
ABSTRACT
Carbonic anhydrase (CA)-IX is an extracellular enzyme that is essential in the adaptation of tumor cells to their increasingly more hypoxic and acidic microenvironment. Within the family of carbonic anhydrases, CA-IX is unique in that it is the only CA with an N-terminal intrinsically disordered region (IDR) containing a proteoglycan (PG)-like domain. This PG-like IDR has been described to be instrumental in CA-IX’s enzyme activity, as well as tumor cell motility and invasion. We have characterized the antibody–epitope interactions of two novel and unique antibodies (11H9 and 12H8) that are specific for the human CA-IX’s IDR. Binding interactions of these antibodies to the intact IDR were studied by surface plasmon resonance and high-resolution nuclear magnetic resonance (NMR) spectroscopy, while the specific epitopes were determined by both NMR and yeast surface display (YSD). Our data show that 12H8 binds to the N-terminus of CA-IX, while 11H9 has a high affinity for an epitope located in the central region of the IDR containing three GEEDLP repeats in a manner that is different from the previously described M75 antibody. Titration NMR spectroscopy using CA-IX’s entire IDR in addition identified a secondary epitope of 11H9 at the beginning of the PG-like domain that remains exposed and available for further binding events after the engagement at its primary epitope at the center of the PG-like domain. Transverse relaxation optimized NMR spectroscopy of 11H9-F(Ab) in complex with the CA-IX IDR outlines structural rigidification of a linear epitope, while the rest of the IDR remains largely unstructured upon complex formation. This study illustrates how high-resolution NMR and YSD are used as complementary tools for a comprehensive characterization of antibody–epitope interactions involving intrinsically unstructured antigen domains with highly repetitive sequences.
Introduction
Carbonic anhydrases (CA) are a family of 15 ubiquitous enzymes that catalyze the reversible hydration of CO2, resulting in the generation of HCO3− and H+ with differing efficiency in addition to a variety of other reactions. CAs can be found intracellularly in several compartments and attached to cells via a transmembrane domain or glycosylphosphatidylinositol anchor. When secreted,Citation1,Citation2 CAs are involved in pH homeostasis of normal cells and tissues and some are of critical importance for the survival of cancer cells.Citation3,Citation4 As tumors grow the diffusion distances to the nearest blood vessels increase, and this not only limits access to oxygen and nutrients but also prevents the proper removal of metabolic waste. Low oxygen levels cause inhibition of oxidative phosphorylation and tumor cells survive by switching to a glycolytic metabolism, resulting in the intracellular accumulation and subsequent extrusion of lactate, which in turn leads to an acidic extracellular pH (pHe). Acidification of the tumor microenvironment (TME), also known as acidosis, disrupts the intracellular pH (pHi) homeostasis, which is detrimental to many cellular functions. Tumor cells adapt to acidosis by activating a very efficient pH-regulating system that uses CAs to prevent intracellular acidification. The hypoxia-driven upregulation of CA-IX together with the activation and upregulation of acid/base transporters (i.e., Na+/H+ exchangers, Na+/HCO3− cotransporters, anion exchangers, and monocarboxylate transporters) form very efficient transport metabolons that are critical for maintaining the proper acid/base balance in tumor cells.Citation5
Of the three exofacial CAs (CA-IV, CA-VII, and CA-IX), CA-IX is structurally unique in that it contains an N-terminal proteoglycan (PG)-like domain (residues 53–111) rich in negatively charged amino acids. It has been suggested that this PG-like domain aids the catalytic efficiency of CA-IX in the acidic microenvironmentCitation6 by acting as a proton buffer in solid tumors that optimizes the hydration of CO2.Citation7 The functional importance of CA-IX’s PG-like domain is further illustrated by the reduced adhesion and increased spreading of cancer cells expressing a mutant form of CA-IX that lacks the entire PG-like domain, or in the presence of antibodies that target a specific region of the PG-like domain.Citation8–10
The primary sequence of the PG-like domain contains a number of prolines which delineate a series of negatively charged glutamic and aspartic acids. This sequence pattern is very similar to that of the keratan sulfate enriched region of aggrecan, a large proteoglycan, hence the name PG-like domain.Citation11 It is interesting to note that a recent study identified CA-IX’s PG-like domain as a novel member in a large class of intrinsically disordered proteins and protein regions (IDPs/IDRs) that are devoid of a folded three-dimensional (3D) structure.Citation12 Another feature of CA-IX’s PG-like domain is the presence of a tandem repeat of three identical GEEDLP motifs, which is preceded by four motifs that are highly homologous to GEEDLPCitation11,Citation12 with a repetitive sequence that is also characteristic of many other IDPs/IDRs.Citation13,Citation14
In this study, we characterized the molecular interactions of two novel antibodies (11H9 and 12H8)Citation15 that specifically target the intrinsically disordered region of CA-IX (residues 37–136; see ). To this end we exploited surface plasmon resonance (SPR), yeast surface display (YSD) and NMR spectroscopy using double (Citation15N and 13C) and triple (Citation2H, 15N and 13C) isotope labeled recombinantly expressed CA-IX’s intact IDR to examine this IDR’s detailed molecular interactions and to determine the binding epitopes of the 11H9 and 12H8 monoclonal antibodies (mAbs). The data presented here identify the N-terminus of CA-IX’s IDR as the binding epitope for 12H8, while a region spanning two negatively charged GEEDLP tandem repeats at the center of the PG-like domain as the primary binding epitope for 11H9. Further titration studies using high-resolution NMR identified a secondary epitope located in the N-terminal region of the PG-like domain for the 11H9 mAb. Here we also show evidence that the 11H9 mAb is capable of simultaneously binding both epitopes in the CA-IX IDR only in an intermolecular fashion, leading to the formation of multi-molecular cross-linked antibody aggregates.
Table 1. Sequence of the ePG-1 polypeptide containing an N-terminal methionine (M) from the bacterial expression vector and a C-terminal linker-His-tag for purification purposes.
Results
11H9 and 12H8 CA-IX mAbs specifically bind distinctive epitopes within hCA-IX’s intrinsically disordered region
Antibodies 12H8 and 11H9 were among a pool of six novel mouse mAbs that were generated by immunization of mice with the entire extracellular domain of human CA-IX as an immunogen and selected for their potential as the antibody component of an antibody–drug conjugate.Citation15 MAbs 11H9 and 12H8 did not interfere with the catalytic activities of CA-IX and both exhibited specific binding to CA-IX’s N-terminal region as demonstrated through Pepscan evaluation.Citation15 To further characterize the binding epitopes of these two antibodies, we recombinantly produced the ePG-1 and ePG-2 polypeptides (; SupplTable S1) which cover the IDR of CA-IX. Before initiating NMR experiments, we evaluated the mAb binding characteristics of these polypeptides by SPR. First, we used two control antibodies: M75 mAb, which is known to bind CA-IX’s PG-like domain,Citation8 and 2D7, which is a catalytic domain-specific mAbCitation15,Citation16 and therefore should not bind. Our data ( and SupplFig.1A) clearly show the expected binding by M75 and the lack thereof by 2D7, indeed confirming that the mAb binding characteristics of CA-IX’s PG-like domain have been retained in the ePG-1 and ePG-2 polypeptides. We then evaluated the binding of the 11H9 and 12H8, in both a mAb and F(Ab) format and found that 11H9 binds both the ePG-1 () and ePG-2 (SupplFig. S1B) polypeptides. However, we also observed that 12H8 only binds the ePG-1 () and not the ePG-2 (SupplFig. S1C) polypeptide, indicating that 12H8 binding relies on the presence of four amino acids (PQRL) located at the exposed N-terminus of CA-IX that are present in the ePG-1 () but not in the ePG-2 (SupplTable S1) polypeptide. The 12H8 and 11H9 F(Ab) fragments display binding profiles similar to the full-length mAb equivalents ( and SupplFig. S1B, C). However, due to their monovalent character, F(Ab)s bind, in general, with reduced affinity resulting from their lack of avidity. This characteristic can be observed for both the 12H8 and 11H9 F(Ab) when binding to the ePG-1 polypeptide ( and ). Interestingly, the 11H9 F(Ab) has an apparently slow rate of dissociation from both surface-immobilized ePG-1 (, panel b) and ePG-2 (SupplFig. S1, panel b) in contrast to the 12H8 F(Ab) (, panel c).
Figure 1. SPR binding studies of the CA-IX mAbs and F(Ab)s to immobilized ePG-1 polypeptide. SPR sensorgrams of (a) mAb M75 and mAb 2D7, (b) mAb 11H9 and its F(Ab) fragment, and (c) mAb 12H8 and its F(Ab) fragment to the immobilized ePG-1 polypeptide. MAb 2D7, which binds to CA-IX’s catalytic domainCitation15, and mAb M75, which binds CA-IX’s PG domainCitation8 were used as a negative and positive control, respectively. MAbs were flowed at 1.23, 3.7, 11.1, and 33.3 nM whereas F(Ab)s were flowed at 1.23, 3.7, 11.1, 33.3 and 100 nM over the chip surface with immobilized ePG-1.
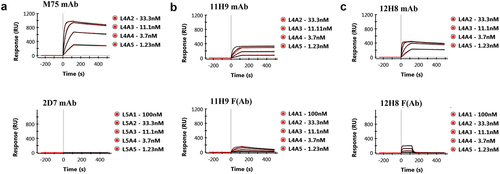
Table 2. Phenomenological kinetic values obtained by fitting a 1:1 binding model to the SPR data for the 11H9, 12H8, and M75 anti-CA-IX mAbs and the F(Ab)s of 11H9 and 12H8 to the ePG-1 and ePG-2 polypeptides.
