four-in-one antibodies have superior cancer inhibitory activity against EGFR, HER2, HER3, and VEGF through disruption of HER/MET crosstalk. Cancer Res. 2015 Jan 1;75(1):159–70. PubMed PMID: 25371409. doi:10.1158/0008-5472.CAN-14-1670
- Hu S, Fu W, Li T, et al. Antagonism of EGFR and Notch limits resistance to EGFR inhibitors and radiation by decreasing tumor-initiating cell frequency. Sci Transl Med. 2017 Mar 8;9(380):eaag0339. PubMed PMID: 28275151. doi:10.1126/scitranslmed.aag0339
- Fu W, Lei C, Yu Y, et al. EGFR/notch antagonists enhance the response to inhibitors of the PI3K-Akt pathway by decreasing tumor-initiating cell frequency. Clin Cancer Res. 2019 May 1;25(9):2835–47. PubMed PMID: 30670492. doi:10.1158/1078-0432.CCR-18-2732
- Lim B, Greer Y, Lipkowitz S, et al. Novel Apoptosis-Inducing Agents for the Treatment of Cancer, a New Arsenal in the Toolbox. Cancers (Basel). 2019 Jul 31;11(8):1087. PubMed PMID: 31370269; PubMed Central PMCID: PMCPMC6721450. doi:10.3390/cancers11081087
- Carneiro BA, El-Deiry WS. Targeting apoptosis in cancer therapy. Nat Rev Clin Oncol. 2020 Jul;17(7):395–417. PubMed PMID: 32203277; PubMed Central PMCID: PMCPMC8211386. doi:10.1038/s41571-020-0341-y.
- Davidson S, Coles M, Thomas T, et al. Fibroblasts as immune regulators in infection, inflammation and cancer. Nat Rev Immunol. 2021 Apr 28 [ PubMed PMID: 33911232]. doi:10.1038/s41577-021-00540-z.
- Kratochwil C, Flechsig P, Lindner T, et al. (68)Ga-FAPIPET/CT: tracer uptake in 28 different kinds of cancer. J Nucl Med. 2019 Jun;60(6):801–05. 10.2967/jnumed.119.227967: PubMed PMID: 30954939; PubMed Central PMCID: PMCPMC6581228
- Locksley RM, Killeen N, Lenardo MJ. The TNF and TNF receptor superfamilies: integrating mammalian biology. Cell. 2001 Feb 23;104(4):487–501. PubMed PMID: 11239407. doi:10.1016/s0092-8674(01)00237-9.
- Labiano S, Roh V, Godfroid C, et al. CD40 agonist targeted to fibroblast activation protein alpha synergizes with radiotherapy in murine hpv-positive head and neck tumors. Clin Cancer Res. 2021 Apr 26;27(14):4054–65. PubMed PMID: 33903200. doi:10.1158/1078-0432.CCR-20-4717
- Tung HY, Su YC, Chen BM, et al. Selective delivery of PEGylated compounds to tumor cells by anti-PEG hybrid antibodies. Mol Cancer Ther. 2015 Jun;14(6):1317–26. 10.1158/1535-7163.MCT-15-0151: PubMed PMID: 25852063
- Herrera-Camacho I, Anaya-Ruiz M, Perez-Santos M, et al. Cancer immunotherapy using anti-TIM3/PD-1 bispecific antibody: a patent evaluation of EP3356411A1. Expert Opin Ther Pat. 2019 Aug;29(8):587–93. 10.1080/13543776.2019.1637422: PubMed PMID: 31241380
- Cebada J, Flores A, Bandala C, et al. Bispecific anti-PD-1/LAG-3 antibodies for treatment of advanced or metastatic solid tumors: a patent evaluation of US2018326054. Expert Opin Ther Pat. 2020 Jul;30(7):487–94. 10.1080/13543776.2020.1767071: PubMed PMID: 32397849
- Dougall WC, Roman Aguilera A, Smyth MJ. Dual targeting of RANKL and PD-1 with a bispecific antibody improves anti-tumor immunity. Clin Transl Immunology. 2019;8(10):e01081. doi:10.1002/cti2.1081. PubMed PMID: 31572609; PubMed Central PMCID: PMCPMC6763724
- De Miguel M, Umana P, Gomes de Morais AL, et al. T-cell-engaging Therapy for Solid Tumors. Clin Cancer Res. 2021 Mar 15;27(6):1595–603. PubMed PMID: 33082210. doi:10.1158/1078-0432.CCR-20-2448
- Singh A, Dees S, Grewal IS. Overcoming the challenges associated with CD3+ T-cell redirection in cancer. Br J Cancer. 2021 Mar;124(6):1037–48. PubMed PMID: 33469153; PubMed Central PMCID: PMCPMC7960983. doi:10.1038/s41416-020-01225-5.