In-depth characterization of the interactions of 12H8 and 11H9 by NMR spectroscopy
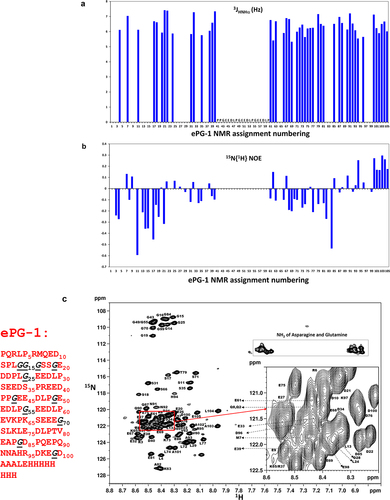
Using high-resolution NMR spectroscopy, the ePG-1 polypeptide was, as expected, found to be unfolded without any significant secondary structure preferences. For example, the 3JHN,HA coupling constants were determined to be 6–7 Hz for almost all (assigned) residues of the ePG-1 peptide (), which is indicative of random-coil conformations.Citation17 Proton-Citation15N heteronuclear NOEs (with 15N-labeled ePG-1) were very small (<0.3) and negative for many residues (), which are characteristic of unstructured/unfolded proteins.Citation18 Such a lack of folding for ePG-1 is not surprising given the highly negatively charged nature of the PG-like domain and the low sequence complexity of the N- and C-terminal regions beyond the charged PG-like sequence.Citation12 Regardless, detailed NMR 1H/Citation15N-heteronuclear single quantum coherence (HSQC) assignments of ePG-1 () and ePG-2 (SupplFig. S2A) showed a significant spectral resolution of 8 of the 11 Gly residues in the highly flexible conformations of these IDR fragments of CA-IX. Interestingly, the three remaining Gly residues with heavily overlapping NMR signals, i.e., Gly43, Gly49, and Gly55 (in ePG-1 NMR assignment numbering, see and SupplFig S2A), are located at the center of the PG-like domain, each at the beginning of the three tandem repeats of the GEEDLP sequence. Another feature of ePG-1 (and ePG-2) is the clustering of many residues at the center of the 1H/Citation15N HSQC spectra in which one finds some Asp and Glu residues and others, e.g., Asp21, Asp22, Leu24, Glu27, and Asp28, from the GEDDPLGEEDLP sequence containing the fourth GEEDLP motif ( and SupplFig. S2A).
Figure 2. Solution conformation of ePG-1 characterized by NMR using backbone 3JHN,HA coupling constants, 15N{Citation1H} heteronuclear NOE measurements and detailed resonance assignments. (a) essentially uniform 3JHN,HA coupling constants (of 6–7 Hz) indicate largely random coil conformations for the ePG-1 backbone. (b) Small 15N{Citation1H} heteronuclear NOE values with excursions to negative values are also characteristic of unfolded polypeptide chains for ePG-1. NMR data for the central region of ePG-1 could not be analyzed due to overlaps of the 1H/Citation15N-HSQC signals of the highly repetitive sequence PPGEEDLPGEEDLPGEEDLP. The companion 15N longitudinal (T1) and transverse (T2) relaxation data are shown in SupplFig. S3B and SupplFig. S3C, respectively. (c) assigned HSQC spectrum of ePG-1. The amino acid sequence of ePG-1 () is reproduced here with Pro (P37 of the hCA-IX sequence) as the first residue for the NMR assignment numbering. The italicized black font indicates the locations of the 11 Gly residues in the sequence of ePG-1. The inset details some signal assignments of the heavily overlapping region in the HSQC spectrum. A rectangular box outlines the side-chain amide HSQC signals of Asn and Gln residues. Two Gln residues, Q2 and Q8, and two Asn residues, N91 and N92 are located at the N-terminal and C-terminal regions of ePG-1, respectively.
Residue-specific interactions of the 12H8 and 11H9 mAbs with the CA-IX IDR were then followed by perturbations in the H/Citation15N-HSQC spectra of 15N-labeled ePG-1 (, SupplFig. S3 and SupplFig. S4) induced by adding the full-length antibodies and their respective F(Ab)s. Additions of the full-length 12H8 (SupplFig. S3A) or its F(Ab) (SupplFig. S3B) led to the disappearance of the same set of HSQC signals of ePG-1, demonstrating the equivalence of full-length 12H8 and its F(Ab) (). These disappearing H/Citation15N-HSQC signals were assignable to residues Leu4, Arg6/Asp10, Glu9, and Ser11 ( using NMR assignment numbering; or residues L40, R42, E45, D46, and S47 using CA-IX’s sequence numbering in the single-letter aa code) along with large HSQC peak shifts of residues Gly14, Gly15, and Gly16 (i.e., CA-IX residues G50, G51, and G52) ( and SupplFig. S3C). On the other hand, there are no backbone amide H/Citation15N-HSQC signals for residue Pro1 (which has an N-C bond in place of an amide NH) and for residues Gln2 and Arg3, whose amide protons have weak, and therefore undetectable, proton NMR signals due to fast exchange at neutral pH conditions.Citation19 Residues Gln2 and Gln8 also exhibited a strong response to 12H8 binding, involving their sidechain amide protons (). Binding of 12H8, therefore, is localized to the N-terminal region of ePG-1 (SupplFig. S3C) or within the CA-IX’s N-terminal sequence of PQRL40PRMQE45DSPLG50GG.
Figure 3. (a) 1H/Citation15N-HSQC spectrum of ePG-1 responding to 12H8-F(Ab) binding with assignments of ePG-1 signals that perturbed and disappeared (shown in black) in the 1:1 complex. The inset shows the detailed responses of the 11 Gly signals with the disappearance of the signals of G14 and G16 of free ePG-1 (black) along with the appearance of two new HSQC signals for the complex (red). Signal movements in response to 12H8 binding were assigned tentatively as indicated by arrows in the inset, producing the chemical shift perturbations for G14, G15 and Gl6 in decreasing amplitudes. (b) Gly region of ePG-1 HSQC responding to 11H9-F(Ab) binding; the left-most panels show the superposition of the complex spectra with that of free ePG-1 (black) exemplifying the response of G25 while the right panels detailing Gly signal intensity evolutions from free ePG-1 (black) to the 1:1 (red), 1:2 (blue) and 1:5 (green) complexes, respectively. (c) Ser region of the ePG-1 HSQC spectrum responding to excess 11H9-F(Ab) binding, i.e., From 1:1 (red) to 1:2 (blue) and to 1:5 (green) for the ePG-1/F(Ab) ratio, respectively; free ePG-1 spectra are shown in black.
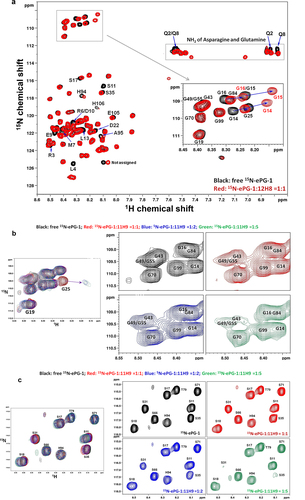
On the other hand, no significant perturbations were evident upon adding the 11H9 mAb to 15N-labeled ePG-1 (SupplFig. S3D). 11H9 F(Ab) titrations also induced limited perturbations in ePG-1 (SupplFig. S3E), mostly concentrated around the area of the first GEEDLP (residues Gly25-Pro30 in NMR assignment numbering; ), with the most evident signal displacements for Gly25 in the presence of excess amounts of F(Ab) beyond the 1:1 stoichiometry (). Interestingly, the signal cluster for residues Gly43, Gly49, and Gly55 has a substantially decreased intensity for the 1:1 complex, which became much more pronounced for the 1:2 ePG-1:F(Ab) and 1:5 complexes (). These are accompanied by identifiable signal perturbations for some assignable Glu, Asp, and Leu residues from the GEEDLP sequence (SupplFig. S3F). The main binding site for 11H9 may therefore lie in this central region of the PG-like domain with three tandem repeats of the same GEEDLP sequence which created heavy resonance overlaps (). After this higher-affinity site is occupied (i.e., at the 1:1 complex), excess amounts of 11H9 F(Ab) become available for binding to lower-affinity sites, as shown by resonance perturbations at Gly25 only under such conditions ( and SupplFig. S3F). Very importantly, the excess 11H9 F(Ab) also produced significant resonance perturbations for Ser31 and Ser35 and clear doubling of HSQC peaks for Ser17 and Ser18 (), all surrounding the first G25EEDLP30 sequence at the beginning of the PG-like domain.