- Crawford A, Chiu D. Targeting solid tumors using CD3 bispecific antibodies. Mol Cancer Ther. 2021 May 27;20(8):1350–58. PubMed PMID: 34045228. doi:10.1158/1535-7163.MCT-21-0073.
- Kaiser J. Forced into battle. Science. 2020 May 29;368(6494):930–33. PubMed PMID: 32467373. doi:10.1126/science.368.6494.930.
- Wu Z, Cheung NV. T cell engaging bispecific antibody (T-BsAb): from technology to therapeutics. Pharmacol Ther. 2018 Feb;182:161–75 . PubMed PMID: 28834699; PubMed Central PMCID: PMCPMC5785550. doi:10.1016/j.pharmthera.2017.08.005.
- Bacac M, Klein C, CEA TCB: UP. A novel head-to-tail 2:1 T cell bispecific antibody for treatment of CEA-positive solid tumors. Oncoimmunology. 2016 Aug 5;5(8):e1203498. PubMed PMID: 27622073; PubMed Central PMCID: PMCPMC5007959. doi:10.1080/2162402X.2016.1203498.
- Schlothauer T, Herter S, Koller CF, et al. Novel human IgG1 and IgG4 Fc-engineered antibodies with completely abolished immune effector functions. Protein Eng Des Sel. 2016 Oct;29(10):457–66. 10.1093/protein/gzw040: PubMed PMID: 27578889
- Lehmann S, Perera R, Grimm HP, et al. In Vivo Fluorescence Imaging of the Activity of CEA TCB, a novel T-cell bispecific antibody, reveals highly specific tumor targeting and fast induction of T-cell-mediated tumor killing. Clin Cancer Res. 2016 Sep 1;22(17):4417–27. PubMed PMID: 27117182. doi:10.1158/1078-0432.CCR-15-2622
- Van De Vyver AJ, Weinzierl T, Eigenmann MJ, et al. Predicting tumor killing and T-cell activation by T-cell bispecific antibodies as a function of target expression: combining in vitro experiments with systems modeling. Mol Cancer Ther. 2021 Feb;20(2):357–66. 10.1158/1535-7163.MCT-20-0269: PubMed PMID: 33298591
- Gonzalez-Exposito R, Semiannikova M, Griffiths B, et al. CEA expression heterogeneity and plasticity confer resistance to the CEA-targeting bispecific immunotherapy antibody cibisatamab (CEA-TCB) in patient-derived colorectal cancer organoids. J Immunother Cancer. 2019 Apr 15;7(1):101. PubMed PMID: 30982469; PubMed Central PMCID: PMCPMC6463631. doi:10.1186/s40425-019-0575-3
- Teijeira A, Etxeberria I, Ponz-Sarvise M, et al. Immunotherapy of cancer visualized by live microscopy: seeing is believing. Clin Cancer Res. 2016 Sep 1;22(17):4277–79. PubMed PMID: 27330056. doi:10.1158/1078-0432.CCR-16-1072
- Sam J, Colombetti S, Fauti T, et al. Combination of T-cell bispecific antibodies With PD-L1 checkpoint inhibition elicits superior anti-tumor activity. Front Oncol. 2020;10:575737. PubMed PMID: 33330050; PubMed Central PMCID: PMCPMC7735156. doi:10.3389/fonc.2020.575737.
- Dudal S, Hinton H, Giusti AM, et al. Application of a MABEL approach for a T-cell-bispecific monoclonal antibody: CEA TCB. J Immunother. 2016 Sep;39(7):279–89. 10.1097/CJI.0000000000000132: PubMed PMID: 27404941
- Kamperschroer C, Shenton J, Lebrec H, et al. Summary of a workshop on preclinical and translational safety assessment of CD3 bispecifics. J Immunotoxicol. 2020 Dec;17(1):67–85. 10.1080/1547691X.2020.1729902: PubMed PMID: 32100588
- Cremasco F, Menietti E, Speziale D, et al. Cross-linking of T cell to B cell lymphoma by the T cell bispecific antibody CD20-TCB induces IFNgamma/CXCL10-dependent peripheral T cell recruitment in humanized murine model. PLoS One. 2021;16(1):e0241091. PubMed PMID: 33406104; PubMed Central PMCID: PMCPMC7787458 and SC declare ownership of Roche stocks. MP, PU, MB, CK, SC, JS are inventors on patent applications related to the CD20-TCB. This affiliation and patent participation do not alter our adherence to PLOS ONE policies on sharing data and materials. doi:10.1371/journal.pone.0241091.
- Prakash A, Diefenbach CS. Immunity War: a Novel Therapy for Lymphoma Using T-cell Bispecific Antibodies. Clin Cancer Res. 2018 Oct 1;24(19):4631–32. PubMed PMID: 29884742. doi:10.1158/1078-0432.CCR-18-1363.
- Hutchings M, Morschhauser F, Iacoboni G, et al. Glofitamab, a novel, bivalent CD20-Targeting T-cell-engaging bispecific antibody, induces durable complete remissions in relapsed or refractory B-cell lymphoma: a phase I trial. J Clin Oncol. 2021 Jun 20;39(18):1959–70. PubMed PMID: 33739857; PubMed Central PMCID: PMCPMC8210975. doi:10.1200/JCO.20.03175
- Killock D. Engaging results with glofitamab. Nat Rev Clin Oncol. 2021 May;18(5):257. PubMed PMID: 33828233. doi:10.1038/s41571-021-00510-3.
- Minson A, Glofitamab DM. CD20-TCB bispecific antibody. Leuk Lymphoma. 2021 Jul 15. 1–11. PubMed PMID: 34263696. doi:10.1080/10428194.2021.1953016.
- Liddy N, Bossi G, Adams KJ, et al. Monoclonal TCR-redirected tumor cell killing. Nat Med. 2012 Jun;18(6):980–87. 10.1038/nm.2764: PubMed PMID: 22561687
- Arenas EJ, Martinez-Sabadell A, Rius Ruiz I, et al. Acquired cancer cell resistance to T cell bispecific antibodies and CAR T targeting HER2 through JAK2 down-modulation. Nat Commun. 2021 Feb 23;12(1):1237. PubMed PMID: 33623012; PubMed Central PMCID: PMCPMC7902842. doi:10.1038/s41467-021-21445-4
- Loo Yau H, Bell E, Ettayebi I, et al. DNA hypomethylating agents increase activation and cytolytic activity of CD8(+) T cells. Mol Cell. 2021 Apr 1;81(7):1469–1483e8. PubMed PMID: 33609448. doi:10.1016/j.molcel.2021.01.038
- Rius Ruiz I, Vicario R, Morancho B, et al. p95HER2-T cell bispecific antibody for breast cancer treatment. Sci Transl Med. 2018 Oct 3;10(461):eaat1445. PubMed PMID: 30282693; PubMed Central PMCID: PMCPMC6498439. doi:10.1126/scitranslmed.aat1445
- Geiger M, Stubenrauch KG, Sam J, et al. Protease-activation using anti-idiotypic masks enables tumor specificity of a folate receptor 1-T cell bispecific antibody. Nat Commun. 2020 Jun 24;11(1):3196. PubMed PMID: 32581215; PubMed Central PMCID: PMCPMC7314773. doi:10.1038/s41467-020-16838-w
- Leclercq G, Haegel H, Schneider A, et al. Src/lck inhibitor dasatinib reversibly switches off cytokine release and T cell cytotoxicity following stimulation with T cell bispecific antibodies. J Immunother Cancer. 2021 Jul 9;9(7):e002582. PubMed PMID: 34326166. doi:10.1136/jitc-2021-002582
- Karches CH, Benmebarek MR, Schmidbauer ML, et al. Bispecific antibodies enable synthetic agonistic receptor-transduced t cells for tumor immunotherapy. Clin Cancer Res. 2019 Oct 1;25(19):5890–900. PubMed PMID: 31285373. doi:10.1158/1078-0432.CCR-18-3927
- Darowski D, Kobold S, Jost C, et al. Combining the best of two worlds: highly flexible chimeric antigen receptor adaptor molecules (CAR-adaptors) for the recruitment of chimeric antigen receptor T cells. MAbs. 2019;11(4):621–31. PubMed PMID: 30892136; PubMed Central PMCID: PMCPMC6601549. doi:10.1080/19420862.2019.1596511. May/Jun.