TROSY NMR characterization of the ePG-1 IDR in complex with the 11H9 F(ab)
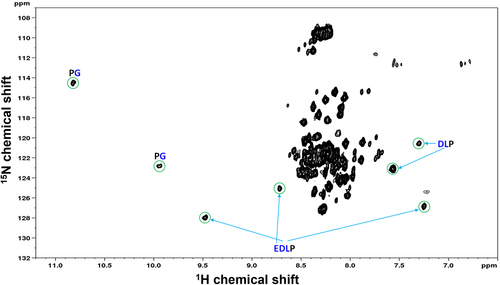
The observation of the 11H9-F(Ab)-induced perturbations at two sites in ePG-1, especially in the central region of the PG-like domain () motivated us to perform a more detailed examination on how three identical ‘GEEDLP’ tandem repeats may interact with the 11H9 antibody. The ePG-1 protein was therefore deuterated (substituting all non-exchangeable carbon attached protons with deuterium) and labeled with the 15N isotopes at the same time. This double-labeling strategy enables the use of TROSY NMR at high magnetic fields, which can detect rigidly structured residues in large complexes (molecular weight of >60 kDa),Citation20 such as ePG-1 bound to the 11H9-F(Ab). Eight additional signals (crosspeaks) appeared in the H/Citation15N-TROSY spectrum of Citation15N/Citation13C/Citation2H-labeled ePG-1 (the additional 13C-labeling for TROSY signal assignments, see Methods Section) in a 1:1 complex with the 11H9-F(Ab) (). Interestingly, the rest of the TROSY spectrum of the 11H9-F(Ab)/ePG-1 complex was essentially identical () to the 1H/Citation15N-HSQC spectrum of free ePG-1 (), as already shown in HSQC titration experiments with selectively perturbed residues (, SupplFig. S3D and SupplFig. S3E). In other words, a 1:1 complexation of the 11H9-F(Ab) does not dramatically influence the disordered conformations of ePG-1 except for those residues that form and surround the direct structural contact between 11H9 and its epitope residues in ePG-1. 3D TROSY-HNCACBCitation21 NMR data with 15N/Citation13C/Citation2H-labeled ePG-1 allowed the assignment of the few TROSY signals to the PGEEDLPG sequence, located within the central region of ePG-1 containing three identical GEEDLP repeats. Specifically, three well-separated TROSY signals were assigned to the E, D, and L residues of the EDLP unit, two assigned to D and L residues of the DLP sequence, and two as originating from two different PG motifs, all based on the unique 13Cα and 13Cβ chemical shifts and the inter-residue correlations in the TROSY-HNCACB spectrum. In other words, TROSY assignments establish that the primary binding epitope of 11H9 is indeed located at the sequence of PGEEDLPGEEDLP with two PG sequences, one EDLP unit and one DLP unit. Furthermore, the resolution of EDLP and DLP by TROSY shows that 11H9 F(Ab) binding to this epitope creates an asymmetry to the two essentially identical EDLP units in the tandem repeats of two GEEDLP motifs. There are still two partially overlapping sequences of PGEEDLPGEEDLP in PGEEDLPGEEDLPGEEDLP with three GEEDLP repeats, but NMR data alone cannot distinguish between these two potential epitopes.
Figure 4. Amide 1H/Citation15N-TROSY NMR spectrum of 13C/Citation15N/Citation2H-labeled ePG-1 in a 1:1 stoichiometric complex with the 11H9-F(Ab) fragment. Well-dispersed signals are labeled with the respective residue assignments in the triple-labeled ePG-1. It should be noted that the remaining signals in the rest of this TROSY spectrum are similar in chemical shift positions to the 1H/Citation15N-HSQC spectra of free ePG-1 (see ).
YSD epitope mapping of the 11H9 and 12H8 mAbs
Fragment scanning of the CA-IX N-terminal fragment: To gain further insights into the binding epitopes of the 11H9 and 12H8 mAbs, we examined the interactions of CA-IX’s N-terminal IDR with these and the M75 control antibody by using yeast S. cerevisiae displaying the full-length CA-IX IDR and overlapping peptides (). All three antibodies (11H9, 12H8, and M75) were able to bind the native and heat-denatured yeast-displayed full-length IDR, suggesting either linear/contiguous binding epitope(s) for these antibodies or the lack of a defined 3D structure for CA-IX’s N-terminal region, the latter of which is shown conclusively by NMR spectroscopy.
Figure 5. Epitope mapping by yeast surface display strategy. (a) Cartoon depicting the CA-IX N-terminal fragments that were expressed on the cell surface of yeast. (b) ELISA results showing the yeast-displayed CA-IX fragments binding to mAb 11H9 (red), 12H8 (blue) and M75 (gray).
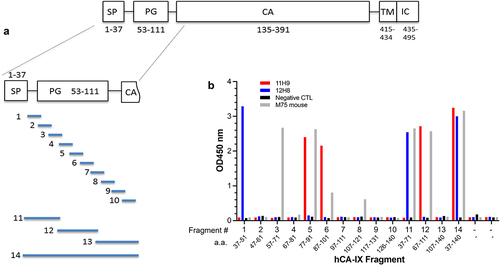
To obtain a higher resolution epitope map, peptide fragments of 15 amino acid (aa) residues (with a five aa tiling overlap) and covering residues 37–140 (Fragment #14) of hCA-IX were expressed on the yeast cell surface display (). Cell ELISA data ( and SupplTable S2) show that 12H8 binds to a distinct 15-aa epitope (Fragment #1; aa 37–51, PQRLPRMQEDSPLGG). 11H9, however, binds to an epitope whose sequence is shared by peptide fragments PPGEEDLPGEEDLPG (Fragment #5; aa 77–91) and EDLPGEEDLPEVKPK (Fragment #6; aa 87–101), while the commercial M75 antibody binds strongly to an epitope whose sequence is shared by peptides DDPLGEEDLPSEEDS (Fragment #3; aa 57–71) and PPGEEDLPGEEDLPG (Fragment #5; aa 77–91). Yeast display data also indicate some level of M75 binding with two additional peptide fragments, i.e., EDLPGEEDLPEVKPK (Fragment #6; aa 87–101) and SLKLEDLPTVEAPGD (Fragment #8; aa 107–121) with weaker ELISA signals ( and SupplTable S2). Such different binding characteristics of 11H9 and M75 indicate that these two antibodies recognize distinct epitopes, albeit with some degree of sequence similarity, especially in the central region of CA-IX’s PG-like domain, while 12H8 binds to a 15-aa peptide fragment at the N-terminal segment of CA-IX (). The commercial antibody M75, therefore, emerged the most promiscuous, having multiple epitopes, in the central region with its known linear epitope GEEDLPGCitation8, in the homologous repeat GEEDLPS preceding the primary epitope as well as two sequences GEEDLPE and KLEDLPT having four identical residues EDLP.
Figure 6. Fine epitope mapping using yeast surface display (YSD). (a) Summary of antibody binding of yeast-displayed 15-residue tiling fragments of the CA-IX N-terminal region, residues 37–140. The colored sequences (12H8, green; 11H9, red, M75, blue) are the fragments within CA-IX IDR that showed the highest binding to each of the indicated antibodies by yeast cell ELISA (see ). (b) Fine epitope mapping on 15-residue fragments that were identified to bind the antibodies through N-and C-terminal truncations to determine the “minimal” epitope followed by mutagenic alanine scan. Residues in large font and bold are those that do not tolerate alanine substitution indicating their critical role in antibody binding (see text for details).
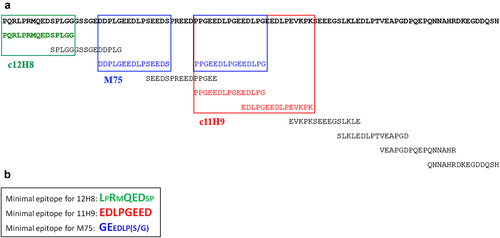
Fine epitope mapping of the CA-IX IDR-binding mAbs: We then carried out a ‘fine epitope mapping’ experiment, focusing on those fragments that had strong ELISA signals (), through truncation and mutagenic alanine scan (). The truncation analysis defined the “minimal” peptides that are both necessary and sufficient for the binding to these antibodies (): LPRMQEDSP for 12H8; EDLPGEED for 11H9; and GEEDLP(S/G) for M75. This analysis also showed that replacing one of residues L, R, Q, E, and D, in the 12H8 epitope LPRMQEDSP with an alanine abolished binding of 12H8 (as defined by yeast ELISA readings). Intriguingly, none of the amino acids of the 11H9 epitope EDLPGEED could tolerate substitution for alanine, whereas in the M75 epitope GEEDLP(S/G), alanine replacement of only two residues (G and E) results in loss of binding (). It should be noted that all five epitope residues identified by epitope fine mapping as essential for 12H8 binding also showed severe NMR HSQC signal perturbations leading to obvious loss of signal intensities (i.e., L40, R42, E45, and D46 and the sidechain amide of Q44, which correspond to NMR assignments of L4, R6, E9, D10, and the sidechain amide of Q8, respectively, in ) in NMR titration experiments. The primary epitope for 11H9 lies at the junction of two consecutive repeats of GEEDLP, which is in complete agreement with TROSY NMR data showing high structural rigidification of the PGEEDLPG sequence moiety upon binding of this mAb’s F(Ab) (). It is interesting that YSD data identified the high-affinity (primary) binding epitope EDLPGEED for 11H9 from the central region of the PG-like domain but did not locate the weaker and secondary binding site around the first G25EEDLP30 sequence which was identified by NMR titration experiments (Figure 3b/c and SupplFig. S3F). Indeed, 11H9 showed no binding to yeast cells displaying peptide 57–71 (Fragment #3), 67–81 (Fragment #4) and 37–71 (Fragment #12) (), despite that these peptide fragments contain DDPLGEED and EDLPSEED (Fragment #3 and #12), and EDSPREED (Fragment #4), each being a variant of the high-affinity epitope EDLPGEED.
Higher-order interactions between the CA-IX IDR and the mAb 11H9
With TROSY NMR data showing a largely unstructured IDR domain in a complex with 11H9 F((Ab) (), the question arises as to how the full-length, bivalent 11H9 mAb may interact with CA-IX’s IDR containing two 11H9 epitopes. The 1H/Citation15N-HSQC spectrum of 15N-labeled ePG-1 remained largely unperturbed in the presence of a less than stoichiometric concentration of the 11H9 mAb with a molar ratio of 6:1 for ePG-1 and 11H9, respectively (SuppFig. S3D), showing the lack of NMR exchange lineshapes due to the slow dissociation of the ePG-1/11H9 complex already identified in SPR studies ( and ). Very interestingly, decreasing the stoichiometry to 3:1 (PG:11H9) caused some intensity decreases of the HSQC signal cluster of the overlapped residues Gly43/Gly49/Gly55 (SupplFig. S4A) along with selective perturbations of some residues in the tail region of ePG-1, namely residues His94, Ala101, Ala102, Ala103, Leu104, and Glu105 (SupplFig. S4B). No further spectral changes were observed (spectra not shown) when a 3:1 complex was prepared with different concentrations, i.e., 65 µM and 22 µM for ePG-1 and 11H9, respectively, instead of the higher 160 uM and 54 uM concentrations, respectively, for the first 3:1 complex (i.e., as in SupplFig. S4A/B).