- Kobold S, Steffen J, Chaloupka M, et al. Selective bispecific T cell recruiting antibody and antitumor activity of adoptive T cell transfer. J Natl Cancer Inst 2015 Jan;107(1):364. 10.1093/jnci/dju364: PubMed PMID: 25424197
- Griessinger CM, Olafsen T, Mascioni A, et al. The PET-tracer (89) Zr-Df-IAB22M2CEnables monitoring of intratumoral CD8 T-cell infiltrates in tumor-bearing humanized mice after t-cell bispecific antibody treatment. Cancer Res. 2020 Jul 1;80(13):2903–13. PubMed PMID: 32409308. doi:10.1158/0008-5472.CAN-19-3269
- Villanueva MT. Building on bispecifics. Nat Rev Drug Discov. 2019 Jul;18(8):582. PubMed PMID: 31367057. doi:10.1038/d41573-019-00117-5.
- Trub M, Uhlenbrock F, Claus C, et al. Fibroblast activation protein-targeted-4-1BB ligand agonist amplifies effector functions of intratumoral T cells in human cancer. J Immunother Cancer. 2020 Jul;8(2):e000238. 10.1136/jitc-2019-000238: PubMed PMID: 32616554; PubMed Central PMCID: PMCPMC7333869
- Gauthier L, Morel A, Anceriz N, et al. Multifunctional natural killer cell engagers targeting NKp46 trigger protective tumor immunity. Cell. 2019 Jun 13;177(7):1701–1713e16. PubMed PMID: 31155232. doi:10.1016/j.cell.2019.04.041
- Zhao L, Xie F, Tong X, et al. Combating non-Hodgkin lymphoma by targeting both CD20 and HLA-DR through CD20-243 CrossMab. MAbs. 2014 May-Jun;6(3):740–48. 10.4161/mabs.28613: PubMed PMID: 24670986; PubMed Central PMCID: PMCPMC4011918
- Rajendran S, Li Y, Ngoh E, et al. Development of a bispecific antibody targeting CD30 and CD137 on hodgkin and reed-sternberg cells. Front Oncol. 2019;9:945. PubMed PMID: 31616638; PubMed Central PMCID: PMCPMC6768943. doi:10.3389/fonc.2019.00945.
- Du K, Li Y, Liu J, et al. A bispecific antibody targeting GPC3 and CD47 induced enhanced antitumor efficacy against dual antigen-expressing HCC. Mol Ther. 2021 Apr 7;29(4):1572–84. PubMed PMID: 33429083; PubMed Central PMCID: PMCPMC8058486. doi:10.1016/j.ymthe.2021.01.006
- Zhao L, Tong Q, Qian W, et al. Eradication of non-Hodgkin lymphoma through the induction of tumor-specific T-cell immunity by CD20-Flex BiFP. Blood. 2013 Dec 19;122(26):4230–36. PubMed PMID: 24178967. doi:10.1182/blood-2013-04-496554
- Panina AA, Rybchenko VS, Solopova ON, et al. Recombinant bispecific antibodies to the human erbb2 receptor and interferon-beta. Acta Naturae. 2020 Apr-Jun;12(2):95–104. 10.32607/actanaturae.10903: PubMed PMID: 32742732; PubMed Central PMCID: PMCPMC7385087
- Montefiori DC. Bispecific Antibodies Against HIV. Cell. 2016 Jun 16;165(7):1563–64. PubMed PMID: 27315470. doi:10.1016/j.cell.2016.06.004.
- Kingwell K. Infectious diseases: two-pronged attack on HIV. Nat Rev Immunol. 2016 Jul 27;16(8):465. PubMed PMID: 27461148. doi:10.1038/nri.2016.87.
- Asokan M, Rudicell RS, Louder M, et al. Bispecific Antibodies Targeting Different Epitopes on the HIV-1 Envelope Exhibit Broad and Potent Neutralization. J Virol. 2015 Dec;89(24):12501–12. 10.1128/JVI.02097-15: PubMed PMID: 26446600; PubMed Central PMCID: PMCPMC4665248
- Galimidi RP, Klein JS, Politzer MS, et al. Intra-spike crosslinking overcomes antibody evasion by HIV-1. Cell. 2015 Jan 29;160(3):433–46. PubMed PMID: 25635457; PubMed Central PMCID: PMCPMC4401576. doi:10.1016/j.cell.2015.01.016
- Bournazos S, Gazumyan A, Seaman MS, et al. Bispecific Anti-HIV-1 antibodies with enhanced breadth and potency. Cell. 2016 Jun 16;165(7):1609–20. PubMed PMID: 27315478; PubMed Central PMCID: PMCPMC4970321. doi:10.1016/j.cell.2016.04.050
- Khan SN, Sok D, Tran K, et al. Targeting the HIV-1 spike and coreceptor with bi- and trispecific antibodies for single-component broad inhibition of entry. J Virol. 2018 Sep 15;92(18): 10.1128/JVI.00384-18. PubMed PMID: 29976677; PubMed Central PMCID: PMCPMC6146690.
- Moshoette T, Ali SA, Papathanasopoulos MA, et al. Engineering and characterising a novel, highly potent bispecific antibody iMab-CAP256 that targets HIV-1. Retrovirology. 2019 Nov 8;16(1):31. PubMed PMID: 31703699; PubMed Central PMCID: PMCPMC6842167. doi:10.1186/s12977-019-0493-y
- Davis-Gardner ME, Alfant B, Weber JA, et al. A Bispecific Antibody That Simultaneously Recognizes the V2- and V3-Glycan Epitopes of the HIV-1 envelope glycoprotein is broader and more potent than its parental antibodies. mBio. 2020 Jan 14;11(1): 10.1128/mBio.03080-19. PubMed PMID: 31937648; PubMed Central PMCID: PMCPMC6960291.
- Guttman M, Padte NN, Huang Y, et al. The influence of proline isomerization on potency and stability of anti-HIV antibody 10E8. Sci Rep. 2020 Aug 31;10(1):14313. PubMed PMID: 32868832; PubMed Central PMCID: PMCPMC7458915. doi:10.1038/s41598-020-71184-7
- Wagh K, Seaman MS, Zingg M, et al. Potential of conventional & bispecific broadly neutralizing antibodies for prevention of HIV-1 subtype A, C & D infections. PLoS Pathog. 2018 Mar;14(3):e1006860. 10.1371/journal.ppat.1006860: PubMed PMID: 29505593; PubMed Central PMCID: PMCPMC5854441
- Padte NN, Yu J, Huang Y, et al. Engineering multi-specific antibodies against HIV-1. Retrovirology. 2018 Aug 29;15(1):60. PubMed PMID: 30157871; PubMed Central PMCID: PMCPMC6114543. doi:10.1186/s12977-018-0439-9
- Wang J, Bardelli M, Espinosa DA, et al. A human bi-specific antibody against zika virus with high therapeutic potential. Cell. 2017 Sep 21;171(1):229–241e15. PubMed PMID: 28938115; PubMed Central PMCID: PMCPMC5673489. doi:10.1016/j.cell.2017.09.002
- De Gasparo R, Pedotti M, Simonelli L, et al. Bispecific IgG neutralizes SARS-CoV-2 variants and prevents escape in mice. Nature. 2021 May;593(7859):424–28. 10.1038/s41586-021-03461-y: PubMed PMID: 33767445
- Jette CA, Cohen AA, Gnanapragasam PNP, et al. Broad cross-reactivity across sarbecoviruses exhibited by a subset of COVID-19 donor-derived neutralizing antibodies. bioRxiv. 2021 Apr 26 [ PubMed PMID: 33948592; PubMed Central PMCID: PMCPMC8095199]. doi:10.1101/2021.04.23.441195.