Surprisingly, a further decrease of the PG:11H9 molar ratio to (approximately) 2:1 led to turbidity of the complex solution (and precipitation, not shown) accompanied by more pronounced HSQC signal losses of the same Gly43/Gly49/Gly55 cluster and intensity losses for residues His94, Ala101, Ala102, Ala103, Leu104 and Glu105 (SupplFig. S4C/D). Together with these, the HSQC signal of Gly99 starts to broaden in this 2:1 complex (SupplFig. S4C). This is in dramatic contrast with the PG:12H8 complex, which had no signs of either sample turbidity or precipitations (SupplFig. S3A). With the molar ratio adjusted to 1:1, a more pronounced sample turbidity and precipitation occurred in an independently prepared sample of the PG/11H9 complex, which was accompanied by essentially a complete loss of the HSQC signal intensities for the Gly43/Gly49/Gly55 cluster and residues His94, Gly99, Ala101, Ala102, Ala103, Leu104, and Glu105 (SupplFig. S4E/F). In addition, residue Gly25 (which is part of the secondary epitope) exhibited HSQC signal perturbations in this 1:1 complex, along with now clearly visible perturbations on residues Gly19 (SupplFig. S4E) and Ser18, Ser31, Ser35, and Asp40, a cluster of residues around the secondary epitope identified in 11H9-F(Ab) titrations with F(Ab) excess (). This latter observation clearly indicates the availability of the secondary epitope in the soluble 2:1 (PG:11H9) molecular complex for interacting with the fraction of the free unoccupied 11H9 molecule ().
Figure 7. A schematic representation of the unique interactions of the 11H9 mAb with the ePG-1 polypeptide containing two epitopes. (a) the 2:1 (PG:11H9) complex is formed preferentially when the two combining sites of 11H9 are occupied by the high-affinity epitope EDLPGEED (shown in black circles), especially when the concentration of ePG-1 is higher than or equal to the 2:1 stoichiometry. With a concentration of ePG-1 at the 1:1 (PG:11H9) ratio, the same 2:1 (PG:11H9) complex is formed for 25% of the 11H9 concentration while 50% of 11H9 forms a 1:1 (PG:11H9) complex via the high-affinity epitope. The fraction of unoccupied 11H9 (25%) interacts with the low-affinity secondary epitope, e.g. DDPLGEED, exposed in the 2:1 (PG:11H9) complex (shown in pink), leading to differential HSQC signal perturbations of residues around this site (SupplFig. S4). Such a 2:1 (PG:11H9) complex can also be formed on SPR sensor chips with the ePG-1 polypeptide immobilized via its His-tag binding to surface-conjugated anti-His antibodies (see Materials and Methods), leading to the extremely slow rate of dissociation of 11H9 mAb from the sensor chip surface ( and SupplFig. S1B), (a) with a concentration of ePG-1 lower than the 2:1 stoichiometry, the insufficient spacing provided by ~ 22 residues between the high-affinity and low-affinity epitopes in ePG-1 favors intermolecular cross-linking or aggregation of the 1:1 PG:11H9 species, as evidenced by turbidity and precipitation in ePG-1/11H9 samples prepared at ~ 2:1 and ~ 1:1 ratios for the concentrations of ePG-1 and 11H9, respectively (see SupplFig. S4 for further details). The 2:1 (PG:11H9) complex species shown in (A) is also present in these (2:1 and 1:1) samples as detected by both 1H/Citation15N-HSQC (SupplFig. S4C/D/E/F) and by size-exclusion chromatography (SupplFig. S5) after removing the cross-linked and insoluble 11H9 aggregates by centrifugation.
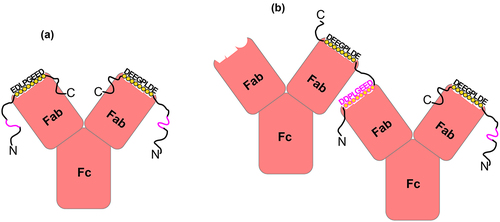
HSQC signal losses for the Gly43/Gly49/Gly55 cluster in the 1:1 and 2:1 (PG:11H9) samples mirror similar responses in titration experiments using the 11H9 F(Ab) (Figure 3b/c), especially with excess concentrations of the F(Ab) (), showing that the observed HSQC signals in these complexes of ePG-1 with 11H9 mAb are now dominated by 15N-labeled ePG-1 bound to the mAb instead of being largely from the uncomplexed ePG-1 as in the 6:1 (PG:11H9) complex (SupplFig. S3D). The intermediate sample with a 3:1 (PG:11H9) molar ratio, leaving only ~1/3 of ePG-1 in the uncomplexed state, also showed similar HSQC spectra as the free 15N-labeled ePG-1 (SupplFig. S4A/B), affirming the largely unstructured nature of ePG-1 whether bound to either the full-length 11H9 mAb or to its F(Ab) (). This lack of folding of ePG-1 whether free or bound to 11H9 can also lead to anomalous migration of PGCitation12 or the PG/11H9 complex through size-exclusion media (SupplFig. S5). PG bound to the 11H9 mAb does distinguish itself in that some tail residues, especially His94, Gly99, Ala101, Ala102, Ala103, Leu104, and Glu105 of ePG-1, appear to be involved in “binding” based on their HSQC signal perturbations (SupplFig. S4E/F). These added tail residues of ePG-1 (), however, may simply have altered conformational and/or dynamic properties in the large ePG-1/11H9 complex. As such, the 2:1 complex between ePG-1 and 11H9 () is also very likely what would be captured on SPR sensor chips with each immobilized anti-His antibody binding two ePG-1 molecules (or ePG2, SupplFig. S1), resulting in further slowed rates of dissociation of bound 11H9 mAbs (top panels of and SupplFig. S1B) in addition to avidity effects related to antigen immobilization.
Discussion
Several antibodies that bind human CA-IX, including M75Citation8 and cG250,Citation22 were described during the past decades. G250 recognizes a determinant present in renal-cell carcinoma and absent from normal kidneys.Citation22 These earlier CA-IX antibodies have been exploited for cancer detection and as candidates for therapy.Citation15 More recently, other anti-CA-IX mAbs,Citation23,Citation24 including the newly characterized 11H9, 12H8, 2D7, 2C7, 9B6, and 4A2,Citation15 have been reported. The unique mAb 12H8 is being developed as a novel candidate immunoconjugate,Citation15 especially as a promising radio-immunotherapeutic (RIT) agent (unpublished data). Several of these antibodies, i.e., M75,Citation8 H7,Citation24 and 11H9 and 12H8,Citation15 have been shown to specifically bind to CA-IX’s IDR, especially the PG-like domain. The epitope of the M75 mAb has been well characterized, binding the GEEDLP(S/G) consensus sequence,Citation8,Citation25 whereas mAb H7 appears to interact with the larger 12-residue GEDDPLGEEDLP epitope.Citation24 Applying the PepScan technology (https://www.pepscan.com) to the entire CA-IX extracellular domain sequence suggested that the 12H8 epitope is located at the N-terminus of the CA-IX sequence, while that of 11H9 is likely to be somewhere in the center of the PG-like domain.Citation15
In this study, we set out to examine the interactions of the entire IDR, residues 37–136 (i.e., ePG-1; ), of hCA-IX by NMR spectroscopy and to characterize the precise binding epitopes of mAb 11H9 and 12H8 in conjunction with yeast surface display. Our comprehensive NMR data, utilizing the 15N/Citation13C-labeled ePG-1 polypeptide, showed that hCA-IX’s N-terminal extension domain is without any 3D structure or even a secondary structure preference ( and SupplFig. S2), which confirms recently published data on the same CA-IX domain using unassigned proton NMR spectroscopy and other biophysical methods.Citation12 This conformational property can be ascribed in part to the low sequence complexity of the N-terminal (aa 37–55) and the C-terminal residues (aa 107–136) of CA-IX’s N-terminal extension, qualifying them as ‘flexible linkers’ between the signal sequence and the PG-like domain and between the PG-like domain and CA-IX’s catalytic domain (). The lack of a 3D fold in the PG-like domain () is related to a highly negatively charged sequence that is rich in aspartic and glutamic acids (), as well as the presence of seven repeats consisting of the following six residue-related sequences: GEEDLP (4), GEDDPL (1), SEEDSP (1), and REEDPP (1). An IDR such as the PG-like domain could thus provide the conformational flexibility that may confer strong avidity effects in binding of antibodies to the native hCA-IX dimer.Citation15 In addition, the negatively charged intrinsically disordered PG-like domain may also serve to modulate the aqueous structure and dynamic hydration at the extracellular membrane surface. This property has been linked to the non-catalytic extruding of lactate from the intracellular cancer cell compartment and its apparent “antenna” function in the pH regulation by CA-IX’s catalytic domain.Citation26 Our SPR data indicated that the M75, 11H9, and 12H8 mAbs all bind with high affinity to the immobilized ePG-1 polypeptide, i.e., residues 37–136 of hCA-IX’s entire N-terminal region ( and ).