- Taylor PC, Williams RO. Combination cytokine blockade: the way forward in therapy for rheumatoid arthritis? Arthritis Rheumatol. 2015 Jan;67(1):14–16. PubMed PMID: 25302944. doi:10.1002/art.38893.
- Buckland J. Rheumatoid arthritis: anti-TNF and anti-IL-17 antibodies–better together! Nat Rev Rheumatol. 2014 Dec;10(12):699. PubMed PMID: 25348041. doi:10.1038/nrrheum.2014.183.
- Fischer JA, Hueber AJ, Wilson S, et al. Combined inhibition of tumor necrosis factor alpha and interleukin-17 as a therapeutic opportunity in rheumatoid arthritis: development and characterization of a novel bispecific antibody. Arthritis Rheumatol. 2015 Jan;67(1):51–62. 10.1002/art.38896: PubMed PMID: 25303306
- Xu T, Ying T, Wang L, et al. A native-like bispecific antibody suppresses the inflammatory cytokine response by simultaneously neutralizing tumor necrosis factor-alpha and interleukin-17A. Oncotarget. 2017 Oct 10;8(47):81860–72. PubMed PMID: 29137228; PubMed Central PMCID: PMCPMC5669854. doi:10.18632/oncotarget.19899
- Heier JS, Singh RP, Wykoff CC, et al. The angiopoietin/tie pathway in retinal vascular diseases: a Review. Retina. 2021 Jan 1;41(1):1–19. PubMed PMID: 33136975. doi:10.1097/IAE.0000000000003003
- Hussain RM, Neiweem AE, Kansara V, et al. Tie-2/Angiopoietin pathway modulation as a therapeutic strategy for retinal disease. Expert Opin Investig Drugs. 2019 Oct;28(10):861–69. 10.1080/13543784.2019.1667333: PubMed PMID: 31513439
- Fleckenstein M, Keenan TDL, Guymer RH, et al. Age-related macular degeneration. Nat Rev Dis Primers. 2021 May 6;7(1):31. PubMed PMID: 33958600. doi:10.1038/s41572-021-00265-2
- Hussain RM, Shaukat BA, Ciulla LM, et al. Vascular endothelial growth factor antagonists: promising players in the treatment of neovascular age-related macular degeneration. Drug Des Devel Ther. 2021;15:2653–65. PubMed PMID: 34188445; PubMed Central PMCID: PMCPMC8232378. doi:10.2147/DDDT.S295223.
- Sharma A, Kumar N, Kuppermann BD, et al. Faricimab: expanding horizon beyond VEGF. Eye (Lond). 2020 May;34(5):802–04. 10.1038/s41433-019-0670-1: PubMed PMID: 31695160; PubMed Central PMCID: PMCPMC7182558
- Jakubiak P, Alvarez-Sanchez R, Fueth M, et al. Ocular pharmacokinetics of intravitreally injected protein therapeutics: comparison among standard-of-care formats. Mol Pharm. 2021 Jun 7;18(6):2208–17. PubMed PMID: 34014104. doi:10.1021/acs.molpharmaceut.0c01218
- Joussen AM, Ricci F, Paris LP, et al. Angiopoietin/Tie2 signalling and its role in retinal and choroidal vascular diseases: a review of preclinical data. Eye (Lond). 2021 May;35(5):1305–16. 10.1038/s41433-020-01377-x: PubMed PMID: 33564135; PubMed Central PMCID: PMCPMC8182896
- Nicolo M, Ferro Desideri L, Vagge A, et al. Faricimab: an investigational agent targeting the Tie-2/angiopoietin pathway and VEGF-A for the treatment of retinal diseases. Expert Opin Investig Drugs. 2021 Mar;30(3):193–200. 10.1080/13543784.2021.1879791: PubMed PMID: 33471572
- Foxton RH, Uhles S, Gruner S, et al. Efficacy of simultaneous VEGF-A/ANG-2 neutralization in suppressing spontaneous choroidal neovascularization. EMBO Mol Med. 2019 May;11(5): 10.15252/emmm.201810204. PubMed PMID: 31040126; PubMed Central PMCID: PMCPMC6505683.
- Wolf A, Langmann T. Anti-VEGF-A/ANG2 combotherapy limits pathological angiogenesis in the eye: a replication study. EMBO Mol Med. 2019 May;11(5): 10.15252/emmm.201910362. PubMed PMID: 31040129; PubMed Central PMCID: PMCPMC6505573.
- Chakravarthy U, Bailey C, Brown D, et al. Phase I trial of anti-vascular endothelial growth factor/anti-angiopoietin 2 bispecific antibody rg7716 for neovascular age-related macular degeneration. Ophthalmol Retina. 2017 Nov - Dec;1(6):474–85. 10.1016/j.oret.2017.03.003: PubMed PMID: 31047438
- Sahni J, Patel SS, Dugel PU, et al. Simultaneous inhibition of angiopoietin-2 and vascular endothelial growth factor-a with faricimab in diabetic macular edema: BOULEVARD phase 2 Randomized Trial. Ophthalmology. 2019 Aug;126(8):1155–70. 10.1016/j.ophtha.2019.03.023: PubMed PMID: 30905643
- Sahni J, Dugel PU, Patel SS, et al. Safety and efficacy of different doses and regimens of faricimab vs ranibizumab in neovascular age-related macular degeneration: the avenue phase 2 randomized clinical trial. JAMA Ophthalmol. 2020 Sep 1;138(9):955–63. PubMed PMID: 32729888; PubMed Central PMCID: PMCPMC7393587. doi:10.1001/jamaophthalmol.2020.2685
- Khanani AM, Patel SS, Ferrone PJ, et al. Efficacy of every four monthly and quarterly dosing of faricimab vs ranibizumab in neovascular age-related macular degeneration: the stairway phase 2 randomized clinical trial. JAMA Ophthalmol. 2020 Sep 1;138(9):964–72. PubMed PMID: 32729897; PubMed Central PMCID: PMCPMC7489851. doi:10.1001/jamaophthalmol.2020.2699
- De La Huerta I, Kim SJ, Sternberg P Jr. Faricimab combination therapy for sustained efficacy in neovascular age-related macular degeneration. JAMA Ophthalmol. 2020 Sep 1;138(9):972–73. PubMed PMID: 32729885. doi:10.1001/jamaophthalmol.2020.2723.
- Sharma A, Kumar N, Parachuri N, et al. Faricimab: two in the bush is proving better than one in the hand? Ocul Immunol Inflamm. 2021 Jul;9:1–3. doi:10.1080/09273948.2021.1931350. PubMed PMID: 34242102.
- Terstappen GC, Meyer AH, Bell RD, et al. Strategies for delivering therapeutics across the blood-brain barrier. Nat Rev Drug Discov. 2021 May;20(5):362–83. 10.1038/s41573-021-00139-y: . PubMed PMID: 33649582
- Bohrmann B, Baumann K, Benz J, et al. Gantenerumab: a novel human anti-Abeta antibody demonstrates sustained cerebral amyloid-beta binding and elicits cell-mediated removal of human amyloid-beta. J Alzheimers Dis. 2012;28(1):49–69. PubMed PMID: 21955818. doi:10.3233/JAD-2011-110977.
- Joshi KK, Phung W, Han G, et al. Elucidating heavy/light chain pairing preferences to facilitate the assembly of bispecific IgG in single cells. MAbs. 2019 Oct;11(7):1254–65. 10.1080/19420862.2019.1640549: PubMed PMID: 31286843; PubMed Central PMCID: PMCPMC6748609
- Gong D, Riley TP, Bzymek KP, et al. Rational selection of building blocks for the assembly of bispecific antibodies. MAbs. 2021 Jan-Dec;13(1):1870058. 10.1080/19420862.2020.1870058: PubMed PMID: 33397191; PubMed Central PMCID: PMCPMC7808324
- Tustian AD, Endicott C, Adams B, et al. Development of purification processes for fully human bispecific antibodies based upon modification of protein A binding avidity. MAbs. 2016 May-Jun;8(4):828–38. 10.1080/19420862.2016.1160192: PubMed PMID: 26963837; PubMed Central PMCID: PMCPMC4966828
- Bogen JP, Carrara SC, Fiebig D, et al. Expeditious generation of biparatopic common light chain antibodies via chicken immunization and yeast display screening. Front Immunol. 2020;11:606878. PubMed PMID: 33424853; PubMed Central PMCID: PMCPMC7786285. doi:10.3389/fimmu.2020.606878.