Interestingly, the 11H9 mAb and F(Ab) were able to bind the ePG-2 polypeptide, but the 12H8 mAb and 12H8 F(Ab) were unable to bind ePG-2, a variant of ePG-1 that lacks the four N-terminal aa residues PQRL40 (SupplFig. S1). We have shown that 12H8 can be used as an in vivo imaging agent for hCA-IX-expressing tumors (unpublished results), which implies that mature hCA-IX expressed by tumor cells thus contains these extreme N-terminal ‘PQRL’ residues. 12H8 titration data with 15N-labeled ePG-1 further delineated N-terminally located residues, especially L40, R42, Q44, E45, and D46 ( and SupplFig. S3C) as the 12H8 binding loci, which is in complete agreement with positive binding of the mAb to the 15-residue peptide fragment PQRLPRMQEDSPLGG displayed on the yeast cell surface () and the earlier published PepScan data.Citation15 Fine epitope mapping by alanine scanning of a 9-residue fragment, LPRMQEDSP, located within the 15-residue peptide fragment PPRLPRMQEDSPLGG, identified the core binding for 12H8 to be sequence LPRMQEDSP within which residues L40, R42, Q44, E45 and D46 cannot be replaced by alanine. It can therefore be concluded that of the four extreme N-terminal residues ‘PQRL’ only L40 is absolutely required for the high-affinity binding of 12H8 to hCA-IX’s N-terminus, which is an important property that would render the binding of 12H8 less sensitive to possible processing variations in signal peptide release.
Compared to the unique epitope of 12H8, those of 11H9 are superficially similar to that of the well-known antibody M75.Citation8,Citation25 First, NMR resonance perturbations on residues Gly43, Gly49, and Gly55 and some Glu and Asp residues in the GEEDLP sequence ( and SupplFig. S3F) indicate a primary binding epitope in the central region of the PG-like domain. However, high-resolution (TROSY) NMR of the ePG-1/F(Ab) complex using a fully deuterated and 15N-labeled ePG-1 polypeptide () revealed that this binding epitope of 11H9 must span two consecutive GEEDLP repeats or the GEEDLPGEEDLP sequence () instead of residing in the single GEEDLP unit like M75.Citation25 It is interesting to note that the antibody H7Citation24 also appears to bind to the tandem of two related 6-residue sequence, i.e., GE20DDPLG25EEDLP30, which overlaps with 11H9‘s secondary or lower-affinity epitope around G25EEDLP30. Our yeast display data showed high-affinity binding of 11H9 with both the PPGEEDLPGEEDLPG and the EDLPGEEDLPEVKPK sequences (). Fine mapping of these putative epitopes identified a continuous stretch of negatively charged residues. i.e., EDLPGEED, as 11H9’s primary binding epitope. None of the amino acids in this sequence can be replaced by alanine (), indicating the critical importance of the first and all glutamic acids (E) in this epitope for high-affinity 11H9 binding, thus setting this antibody apart from M75 and from the H7 antibody the latter of which binds only the GEDDLPGEED epitope. Given the additional residues perturbed by (excess concentrations of) 11H9-F(Ab) (), i.e., Ser17, Ser18, Ser31, and Ser35, there may be a number of low-affinity 11H9 binding sites, namely, D20DLPG25EED, EDLP30SEED, and EDS35PREED40, all of which are variations of the high-affinity epitope EDLPGEED and whose weaker binding to 11H9 could not be detected directly by displaying on the yeast surface smaller fragments containing these sequences (). There are also two EDLPGEED motifs in the GEEDLPGEEDLPGEEDLP region only one of which should be able to bind at a time due to the overlap of two essential residues, i.e., the ED unit in the middle of this 18-residue segment. Either way, 11H9 binding may induce more substantial structural rigidifications in the central region of the otherwise flexible N-terminal IDR of CA-IX.
Antibody–IDR interactions with two complementary binding sites may lead to the formation of a 1:1 bimolecular complex, a 1:2 (11H9:IDR) tri-molecular complex or cross-linked multi-molecular oligomers and/or aggregates. However, a maximum of 22 residues separates the first low-affinity epitope D20DLPG25EED from the last high-affinity epitope EDLPG55EED in the ePG-1 sequence ( or ), which is insufficient for spanning the two symmetrically structured combining sites, leaving the second combining site of 11H9 unoccupied in a bimolecular 1:1 PG:11H9 complex. It is known experimentally that bivalent binding requires a minimum of four residues to span ~1.8 nm,Citation27,Citation28 while bivalent binding to an intact mAb is shown to occur at a spacing of ~16 nm,Citation29 which corresponds to a peptide linker of more than 35 residues. As such, PG-11H9 interactions can only involve the formation of a 2:1 stable complex with both combining sites occupied by the high-affinity epitopes of two separate PG molecules (), as is the case for samples with more than 3:1 excess in PG concentrations (Suppl and SupplFig. S4A/B). Approaching and below the 2:1 molar ratio, unoccupied combining sites in a bivalent 11H9 mAb are also available for interactions with the secondary epitope presented by another PG-11H9 1:1 complex, in other words, the formation of cross-linked antibody aggregates (), as observed for both the 2:1 and 1:1 complex preparation in terms of sample turbidity and precipitations. Cross-linking interactions of a full-length (bivalent) antibody can also occur on the cell-surface or in SPR studies (commonly referred to as avidity effects) depending on the density,Citation15 the chemistry of antigen immobilization ( and SupplFig. S1), and the conformational flexibility of immobilized antigens. Higher-order molecular assembly, a functional feature of intrinsically disordered proteins with multiple binding sites,Citation30 may therefore be exploited in experiments studying the role of CA-IX’s IDR in maintaining pH homeostasis in the hypoxic/acidic cancer microenvironments.
In general, studying antibody–antigen interactions and specifically epitope mapping represents a critical step toward characterizing and understanding the mechanism of action in antibody discovery and therapeutic development.Citation31,Citation32 Established methods for epitope mappingCitation32 include structure determination of antibody-antigen complexes by x-ray crystallography, antigen fragmentation using peptides or peptide libraries, site-directed mutagenesis, hydrogen-deuterium exchange mass spectrometry (HDX-MS),Citation16,Citation33 and to some extent NMR spectroscopy.Citation34 Structure-based methods can be limited,Citation35 often requiring the removal of non-binding sequences/regions, as shown for the M75 studyCitation25 and in other systems.Citation36,Citation37 Epitope mapping efforts also need to recognize the recent identification of a large number of intrinsically unfolded proteins and functionally important intrinsically unfolded protein regions.Citation38 As exemplified by previous workCitation33,Citation37 and by this study, antibody binding to IDPs/IDRs frequently engage the so-called linear or contiguous epitopes,Citation32 which points to potentially high success rates of peptide (fragment)-based methods including the yeast surface display for identifying high-affinity binding sequences. On the other hand, 3D structure determination of full antibody-antigen complexes may fail, especially when an IDP/IPR in complex with the antibody can remain largely flexible, as shown here by the use of TROSY NMR spectroscopy. In this regard, titration NMR spectroscopy has the potential for locating weaker (secondary) binding sites which may function cooperatively with primary epitopes to impart higher-order interactions such as concentration-dependent antibody cross-linking and aggregation. In all, the data presented here demonstrate that high-resolution NMR experiments in combination with the yeast surface display technology provides a powerful method for a comprehensive understanding of antibody–antigen interactions involving intrinsically unstructured proteins or protein regions that may remain largely unfolded and unstructured even in complex with a structure-stabilizing protein such as an antibody.
Materials and methods
Recombinant antibody and F(Ab) production and purification
The 11H9 and 12H8 anti-human CA-IX mouse mAbs were generated, functionally screened, and sequenced as described.Citation15 VH and VL CDR1–3 regions of 11H9 and 12H8 were grafted on a human IgG1 framework. The 11H9 and 12H8 antibodies, and their respective F(Ab) fragments were produced in CHO-3E7 cells. Briefly, cells were grown to a density of 2 × 10Citation6 cell/mL in 150 mL of F17 medium (Catalog No. A1383501, Thermo Fisher Scientific) supplemented with 4 mM L-glutamine and 0.1% Kolliphor P188 in 1.0 L shaker flasks and then transfected with 2 μg total DNA (containing 500 ng each of the heavy and light chain constructs) using PEI MAX™ (Catalog No. 24765, Polysciences, Inc.), then kept at 37°C for 24 h, fed with Tryptone N1 (Catalog No 19,553, Organotechnie, 1% w/v), and maintained at 32°C for another 6 days. Cell culture supernatant was harvested and analyzed by SDS-PAGE. Antibodies and F(Ab)s were captured from the supernatant using MabSelect Sure (Catalog No. 17543802, Cytiva Life Sciences) or Capto L resin (Catalog No. 17547801, Cytivia Life Sciences), respectively, and eluted with elution buffer (mAbs: 100 mM Citrate, 50 mM NaCl, pH 3.0; F(Ab)s: 0.1 M sodium citrate, pH 3.0); eluates were immediately neutralized with 1 M HEPES. Finally, samples were desalted and subjected to a phosphate-buffered saline (PBS) buffer exchange and to UPLC-SEC to ensure that the F(Ab)s are monomeric and finally stored at −80°C.
Polypeptide production, triple isotope labeling and purification
Unlabeled and 13C/15N-labeled polypeptides: An oligonucleotide encoding a human CA-IX N-terminal fragment polypeptide, designated ePG-1 () was codon-optimized, synthesized, and inserted into the pJ401 expression vector (by Atum), while the pJ404 expression vector was used for a variant of ePG-1, ePG-2 (Suppl. ) for recombinant expression in E. coli (Catalog No. 69449, Novagen). Plasmids were electroporated into the BL21 E. coli strain [F – ompT lon hsdSB,rB-,mB-)gal dcm [malB+]K-12(λS)]. One milliliter cultures in LB media were inoculated with these bacterial cell stocks and were incubated for 6 h at 37°C. Cultures were then transferred to 100 mL of filter-sterilized defined media containing M9 salts, 1 mM MgSO4, 5 mg/L thiamine (T4625-25 G, Sigma-Aldrich), 100 µM CaCl2, 2 g/L of glucose (or glucose enriched uniformly with the 13C isotope, i.e., U-Citation13C6-99%, CLM-1396-25, Cambridge Isotope Laboratories, Inc. for 13C-labeling) and 1 g/L of ammonium sulfate (or 15N-labeled ammonium sulfate, i.e., 15N2-99, MLM-713-50, Cambridge Isotope Laboratories, Inc. for 15N-labeling) as the sole carbon and nitrogen source, respectively; kanamycin (BP906–5, Fisher Scientific) (50 µg/mL) (or ampicillin, AMP201.25, Bioshop, at 100 µg/mL for ePG-2) was used as selection markers. After overnight incubation at 37°C, cell cultures were diluted 10-fold in defined media and incubated at 37°C until A600 = 0.8 ~ 1.0. Polypeptide expression was induced by the addition of IPTG (BP1755–10, Fisher Scientific, 1 mM final concentration) and continued incubation for an additional 6 h at 37°C. Cells were then collected by centrifugation at 4,000 rpm for 20 min at 4°C.
To obtainCitation13C/Citation15N/Citation2H-labeled ePG-1, bacteria were adapted to grow in the 15N/Citation13C-labeled defined media (described above). One mL of LB media containing 50 µg/mL kanamycin was inoculated with bacterial cell stocks and incubated at 37°C. After 6 h, the cell cultures were diluted 20-fold in defined media prepared with 100% H2O and incubated at 37°C until A600 ~0.5. Cells were then further diluted fivefold into defined media containing 25% D2OCitation2H-99.9%, DLM-4-99-5000, Cambridge Isotope Laboratories, Inc.) and incubated at 37°C until an A600 of ~0.5 was reached. This process was repeated with defined media containing 50%, 75%, and finally 100% D2O. Cells harvested from defined media containing 100% D2O were then aliquoted in a 30% (v/v) sterile glycerol solution and stored in −80°C. For each production of 2H/Citation13C/Citation15N-labeled ePG-1, stored cell stocks were diluted fivefold in 8 mL of 100% D2O containing 15N/Citation13C-labeled defined media and incubated at 37°C until A600 reached ~0.5. This was followed by three additional steps of fivefold dilution to reach a final volume of 1 L cells with A600 of ~0.8. One mL of 1 M IPTG was then added, and cultures were incubated for 6 h at 37°C. The extent of deuteration for non-exchangeable carbon-attached (CH) protons in ePG-1 was ~98% determined by comparison of proton NMR spectra of un-deuterated and 2H/Citation15N/Citation13C (or D/Citation15N/Citation13C)-labeled ePG-1 samples.
Polypeptide Purification: Bacterially expressed labeled and non-labeled His-tagged ePG-1 (and ePG-2, see Supplementary Materials) were purified in a two-step procedure, first using immobilized metal-affinity chromatography followed by reversed-phase high-performance liquid chromatography. Briefly, cell pellets were suspended in 100 ~ 150 mL of lysis buffer (50 mM Na3PO4, 300 mM NaCl; pH 8) and sonicated on ice for 2 min (10s on, 15s off; power = 6). Cell debris was removed by centrifugation (10,000 rpm, 4°C, 30 min). The cleared supernatant was then mixed with 3 mL of the Ni-NTA resin (30230, QIAGEN) by gentle shaking. After 30 min, the mixture was loaded into a column and washed with 100 mL of the lysis buffer containing 20 mM imidazole. His-tagged ePG-1 (and ePG-2, see Supplementary Materials) was eluted with 15 mL of the lysis buffer containing 250 mM imidazole. The pH of the eluted protein solution was lowered to <4 by adding 2 mL of 10% trifluoroacetic acid (TFA), after which the solution was passed through a 0.45-micron filter before loading onto a reversed-phase C18 column equilibrated with 10% (v/v) aqueous acetonitrile containing 0.1% (v/v) TFA. Purified ePG-1 (and ePG-2) was eluted using a linear gradient of 10%−100% (v/v) aqueous acetonitrile containing 0.1% (v/v) TFA (30 min). Selected peak fractions (UV absorbance at 214 nm) were collected and lyophilized. The protein content of the fractions was confirmed by mass spectroscopy and finally re-dissolved in 450 uL of a mixture of 10% D2O and 90% NMR buffer (50 mM Na3PO4, 50 mM NaCl, 0.2 mM EDTA, and 0.01%-w/v sodium azide; pH 6.8).
Surface plasmon resonance
All SPR assays were carried out at 25°C using a BioRad ProteOn XPR36 (Bio-Rad Laboratories Ltd.). GLC Sensor chips and coupling reagents [10 mM sodium acetate, pH 4.5; sulfo-N-hydroxysuccinimide (S-NHS); 1-ethyl-3-(3-dimethylaminopropyl)-carbodiimide hydrochloride (EDC) and ethanolamine] were purchased from BioRad Inc. PBS containing 3.4 mM EDTA, 0.05% Tween, and 500 nM NaCl was used as running buffer, while 0.85% phosphoric acid (18s, 100 μL/min) was used as regeneration buffer.
GLC sensor chip surfaces were activated by injecting S-NHS/EDC (1:10; 140 s, 100 μL/min). Immediately after, 10 µg/mL anti-His antibody (0.2 mg/mL; Qiagen) was injected (25 µL/min) in running buffer until ~400 to 600 RUs were captured. Remaining active groups were quenched by ethanolamine (1 M; 140s injection at 100 µL/min); mock-activated inter-spots were used for blank referencing. The ePG-1 polypeptide (1.38 mg/ml) was then injected (240s, 25 µL/min), after which serial 11H9 and 12H8 mAb and F(Ab) dilutions (1:3 serial dilutions of 100 nM), along with a buffer blank were injected (120s, 50 μL/min), followed by a buffer injection (300s, 50 μL/min). The M75 antibody (M75, Absolute Antibody Inc.), which is known to bind the human CA-IX PG-like domain was used as positive control, while the in-house generated anti-CA-IX c2D7 mAb, which binds CA-IX’s catalytic domain,Citation15 was used as negative control. Antibody surfaces were regenerated by 0.85% phosphoric acid (18s, 100 μL/min). Sensorgrams were aligned and double-referenced using the buffer blank injection and interspots, analyzed (ProteOn Manager software v3.1) and fitted to the 1:1 binding model to calculate ka, kd, and KD ().
NMR spectroscopy
All NMR experiments were carried out on Bruker Avance-III 600 MHz and 800 MHz NMR spectrometers equipped with a 5 mm PATXI probe with Z-field gradient accessories. Triple resonance experiments for backbone assignments and 3JHNHA measurementsCitation39,Citation40 were performed at 298 K and 800 MHz. The 15N relaxation data, 15N-R1, 15N-R2 and 15N{1 H} NOECitation41 of ePG-1 were recorded at 288 K and 600 MHz. 11H9 titration, ePG-1/11H9-F(Ab) complex TROSY, and TROSY assignment data were recorded at 298 K and 800 MHz. NMR samples of ePG-1 were from 0.2 mM to 1 mM in 450 µL NMR buffer (50 mM in NaPi and 50 mM in NaCl at pH 6.8) with 50 µL D2O. ePG-1 complexes were prepared by adding water-dialysed and dried antibodies or F(Ab) (see below) to the 15N-labeled and 2H/Citation13C/Citation15N-labeled ePG-1 polypeptide in the NMR buffer, respectively.
Resonance assignments: Seven heteronuclear 3D experiments HNCA, HN(CO)CA, HNCACB, CBCA(CO)NH, HNCO, HN(CA)CO,Citation40 and H(NCA)NNHCitation42 were first carried out with a uniformly 15N/Citation13C-labeled ePG-2 polypeptide (SupplTable S1). Backbone 1H and 15N resonance assignments (SupplFig. S2A) of the shorter ePG-2 polypeptide were obtained using six 3D spectra of HNCA, HN(CO)CA, HNCACB, CBCA(CO)NH, HNCO, and HN(CA)CO. All the sequential correlations were verified using a 3D-H(NCA)NNH spectrum that establishes inter-residue connections between those residues separated by Prolines. This H(HCA)NNH spectrum is critical for distinguishing the Gly residue in the first PGEEDL fragment from the following two PGEEDL repeats in the sequence segment PPGEEDLPGEEDLPGEEDL because there are two consecutive Pro residues at the beginning of the first PGEEDL fragment. Based on the sequence-specific resonance assignment of ePG-2 (see Supplementary Materials), backbone resonances of ePG-1 were assigned using HNCA, HN(CO)CA, HNCACB, and CBCA(CO)NH spectra. The TROSYCitation20 spectrum of the ePG-1/11H9-F(aAb) complex was assigned using TROSY-HNCA and TROSY-HNCACBCitation21 recorded with the uniformly 2H/Citation13C/Citation15N-labeled ePG-1 in complex with non-labeled 11H9-F(Ab)
15N NMR relaxation measurements: 15N-R1, 15N-R2, and 15N{Citation1H} NOECitation41 were acquired on a uniformly 15N-labeled ePG-1 at the lower temperature of 288 K. NMR pulse sequences from the (Bruker) spectrometer library, named hsqct2etf3gpsi, hsqct1etf3gpsi, and hsqcnoef3gpsi, were used for the measurement of 15N-R1, 15N-R2, and 15N{Citation1H} NOE, respectively. The 15N-R1 measurements were performed with 10 relaxation delays in the order of 5, 1500, 60, 1200, 140, 950, 220, 700, 330, and 500 ms, and three repetition delays at 60, 700, 1200 ms. The 15N-R2 measurements were performed with 18 relaxation delays in the order of 115.2, 14.4, 345.6, 28.8, 316.8, 43.2, 288.0, 57.6, 259.2, 72.0, 230.4, 86.4, 210.6, 100.8, 172.8, 115.2, 144.0, and 129.6 ms, and three repetition delays at 14.4, 316.8, and 201.6 ms. In 15N{Citation1H} NOE measurements, the reference spectrum was acquired with a relaxation delay of 6s and the 1H saturated spectrum was recorded with a 3s delay followed by a 3s saturation. Residue-specific assignments of the 15N relaxation data for ePG-1 at 288 K were obtained by tracing movements with temperature variations of six 1H/Citation15N-HSQC spectra recorded at 298 K, 296 K, 294 K, 292 K, 290 K, and 288 K, respectively. The 3D-HNHA spectrum recorded with Bruker pulse sequence named hnhagp3d at 288 K and 600 MHz was used to determine the backbone coupling constants of the ePG-1 polypeptide. All 3D spectra and NMR relaxation data were processed with nmrPipe.Citation43 Spectra were analyzed by using nmrView.Citation44 The cross-peak intensities of NMR relaxation measurements were extracted using Non-Linear N-Dimensional Spectral Modeling, a module of NMRPipe.Citation43 15N-R1 and 15N-R2 values were fitted using in-house developed software programs.