- Bogen JP, Carrara SC, Fiebig D, et al. Design of a Trispecific Checkpoint Inhibitor And Natural Killer Cell Engager Based On A 2 + 1 Common Light Chain Antibody Architecture. Front Immunol. 2021;12:669496. PubMed PMID: 34040611; PubMed Central PMCID: PMCPMC8141644. doi:10.3389/fimmu.2021.669496.
- Bogen JP, Storka J, Yanakieva D, et al. Isolation of common light chain antibodies from immunized chickens using yeast biopanning and fluorescence-activated cell sorting. Biotechnol J. 2021 Mar;16(3):e2000240. 10.1002/biot.202000240: PubMed PMID: 32914549
- Chiu D, Tavare R, Haber L, et al. A PSMA-Targeting CD3 Bispecific Antibody Induces Antitumor Responses that Are Enhanced by 4-1BB Costimulation. Cancer Immunol Res. 2020 May;8(5):596–608. 10.1158/2326-6066.CIR-19-0518: PubMed PMID: 32184296
- Crawford A, Haber L, Kelly MP, et al. A Mucin 16 bispecific T cell-engaging antibody for the treatment of ovarian cancer. Sci Transl Med. 2019 Jun 19;11(497): 10.1126/scitranslmed.aau7534. PubMed PMID: 31217340.
- Skokos D, Waite JC, Haber L, et al. A class of costimulatory CD28-bispecific antibodies that enhance the antitumor activity of CD3-bispecific antibodies. Sci Transl Med. 2020 Jan 8;12:525. 10.1126/scitranslmed.aaw7888: PubMed PMID: 31915305
- Waite JC, Wang B, Haber L, et al. Tumor-targeted CD28 bispecific antibodies enhance the antitumor efficacy of PD-1 immunotherapy. Sci Transl Med. 2020 Jun 24;12(549): 10.1126/scitranslmed.aba2325. PubMed PMID: 32581132.
- Geuijen C, Tacken P, Wang LC, et al. A human CD137xPD-L1 bispecific antibody promotes anti-tumor immunity via context-dependent T cell costimulation and checkpoint blockade. Nat Commun. 2021 Jul 21;12(1):4445. PubMed PMID: 34290245. doi:10.1038/s41467-021-24767-5
- Geuijen CAW, De Nardis C, Maussang D, et al. Unbiased combinatorial screening identifies a bispecific igg1 that potently inhibits her3 signaling via her2-guided ligand blockade. Cancer Cell. 2018 May 14;33(5):922–936e10. PubMed PMID: 29763625. doi:10.1016/j.ccell.2018.04.003
- Van Loo PF, Hangalapura BN, Thordardottir S, et al. MCLA-117, a CLEC12AxCD3 bispecific antibody targeting a leukaemic stem cell antigen, induces T cell-mediated AML blast lysis. Expert Opin Biol Ther. 2019 Jul;19(7):721–33. 10.1080/14712598.2019.1623200: PubMed PMID: 31286786
- Shiraiwa H, Narita A, Kamata-Sakurai M, et al. Engineering a bispecific antibody with a common light chain: identification and optimization of an anti-CD3 epsilon and anti-GPC3 bispecific antibody, ERY974. Methods. 2019 Feb 1;154:10–20. 10.1016/j.ymeth.2018.10.005: PubMed PMID: 30326272
- Bonisch M, Sellmann C, Maresch D, et al. Novel CH1: cLinterfaces that enhance correct light chain pairing in heterodimeric bispecific antibodies. Protein Eng Des Sel. 2017 Sep 1;30(9):685–96. PubMed PMID: 28981885; PubMed Central PMCID: PMCPMC5914326. doi:10.1093/protein/gzx044
- Dillon M, Yin Y, Zhou J, et al. Efficient production of bispecific IgG of different isotypes and species of origin in single mammalian cells. MAbs. 2017;9(2):213–30. PubMed PMID: 27929752; PubMed Central PMCID: PMCPMC5297516. doi:10.1080/19420862.2016.1267089. Feb/Mar.