NMR titration experiments: Interactions of the ePG-1 polypeptide with antibodies 12H8 and 11H9 were followed by NMR signal perturbations in the 1H-Citation15N HSQC spectrum (at 800 MHz) of 15N-labeled (including 15N/Citation13C-labeled and 15N/Citation13C/Citation2H-labeled) ePG-1 samples after addition of less than stoichiometric amounts of the respective antibody. Both full-length mAbs 12H8 and 11H9 and their F(Ab) fragments were used for the titration experiments. The F(Ab) titrations proceeded beyond stoichiometric (1:1) ratios to excess for the respective F(Ab) in order to determine the existence of secondary binding sites and/or relative binding affinities in the case of multiple binding epitopes (i.e., for 11H9). For all experiments, the antibodies were dialyzed into pure water followed by lyophilization and reconstitution either in pure water (to ~100–150 mg/mL) or in the phosphate buffer of 50 mM in NaPi and 50 mM in NaCl at pH 6.8 (or NMR buffer). Small volumes of the concentrated antibody solutions were introduced into prepared samples of ePG-1 under the appropriate buffer and pH conditions. For 11H9 mAb titrations specifically, a sample of the 15N-labeled ePG-1 was first prepared at 1.6 mM in a volume of 100 µL with 60 µL of D2O and 40 uL of the NMR buffer and the 11H9 mAb solution at 120 µM by dissolving the dried 11H9 powder (see above) in 450 µL of the NMR buffer. Appropriate portions of these two solutions were mixed to produce the various samples of the PG:11H9 complexes with approximate molar ratios of 6:1, 3:1, 2:1, and 1:1, respectively.
Epitope mapping by yeast surface display
The CA-IX fragments covering the PG-like domain were cloned in the pPNL6 vector (provided by the Pacific Northwest National Laboratory, USA), expressed as fusion proteins [Aga2-HA-(CA-IX)-MYC] and covalently displayed on the cell wall of the yeast surface.Citation45 The relative amount of displayed fusion protein was measured by probing yeast cells with a chicken polyclonal Anti-c-Myc IgY antibody (ab19233; Abcam), followed by a horseradish peroxidase (HRP)-conjugated goat anti-chicken IgY (H+L) secondary antibody (ab97135, Abcam), and used to normalize the binding signal for anti-hCA-IX mAbs. Binding of the 11H9, 12H8, and M75 mAbs to the yeast cells was assessed using a whole yeast cell ELISA.Citation46 Briefly, various clones of the induced yeast cells were loaded into a 96-well filter plate (1×10Citation7 cells per well), blocked with PBS containing 1 mg/ml bovine serum albumin, incubated with the anti-hCAIX antibodies, after which the binding of 11H9, 12H8, and M75 mAbs was revealed by using an HRP-conjugated secondary antibody of either goat anti-mouse IgG (ab6789, Abcam) or goat anti-human IgG-Fc (ab97225, Abcam) followed by incubation with the HRP substrate tetramethyl benzidine, according to the manufacturer’s conditions, and read at OD450. To determine whether the binding epitopes are linear or structured, yeast cells were in addition heated at 80°C for 30 min to denature displayed hCA-IX fragments, then chilled on ice for 20 min prior to incubation with the antibodies.
Fine epitope mapping: Tiling fragments of 15 amino acids in length with five amino acids overlapping on each end covering the entire CA-IX N-terminal fragment (residues 37–140) were cloned, expressed, and displayed on the yeast surface. The binding of the 11H9, 12H8, and M75 mAbs to these yeast cell-expressed fragments was analyzed by ELISA as described above. Systematic deletion mutagenesis from either the N- or C-terminus of the mAb-binding fragment(s) was applied to delineate the “minimal” binding epitope, and alanine scanning mutagenesis was applied to assess the contribution of each individual amino acid within the minimal epitope to the binding of the mAbs.
List of Abbreviations:
Supplemental Material
Download Zip (29.3 MB)Acknowledgments
We thank Mimi Simmons and Maurizio Achione of the Quality Attributes Characterization Team for collecting the size-exclusion chromatography elution profiles of ePG-1, 11H9 and their 2:1 and 1:1 molecular complexes.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Supplementary material
Supplemental data for this article can be accessed online at https://doi.org/10.1080/19420862.2023.2248672.
Additional information
Funding
References
- Pastorekova S, Parkkila S, Pastorek J, Supuran CT. Carbonic anhydrases: current state of the art, therapeutic applications and future prospects. J Enzyme Inhib Med Chem. 2004;19(3):199–15. PMID: 15499993. doi:10.1080/14756360410001689540.
- Thiry A, Dogne JM, Masereel B, Supuran CT. Targeting tumor-associated carbonic anhydrase IX in cancer therapy. Trends Pharmacol Sci. 2006;27:566–73. doi:10.1016/j.tips.2006.09.002. PMID: 16996620.
- Ihnatko R, Kubes M, Takacova M, Sedlakova O, Sedlak J, Pastorek J, Kopacek J, Pastorekova S. Extracellular acidosis elevates carbonic anhydrase IX in human glioblastoma cells via transcriptional modulation that does not depend on hypoxia. Int J Oncol. 2006;29:1025–33. PMID: 16964400 https://www.ncbi.nlm.nih.gov/pubmed/16964400.
- Andreucci E, Peppicelli S, Carta F, Brisotto G, Biscontin E, Ruzzolini J, Bianchini F, Biagioni A, Supuran CT, Calorini L. Carbonic anhydrase IX inhibition affects viability of cancer cells adapted to extracellular acidosis. J Mol Med (Berl). 2017;95:1341–53. doi:10.1007/s00109-017-1590-9. PMID: 28929255.
- Becker HM, Deitmer JW. Transport metabolons and acid/base balance in tumor cells. Cancers Basel. 2020;12(4):899. PMID: 32272695. doi:10.3390/cancers12040899.
- Pastorekova S, Gillies RJ. The role of carbonic anhydrase IX in cancer development: links to hypoxia, acidosis, and beyond. Cancer Metastasis Rev. 2019;38:65–77. doi:10.1007/s10555-019-09799-0. PMID: 31076951.
- De Simone G, Supuran CT. Carbonic anhydrase IX: Biochemical and crystallographic characterization of a novel antitumor target. Biochim Biophys Acta. 2010;1804:404–09. doi:10.1016/j.bbapap.2009.07.027. PMID: 19679200.
- Zavada J, Zavadova Z, Pastorek J, Biesova Z, Jezek J, Velek J. Human tumour-associated cell adhesion protein MN/CA IX: identification of M75 epitope and of the region mediating cell adhesion. Br J Cancer. 2000;82(11):1808–13. PMID: 10839295. doi:10.1054/bjoc.2000.1111.
- Csaderova L, Debreova M, Radvak P, Stano M, Vrestiakova M, Kopacek J, Pastorekova S, Svastova E. The effect of carbonic anhydrase IX on focal contacts during cell spreading and migration. Front Physiol. 2013;4:271. doi:10.3389/fphys.2013.00271. PMID: 24101905.
- Swayampakula M, McDonald PC, Vallejo M, Coyaud E, Chafe SC, Westerback A, Venkateswaran G, Shankar J, Gao G, Laurent EMN, et al. The interactome of metabolic enzyme carbonic anhydrase IX reveals novel roles in tumor cell migration and invadopodia/MMP14-mediated invasion. Oncogene. 2017;36(45):6244–61. doi:10.1038/onc.2017.219. PMID: 28692057.
- Opavsky R, Pastorekova S, Zelnik V, Gibadulinova A, Stanbridge EJ, Zavada J, Kettmann R, Pastorek J. Human MN/CA9 gene, a novel member of the carbonic anhydrase family: structure and exon to protein domain relationships. Genomics. 1996;33(3):480–87. PMID: 8661007. doi:10.1006/geno.1996.0223.
- Langella E, Buonanno M, Vullo D, Dathan N, Leone M, Supuran CT, De Simone G, Monti SM. Biochemical, biophysical and molecular dynamics studies on the proteoglycan-like domain of carbonic anhydrase IX. Cell Mol Life Sci. 2018;75:3283–96. doi:10.1007/s00018-018-2798-8. PMID: 29564477.
- Jorda J, Xue B, Uversky VN, Kajava AV. Protein tandem repeats - the more perfect, the less structured. FEBS J. 2010;277(12):2673–82. PMID: 20553501. doi:10.1111/j.1742-464X.2010.07684.x.
- Langella E, Buonanno M, De Simone G, Monti SM. Intrinsically disordered features of carbonic anhydrase IX proteoglycan-like domain. Cell Mol Life Sci. 2021;78:2059–67. doi:10.1007/s00018-020-03697-3. PMID: 33201250.
- Lenferink AEG, McDonald PC, Cantin C, Grothe S, Gosselin M, Baardsnes J, Banville M, Lachance P, Robert A, Cepero-Donates Y, et al. Isolation and characterization of monoclonal antibodies against human carbonic anhydrase-IX. MAbs. 2021;13(1):1999194. doi:10.1080/19420862.2021.1999194. PMID: 34806527.
- Sheff JG, Kelly JF, Robotham A, Sulea T, Malenfant F, L’Abbe D, Duchesne M, Pelletier A, Lefebvre J, Acel A, et al. Hydrogen-deuterium exchange mass spectrometry reveals three unique binding responses of mAbs directed to the catalytic domain of hCAIX. MAbs. 2021;13:1997072. doi:10.1080/19420862.2021.1997072. PMID: 34812124.
- Pardi A, Billeter M, Wuethrich K. Calibration of the angular dependence of the amide proton-Cα proton coupling constants, 3JHNα, in a globular protein. J Mol Biol. 1984;180(3):741–5110.1016/0022-2836(84)90035-4
- Ota M, Koike R, Amemiya T, Tenno T, Romero PR, Hiroaki H, Dunker AK, Fukuchi S. An assignment of intrinsically disordered regions of proteins based on NMR structures. J Struct Biol. 2013;181:29–36. doi:10.1016/j.jsb.2012.10.017. PMID: 23142703.
- Pascal SM. NMR primer: An HSQC-based approach with vector animations. West Sussex, UK: IM Publications LLP; 2008. p. 31.
- Pervushin K, Riek R, Wider G, Wuthrich K. Attenuated T 2 relaxation by mutual cancellation of dipole–dipole coupling and chemical shift anisotropy indicates an avenue to NMR structures of very large biological macromolecules in solution. Proc Natl Acad Sci U S A. 1997;94(23):12366–71. PMID: 9356455. doi:10.1073/pnas.94.23.12366.
- Salzmann MW, Wider G, Pervushin K, Senn H, Wüthrich K. TROSY-type triple-resonance experiments for sequential NMR assignments of large proteins. J Am Chem Soc. 1999;121(4):844–48. doi:10.1021/ja9834226.
- Oosterwijk E, Ruiter DJ, Hoedemaeker PJ, Pauwels EK, Jonas U, Zwartendijk J, Warnaar SO. Monoclonal antibody G 250 recognizes a determinant present in renal-cell carcinoma and absent from normal kidney. Int J Cancer. 1986;38(4):489–94. PMID: 2428759. doi:10.1002/ijc.2910380406.
- Xu C, Lo A, Yammanuru A, Tallarico AS, Brady K, Murakami A, Barteneva N, Zhu Q, Marasco WA. Unique biological properties of catalytic domain directed human anti-CAIX antibodies discovered through phage-display technology. PLoS One. 2010;5(3):e9625. PMID: 20224781. doi:10.1371/journal.pone.0009625.
- Stravinskiene D, Imbrasaite A, Petrikaite V, Matulis D, Matuliene J, Zvirbliene A. New monoclonal antibodies for a selective detection of membrane-associated and soluble forms of carbonic anhydrase IX in human cell Lines and biological samples. Biomolecules. 2019;9(8):304. PMID: 31349673. doi:10.3390/biom9080304.
- Král V, Mader P, Collard R, Fábry M, Horejsí M, Rezácová P, Kozísek M, Závada J, Sedlácek J, Rulísek L, et al. Stabilization of antibody structure upon association to a human carbonic anhydrase IX epitope studied by X-ray crystallography, microcalorimetry, and molecular dynamics simulations. Proteins. 2008;71(3):1275–87. doi:10.1002/prot.21821. PMID: 18041760.
- Ames S, Pastorekova S, Becker HM. The proteoglycan-like domain of carbonic anhydrase IX mediates non-catalytic facilitation of lactate transport in cancer cells. Oncotarget. 2018;9:27940–57. doi:10.18632/oncotarget.25371. PMID: 29963253.
- DiMaio J, Gibbs B, Munn D, Lefebvre J, Ni F, Konishi Y. Bifunctional thrombin inhibitors based on the sequence of hirudin45-65. J Biol Chem. 1990;265(35):21698–703. PM:2254323. doi:10.1016/S0021-9258(18)45796-1.
- Maraganore JM, Bourdon P, Jablonski J, Ramachandran KL, Fenton JW. Design and characterization of hirulogs: a novel class of bivalent peptide inhibitors of thrombin. Biochemistry. 1990;29:7095–101. doi:10.1021/bi00482a021. PM:2223763.
- Alan SI, Smyrlaki I, Rosa J, Grevys A, Bratlie D, Bratlie D, Sandlie I, Michaelsen TE, Andersen JT, Högberg B. Inger Sandlie, Terje Einar Michaelsen, Jan Terje Andersen and Björn Högberg. Binding to nanopatterned antigens is dominated by the spatial tolerance of antibodies. Nature Nanotech. 2019;14(2):184–90. doi:10.1038/s41565-018-0336-3.
- Banani SF, Lee HO, Hyman AA, Rosen MK. Biomolecular condensates: organizers of cellular biochemistry. Nat Rev Mol Cell Biol. 2017;18:285–98. doi:10.1038/nrm.2017.7. PMID: 28225081.
- Abbott WM, Damschroder MM, Lowe DC. Current approaches to fine mapping of antigen-antibody interactions. Immunology. 2014;142:526–35. doi:10.1111/imm.12284. PMID: 24635566.
- Nilvebrant J, Rockberg J. An Introduction to epitope mapping. Methods Mol Biol. 2018;1785:1–10. doi:10.1007/978-1-4939-7841-0_3. PMID: 29714010.
- Sheff J, Wang P, Xu P, Arbour M, Masson L, van Faassen H, Hussack G, Kemmerich K, Brunette E, Stanimirovic D, et al. Defining the epitope of a blood–brain barrier crossing single domain antibody specific for the type 1 insulin-like growth factor receptor. Sci Rep. 2021;11(1):4284. doi:10.1038/s41598-021-83198-w. PMID: 33608571.
- Simonelli L, Pedotti M, Bardelli M, Jurt S, Zerbe O, Varani L. Mapping antibody epitopes by solution NMR spectroscopy: Practical considerations. Methods Mol Biol. 2018;1785:29–51. doi:10.1007/978-1-4939-7841-0_3. PMID: 29714010.
- King MT, Brooks CL. Methods in molecular Biology. Epitope mapping of antibody-antigen interactions with X-ray crystallography. Methods Mol Biol. 2018;1785:13–28. doi:10.1007/978-1-4939-7841-0_3. PMID: 29714010.
- Menting JG, Lawrence CF, Kong GK, Margetts MB, Ward CW, Lawrence MC. Structural congruency of ligand binding to the insulin and insulin/type 1 insulin-like growth factor hybrid receptors. Structure. 2015;23(7):1271–82. PMID: 26027733. doi:10.1016/j.str.2015.04.016.
- Morales RA, MacRaild CA, Seow J, Krishnarjuna B, Drinkwater N, Rouet R, Anders RF, Christ D, McGowan S, Norton RS. Structural basis for epitope masking and strain specificity of a conserved epitope in an intrinsically disordered malaria vaccine candidate. Sci Rep. 2015;5:10103. doi:10.1038/srep10103. PMID: 25965408.
- Dunker AK, Oldfield CJ. Back to the future: Nuclear magnetic resonance and bioinformatics studies on intrinsically disordered proteins. Adv Exp Med Biol. 2015;870:1–34. doi:10.1007/978-3-319-20164-1_1. PMID: 26387098.
- Bracken C, Carr PA, Cavanagh J, Palmer AG. Temperature dependence of intramolecular dynamics of the basic leucine zipper of GCN4: implications for the entropy of association with DNA. J Mol Biol. 1999;285(5):2133–46. PM:9925790. doi:10.1006/jmbi.1998.2429.
- Cavanagh J, Fairbrother WJ, Palmer AG III, Skelton NJ. Protein NMR spectroscopy: principles and practice. San Diego, USA: Academic Press; 1996.
- Kay LE, Torchia DA, Bax A. Backbone dynamics of proteins as studied by 15N inverse detected heteronuclear NMR spectroscopy: application to staphylococcal nuclease. Biochemistry. 1989;28(23):8972–79. PMID: 2690953. doi:10.1021/bi00449a003.
- Weisemann R, Ruterjans H, Bermel W. 3D triple-resonance NMR techniques for the sequential assignment of NH and 15N resonances in 15N- and 13C-labelled proteins. J Biomol NMR. 1993;3(1):113–20. PMID: 8448431. doi:10.1007/BF00242479.
- Delaglio F, Grzesiek S, Vuister GW, Zhu G, Pfeifer J, Bax A. NMRpipe: a multidimensional spectral processing system based on UNIX pipes. J Biomol NMR. 1995;6(3):277–93. PM:8520220. doi:10.1007/BF00197809.
- Johnson BA, Blevins RA. NMRview: A computer program for the visualization and analysis of NMR data. J Biomol NMR. 1994;4:603–14. doi:10.1007/BF00404272. PMID: 22911360.
- Feldhaus MJ, Siegel RW, Opresko LK, Coleman JR, Feldhaus JM, Yeung YA, Cochran JR, Heinzelman P, Colby D, Swers J, et al. Flow-cytometric isolation of human antibodies from a nonimmune Saccharomyces cerevisiae surface display library. Nat Biotechnol. 2003;21:163–70. doi:10.1038/nbt785. PMID: 12536217.
- Cochran JR, Kim YS, Olsen MJ, Bhandari R, Wittrup KD. Domain-level antibody epitope mapping through yeast surface display of epidermal growth factor receptor fragments. J Immunol Methods. 2004;287:147–58. doi:10.1016/j.jim.2004.01.024. PMID: 15099763.